
Abstract
A palynological reconstruction (n = 25 profiles) suggests that the northern extent of lodgepole pine (Pinus contorta var. latifolia Engelmann ex S. Watson) occurred between 59° and 60°N latitude in northwest North America from 10,000 to 7000 calendar years before present (cal. yr BP) prior to entering Yukon. Although specific migration pathways could not be resolved with the available palynological data, mountains along the southern edge of Yukon appear to have diverted lodgepole pine migration through the Carcross and Frances Lake areas in southwest and southeast Yukon, respectively. Migration in the southwest (70 m/yr) was likely confined to lower elevations of the Yukon and Teslin river valleys, with lodgepole pine reaching 61°N ~2000 cal. yr BP. Along the eastern route, migration was channeled through a 15–20 km wide pass in a 200 km mountainous front. After breaching the Liard drainage divide north of Frances Lake ~4000 cal. yr BP, migration progressed northwest (160–220 m/yr) along the Tintina Trench. Lodgepole pine was estimated to have reached its near present-day northern limit (~63°N) ~1790 cal. yr BP, which is ~1290 years earlier than previously thought. This difference in arrival dates is due the use of a >5% rather than a >15% pine pollen content threshold, which appears to correspond with >1% pine tree cover in the landscape. Climatic cooling after 1000 cal. yr BP that caused a population decline at higher elevations is hypothesized to explain the present-day sparse and disjunct distribution of pine across Yukon north of 61°N.
Introduction
Lodgepole pine (Pinus contorta var. latifolia Engelmann ex S. Watson) is a common western North American cordilleran tree species with a distribution that ranges from central Colorado in the western USA (38°N latitude) to central Yukon (~63.5°N) in northwest Canada (Lotan and Critchfield, 1990). During the Wisconsinan glacial maximum at ~21,400 calendar years (cal. yr BP) or 18,000 radiocarbon years before present (14C yr BP), however, its northern distribution was south of 48°N latitude with populations occurring both west and east of the Rocky Mountains from where it migrated northward following deglaciation (Godbout et al., 2008; Strong and Hills, 2006). East of the Rocky Mountains, lodgepole pine was present at the Canada border (49°N) about 16,300 cal. yr BP (14,000 14C yr BP) and continued moving northward through Alberta and northeast British Columbia between retreating Laurentide and Rocky Mountain cordilleran glacial fronts (Strong and Hills, 2005). As cordilleran glaciers retreated, lodgepole pine also probably migrated westward into higher elevations along the eastern slopes of the Rocky Mountains. MacDonald and Cwynar (1985) suggested lodgepole pine reached its near present-day northern limit about 500 cal. yr BP (430 14C yr BP), although range expansion continues to occur (Johnstone and Chapin, 2003).
The postglacial migratory history of lodgepole pine is not well understood. Despite numerous palynological studies in western North America, fundamental questions still exist such as – Where exactly did various populations reside during the glacial maximum, and what were the routes and the chronology of Holocene migration from each population center? The latter will likely be a complex palaeobiogeographical problem to unravel whether based on comparisons of morphological attributes (LV Hills, unpublished data, 2012) or genetic analysis (e.g. Godbout et al., 2008; Wheeler and Guries, 1982). To initially address the latter question, MacDonald and Cwynar (1985) used palynology to reconstruct the arrival dates of lodgepole pine at 14 sites along the eastern slopes of the Rocky Mountains between the southern Canadian border and central Yukon. A review of their 14C lodgepole pine arrival dates suggested a continual but varied rate of north-northwest migration toward Yukon; however, there were inconsistencies in the data. For example, a reversal in arrival dates occurred in the vicinity of southeastern Yukon, i.e. pine arrived at John Klondike Bog (6400 cal. yr BP [5600 14C yr BP], 60°21′N) 700 calendar years before a more southern site (Lac Ciel Blanc Lake, 5700 cal. yr BP [5000 14C yr BP], 59°33′N). There also appeared to be a trend of later arrival in southwestern than southeastern Yukon at near the same latitude (MacDonald and Cwynar, 1985: 1040). The purpose of this analysis is to: (1) re-evaluate the chronology of lodgepole pine migration into Yukon based on archival palynological analyses, mostly compiled after publication of the MacDonald and Cwynar (1985) study; and (2) identify likely migratory routes of lodgepole pine into and through southern Yukon based on arrival dates in conjunction with terrain and ecological considerations. A more comprehensive understanding than currently exists of mid- to late-Holocene lodgepole pine migration patterns in Yukon would help to explain the present-day discontinuous distribution, and what appears in some cases to be disjunct occurrences in the northernmost portion of its range.
Materials and methods
Study area
Mountains and foothills occur throughout most of southern Yukon, with local valley to ridge relief typically ranging from 500–1000 m and upper elevations often approximating 2000 m. South of 64°N latitude, only the northwest corner of the territory escaped Wisconsinan glaciation. Jackson et al. (1991) estimated that regional deglaciation was complete in southern Yukon prior to 11,400 cal. yr BP (10,000 14C yr BP), with high-elevation source areas ice-free by 10,200 cal. yr BP (9000 14C yr BP); except the St. Elias Mountains in southwest Yukon where extensive cordilleran glaciers still exist (Strong, 2013). Boreal forest dominates most moderately well- to well-drained lower-elevation areas in the present-day landscape. Along the Yukon–British Columbia border, lodgepole pine forests are a common component of the vegetation, whereas western white spruce (Picea albertiana ssp. albertiana S. Brown emend. Strong and Hills, 2006) and black spruce (Picea mariana (Mill.) BSP.) predominate farther north. An exception to the latter is the general occurrence of aspen (Populus tremuloides Michx.) in a broad triangular area south and southeast of Pelly Crossing (Figure 1), north of 61°N latitude (NCb ecoclimatic region, Strong, 2013). Mid- and high-elevation areas in southern Yukon are dominated by open-growing spruce/willow-birch (Picea/Salix-Betula) subalpine shrublands and alpine ecosystems, respectively.

