
Abstract
Recent archaeological and archaeobotanical data suggest a very long tradition for the broad-spectrum subsistence economy in North China. It can be traced back at least to the Upper Paleolithic, in the last glacial maximum, and it continued into the early Neolithic period. Subsistence strategies also show great regional variation, suggesting a complex mosaic of adaptations in the transition to agriculture. The research reported here focuses on the plant-derived subsistence economy of the earliest Neolithic communities in the Daihai Lake area, Inner Mongolia, where the ecosystem was sensitive to climatic fluctuations. Neolithic groups likely migrated to the region as part of population expansion from the Central Plain. Previous scholars have suggested that this expansion was due to a search for agricultural land for millet farming. By examining residue remains and usewear patterns on sandstone grinding stone tools unearthed from the Shihushan I and Shihushan II sites, dating to the mid-5th millennium
Introduction
Recent archaeological and archaeobotanical data, particularly based on residue and usewear analyses on grinding stone tools, suggest a very long tradition for the broad-spectrum subsistence economy in North China. It can be traced back at least to the Upper Paleolithic period, c 23,000 cal. BP, during the last glacial maximum of the late Pleistocene, in the Yellow River region of North China. People exploited many types of plant resources, including wild millet, wheat grass, beans, tubers, and nuts (Guan et al., 2012; Liu et al., 2011, 2013a). This subsistence pattern seems to have continued in the region during the early Neolithic period (c. 9000–7000 cal. BP) when sedentary villages were built and millet domestication was underway, suggesting a persistent practice of plant gathering as well as low-level food production (Liu et al., in press, 2011; Tao et al., 2011). These observations are also in line with the previous studies suggesting that the transition from the first emergence of cultigens to intensive agriculture was a very long process in many regions worldwide (Piperno and Pearsall, 1998; Smith, 2001). Since China’s climate and ecosystems are very diverse, various regions are likely to have gone through different pathways to agriculture. It is important, therefore, to examine adaptation strategies at a micro-regional level, to understand how Neolithic people exploited and managed resources in particular environments.
The current research investigates the subsistence strategies of Neolithic communities in the Daihai Lake area, central-south Inner Mongolia, a region where the ecosystem has been sensitive to climatic fluctuations and agriculture has been marginal in the past. Using residue remains and usewear patterns on grinding stone tools unearthed from two sites, Shihushan I and Shihushan II (SHSI and SHSII hereafter), we examined the exploitation of plants by the earliest Neolithic populations there. The results of our research show that the earliest Neolithic settlers in the Daihai region appear to have enjoyed a way of life including the use, and possibly management, of a wide range of wild plants, including various underground storage organs (USOs; tubers, roots, rhizomes, and bulbs), nuts, and wild grasses, while engaged in a limited level of millet production. This study adds to a growing literature that questions the economic significance of early cereal crops in subsistence system (Bowles, 2011; Greaves and Kramer, 2014; Jones et al., 2013), and suggests the consideration of the role of other plants during the period when millets were domesticated in North China.
Environmental and archaeological background
SHSI and SHSII were discovered in 1993 and are currently the earliest Neolithic settlements in the Daihai region (Figure 1). They were excavated by the Inner Mongolia Institute of Archaeology and a Sino-Japanese archaeology team in 1995 and 1996, in order to understand the beginning of farming in this region (Daihai Region Investigation Team, 2001). The sites, 300 m apart, are located on low hills, 2.5 km south of the Daihai Lake (Figure 1). The lake is situated in the transitional region between semi-humid and semiarid areas in the middle temperate zone of China. Mean annual temperature is 5.1°C, and mean annual precipitation is 423 mm. The modern natural vegetation of the Daihai basin belongs to the southern temperate steppe, composed of forest on the mountains, alpine meadows, grasses and herbs on the hilly areas, and meadows over the lakeshore plain and the frontal fringes of diluvial fans. Economic herbs growing in the region and relevant to this study include Lilium sp., Setaria viridis, and Elymus dahuricus.
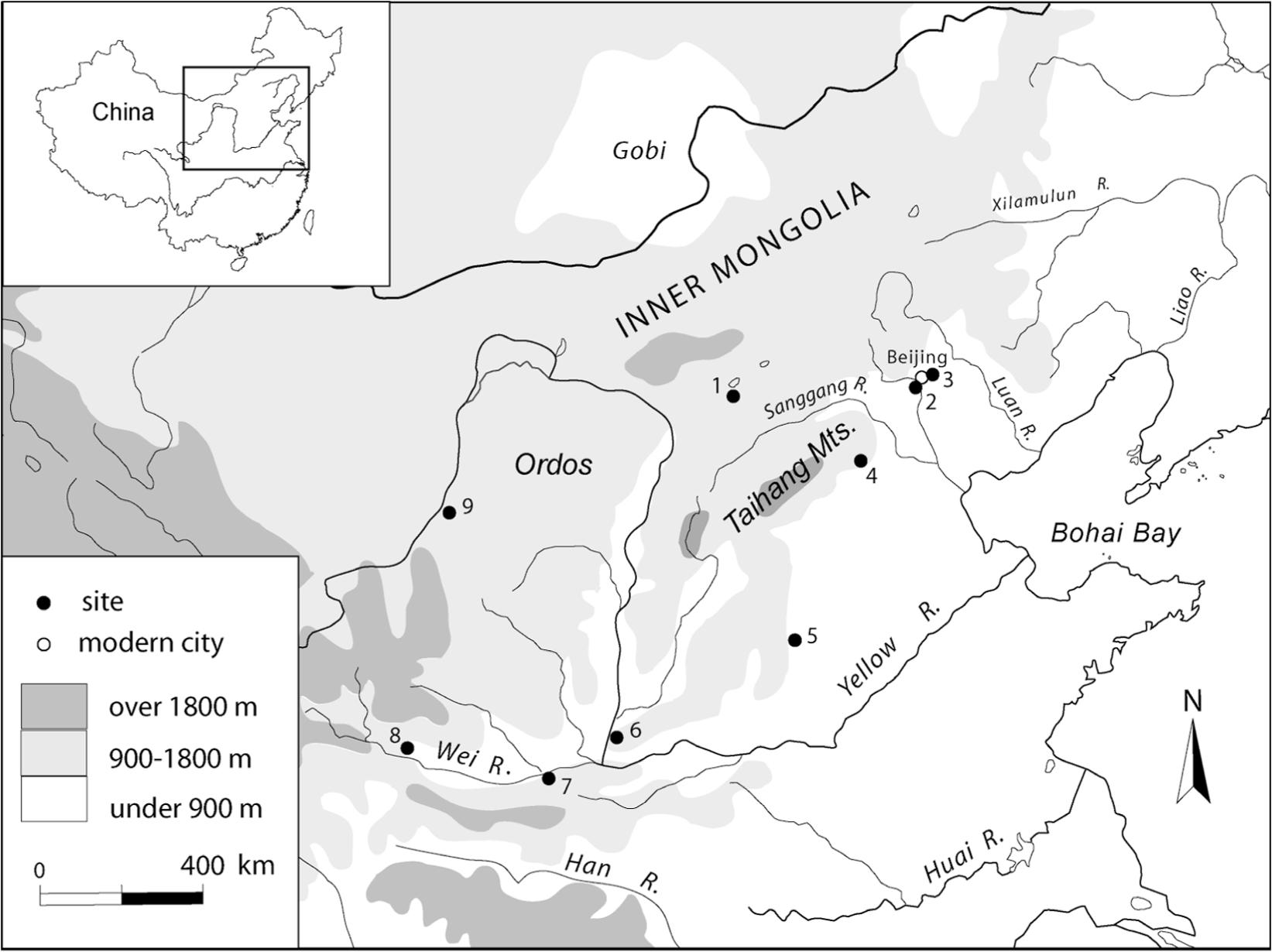
Location of sites mentioned in this paper. (1) Shihushan, (2) Zhenjiangying, (3) Shangzhai, (4) Beifudi, (5) Cishan, (6) Shizitan, (7) Banpo, (8) Dadiwan, and (9) Shuidonggou.
