
Abstract
Micropaleontological analyses of a sediment core raised in Disko Bugt (West of Greenland) were undertaken in order to document paleoceanographical changes in the eastern Baffin Bay during the Holocene. The modern analogue technique (MAT) applied to dinocyst assemblages provided information on paleo-sea-surface conditions, whereas isotopic analyses of benthic foraminifers aimed at documenting the ‘deep’ water mass occupying the site. During the earlier interval recorded (~10 to ~7.3 cal. kyr BP), important discharge of ice and meltwater from the Greenland Ice Sheet (GIS) margin, notably through the Jakobshavn Isbrae, resulted in harsh conditions with a dense sea-ice cover and low temperatures, productivity, and foraminiferal abundances. Postglacial conditions settled at ~7.3 cal. kyr BP, with a sharp rise in dinocyst abundance and species diversity, which led to reconstruct increase in summer temperatures. We link this transition to the advection of West Greenland Current waters in the upper part of the water column after the reduction of meltwater inputs from GIS. Optimal temperature conditions reaching up to >10°C were finally achieved in surface waters at ~6 cal. kyr BP. Slight cooling pulses were then recorded at ~4.2–4 and ~1.5–1 cal. kyr BP, and the final optimum recorded in surface temperature from ~1 to 0.8 cal. kyr BP is associated with the ‘Medieval Warm Period’. Throughout the postglacial interval, the data suggest an opposition between sea-surface temperatures and salinity, with warmer intervals being characterized by lower salinity waters, probably as a result of the higher freshwater discharge along the ice margin and notably the Jakobshavn Isbrae.
Introduction
Disko Bugt is an area of interest with regard to ocean-ice dynamics because it is located on the western margin of Greenland, where flows the Jakobshavn Isbrae, the largest and fastest ice stream of Northern Hemisphere (Bindschadler, 1984). Disko Bugt is bathed by the West Greenland Current (WGC), constituted of mixed waters from the East Greenland Current (EGC) and Atlantic waters from the Irminger Current (IC). A warming trend of the WGC, accompanied by significant acceleration of glacier retreats and meltwater discharges, has recently been recorded (Holland et al., 2008; Rignot and Kanagaratnam, 2006). The Jakobshavn Isbrae currently drains 7% of the Greenland Ice Sheet (GIS; Roberts and Long, 2005) and produces a large amount of icebergs (Weidick and Bennike, 2007). In 2003, the velocity of the Jakobshavn Isbrae reached 12.6 km/yr (Joughin et al., 2004). Ongoing ice discharges from the GIS contribute significantly to the sea level rise, with values of 0.34 mm/yr from 1996 to 2005 (Rignot and Kanagaratnam, 2006).
In order to explore the relationship between the Jakobshavn Isbrae dynamics and the ocean conditions during the Holocene, we have analyzed a marine sediment core from Disko Bugt (core MSM343300; Figure 1). A first objective was to document long-term changes of the WGC, in particular that of the subsurface layer, which is mostly influenced by the northward flow of North Atlantic waters. A second objective was to reconstruct sea-surface temperature (SST) and sea-surface salinity (SSS) to assess the impact of meltwater discharge from Jakobshavn Isbrae on marine conditions at mouth of Disko Bugt. To meet these objectives, we used organic-walled and calcareous microfossils as tracers of surface-water masses properties. Palynological analyses were performed with special attention to dinoflagellate cysts, which permit reconstruction of summer SST, SSS, sea-ice cover, and primary productivity (e.g. de Vernal and Rochon, 2011). In addition, isotopic analyses (δ18O and δ13C) of benthic foraminifer shells were made on two different species, Islandiella norcrossi and Nonionellina labradorica, as a means to document the properties of bottom waters, which correspond to the deeper part of the WGC, mostly its ‘warm’ component from the North Atlantic.