Palynological analysis sites in Yukon and surrounding areas, and the present-day distribution of lodgepole pine (Pinus contorta var. latifolia) in Yukon. See Table 1 for the site names that were used in the study.
Lodgepole pine in the year 2009 occupied ~12,680 km2 of land in Yukon, based on forest cover mapping (Yukon Energy Mines and Resources, 2009), these trees were spread across 45.8% (221,123 km2) of the territory (Figure 1). Only along the southern Yukon border in the Watson Lake area and in the vicinity of Carcross does lodgepole pine abundance on the landscape exceed 25% (WL Strong, unpublished data, 2012). Lodgepole pine decreases rapidly in abundance northward. The area north of 61°N latitude represents 73% of the species’ range in Yukon, but includes only 26% of its abundance, with an average density of 2 ha/km2. The median latitudinal width of the range is 440 km, but most lodgepole pine stands occur within the vicinity (e.g. ±15 km) of the Yukon River and the lower reach of the Pelly River (Figure 1). The most northerly and westerly stand of lodgepole pine in North America occurs ~7 km north of Dawson (64°8′N, 139°26′W, Yukon Energy Mines and Resources, 2009).
Ecologically, lodgepole pine is a fire-dependent shade-intolerant seral species, and as a result, it is most common in environments where wildfires periodically rejuvenate the vegetation. Monocultural stands are common, particularly on higher elevation sites in mid-latitudinal areas (Strong, 1992) and in more northerly latitudes, but stands mixed with aspen can occur at low elevations north of 51°N. Shade-tolerant late-successional trees such as western white spruce, black spruce, and firs (e.g. Abies lasiocarpa (Hook.) Nutt. and A. balsamea (L.) Mill.) are often a component of older lodgepole pine stands, and eventually replacing them through ecological succession. Black spruce is a common stand component at high elevations in mountainous areas and northern latitudes in Yukon. Lodgepole pine occurs primarily in boreal and subalpine forests throughout its distribution, and can occupy a broad spectrum of site conditions ranging from hot dry south-facing slopes to cool damp north aspects. Soils can include fine- to coarse-texture substrates, with rapid to imperfect drainage. Lodgepole pine stands in Yukon tend to have overstoreys with 30–50% canopy cover, and understoreys dominated by mosses and lichens with a modest vascular plants cover. Tree height decreases from ≤30 m in the mid-portion of its range in western Alberta to <10–15 m near its northern limit in Yukon.
Data sources and chronology
Palynological studies in Yukon, eastern Alaska (east of 146°W), western Northwest Territories (west of 120°W), and northern British Columbia (north of 55°N) were reviewed for their suitability for inclusion in the analysis. Criteria for inclusion included the occurrence of radiocarbon or other reliable stratigraphic dating (e.g. White River volcanic ash with a known age − 1147 cal. yr BP, Clague et al., 1995), and continuous stratigraphic pollen percentage values.
A constant rate of sedimentation was assumed between 14C stratigraphic dates when estimating a lodgepole pine arrival date within a palynological profile. Arrival was considered to occur when pine pollen first exceeded 5%. This threshold value was chosen because near-surface pollen spectra in Yukon profiles contained ≤5% pine pollen when lodgepole pine tree abundance was ≤1% in the landscape. This choice was also supported by a polynomial regression (Figure 2), which predicted a 5.4% lodgepole pine pollen content when 1% tree cover occurred within 30 km of a deposition site. Estimates of tree abundance were based on the assumption that wind-dispersed pine pollen, which is shed during the June to early July period, largely originates within a 30–60 km radius of a deposition site (McLeod and MacDonald, 1997; Prentice, 1985). A lodgepole pine tree abundance value for each pollen source area was derived from forest cover data summarized according to a 0.5° latitude × 1° longitude grid that covered all of southern Yukon (WL Strong, unpublished data, 2012). Because pollen source areas overlapped grid cell boundaries, an average was determined by weighting cell values according to their proportional occurrence. Grid cell values (i.e. Σ (polygon area × crown closure × proportional of lodgepole pine)) were compiled from 1:50,000-scale Yukon Energy Mines and Resources (2009) forest cover mapping.