The pollen record obtained from the Holocene sediments in the lake indicates that the middle Holocene period between c. 7900 and 4450 cal. BP was marked by a warmer and more humid climate than the preceding period. The surrounding hilly regions were covered with large-scale mixed coniferous and broad-leaved forests, dominated by Pinus and accompanied by Quercus and Ostryopsis. Aquatic plants include Typha sp. (Xiao et al., 2004; Xu et al., 2004). The average annual temperature was 2–3 °C higher, and the water level of the lake was 20 m higher than at present. This mid-Holocene Climatic Optimum period, however, was punctuated by several episodes of dryer and colder conditions, dating to c. 6400–6000, 5000–4800, and 4400 cal. BP (Tian and Guo, 2004).
SHSI (6530–6440 cal. BP) dates slightly later than SHSII based on ceramic typology. These two settlements appeared in the region during a warmer and slightly more humid episode, as identified in the lake sediments. The percentage of tree pollen decreased, and the herb pollen percent increased compared with the previous period. Pinus pollen accounts for 9.5–36.5%, Quercus for 0.4–13.1%, and Artemisia for 35.9–69.2% of the pollen sum, suggesting the expansion of grassland vegetation (Xiao et al., 2004; Xu et al., 2004). Periodic abandonment of the settlements in the region appears to have coincided with climatic fluctuations (Tian and Guo, 2004).
SHSII measures 0.3 ha in size. Within the site, archaeologists discovered 14 house foundations, 23 pits, and one burial within the excavated area of 1545 m2. At SHSI (1.5 ha), the excavated area of 1270 m2 revealed 6 house foundations, 12 ash pits, 1 sacrificial pit, and a surrounding moat. At both sites, numerous stone, pottery, bone, and shell artifacts have also been found. Stone tools include spades, axes, adzes, knives, grinding stones, and microliths.
The sites were partially destroyed by soil erosion, and the remaining occupational deposits were thin, measuring 0.15–0.35 m at SHSII and 0.1–0.4 m at SHSI in thickness. Residential houses were rarely overlapping, suggesting that settlement occupation was relatively short (Daihai Region Investigation Team, 2001).
The material assemblages of SHSII are similar to those of the Neolithic sites at Zhenjiangying in Beijing and Beifudi II in Hebei (5000–4700
In these two regions, which appear to have been the ancestral homes of the SHS populations, Neolithic communities were already engaged in the domestication of millet, pig, and dog prior to the Yangshao Period (Liu and Chen, 2012: 75–122; Zhao, 2011). For example, in the North China Plain, considerable quantities of millet remains have been uncovered from ash pits at Cishan in Hebei (6100–5750
Like the Yangshao in the Central Plain, the Shihushan people raised pigs and dogs. However, pig and dog bones only account for a small percentage of the faunal remains (210 of 2306, 9% in NISP; 18 of 138, 13% in MNI). The sources of animal protein for the populations seem to have been primarily derived from hunting, including several species of deer and water buffalo (1927, 83.5% in NISP; 98, 69.6% in MNI; Huang, 2001).
Excavations at both sites have yielded significant numbers of sandstone grinding stones (mopan slabs and mobang elongate handstones), mostly from pits, house floors, and the moat. At SHSI, 55 of the 157 stone tools are grinding stones (35%), so are the majority of the 83 stone tools from SHSII (no numerical information is available in the report). In some houses, multiple grinding stones were found on the floors (Daihai Region Investigation Team, 2001). All the slabs are fragments, and many show a concave surface, apparently due to use. Handstones are mostly elongate in shape, also showing clear usewear traces. Similar mopan and mobang grinding stones have been widely found in Upper Paleolithic and early Neolithic sites of northern China, and previous studies have demonstrated that they were used primarily for processing plant foods (Liu et al., in press, 2010a, 2010b, 2011, 2013b; Tao et al., 2011; Yang et al., 2012).
It has been suggested that the Shihushan people practiced agriculture, as indicated by the presence at the sites of large numbers of grinding stones and spades (Daihai Region Investigation Team, 2001: 140). Nevertheless, none of these tools have been analyzed, and no flotation was done during the excavations (no macrobotanical samples were systematically collected). Thus, there is as yet no direct evidence of the plant-derived subsistence economy of these Neolithic people.
In view of these observations, the current project, using residue and usewear analyses to examine the grinding stones, aims to investigate the function of these tools, and furthermore, to provide a better understanding of plant-based subsistence strategies of the Shihushan communities, who were the Neolithic pioneers to this environmentally fragile region, the northernmost area of the Yangshao cultural distribution.
Analytical methods
The SHS artifacts are stored in the Jining Museum, Inner Mongolia, where we collected residue and usewear samples from 10 grinding stones in the summer of 2012 for laboratory examination (Figure 2 and Table 1).

Grinding stones analyzed. SHSI: GS1 and GS2; R1 and R5. SHSII: GS3–GS5; R2–R4.
Shihushan grinding stone sampling record.

Residue analysis
A total of 10 residue samples were extracted from the tools either by applying a small amount of distilled water to the tool and then extracting the sample with a pipette, or by submerging a part of the tool in distilled water in a plastic bag, and shaking the bag in an ultrasonic bath for 3 min to remove the residues on the tool surface (Fullagar, 2006b; Loy and Fullagar, 2006). Residue samples were processed for starch and phytolith extraction using the heavy liquid sodium polytungstate (in a specific gravity of 2.35). Extractions obtained from residue samples were mounted in 25% glycerol and 75% distilled water on glass slides.
For starch analysis, the slides were scanned under a Zeiss Axio Scope A1 fitted with polarizing filters and differential interference contrast (DIC) optics. Images were taken using a Zeiss Axiocam HRc3 digital camera and Zeiss AxioVision software ver 4.8. There are over 900 plant specimens in our modern reference collection at the Stanford Archaeology Center. We have specifically analyzed those starch-rich and economically important samples relevant to the research area, including more than 160 samples belonging to 84 species in 46 genera of 19 families.
Phytoliths were counted using the same slides prepared for starch grain analysis. A Zeiss Axioskop with polarizing filters and a QI-Imaging camera were used to identify and record the phytoliths. The phytolith reference collection includes more than 1200 modern plant specimens, selected both for the economic uses and their habitat specificity for various regions in Southeast Asia and China. Collections focus on the families and genera that produce the most abundant phytoliths (Poaceae, Arecaceae, Zingiberaceae, Magnoliaceae, etc.; Kealhofer and Piperno, 1998; Piperno, 1988, 2006).
Usewear analysis
Polyvinyl siloxane (hereafter PVS or peel) was applied to the grinding stones to provide portable and durable records for microscopic analysis. Previous research, using PVS on usewear patterns from grinding stones in China (Liu et al., 2010a, 2010b, 2011) and other parts of the world (Dubreuil, 2004; Field and Fullagar, 1998; Fullagar, 1991, 2006a), established valuable reference data for the study of ancient tools.
Usewear analysis does not always determine specific materials being processed on the tools, and researchers have used blind-test methods to check the basic parameters and improve the methodology (Stevens et al., 2010). Hamon and Plisson (2008) employed blind tests, using a stereoscopic microscope, to study grinding stone usewear. Their results suggest that actions (grinding, crushing, etc.) constitute the most reliable parameters of determination, followed by the texture of material being processed (soft, hard, powder, etc.).