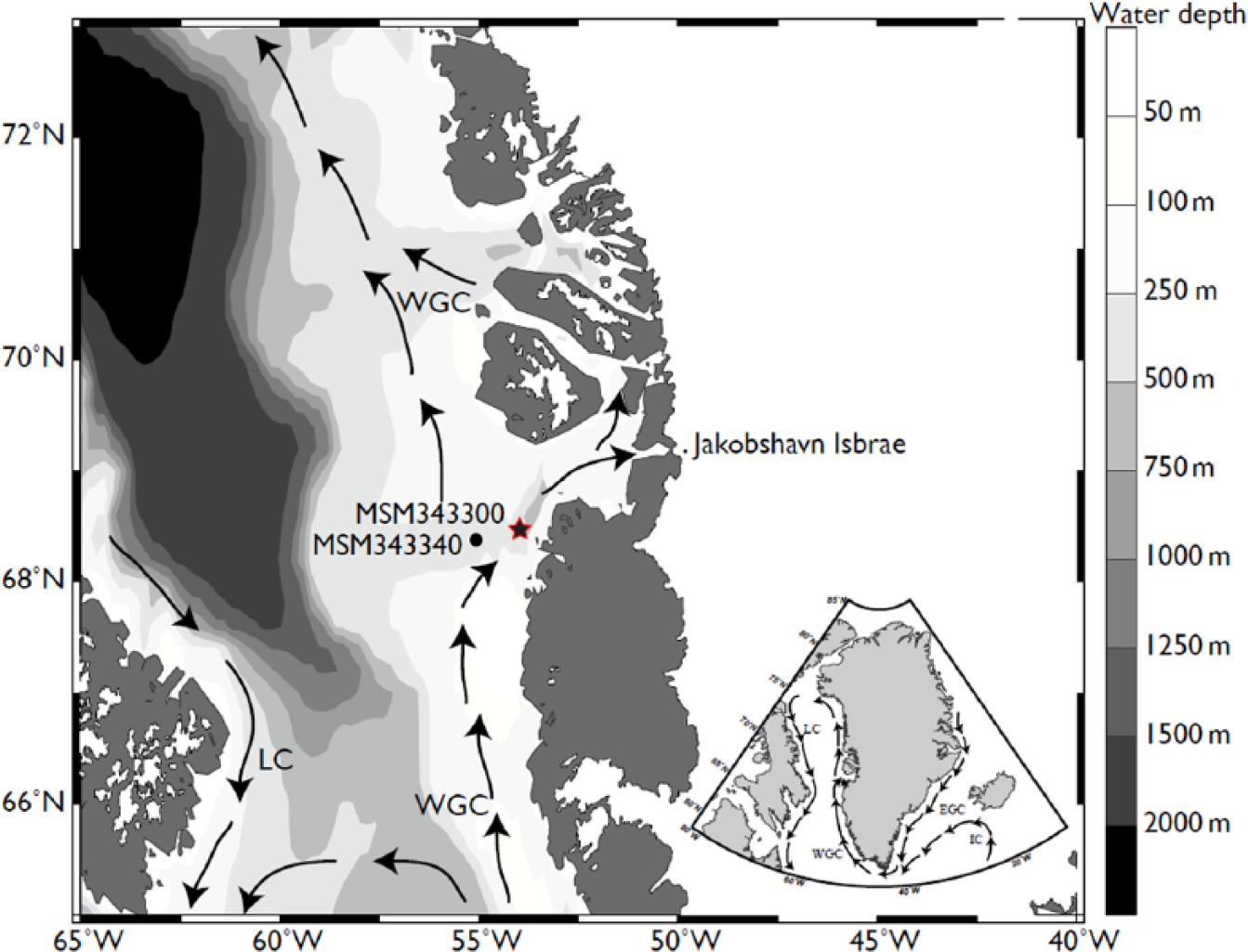
Map of the study area. A star indicates the location of core MSM343300.
Study area
Disko Bugt is a large marine embayment with water depths ranging mostly from 200 to 400 m and reaching up to 900 m in the Egedesminde Dyb trough (Hogan et al., 2012; Long and Roberts, 2003; Ó Cofaigh et al., 2013; Perner et al., 2011; Roberts and Long, 2005). The trough is associated with the Jakobshavn Isbrae ice stream route and GIS melting path during glacial times (Long and Roberts, 2003). In the Disko Bugt area, the WGC constitutes a combination of the IC formed of warm and saline Atlantic waters from the North Atlantic Current (NAC) and the EGC, which consists of polar and low salinity waters (Buch, 1982; Tang et al., 2004). The area is also characterized by buoyant low saline surface waters resulting from GIS meltwater supplies (Andersen, 1981). The WGC today occupies subsurface waters below 200 m, where temperature and salinity are 2.5–3.5°C and 34.5–34.8 psu, respectively (Andersen, 1981). The surface waters are usually colder as they are influenced by the polar water from the EGC in addition to the regional runoff and meltwater discharge. Data compiled from the National Oceanographic Data Center (NODC, 2001) indicate summer SST of 4.4 ± 1.24°C and salinity ranging between 32.9 and 33.4 at the study site. The modern sea-ice cover averages 3.8 ± 1.3 months/yr for the period 1953–2003 (National Snow and Ice Data Center (NSIDC), 2003).