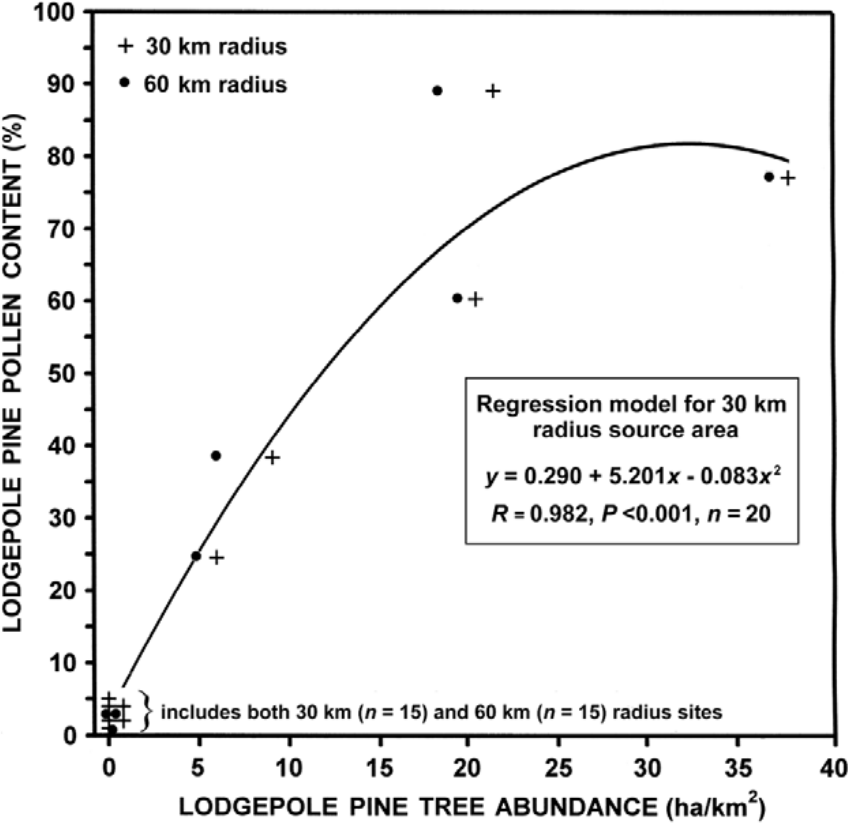
A comparison of lodgepole pine (Pinus contorta var. latifolia) tree and near-surface pine pollen abundance in Yukon (n = 20 sites) based on 30 km and 60 km radius source areas, with a polynomial regression model. Tree abundance was compiled from 1:50,000-scale forest cover maps (Yukon Energy Mines and Resources, 2009) based on a 0.5° latitude × 1° longitude (~56 km × 53 km) grid. Pollen percentages were based on data from: Antifreeze and Eikland Pond (Vermaine and Cwynar, 2010); Buggy Pond (MacDonald and Cwynar, 1985); Candelabra and Hail Lake (Cwynar and Spear, 1995); Grayday, Honeymoon, and Monkhood Pond (Cwynar and Spear, 1991); Gull Lake (Birks, 1980); Ittlemit Lake (Wang and Geurts, 1991); Jenny Lake (Stuart et al., 1988); Kettlehole Pond (Cwynar, 1988); Lake WA01 (Rainville and Gajewski, 2012); Long Last Lake and Two Horsemen Pond (Keenan and Cwynar, 1992); Shaky Hand Creek (Campbell, 1987); Sulphur Lake (Lacourse and Gajewski, 2000); Tourbière Volcano (Bourgeois and Geurts, 1983); Upper Fly Lake (Bunbury and Gajewski, 2009); and 16 Mile Bog (Anderson, 1970).
To maximize the comparability of arrival dates among palynological sites and other information, radiocarbon dates were converted to calendar years (Fairbanks et al., 2005; see http://radiocarbon.ldeo.columbia.edu/research/radiocarbon.htm). Once determined, calendar year arrival dates were plotted and an isochron (lines of equal time) map was constructed to illustrate migration patterns. Boone Lake, immediately east of the southeast corner of the study area in Alberta, with a lodgepole pine arrival date of 12,900 cal. yr BP (11,000 14C yr BP) (White and Mathewes, 1986) was used in conjunction with British Columbia sites to facilitate the placement of isochrons older than 10,000 cal. yr. Initially, three-dimensional contouring (Statsoft, 1995) was used to model the chronology of lodgepole pine migration based on arrival dates, but this method only provided a generalized perspective. For the presented migration models, isochrons were manually positioned based on an assumed constant rate of migration between adjacent palynological sites in lowland areas. Although sophisticated computer-based approaches are possible, they do not necessarily yield substantially better results (King and Herstrom, 1997), but can be convenient when handling large data sets. If an inconsistency in arrival dates occurred among nearby sites, preference was given to the most recently measured 14C samples.
Results
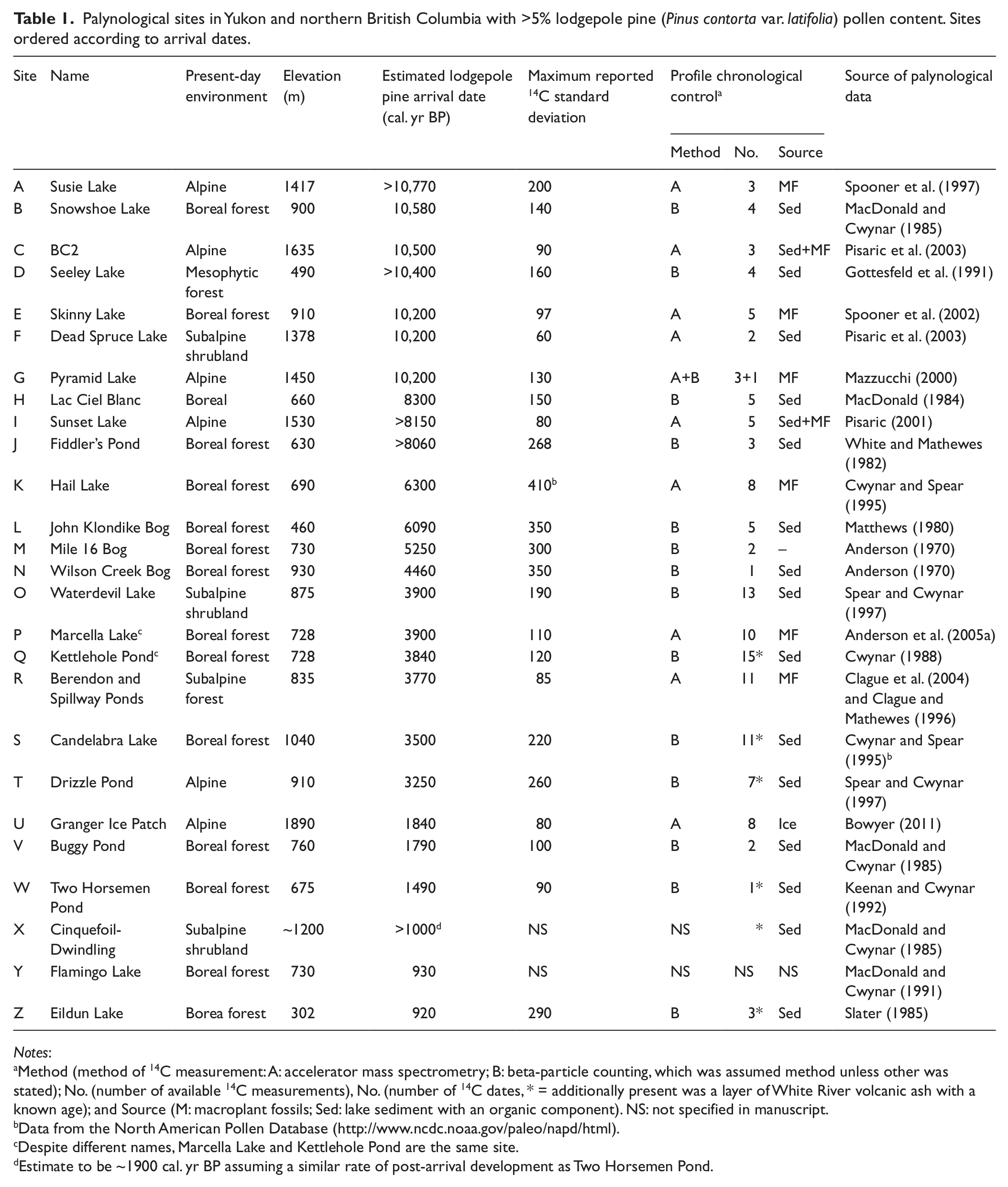
Among 126 reviewed pollen profiles, 75 were initially considered suitable (Figure 1). Of the latter, 26 had a >5% pine pollen content at some point in their stratigraphy (Table 1). All sites outside the contemporary geographical range of lodgepole pine had less pine pollen content, except Eidlun Lake (site Z, Figure 1 and Table 1). This site was excluded from analysis because it occurred within the present-day geographical range of jack pine (Pinus banksiana Lamb.) rather than lodgepole pine (Porsild and Cody, 1980). Among the