In recent years, we have also conducted a series of experimental studies, although not blind testing, on sandstone grinding stones which were recorded with PVS to create a permanent record of the experimental results of usewear derived from processing seeds, tubers, nuts, wooden objects, minerals, shells, and stone implements (Fullagar et al., 2012; Liu et al., 2011, 2013a). Our results also show that usewear traces from processing soft plants, hardwood, and lithics are clearly distinguishable (see below). Based on these studies, the analytical variables examined in the current project include the following: stage of polish development (low, medium, and high), polish reticulation pattern, polish topography, striations (furrow, or V-shape in cross section; sleek, or U-shape in cross section; fine), pitting and pecking, and surface micro-topography.
Usewear analysis was employed here to help understand both the type of action for the work undertaken (grinding, pounding, etc.), and the general category of materials being processed (soft plants, wood, stone, etc.). The results are compared with the residue analysis to provide a more detailed identification of the types of materials that were worked on the grinding stones at the SHS sites.
PVS samples were taken from different locations on the tool to document used surfaces. The peels were examined under a compound (reflected light) Zeiss microscope at magnifications of 50×, 100×, 200×, and 500× and compared with reference peels. Images were collected with a Zeiss Axiocam HRc3 digital camera and Zeiss AxioVision software ver 4.8.
Starch grain analysis
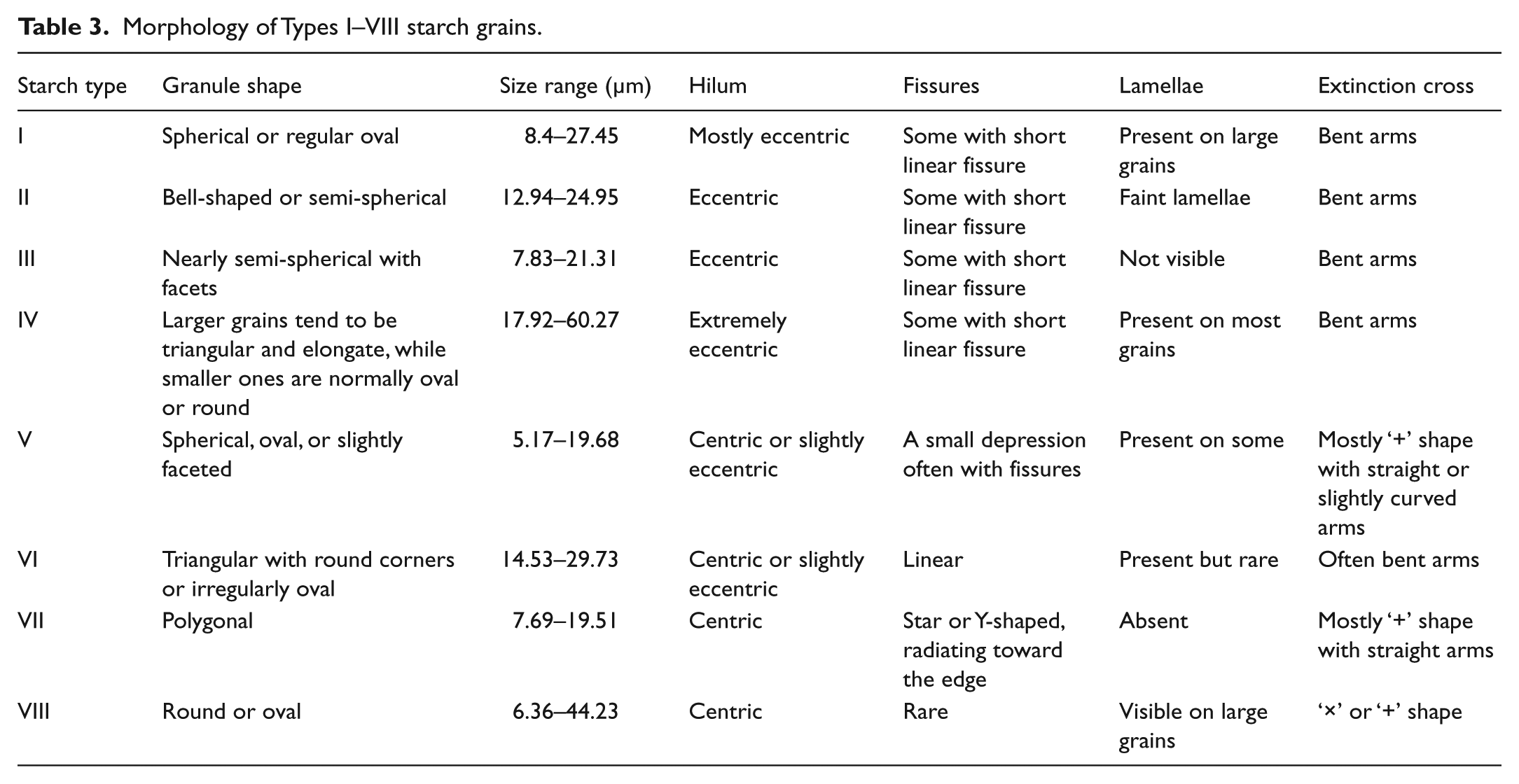
A total of 485 starch grains have been uncovered from the tools (363 from SHSII and 122 from SHSI). Of these, 468 (96% of the total) are identifiable to various taxonomic levels when compared with our reference material. Nearly half of the starch grains show traces of damage (220; 45% of the total), characterized by broken edges, deep fissures, pronounced lamellae, rough surface, and/or a dark central area on the extinction cross (Table 2). These features are similar to those of ground plant samples in our reference collection, indicating that the starch remains are mostly associated with the tool function, an inference consistent with previous studies (e.g. Babot, 2003; Haslam, 2004; Liu et al., 2013a). The ancient starch grains are classified into eight types on the basis of their morphology and size, and Table 3 provides detailed description of the diagnostic features for each type. The unidentifiable starch grains lack diagnostic features comparable to available reference material.
Shihushan starch counts.

Morphology of Types I–VIII starch grains.
Types I–III starch
Type I starch grains (N = 82) are spherical or regular oval in shape, Type II (N = 11) is characterized by a bell or semi-sphere shape, and Type III (N = 13) consists of compound grains of twins, triplets, and quadruples (Figure 3 and Table 3). Types I–III starch grains are most comparable in morphology and size with the root of snakegourd (Trichosanthes kirilowii in the Cucurbitaceae family) in our reference data. The starch grains from this plant are varied in shape, and include spherical, oval, semi-spherical, bell-shaped, and polygonal morphologies. Compound starch is composed of various numbers of grains, mostly between two and four. The hilum is eccentric in most cases, fissures in various forms are sometimes present, lamellae are visible on some large grains, and the extinction cross often has bent arms (Figure 4). When sizes are compared, Types I–III also closely resemble the range of ground T. kirilowii dry root (Figure 5). Types I–III grains together, therefore, are identifiable as T. kirilowii root, accounting for 22% of the starch assemblage (N = 106).

Starch types from Shihushan (under DIC and polarized filters for each granule). (1 and 2) Type I, spherical and regular oval, showing damaged granule (2); (3) Type II, bell-shaped; (4–6) Type III, cluster of granules in twins, triplets, and quadruples (Types I–III: Trichosanthes kirilowii snakegourd root); (7–9) Type IV, oval and triangular shaped with very eccentric cross (Lilium bulb and/or Dioscorea yam), showing damaged granule (8); (10–12) Type V, compound and individual spherical and oval (Typha sp.); (13) Type VI, triangular shaped with round corners (Quercus acorn); (14) Type VII, polygonal with centric extinction cross (Panicum millet); and (15) Type VIII, regular oval with centric X-shaped cross (Triticeae grass) (scale bar 8 and 9: 20 µm; others: 10 µm).