Material and methods
Sediment core and chronology
The study core MSM343300 (68°28,311′N, 54°00,118′W, Figure 1) was collected by gravity coring during the June 2007 Maria S. Merian expedition. It was retrieved from a 519 m water depth (Harff et al., 2007), southwest of Disko Bugt in the direct flow path of the WGC.
The chronology was established from 25 accelerator mass spectrometry (AMS) 14C dates in mollusc shells and benthic foraminifers (Table 1, Figure 2). The AMS radiocarbon dates were calibrated using the Marine09 (Reimer et al., 2009) calibration curve in CALIB 6.0.2 (Stuiver and Reimer, 1993). A total air-sea reservoir correction of 550 years was applied. It includes the usual correction of 410 years and a ΔR of 140 ± 35 years established from two measurements on modern Astarte collected at 60–70 m of water depth (McNeely et al., 2006). Such reservoir correction is consistent with the one applied by, for example, Lloyd et al. (2011) and Perner et al. (2011, 2013).
Radiocarbon dates for core MSM343300. Calibrated years include one standard deviation.
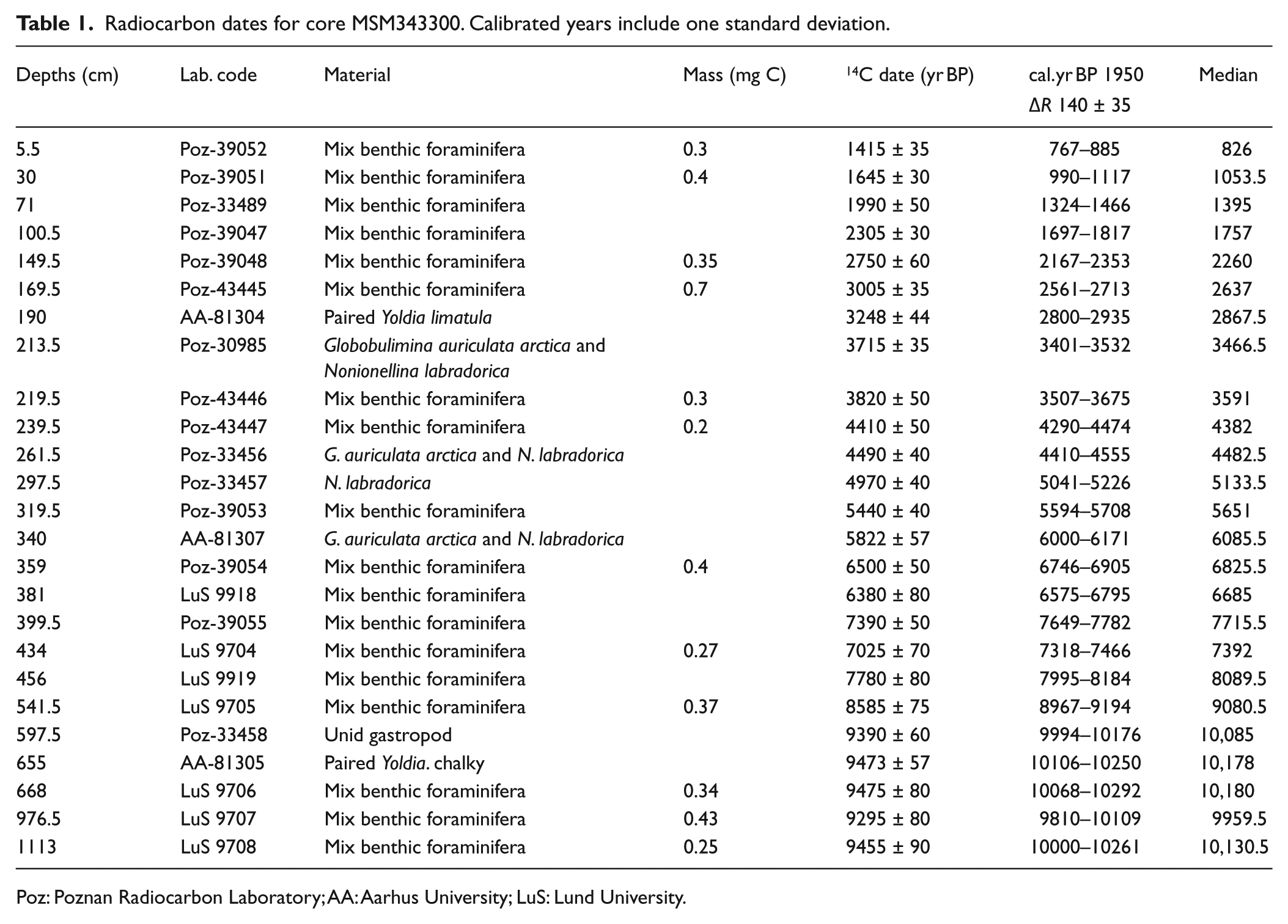
Poz: Poznan Radiocarbon Laboratory; AA: Aarhus University; LuS: Lund University.