remaining sites, 15 sites had four or more radiocarbon profile dates (n = >135, Table 1). Five sites contained White River tephra, which has a known age. Most profiles also included the uppermost portion of sediment with an age of zero years. Ten studies used accelerator mass spectrometry for measuring 14C content, with dating primarily based on plant macrofossils (Table 1). Bulk lake sediments with a high organic content were primarily used for 14C dating at 16 sites. Maximum variation around a 14C date within a stratigraphic profile was ≤200 years for two-thirds of the sites. More extreme deviations of 300–400 years tended to be associated with 14C dates determined prior to the mid 1980s (Table 1).
Palynological sites in Yukon and northern British Columbia with >5% lodgepole pine (Pinus contorta var. latifolia) pollen content. Sites ordered according to arrival dates.
Notes:
Method (method of 14C measurement: A: accelerator mass spectrometry; B: beta-particle counting, which was assumed method unless other was stated); No. (number of available 14C measurements), No. (number of 14C dates, * = additionally present was a layer of White River volcanic ash with a known age); and Source (M: macroplant fossils; Sed: lake sediment with an organic component). NS: not specified in manuscript.
Data from the North American Pollen Database (http://www.ncdc.noaa.gov/paleo/napd/html).
Despite different names, Marcella Lake and Kettlehole Pond are the same site.
Estimate to be ~1900 cal. yr BP assuming a similar rate of post-arrival development as Two Horsemen Pond.
Lodgepole pine arrived at ~59°N latitude in north-central British Columbia about 10,000 cal. yr BP from the southeast (Figure 3). Its arrival at the same latitude in northeast British Columbia was delayed by an additional 1000–1500 cal. yr. Migration after 10,000 cal. yr BP continued to the north and northwest (Figure 3). Post-10,000 to 7000 cal. yr BP migration rates ranged from ~30 to 60 m/yr, with the rate decreasing from east to west. In contrast, from 12,000–10,000 cal. yr BP, migration rates were as much as 190 m/yr. Continued northward migration resulted in the arrival of lodgepole pine at the eastern Yukon border ~7000 cal. yr BP (i.e. eastern migration route). About 5000 cal. yr BP, migration in the eastern portion of Yukon was likely physically blocked by 200 km of mountainous terrain and presumably colder climatic conditions in higher elevation areas, except through a 15–20 km wide gap in the Frances Lake area (Figure 3). After passing through this gap, lodgepole pine likely migrated northwest down the Tintina Trench, which is occupied by the main stem of the Pelly River, during the subsequent 2200 calendar years. At least some migration probably also extended to the east and northeast up tributaries of the Pelly River. Lodgepole pine arrived at Buggy Pond near its present-day northern geographical range (site V, Figure 1) about 1790 cal. yr BP (1850 14C yr BP). The pace of migration accelerated from ~80 to 220 m/yr during the 6000–4000 and 3000–2000 cal. yr BP periods, respectively (Figure 3).