Starch references relevant to this study (under DIC and polarized filters). (1–3) Trichosanthes kirilowii (Henan, Shanxi), showing various starch morphology and compounds, for example, (1) spherical, semi-spherical, bell-shaped; (2) twins; (3) triplets; (4) Lilium pumilum (Yan’an, Shaanxi); (5) wild Dioscorea opposita (Yanshi, Henan); (6) Setaria italica ground (Shanxi); (7) Agropyron cristatum, ground (Inner Mongolia); and (8) Typha orientalis (Australia; scale bar 2, 3, and 6: 10 µm; others: 20 µm).

Ancient starch size compared with modern samples.
Type IV starch
Type IV starch grains (N = 142, 29% of the total) are characterized by a large grain size range with various shapes and a very eccentric hilum on large granules (Figure 3 and Table 3). These starch grains resemble those from Lilium sp. bulb and Dioscorea opposita yam (Figure 4). These plants have both wild and cultivated variations in North China. In Inner Mongolia today, only cultivated D. opposita grows in Wulanchabu, but five wild species of Lilium are widely distributed (Ma, 1985), and at least two species (L. concolor var. and L. pumilum) are found in the Daihai area (Ma, 1985; Xiao et al., 2004). When compared with our reference data, Type IV starch grains best match, in morphology and size, L. pumilum and L. tigrinum samples collected from Shaanxi. Some of the large triangular-shaped Type IV grains also resemble wild D. opposita samples, thus we cannot rule out the possibility that some Type IV starch grains also belong to D. opposita. We therefore identify Type IV starch as Lilium sp. and/or D. opposita (lily/yam).
Type V starch
Type V starch grains (N = 107, 22% of the total) are spherical, oval, or slightly faceted in shape. The grains appear either as compound or as individual. The compound grains are often blurry, but when the surface morphology is visible on some grains, it is consistent with those of individual ones (Figure 3 and Table 3). Type V starch grains best match those of the rhizome of Typha spp. in our reference data, which contains both compound and individual grains (Figure 4). The size range of Type V also falls into the ranges of two modern Typha species (T. orientalis, 4.52–19.09 µm; T. latifolia, 3.47–19.4 µm, after grinding; Figure 5), both can be found in Inner Mongolia today. Typha was also present in the Daihai pollen data dating to mid-Holocene (Xu et al., 2004).
Type VI starch
Four grains (1% of the total) are classified as Type VI starch, which are either triangular or irregularly oval in shape (Figure 3 and Table 3). These features mostly resemble several Quercus species (Figure 4), such as Q. variabilis, Q. liaotungensis, and Q. mongolica, which grow in North China today (Yang et al., 2009b). When sizes are compared, Type V starches show best matches to a ground Q. variabilis in our reference collection (Figure 5).
Type VII starch
Type VII starch grains (N = 9, 2% of the total) are characterized by small sizes and polygonal shapes (Figure 3 and Table 3). Damaged grains are similar in morphology to ground foxtail and broomcorn millets (Setaria italica ssp. italica and Panicum miliaceum) in our reference collection (Figure 4). The size range also falls into data for the experimental ground foxtail millet (Figure 5). Given that two millets were domesticated several thousand years before the SHS occupations (Crawford, 2006; Zhao, 2011), it is likely that Type VII grains include domesticated foxtail millet.
Type VIII starch
Type VIII starch grains (N = 59, 12% of the total) are round or oval in shape, with a broad size range (Figure 3 and Table 3). These starch grains, in morphology and size, resemble many taxa in the Triticeae tribe of the grass family indigenous to North China. This includes genera of Agropyron, Roegneria, and Leymus in our references. Several species of Agropyron and Leymus can be found in the Daihai area today based on our fieldwork and published data (Xu et al., 2004). The characteristics of damaged grains are also consistent with those found in ground Leymus and Agropyron from our references. It is possible that Type VIII starch grains belong to more than one genus in the Triticeae tribe, and the large grains resemble Agropyron cristatum and Agropyron desertorum in form and size, particularly after grinding (Figures 4 and 5).
Other starch grains
A total of 37 starch grains (12%) were identified as USOs. They show general features, which are shared by Trichosanthes, Lilium, and Typha, such as round shape, eccentric hilum, and bend arms of extinction cross. A total of 17 grains (4%) are unidentifiable due to the lack of diagnostic features which match to our reference samples.
Phytolith/microfossil analysis
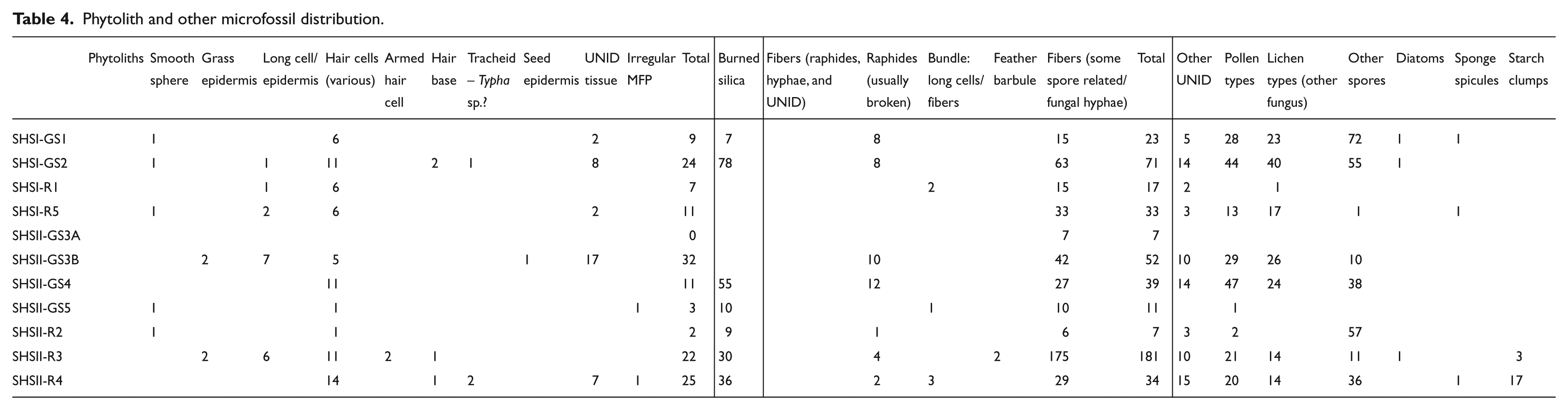
Beyond starch grains, the microfossil distribution for the grinding stones is provided in Table 4. These microfossils provide two discrete types of information, the first related to artifact use and the second related to artifact taphonomy. The phytoliths, starch grains, and fibers appear to be directly related to usewear based on the assemblage patterns. The spore, lichen, and pollen evidence is arguably more difficult to interpret, but it seems likely that the spores, at least, are related to post-depositional processes either on site or while in storage. The distribution of burned silica generally parallels the abundance of phytoliths on the grinding stones. The presence of burned plant material would seem to reflect both preservation and the domestic context of tool use (e.g. near hearths), as plant processing often occurs prior to cooking/roasting.
Phytolith and other microfossil distribution.
Residues
The phytolith counts for most of the artifacts were quite low (<30 per sample). In most cases, the phytoliths were not diagnostic. The dominant phytolith type was the hair cell, in several different forms (Table 4 and Figure 6a–c). Hair cells were most common on the SHSII artifacts, specifically GS4, R3, and R4, but SHSI-GS2 had a comparable number. R3 also had a few more diagnostic hair cells (‘armed hair cells’) that in some cases are consistent with the Trichosanthes sp. identifications based on the starch grains (Kealhofer and Piperno, 1998); however, armed hair cells are present in a variety of other taxa (also present in the regional pollen profiles). Both R3 and R4 had hair base phytoliths that also are consistent with Cucurbitaceae types, but occur in other taxa as well (e.g. Kealhofer and Piperno, 1998; Morris et al., 2009: Plate 1; Piperno, 2006). Small pieces of unidentified tissue were also common on several of the artifacts. The remaining phytolith types included smooth spheres, tissue fragments (Figure 6d), tracheids, and long cells (Figure 6f). While, in general, tracheids are not particularly diagnostic, those found on GS2 are very similar to Typha sp. tracheids (Guo et al., 2012). Very few grass-related phytoliths (particularly from glumes) were identified.