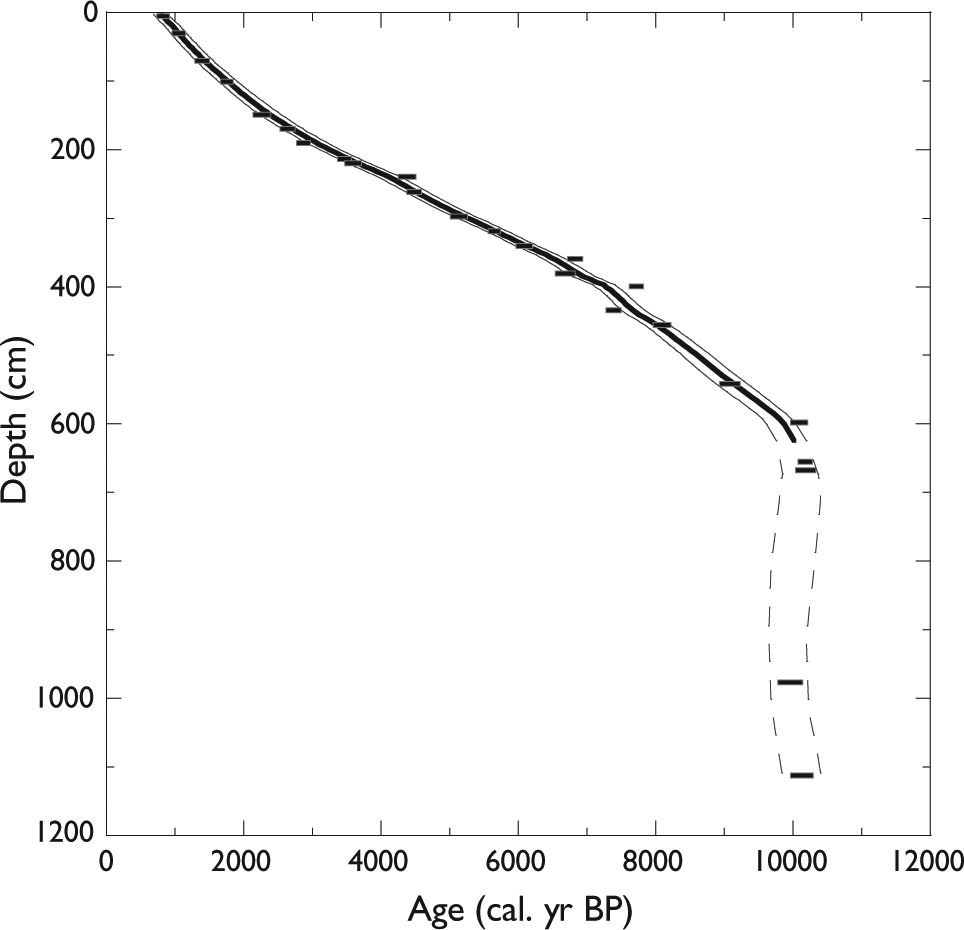
Age versus depth relationship in core MSM343300. 1-sigma range calibrated age 14C is represented in black, smooth interpolation on the median values is represented by a black line, and 2-sigma range smooth interpolations are represented by a gray line.
Age–depth profile
The age versus depth profile was established based on interpolation (Figure 2). For the interval younger than 7500 cal. yr BP, the age to depth relationship is the same that reported by Perner et al. (2013). Based on the age model, sedimentation rates at the coring site ranged from 0.5 to 0.9 mm/yr during the last 10,000 years: they were approximately 0.82 mm/yr between ~10,000 and ~7300 cal. yr BP, 0.5 mm/yr between 7300 and 2500 cal. yr BP, and 0.9 mm/yr between 2500 and 800 cal. yr BP. In the lower part of the core, from 597 to 1113 cm, five AMS 14C dates yielded similar ages of about 10,000–10,200 cal. yr BP, which indicate extremely rapid sediment accumulation.
Microfossil analyses
Sub-sampling was done at 4-cm intervals throughout the core, which provide a time resolution of 50–80 years. The sediment samples were processed according to the protocol of de Vernal et al. (1996). A volume of 1–3.5 cm3 of dry sediment was sieved on 106 µm and 10 µm mesh sieves. The fraction between 10 and 106 µm was used for palynological preparation and the fraction >106 µm was kept for the hand-picking of foraminifers.