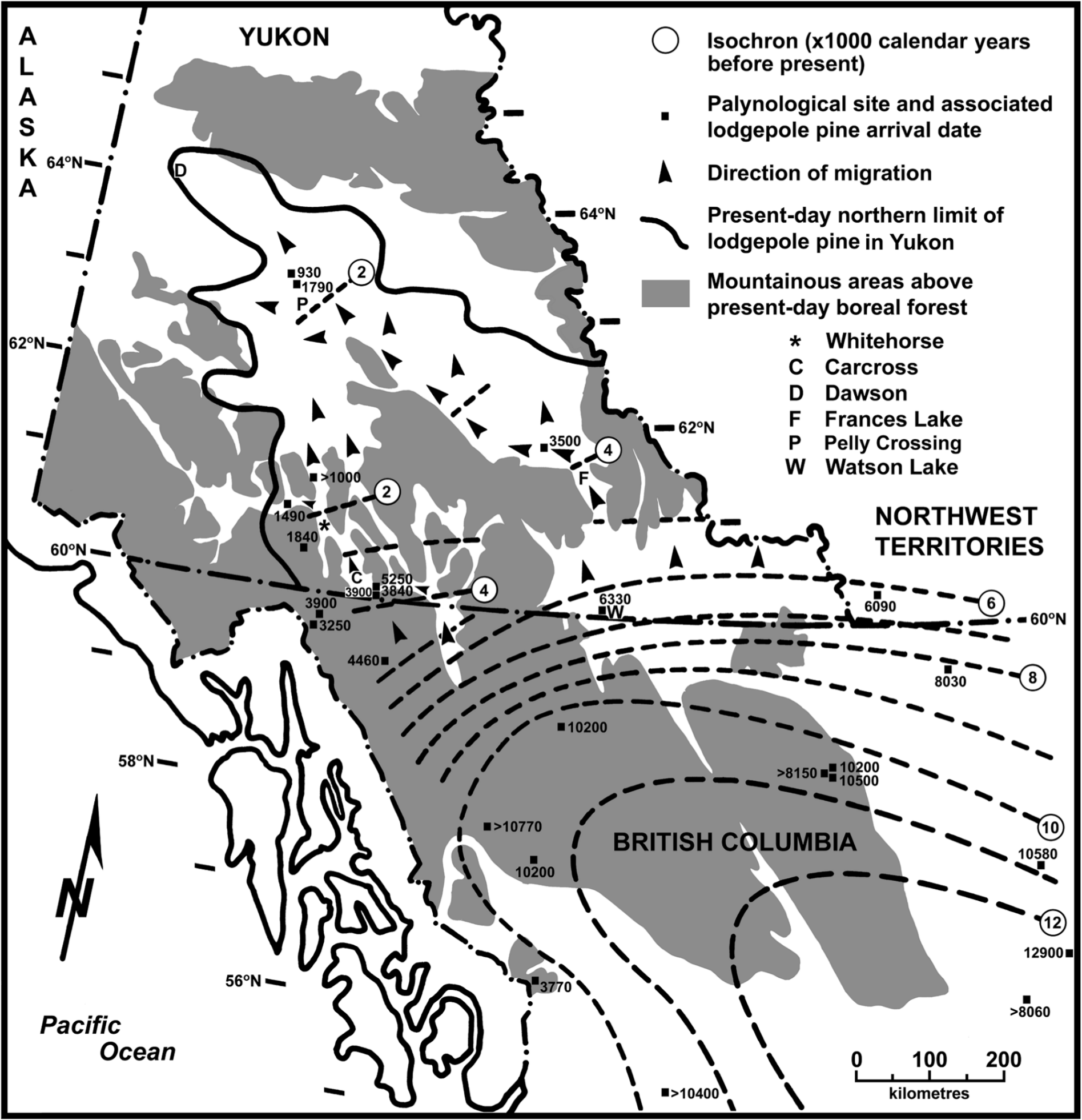
Holocene lodgepole pine (Pinus contorta var. latifolia) arrival dates in northern British Columbia and Yukon as represented by ≥5% pine pollen occurrence. Generalized isochrons represent calendar years before present as determined from 14C dating of depositional basin sediments. Shaded areas represent high-elevation topography where subalpine shrubland and alpine vegetation occur in the present-day environment (Meidinger and Pojar, 1991; Strong, 2013). Present-day occurrences of lodgepole pine are largely confined to lower elevations in shaded areas.
In northwestern British Columbia, the migration rate was ~30 m/yr between 10,000 and 5000 cal. yr BP. Lodgepole pine reached the Yukon border ~4000 cal. yr BP. Its subsequent migration into southwest Yukon (western migration route), like its eastern counterpart, encountered mountainous terrain with colder than ecologically acceptable temperatures at mid and high elevations. Therefore, further northward advance was probably confined to the lower elevations of the Yukon River and Teslin River valleys (Figure 3), which have several locations where the distance between mountain ridges is ≤15 km wide. Northward migration down these valleys was slower than along the eastern route (~70 m/yr) from 4000 to 2000 cal. yr BP. Lodgepole pine appears to have reached the Whitehorse area ~2200 cal. yr BP (Figure 3). From this location, it continued northward, but also spread westward up the Takhini Valley to arrive at Two Horsemen Pond (site W) 1490 cal. yr BP (1600 14C yr BP) (Figures 1 and 3). If the post-arrival development of lodgepole pine was similar at the Two Horsemen Pond and Cinquefoil-Dwindling sites (site X, Figure 1), lodgepole pine probably arrived in the Cinquefoil-Dwindling area ~1900 cal. yr BP (1950 14C yr BP).
Discussion
Arrival dates
MacDonald and Cwynar (1985: 1041) justified the use of 15% pine pollen content as an arrival threshold value based on prior work by MacDonald (1984: 163), who cited Ritchie (1974, 1984) as sources, and an unpublished document. The latter document was presumably later published by MacDonald and Ritchie (1986), who presented an extensive modern pollen rain data set for western Canada. None of these studies directly compared pollen rain composition and tree abundance; only regional plant presences were given in the MacDonald (1984) and MacDonald and Ritchie (1986) studies. Unfortunately, the limited number (n = 5) of near-surface limnic sediments in southern Yukon with >5% pine pollen content creates some uncertainty regarding the threshold level needed to represent the early occurrence of lodgepole pine trees within 30 km of a deposition site, although a good regression line fit occurred between percent pollen content and tree abundance when the former variable was <40% (Figure 2). Other studies have more rigorously compared pine pollen and tree abundances in proximity of the pollen sampling site. For example, (1) Jensen et al. (2002) reported 10% pine pollen content where trees formed 7% of the vegetation within 100 km of the sampled site (cf. Figure 2); (2) Webb et al. (1981) found through regression analysis that a 4.4% pine pollen content was associated with basins <9 ha in surface area (i.e. same as ≥21 of 25 analyzed sites in Table 1) when pine trees were absent within 30 km of a basin, and (3) by similar analysis methods, Prentice et al. (2008) reported a 5.5% pine pollen content in limnic surface sediments in the absence of pine trees within 50 km, although the content increased to ~15% for 5–10 km radius source areas (basins typical 20–70 ha); and (4) Froyd (2005) and Froyd and Bennett (2006) reported fossilized pine leaf stomata occurrences when pollen content values were as low as 0.4–1%. These studies strongly suggest that a pollen threshold value of 15% is much too high for recognizing pine arrival in a 30–60 km radius area, with a value between 0.4 and 10% appearing appropriate for small sedimentation basins in boreal forest environments. Based on Figure 2, the use of any pine pollen threshold value between 5% and 15% appears to represent a difference in tree abundance values that range from 1% to 3% within 30 km of a deposition sites, respectively.