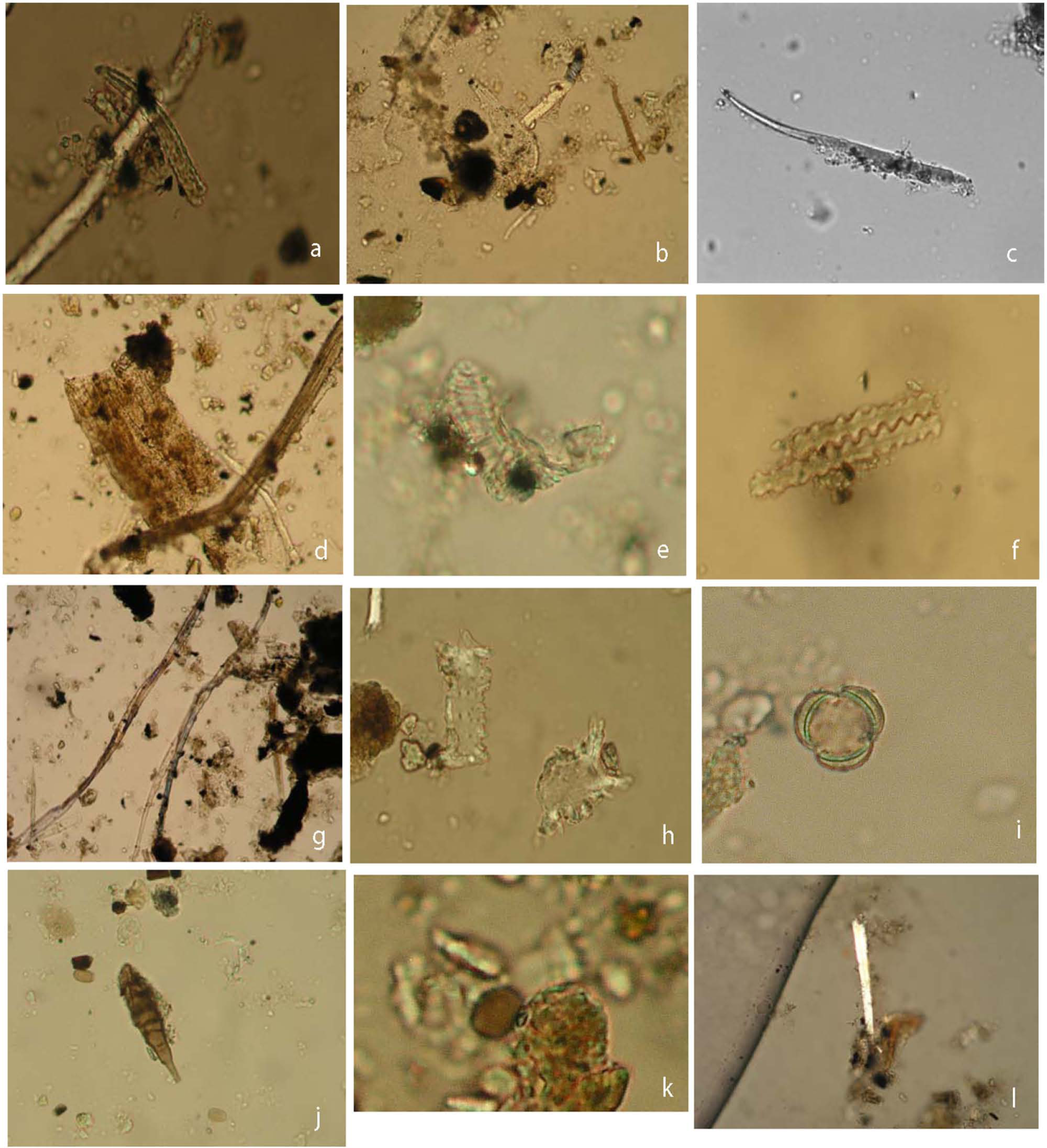
Phytoliths and other microfossil remains recovered from Shihushan grinding stones. (a) SHSII-R3 armed hair cell 60 µm (400×) enlarged, (b) SHSII-R3 hair cell 125 µm (400×), (c) SHSII-R4 hair cell 150 µm (400×), (d) SHSII-R3 UNID tissue (200×), (e) SHSI-GS2 tracheid (400×) enlarged, (f) SHSI-R5 epidermal long cells 90 µm (400×), (g) SHSII-R3 fibers (200×), (h) SHSII-GS3b seed (?) epidermis (2; 400×), (i) SHSII-R3 Brassica-type pollen 15 μm (400×) enlarged, (j) SHSI-GS1 lichen-type spore (cf. Alternaria sp.; 400×), (k) SHSII-GS4 mushroom-type spore (cf. Agaricus sp.; 400×) enlarged, and (l) SHSII-R3 raphide 125 µm (400×).
The plant ‘fibers’ present are not specifically diagnostic (Figure 6g; https://fril.osu.edu/index.cfm, Fiber Reference Image Library); however, their distribution is strongly patterned. Their morphology, in most cases, is consistent with root hairs (e.g. Typha sp., see Acharya et al., 2013), while others are similar to fungal hyphae. Comparisons with reference material from Typha, Trichosanthes, Lilium, and Dioscorea, all of which produce fibers, indicate that all but Lilium are likely sources of the archeological fibers. However, other sources cannot be ruled out. Fibers are present on all of the SHSI artifacts, but most abundant on GS2. They are also present on all SHSII artifacts, but clearly dominant on R3. A few broken raphides were found in association with the grinding stones. Both Dioscorea yam and Typha rhizome produce abundant raphides. The presence of both root hairs and raphides, likely in association, support the processing of USOs on these grinding stones. In terms of intensity, GS2 and R3 reveal the most evidence for USO processing.
Other microfossils
In addition to phytoliths and starch, a variety of other types of microfossils are present on tools, particularly pollen, lichen types, and other spores. The pollen assemblage includes one Pinaceae grain, a few grass pollen grains (Poaceae), and a high frequency of 3C3P type pollen (Figure 6i). Several different types of spores are common on nearly all of the grinding stones from both sites (Table 4). Given the long-term storage of these artifacts in a field station near the site before being moved to the museum, it seems likely that most of these are not related to their mid-Holocene use.
Usewear patterns, residues, artifact typology, and tool functions
The 10 tools examined can be divided into three groups typologically: (1) normal grinding stones, which are commonly found in Neolithic sites of North China and include slabs in rectangular shape with round corners and elongate handstones with a multi-faceted cross section; (2) grinding stones similar to group 1, but small depressions are present on one side of the tool; and (3) grinding stones with special forms that are uncommon in the tool assemblages.
Group 1. Normal grinding stones
Six tools analyzed here are normal grinding stones, including GS1, GS2, and GS4 and R1, R2, and R5. GS1, GS2, and GS4 are slab fragments with similar shape and breakage. Peels taken from the used surfaces show very similar usewear traces: The surface is uneven with low- to medium-level development of polish on crystal grains; some very fine parallel striations are visible occasionally (Figure 7(1 and 2)). The low- to medium-level polish is similar to plant processing in our experimental reference collection, and the fine striations particularly resemble the traces from grinding USOs (Figure 8(1 and 2) and Table 5).