Palynological analyses
In order to concentrate the organic remains, carbonate and silicate particles were dissolved from repeated chemical treatment with HCl (10%) and warm HF (48%). A small drop of the final residue was mounted on a microscope slides with glycerine jelly. Before sieving and chemical treatments, Lycopodium clavatum spore tablets were added to estimate palynomorph concentration (Matthews, 1969; Mertens et al., 2009). Counting and identification were performed with an Orthoplan Leitz optical microscope. All palynomorphs were counted (dinoflagellate cysts, pollen grains, spores, Halodinium and organic linings of foraminifer). A special attention was paid to dinoflagellate cysts (hereafter dinocysts). At least 300 dinocysts were counted per sample, which provide statistically reliable data (Mertens et al., 2009). The taxonomic nomenclature was based on Rochon et al. (1999) and Radi et al. (2013).
Quantitative reconstructions of sea-surface conditions from dinocyst assemblages
Reconstruction of sea-surface parameters were made based on the modern analogue technique (MAT), which relies on similarities between fossil and modern assemblages instead of calibrations between sea-surface parameters and assemblages (Guiot and de Vernal, 2007). Hence, MAT can be applied for simultaneous reconstruction of different parameters and appears appropriate in the case of non-linear relationship between assemblages and climate parameters. MAT was applied using dinocyst assemblages to reconstruct SST and SSS in summer, in addition to sea-ice cover extent and mean annual productivity. All these parameters play a determinant on the distribution of dinocyst assemblages as shown from multivariate analyses (e.g. de Vernal et al., 2001, 2005, 2013; Radi and de Vernal, 2008; Rochon et al., 1999). Here, we have used the updated Geotop dinocyst database that includes 66 taxa after taxonomic standardization and 1492 reference sites from mid-high latitude of the Northern Hemisphere (de Vernal et al., 2013). We have followed the procedure as described by de Vernal et al. (2005). The relative occurrence of taxa was logarithmic transformed in order to emphasize the weight of accompanying taxa as they have usually a more narrow range of ecological affinities than dominant taxa, which are often opportunistic and cosmopolitan (de Vernal et al., 2005). After the log transformation of the taxa occurrence, the five closest modern analogues were identified to estimate past sea-surface conditions. Reconstructions are given as the most probable value, which is the mean of the sea-surface parameter for the five best analogues, weighted inversely to the distance between the fossil and modern spectra. Minimum and maximum possible according to the set of analogues identified are also reported. In order to assess the error of prediction of the reconstructions, validation tests are made after splitting of the database, with 1/6 of the data points taken randomly being used to verify the ability of the approach to reconstruct the sea-surface parameters. The errors of prediction at ±1σ level established at ±1.6°C for summer SST, ±2.6 for SSS, ± 1.4 months/yr for the sea-ice cover, and ± 55 gC/m2 for productivity. It is noted that the SSS error of prediction is large because the database includes low salinity environments (down to 5 psu), where the variability of surface conditions is particularly large. When considering only the >30 and >33 salinity domains, the summer SSS errors of prediction are ±1.3 and ±0.8 psu, respectively.
In this study, a special attention was paid to the genus Islandinium, as the specie cezare appears to have an important weight in salinity reconstruction where dinocyst diversity is low (cf. Ouellet-Bernier, 2014).
Foraminifer processing
The >106 µm fraction was used for foraminifer analyses. We did not analyze benthic foraminifer assemblages, as they were previously described by Perner et al. (2013). The two dominant calcareous taxa, I. norcrossi and N. labradorica were selected for isotopic measurements. These two species have different ecological affinities. I. norcrossi is a shallow infaunal species common in Arctic environment characterized by high and stable salinities, in addition to relatively warm subpolar waters conditions (Hald and Steinsund, 1996; Knudsen et al., 2012; Polyak et al., 2002). The foraminifer N. labradorica is a deeper infaunal species, which is associated with episodic fresh phytodetritus production (Knudsen et al., 2012; Murray, 2006; Polyak et al., 2002; Rytter et al., 2002; Steinsund, 1994).
In the study core, the foraminifer shells of I. norcrossi occur sporadically prior to ~6100 cal. yr BP and continuously afterward (above 341 cm). Shells of N. labradorica record sporadic occurrences at ~8200, ~6000, ~5500–5100, ~4000–2800, and ~1900–1200 cal. yr BP. Although N. labradorica was the second most abundant specie after I. norcrossi, it occurred in low abundance. Hence, two size fractions were used, 150–250 µm and >250 µm, to collect enough specimens for isotope analyses.