Lodgepole pine appears to have been present near its northern limit in quantities of ≥1%, as represented by Buggy Pond data, ~1300 cal. yr earlier than previously estimated by MacDonald and Cwynar (1985); and 500 to 2570 cal. yr earlier at Waterdevil Lake, Kettlehole Pond, Snowshoe Lake, and Lac Ciel Blanc. MacDonald and Cwynar (1985) reported a lodgepole pine arrival date of 6375 cal. yr BP (5600 14C yr BP) at John Klondike Bog. Based on the profile data presented by Matthews (1980, Figure 2), however, pine pollen never exceeded 15% (MacDonald and Cwynar, 1985) to indicate arrival, except in surface samples. The reason for this discrepancy is unknown.
The differences in the first four previously indicated arrival dates stem from the use of >5% rather than a 15% pine pollen content as the indicator level. The two threshold levels potentially have different ecological implications. Pollen values slightly greater than 5% (i.e. long-distance transport plus local tree input) appear to denote the presence of 1% lodgepole pine trees in the Yukon landscape; thereby signifying an early stage of tree colonization, although not necessarily the very beginning. In contrast, 15% pine pollen content likely represents the common presence of lodgepole pine trees, which implies prior establishment and a prolonged period of local population expansion. Therefore, use of the higher threshold would underestimate the geographical extent of lodgepole pine trees (MacDonald, 1995: 39) and underestimate the species arrival date. The common occurrence of spruce (1) within the immediate proximity (e.g. <500 m, Sugita, 1994) of an assessed basin, because of site conditions that favor spruce but inhibit lodgepole pine (i.e. imperfect to poor drainage), and (2) in the surrounding landscape likely masked the presence and abundance of lodgepole pine within several kilometers of a pollen profile site. The proximity of spruce to the assessed basins is indicated by the occurrence of fossil spruce needles at sites A and E. The conclusion by MacDonald and Cwynar (1985: 1042) that the geographical range of lodgepole pine did not change whether a >5%, 10%, or 15% threshold was used was not refuted by the current analysis, primarily due to the low density of palynological studies that precluded a comprehensive delineation of the species’ palaeodistribution.
Several arrival dates among the analysed sites superficially appear to conflict (Figure 3), but are not necessarily inconsistent, with the general spatial pattern of lodgepole pine migration. For example, Berendon/Spillway Ponds (site R), Granger Ice Patch (site U), Sunset Lake (site I), and Drizzle Pond (site T) had late arrival dates compared to other nearby sites or adjacent isochron dates (Figure 3), but all occurred at high elevations on alpine sites. Therefore, more time was required for suitable environmental conditions to develop and for pine to colonize the subalpine areas below each site before sufficient pollen reached the deposition basin. The differences in arrival dates between Mile 16 Bog (site M, Anderson, 1970), and Kettlehole Pond (site Q, Cwynar, 1988) or Marcella Lake (site P, Anderson et al., 2005a) may simply reflect technological improvements in the accuracy of radiocarbon dating (Table 1) and sample collection procedures, or biases associated with the source material (e.g. MacDonald et al., 1991). Inadequate palynological data and radiocarbon dating information for Flamingo Lake (site Y) limited critical comparison of the site with Buggy Pond (site V, 8 km apart at similar elevations). In this case, the earlier date of arrival was used because no data refuted its validity. Except for Flamingo Lake and Mile 16 Bog, no major inconsistencies occur between the estimated arrival dates (n = 25 sites) and the general pattern of lodgepole pine migration presented in Figure 3.
Most post-1980 palynological studies in Table 1 used similar sediment coring and palynomorph processing techniques, so potential differences resulting from such mechanical and technical considerations should not be a major concern with respect to the presented migration scenario. One important consideration, however, might be the intensity of the original core analysis. Sampling along individual cores ranged from 1 to 20 cm intervals among studies. This range in sampling intervals was often related to core length (i.e. shorter cores had shorter intervals), with most studies typically having 40–50 samples per core (i.e. core length divided by sampling interval). Therefore, the accuracy of determining when pine exceeded 5% abundance was probably similar among pollen profiles.
Eastern Yukon migration and relative climatic conditions
The migration of lodgepole pine from the Canada–USA border to the northern-most portion of British Columbia (1520 km) took about 6340 cal. yr and represents an average advance of 240 m/yr, which is at least twice the normally expected seed-dispersal distance of 50–120 m (Despain, 2001; Lotan and Critchfield, 1990). However, an additional 8200 cal. yr were required for lodgepole pine to traverse the remaining 700 km to Buggy Pond. Only along the eastern route after 3000 cal. yr BP did migration approach the pre-10,000 cal. yr BP rates of advance. The slow migration after 9000 cal. yr BP, or post-Holocene Thermal Maximum period in northwestern North America (Kaufman et al., 2004), was followed by a 6–7°C decrease in summer temperatures (Clark et al., 2004; Lauriol et al., 2002), which was also reflected in colder lake water temperatures based on shifts in diatom assemblages in southwest Yukon (Chakraborty et al., 2010). As well, cordilleran glaciers retreated in the St. Elias Mountains (Denton and Karlén, 1973) and a substantial reduction of alpine ice patch abundance occurred from 8270 to 5530 cal. yr BP (7440–4780 14C yr BP) in south-central Yukon (Farnell et al., 2004). In combination, these environmental conditions suggest a colder and possibly more arid climate in southern Yukon and northern British Columbia after the thermal maximum (Pisaric et al., 2003), when migration was essentially static. Post-7000 cal. yr BP climatic warming would be needed to account for the resurgence of migration rates, although not well represented from 5000–4000 cal. yr ago in Figure 3 because of dampening of the rate by cooler local temperatures as lodgepole pine ascended to the Liard–Pelly river drainage divide. The occurrence of treelines above present-day elevations in and north of the St. Elias Mountains between 7000 and 4000 cal. yr BP provides evidence for warmer than present conditions (Denton and Karlén, 1973; Rampton, 1971).