Usewear patterns from Shihushan grinding stones (magnifications 1–10: 200×; 11 and 12: 500×). Group 1 tools: (1 and 2) GS2, medium level polish and very fine striations running diagonally; (3 and 4) side of R1, extensive high-level polish and very fine striations; and (5) end of R5, uneven surface, highly polished areas on high plateau, and very fine striations. Group 2 tools: (6) from a depression on GS3, showing high-level polish with small pitting marks on a crystal grain; (7) R3, highly polished areas with fine striations and small pitting marks. Group 3 tools: (8–10) GS5, showing furrow striations on the flat surface of Side A (8), a small highly polished area with very fine striations in the depression (9), and reticulate highly polished areas with straight and short striations on a lateral side (10); (11 and 12) R4, lateral sides show medium-level polish with fine striations, and the tool end exhibits much more extensive polish and striations, respectively.
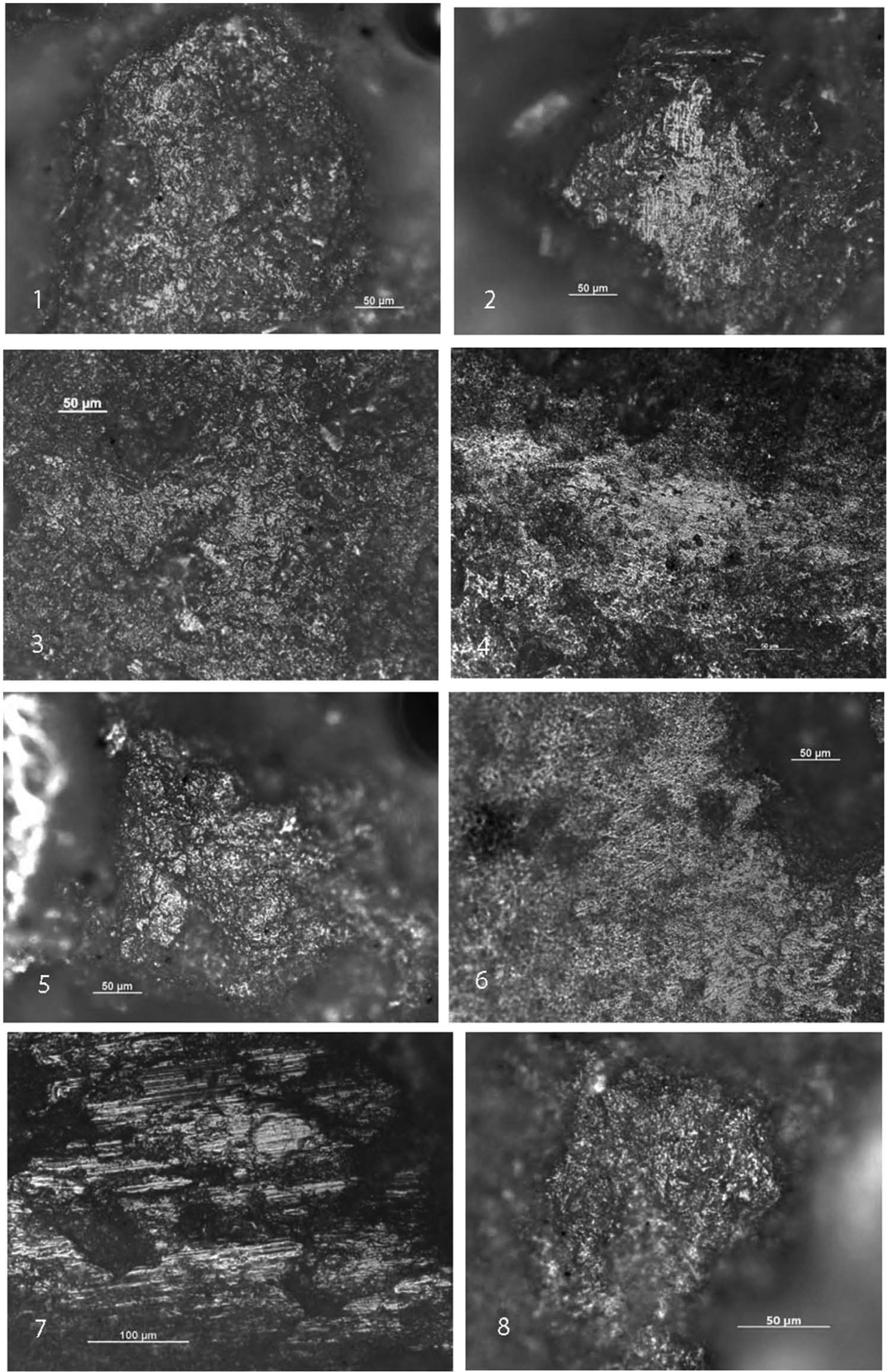
Usewear patterns from experimental study (magnifications 7: 100×; others: 200×). (1 and 2) Medium-level polish and very fine striations on slab after grinding dry snakegourd root for 2 h; (3 and 4) high-level polish and extensive fine striations on handstone after grinding snakegourd root for 2 h; (5) medium-level polish on slab after grinding dry yam for 1 h; (6) extensive fine striations on a handstone after grinding dry yam for 1 h; (7) furrow striations on slab after abrading stone tool for 8 h; and (8) low-level polish on slab after grinding dehusked millet for 2.5 h.
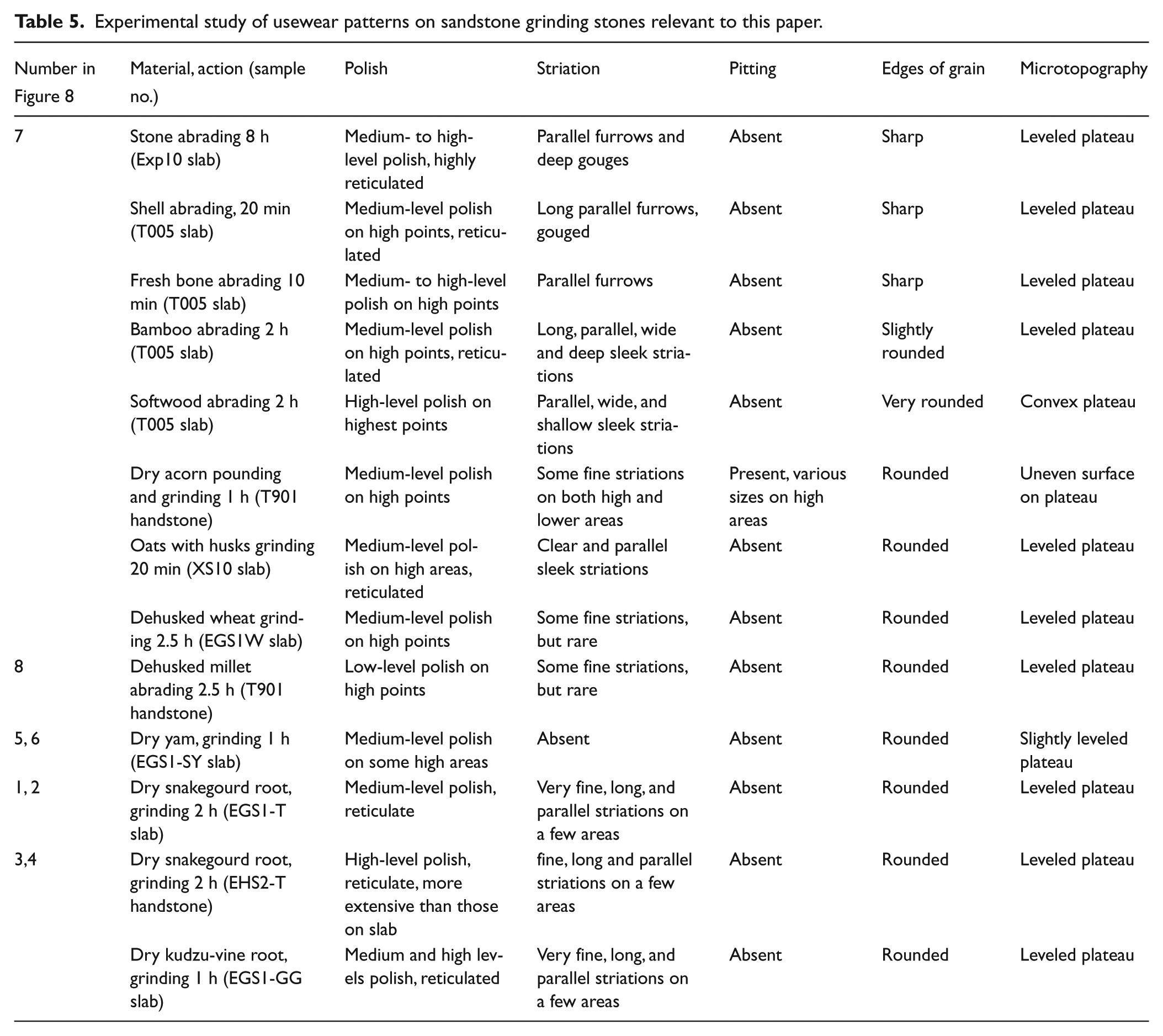
Experimental study of usewear patterns on sandstone grinding stones relevant to this paper.