Oxygen and carbon isotope analyses (δ18O, δ13C) in foraminifer shells
Approximately 15 tests of I. norcrossi (150–250 µm) and 10 tests of N. labradorica (>250 µm) were roasted under vacuum at 250°C for 1 h. Then, they were analyzed with a Micromass Isoprime™ isotope ratio mass spectrometer in dual inlet mode coupled with a MultiCard™ preparation system. Samples were heated to a constant temperature (90°C) for at least 30 min. CO2 from foraminifer shells was extracted after dissolution with H3PO4 (102%). An internal reference carbonate material (UQ6) calibrated on the V-PDB scale was used. The overall analytical uncertainty (±1σ) is better than ±0.05‰ for both δ13C and δ18O, as determined from replicate measurements of the home standard material.
Results
Palynological assemblages and pelagic fluxes
The palynological assemblages were largely dominated by dinocysts. Their concentration ranged between 2040 and 171,500 cysts/g, suggesting very high pelagic productivity at least during the middle and late Holocene (Figure 3). Average fluxes of 3400 cysts/cm2/yr were calculated in the interval spanning ~10,000 to ~7300 cal. yr BP (625–405 cm), while they ranged from ~2000 to ~20,000 cysts/cm2/yr in the upper part of the sequence.
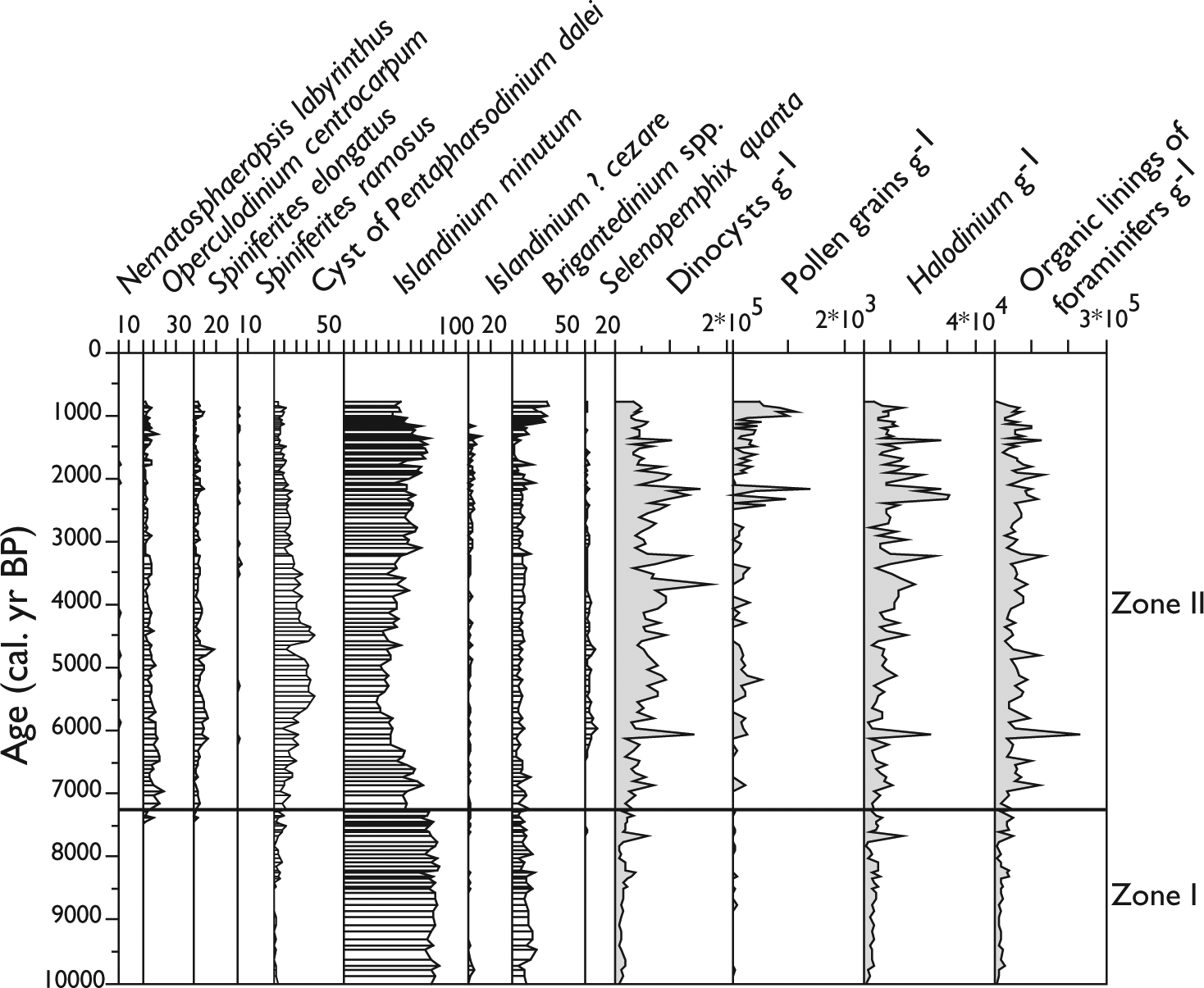
Percentages of main dinocyst taxa in core MSM343300 and concentrations of dinocyst, pollen grains Halodinium and organic linings of foraminifers.