The relatively rapid migration rates from 4000 to 2000 cal. yr ago in eastern Yukon imply even more favorable environmental conditions for lodgepole pine development than had previously occurred any time after 9000 cal. yr BP. From a broad perspective, the accelerated rate of migration might be at least partially linked to an eastward shift of the Aleutian Low pressure system in the North Pacific during the 4500–2800 cal. yr BP period, which would have created more arid conditions in interior Yukon than previously occurred (Anderson et al., 2005b). The 4000–2000 cal. yr BP period based on palynological evidence from within Yukon, however, has been described as having temperatures cooler (Anderson et al., 2005a – also see Cwynar et al., 2012; Bunbury and Gajewski, 2009; Kurek et al., 2009), comparable with (Rainville and Gajewski, 2012), or warmer (Szeicz and MacDonald, 2001), and moister (Anderson et al., 2005a; Bunbury and Gajewski, 2009) or drier (Cwynar, 1988; Cwynar and Spear, 1995; Pompeani et al., 2012) than those of the pre-4000 cal. yr BP period. The lack of a consistent pattern of change in the regional Yukon climatic regime contributes little to explaining why lodgepole pine migration rates more than doubled after 4000 cal. yr BP. Other possible explanations, regardless of the regional climatic regime, might be warmer local temperatures north of the Liard–Pelly river drainage divide because of low elevation terrain (e.g. ~2°C based on a normal temperature lapse rate of 0.65°C per 100 m decline in elevation), and possibly, the occurrence of the area in rainshadow (Jatzold, 2000) that might have been prone to wildfire and, therefore, more favorable for lodgepole pine. These effects are represented in the present-day landscape by the common occurrence of aspen (a warm site species) rather than just spruce, and by 1°C warmer annual temperatures than occurs in the surrounding areas at the same latitude (Strong, 2013). On a relative basis, present-day temperatures along the late-Holocene migration route (4000–2000 cal. yr BP) are only ~1°C colder than immediately north of the British Columbia border where lodgepole pine is most abundant in Yukon. Therefore, if the climate were only 1° or 2°C warmer than present, it is realistic to envision the common occurrence of lodgepole pine along the eastern migration route 4000–2000 cal. yr BP.
Relatively rapid northwest migration during the 4000–2000 cal. yr BP period suggests climatic conditions may have been favorable for lodgepole pine expansion beyond the lower elevations of the Tintina Trench, particularly northeast of the Pelly River. If lodgepole pine was once more abundant, this could explain the very broad present-day distribution of the species. However, a subsequent reduction in abundance would have had to occur to explain its present-day sparse and disjunct occurrence in peripheral as well as climatically marginal portions of its current range, and along the 4000–2000 cal. yr BP section of its migration route. Such a reduction appears to have occurred at Candelabra Lake (Cwynar and Spear, 1995) where pine pollen abundance decreased from 10–11% prior to 650 cal. yr BP to a present-day level of 4%, i.e. the equivalent of lowering treeline. At the same time, pine pollen abundance continuously increased after 1790 and 930 cal. yr BP at Buggy Pond and Flamingo Lake, respectively, which occur at lower elevations (Table 1). The date of decline at Candelabra Lake roughly corresponds with the beginning of the ‘Little Ice Age’ at ~600 cal. yr BP and extended to ~100 cal. yr BP (Cronin, 2009). Outside the present-day geographical range of lodgepole pine, as well as at Candelabra Lake, parallel decreases in (western white) spruce pollen abundance occurred after ~1000 cal. yr BP ago at high-elevation palynological sites such as Ittlemit Lake (1180 m, Wang and Geurts, 1991) and Honeymoon Pond (1160 m, Cwynar and Spear, 1991), and to a lesser extent Upper Fly Lake (1325 m, Bunbury and Gajewski, 2009); with reduced treeline elevations noted by Rampton (1971) in the Antifreeze Pond area. This suggests less favorable conditions for a species adapted to cold environments. The simultaneous increase of lodgepole pine at low while decreasing at higher elevation is not necessarily ecologically inconceivable, if climatic conditions remained above acceptable minima and suitable vacant niches were available at lower elevations (e.g. post-fire sites). Remnants of a possibly once larger lodgepole pine population at higher elevations would have been isolated on favorable sites such as ridge crests, southfacing slopes, and coarse-texture substrates as seen in the current landscape when temperatures cooled. If present-day climatic conditions are the benchmark for comparing palaeoclimates, it should be noted that average annual and summer temperatures increased ~3°C and 1.4°C between 1956 and 2010 (r = 0.57 and 0.50, P <0.01), respectively, based on the Pelly Ranch meteorological station located ~55 km southwest of Buggy Pond (ID 2100880, 62°49′N, 137°22′W, Environment Canada, 2012), although warming likely began after the start of the 20th century (Szeicz and MacDonald, 1995: 265). Therefore, cooling during the ‘Little Ice Age’ could have substantially reduced lodgepole pine abundance at higher elevations, if conditions were even warmer than present-day from 4000 to 2000 cal. yr BP.