Starch remains from the three slabs are similar in composition. Among a total of 144 starch grains, 113 are from snakegourd and lily/yam and 16 are USOs. Only three starch grains are identified as non-USO plants, including Quercus acorn, Paniceae millet, and Triticeae grasses (Table 2). These slabs were likely used for processing mainly USOs. Phytoliths are not abundant on these grinding stones, as might be expected if processing starchy roots. Hair cells are the one phytolith type shared by all three grinding stones and are most common on GS2. The burned silica distribution is comparable to that of the hair cells, most abundant on GS2 and GS4. GS2 has a large assemblage of fibers, possibly related to Typha sp. root processing. Although not abundant, raphides are most common on these three grinding stones (and GS3). The combination of starch and phytolith data suggests that these artifacts were likely used to process roots and tubers.
R1, R2, and R5 are elongate handstones, with various shapes in cross section (oval, nearly rectangular, or nearly square). They also show very similar usewear traces with one another, characterized by medium- to high-level of polish and very fine striations sometimes visible on polished areas (Figure 7(3 and 4)). R5 appears to have also been used as a pestle, indicated by the presence at the tool ends of uneven surface, highly polished areas on high plateau, and very fine striations (Figure 7(5)). Compared with the slabs, the polish level is higher, and striations are more extensive on handstones, all consistent with tuber processing in our experimental study (Figure 8(3 and 4) and Table 5).
Consistent with tuber-processing usewear traces, starch remains from these three handstones (N = 62) are almost entirely derived from snakegourd root, lily/yam, and possibly Typha (in the category of USOs), except for one millet starch grain (Table 2). All three handstones had only a few phytoliths (hair cells), limited evidence of burning, no raphides, and fairly low counts of fibers.
Group 2. Grinding stones with small depressions
Two tools, GS3 and R3, were made with small depressions on their surface. GS3 is a slab fragment. Side A is flat, while Side B has several small depressions. Peels from Side A exhibit a low-level polish with very fine striations on some small areas, similar to those on GS1, GS2, and GS4. Peels taken from the depressions on Side B exhibit a very uneven surface, and high-level polish with very fine striations is visible on the high plateau of crystal grains (Figure 7(6)). Small pitting marks are also present.
A total of 25 starch grains were recovered from GS3, including snakegourd root, lily/yam, acorn, millet, and Triticeae grasses. This tool appears to have been used for processing various plant foods. The presence of acorn starch suggests that the high polish and pitting marks on the depression are probably related to cracking shells of acorn and other nuts. The phytolith types present in GS3(B) were similar to GS1 and GS2, with the additional presence of a few grass epidermal cells and some non-diagnostic epidermal long cells. GS3 had the most abundant and diverse assemblage of tissue fragments. One phytolith form from a seed epidermis may also suggest the processing of a plant not found on other tools. GS3(B) is also likely to have been used for more siliceous material, supporting the high-level polish in usewear traces. These differences highlight the likelihood of a different function for this grinding stone.
R3 is an elongate handstone with three small depressions on a lateral side. The general usewear traces include medium- to high-level polish and very fine striations, similar to the handstones in group I, indicating plant processing. The peel from one of the depressions shows highly polished areas with fine and long striations, and pitting marks are also present (Figure 7(7)). This handstone appears to have also been used as a pestle, indicated by the usewear traces similar to those on the end of R5, as described above.
Most starch grains uncovered from R3 are identifiable as USOs (42 of the total 46), including snakegourd root, lily/yam, and possibly Typha (in the category of USO) in addition to a very few grains from acorn (one grain) and millet (three grains), suggesting that its primary function was associated with tuber/root processing. The small depressions on this tool may have been used for cracking acorn shells based on comparison with reference materials (Table 5), a function similar to the depressions on GS3.
R3 had a relatively large phytolith assemblage, including a range of different hair cells. The armed hair and hair cell bases are consistent with Cucurbitaceae species identified in the starch (but, as noted, armed hair cells are also found in other taxa from Ulmaceae and Asteraceae). R3 also had a superabundance of fibers, most likely related to processing tubers/roots, possibly snakegourd and Typha sp. Only a few raphides were identified for this handstone, perhaps associated with yam and/or Typha. Only two small pieces of silicified grass epidermis were present in this assemblage, confirming the limited usage for grain processing.
Group 3. Grinding stones in special shapes
GS5 and R4 can be classified into this group, based on their unusual morphology. GS5 is a nearly square-shaped stone block with a large depression on one side (Side A). Peel 1, taken from the flat surface of Side A, shows many small spots of polish, some with furrow striations (angular in cross section), resembling usewear traces from abrading hard objects, such as stone, in our experimental reference collection. Peel 2, taken from the depression, exhibits highly polished spots, some with very fine striations, similar to plant processing. Peel 3, taken from a lateral side, shows reticulate and highly polished areas with very fine, straight, and short striations, a pattern which does not match to any of our experimental samples. This side of the tool may have been used for polishing unknown materials with flat surfaces, a function rather unique in the tool assemblage examined here. GS5 was used for processing and abrading materials with different hardness (Figure 7(8–10)).
Three starch grains were recovered from this tool, all identifiable as snakegourd root. This tool evidently had multiple functions, one of which was tuber processing. GS5 revealed very little in terms of any microfossil remains (a few fibers), also perhaps indicating that non-plant-related processes were the most common function of this tool.
R4 is a relatively short handstone with a square-shaped cross section. PVS peels taken from surfaces on the lateral sides show medium-level polish and very fine striations. The peel from the tool end exhibits many more reticulate areas of polish and extensive fine striations, clearly visible under 500× magnification, suggesting actions of pounding and grinding plants (Figure 7(11 and 12)). All these traces are consistent with processing tubers/roots with high contents of fiber (Figure 8(1–4) and Table 5).
The starch assemblage (N = 205) recovered from this tool is unique, composed of mainly Typha rhizome (N = 107) and Triticeae grasses (N = 56), followed by snakegourd root and lily/yam (N = 19) and acorn (N = 1). Most starch grains from R4 (N = 150) show clear traces of damage, suggesting that these plants were extensively processed.