Pollen grains were present in low numbers (<1400 grains/g). Picea, Pinus, and Betula were the most common taxa. Low numbers of monolete and trilete spores, including Lycopodium spp., were observed. Concentrations of organic linings of foraminifers (cf. de Vernal et al., 1992) were relatively high, ranging between 550 and 234,000 linings/g, with maximum abundance during the middle and late Holocene. Concentrations of organic linings were significantly higher than those of calcareous shells of benthic foraminifers. However, linings were observed in the 10–106 µm fraction, whereas shells were counted in the >106 µm fraction. Thus, the higher lining abundances might either indicate some calcium carbonate dissolution in the sediment (cf. de Vernal et al., 1992) or a much higher abundance of small lining-bearing benthic foraminifers in the <106 µm fraction and probably both. High concentrations of the acritarch Halodinium were recorded with up to 31,200 specimens/g. Halodinium has still unknown biological affinities, but it was generally associated with shallow marine and estuarine environment (e.g. Verhoeven et al., 2014), notably in cold Arctic–subarctic settings marked by freshwater–meltwater inputs (de Vernal et al., 1989; Richerol et al., 2008).
Dinocyst assemblages
Dinocyst assemblages showed moderately high species diversity, with a maximum of 15 species, 9 being common to abundant (Table 2, Figure 3). The assemblages were dominated by Islandinium minutum, which was accompanied by the cyst of Pentapharsodinium dalei, Brigantedinium spp., Operculodinium centrocarpum, Spiniferites elongatus, Selenopemphix quanta, and Islandinium? cezare. Nematosphaeropsis labyrinthus and Spiniferites ramosus occurred in very low number. The dinocyst assemblages permit to identify two main zones (Figure 3).
List of dinoflagellate cysts.

A: autotrophic; H: heterotrophic.
Zone I covers from ~10,000 to ~7300 cal. yr BP. The assemblage was composed almost exclusively of heterotrophic taxa, notably I. minutum, which largely dominates, and Brigantedinium. I. minutum occurs mostly in neritic environments, whereas it occupies a wide range of temperature and salinity conditions, and dominates in environments characterized by cold conditions and dense seasonal sea-ice cover (Buck et al., 1998; de Vernal et al., 2001, 2008, 2013; Rochon et al., 1999). I. minutum has recently been shown to live under sea ice (Potvin et al., 2013). Brigantedinium spp. is a cosmopolitan taxon, which can be abundant in polar environments (Kunz-Pirrung, 2001; Matthiessen et al., 2005; Rochon et al., 1999).
Zone II covers from ~7300 to ~800 cal. yr BP and was defined from the increase of the P. dalei (Dale, 1996; Rochon et al., 1999), which reaches up to 40%, and the occurrence of other taxa such as S. elongatus, S. ramosus, N. labyrinthus, and S. quanta. The taxa S. elongatus and S. ramosus are generally found in association with Atlantic waters (Marret and Scourse, 2003; Rochon et al., 1999; Zonneveld et al., 2001). S. elongatus prefers cool to temperate conditions (Rochon et al., 1999), whereas S. ramosus is more cosmopolitan and occurs in tropical to subpolar environments (e.g. Rochon et al., 1999; Zonneveld et al., 2001). N. labyrinthus develops in temperate to subpolar latitudes and prefers open oceanic environment (de Vernal et al., 2001; Kunz-Pirrung, 2001). S. quanta is cosmopolitan and occurs in temperate to subpolar environments (Rochon et al., 1999). It is abundant in upwelling zones and in areas of high nutrient concentration (Dale and Fjellså, 1994; Sangiorgi et al., 2002).