West Yukon migration
If present-day vegetation and climatic conditions can be used as a guide, lodgepole pine migration in northwest British Columbia and southwest Yukon was constrained by mountains and cold climatic conditions at mid and high elevations. Specific routes through northwest British Columbia are unknown. The increased rate of migration after 5000 cal. yr BP may reflect movement of lodgepole pine from a strongly cordilleran to a more boreal climatic regime, which was ecologically more favorable. Once lodgepole pine reached Yukon, its northward progress continued to be slow compared with the eastern route, most likely because of generally colder summers (cf. Strong, 2013: figure 2). Progress was sufficiently delayed that lodgepole pine passed through the Whitehorse area as its eastern counterpart approached Buggy Lake, 270 km further north (Figure 3). As a result, most of the lodgepole pine range in Yukon appears to have been populated by trees that past through the Watson Lake area. Although the data are sparse, leaf oil terpenes (von Rudloff and Nyland, 1979) and genetics (Fazekas and Yeh, 2006: figure 3; Godbout et al., 2008, i.e. 2b versus 3a group) of lodgepole pine in the western migration route hint at a deviation from those present along the eastern route. Such differences tend to support the likelihood of two spatially separated northward-migrating segments of the population in northern British Columbia, rather than a lateral transfer of plants from the Watson Lake to Carcross areas.
Migration rates and ecology
During more than half the migration time between 49° and 63°N latitude, lodgepole pine advanced northward at 220–240 m/yr, with an overall average of ~153 m/yr. The latter rate was greater than the maximums determined for other tree species such as beech (Fagus grandifolia Ehrh.) and red maple (Acer rubrum L.) in the eastern USA (McLachlan et al., 2005) and Norway spruce (Picea abies (L.) Karst., Ravazzi, 2002) in Europe at ≤100 m/yr. The typical migration rate for lodgepole pine was similar to the maximum determined for Douglas-fir (Pseudotsuga menziesii (Mirb.) Franco, ≤220 m/yr, Gugger and Sugita, 2010), which often co-occurs with lodgepole pine in present-day forests and might have occupied similar mountainous areas of the western USA during the Wisconsinan glacial maximum. In contrast, jack pine (Pinus banksiana Lamb.), which is a fire-dependent species with similar morphological and ecological attributes as lodgepole pine, appears to have migrated across the low relief terrain of central Canada at a rate of ~380 m/yr (McLeod and MacDonald, 1997). These differences in migration rates suggest lodgepole pine was typically a moderately fast spreading species during the mid and late Holocene. The principal anomaly in its migration rate was a substantial slowing (e.g. <50 m/yr) from 10,000 to 6000 cal. yr BP. Although this period of slow advance can largely be attributed to an abrupt cooling of the regional climate following the Holocene Thermal Maximum (Clark et al., 2004), other factors such as its high latitudinal location with an associated cooler northern climate during this period of advance and higher topographic elevations may have also been contributing factors. The latter factor might explain the much slower pace of migration in northwest British Columbia compared with its more eastern counterpart (Figure 3).
Lodgepole pine migration north of 56°N latitude did not directly follow deglaciation, but rather occurred several thousand calendar years after an advance and domination of the landscape by white spruces (e.g. Picea albertiana and P. glauca (Moench) Voss) (e.g. see Hail Lake – Cwynar and Spear, 1995). The rate of early-Holocene spruce migration was likely determined by the rate of deglaciation and probably involved the colonization of pre-existing non-arboreal vegetation. In contrast, the shade-intolerant nature of lodgepole pine would not have allowed the species to replace established shade-tolerant spruce forests through direct competition. It is unlikely that the environment became ecologically unfavorable, thereby causing the demise of spruce, because white spruce species have a broad ecological amplitude and are capable of growing on the same site conditions as lodgepole pine. Therefore, some form of landscape perturbation, which would favor lodgepole pine, such as an increase in aridity and wildfire frequencies (Cwynar, 1988; Pisaric et al., 2003) would have been necessary to at least temporarily eliminate spruce forests from a portion of the landscape to allow lodgepole pine colonization. The lack of substantial lodgepole pine range expansion from 10,000 to 6000 cal. yr BP suggests a period of sufficiently moist conditions (e.g. Cwynar, 1988) occurred that minimized wildfire frequencies in northern British Columbia.
Conclusion
The proposed migration scenario accounts for most of the variance among known lodgepole pine arrival dates in northern British Columbia and Yukon. The available palynological data indicate that lodgepole pine first arrived in Yukon at least 7000 cal. yr ago and appears to have moved northward by separate east and west routes because of obstruction of migration by mountains in the southern portion of the territory. Lodgepole pine reached its near northern limit at Buggy Lake at least by 1790 cal. yr BP, which is earlier than previously estimated. Some of the differences in migration rates in northern British Columbia and eastern Yukon correspond with palaeoenvironmental changes reported in other studies. Explanation of the broad current range of lodgepole pine in Yukon depends upon whether a slow systematic spread, or a rapid expansion and subsequent contraction of the population occurred after 4000 cal. yr BP. Based on the establishment ecology lodgepole pine, the intervention of wildfire was likely necessary to displace spruce along its northern British Columbia–Yukon migration route. The influence of topography and elevation-induced climates should be considered when attempting to interpret plant migration patterns and directions, particularly in mountainous terrain. Several detailed pollen stratigraphy studies that extend back in time at least 5000 cal. yr BP will be needed to understand the potentially interesting history of lodgepole pine along the 4000–2000 cal. yr BP section of its eastern migration route. Such analyses might answer the question of whether lodgepole pine was once much more extensive than it is on the current landscape. More palynological research is also needed to elucidate the migration routes of lodgepole pine in northwestern British Columbia during the mid Holocene.
Footnotes
Acknowledgements
Yukon Forest Management Branch provided forest cover mapping and the Yukon Energy Mines and Resources Library acquired various locally unavailable publications for this study. Dr Jennifer Galloway (Geological Survey of Canada) provided constructive comments on an earlier draft of the manuscript, Dr Cindy Froyd (Swansea University) provided helpful input, and two anonymous reviewers provided comments that improved the clarity of the manuscript.
Funding
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.