R4 had a somewhat larger and more diverse phytolith assemblage. In addition to hair cells and hair bases, R4 had tracheids consistent with those found in Typha sp. Based on the range of tissue fragments, the phytolith assemblage on R4 suggests some diversity in the range of plants processed with this handstone. If grasses were processed with this tool, it would appear that they were already hulled, as there are no grass phytoliths present in the assemblage. This tool seems to have been used for abrading and pounding plants, mainly Typha rhizome and Triticeae grasses.
Summary
Tool types, usewear patterns, and microbotanical residues are supplementary to one another for understanding general tool function. Grinding tools from Groups 1 and 2 seem to have performed very similar tasks: processing mainly tubers/roots (lily/yam, snakegourd, and possibly Typha), and, to a much lesser extent, Triticeae grasses, millet, and acorn. It is notable that very few acorn starch grains are present in the assemblage. Acorn was apparently shelled, as indicated by Group 2 tools, but not ground in large quantities with the tools examined. The grinding tools with special forms appear to have been either used as a multipurpose tools working on plant and non-plant materials (GS5), or used for processing mainly Typha rhizome and Triticeae grasses (R4). The limited phytolith assemblage is consistent with plant food processing: most edible portions of plants, including roots, are low in silica.
Despite the limited diagnostic material in the phytolith assemblage, form diversity can also provide some evidence for tool use, as long as the forms are from different taxa. In terms of the number of phytolith types present, GS2 and R4 have the most diverse assemblage, with R3 and GS3B also showing some evidence of a wider range of types. This is consistent with the starch analysis, particularly for R4.
Comparisons of tool use, based on the phytoliths, between the two sites reveal very little difference beyond the proportions of individual forms. There are a few more types represented at SHSII, but the assemblages are too small to draw strong conclusions from this. Preservation appears to be a bit better at the earlier SHSII site. The similarity in the assemblages suggests that subsistence practices change little over time (at least in relation to these grinding stones).
In general, the results based on the analyses of starch, fibers, and raphides are complimentary to one another, providing greater insight to the tool function and reinforcing the interpretation that most of these grinding stones were used to process USOs.
Discussion and conclusion
Grinding stones from the Daihai region were evidently used for processing a wide range of starchy plants, including lily/yam, snakegourd root, Typha rhizomes, wild Triticeae grasses, acorn, and millet. Contrary to previous assumptions, it was USOs, not millet, that were likely the main staple foods processed on these grinding stones. Most of these plants were exploited by people long before the Neolithic period in North China.
Starch grains resembling Dioscorea yam, Lilium bulb, Typha roots, and Triticeae grasses have been found on 28,000-year-old flaked tools at Shuidonggou in Ningxia (Guan, 2011). The earliest direct evidence for human processing of Dioscorea yam and Trichosanthes snakegourd root in China comes from starch residues found on grinding stones, dating to c. 23,000 at Shizitan Locality 14 in Shanxi (Liu et al., 2013a).
These plants still grow in North China today (Lu and Chen, 1986; Pei and Ding, 1985). Lilium bulb and Dioscorea yam have been commonly used as food and in traditional Chinese medicine (Li, 1981). They can be eaten as fresh vegetables, or dried and ground into flour to make various starchy foods. As recorded in ethnographic accounts, wild lilies and yams were among the various tubers and roots collected as staple food by many ethnic groups in Yunnan. Lilies were ground and filtered to obtain starch (Li and Lu, 1987: 180–181). Both lily and yam have been domesticated, but the processes of their domestication are unclear. Snakegourd root, also called tian hua fen, has been used as a traditional medicine as well as for famine food as recorded in ancient texts. Unlike lily and yam, this tuber needs to be ground into flour and leached to obtain starch (Zhu, 1406).
There is no evidence of Typha being domesticated in China, but its stems and rhizomes were used as famine food in North China during the historical times. The fresh stems were cooked directly, while the rhizomes were dried and then ground into flour for making pancakes or steamed buns (Zhu, 1406). Typha produces much fiber, which has been used by people around the globe for food as well as for making strings, baskets, and so on. Among Australian Aborigines, Typha starch is extracted from fiber by beating (Gott, 1999). This scenario helps to explain why the Typha starch grains show much damage and fibers are abundant in the residue samples.
Starch grains from USOs account for the great majority of the Shihushan assemblage (81%) and also appear on every tool examined. This phenomenon is consistent with the common presence of fibers and raphides, which are likely derived from snakegourd roots, yam, and Typha. Given that grinding stones were the most common tools at the sites, it is reasonable to infer that these tubers and roots were the staple foods of the earliest Neolithic people in the Daihai region.
Triticeae starch grains have also been found on stone tools (flaked tools and grinding stones) from several Upper Paleolithic and Neolithic sites in North China (Guan, 2011; Liu et al., in press, 2011, 2013a; Yang and Perry, 2013). Many species in the Triticeae tribe are indigenous in North China, but none has been domesticated. These grasses were apparently used widely as food before domesticated wheat and barley, also belonging to the Triticeae tribe, were introduced to China from West Asia during the 3rd millennium
Acorn starch grains have been found on grinding stones dating to the Upper Paleolithic (c. 11,600 cal. BP; Liu et al., 2011), Epipaleolithic (Liu et al., 2010b), early Neolithic (Liu et al., in press, 2013b; Tao et al., 2011), and middle Neolithic (Yang et al., 2009), all in northern China. This nut seems to have been commonly used as food in prehistory. Oak trees appear to have been relatively abundant in the Daihai region, based on the Quercus counts in the pollen sum (13%) during the 7th millennium
Millet starch grains were only found on four SHSII tools, accounting for 2% of the total starch count. Millet can be cooked as whole grains without being ground; therefore, it is difficult to estimate how much it may have been consumed based on starch remains on grinding stones. Nevertheless, given that millet starch grains are rare on grinding stones, which dominate the tool kit, and typical harvesting knives were rarely found at the sites, it is likely that the level of millet production was rather low.
The Shihushan people apparently relied on a wide range of plants, which were available in abundance around the Daihai region. The easy access to highly ranked plants, particularly USOs, during a period of climatic amelioration, and low population density, might be among the reasons that the sedentary Neolithic people did not practice a cereal-based intensive agricultural economy in this region. The extent to which these USOs may have been domesticated is currently unknown, but some of them may have been managed, if not cultivated, as staple foods. Lilies, yams, and snakegourd were exploited by people in North China more than 20,000 years ago and were recorded as cultivated/domesticated plants in later historical times, as discussed above. Given that the SHS populations already possessed the knowledge of farming, it is plausible that these tubers and roots were intentionally managed or cultivated to maintain high productivity and therefore provide staple foods. Strategies of plant management may be comparable to those documented in the ethnographic and archeological record from Australia (Denham, 2008; Gott, 1982), among native Americans in California (Anderson, 2005), and the Jomon in Japan (Crawford, 2011).
In conclusion, recent microbotanical studies on plant residues recovered from tools have revealed a considerable amount of new data, indicating that a variety of plants, including tubers/roots and cereals, were intensively exploited throughout the Upper Paleolithic and early Neolithic times, as discussed previously. The current project on Shihushan grinding stones extends this picture to the Yangshao culture of the middle Neolithic period, suggesting that a broad-spectrum subsistence strategy continued in some regions when millet agriculture was thought to have been well established in northern China. Since tubers/roots normally do not survive well as macrobotanical remains and are not easily identifiable, microfossil analyses are usually the most effective way to recover and identify these plants, contributing new insights into human diets in the past.
Footnotes
Acknowledgements
We thank Mr Wang Xinyu and other archaeologists in the Jining Museum who generously assisted our sample collection. We also appreciate the constructive comments made by two anonymous reviewers. The research was supported by the Stanford Archaeology Center. The underlying research materials related to this paper (data and samples) can be accessed by contacting the first author.
Funding
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.