Reconstruction of sea-surface conditions
Close modern analogues of dinocyst assemblages were found for all samples, mostly in the Arctic Ocean and/or subpolar North Atlantic. High concentration of dinocysts allowing statistically reliable counts (>300 counted specimens/sample) and high degree of similarity between the fossil assemblages and their modern analogues permit to be as confident as possible with the reconstruction from MAT
In Zone I, reconstructions indicated cold conditions, with summer SST averaging 1.5°C, almost 9 months/yr of sea-ice cover, and a very low primary productivity (100 gC/m2/yr). SSS was relatively high (~31.5 psu) during the interval (Figure 4).
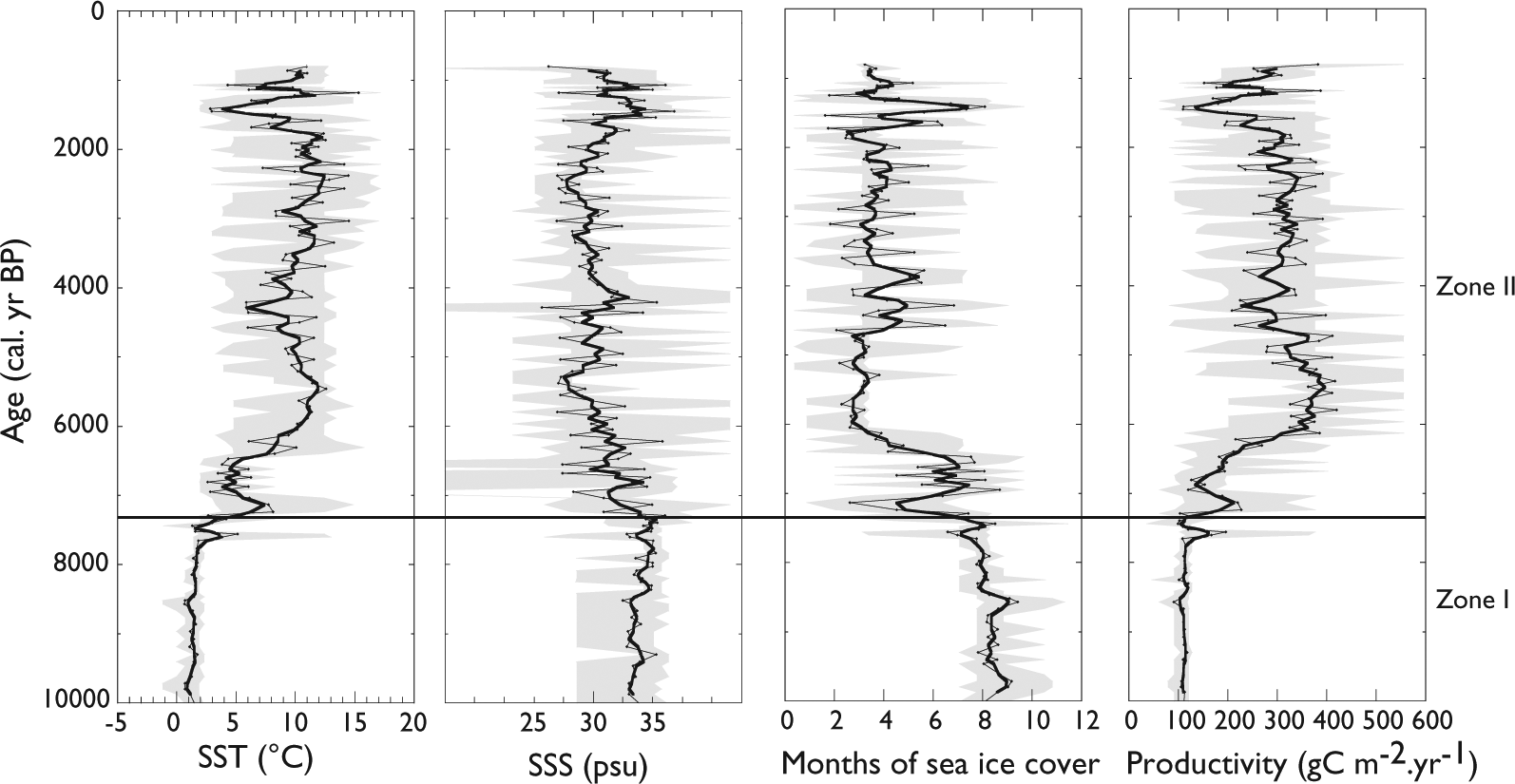
Reconstruction of summer sea-surface temperature (SST), salinity (SSS), seasonal sea-ice cover in months/year, and productivity (in gC/m2/yr) for the last ~10,000 years based on modern analogue technique (MAT) applied to dinocyst assemblages in core MSM343300. Data points are represented by dots, 3-points running mean in dark gray and the maximum and minimum possible according to the set of five analogues are shown in light gray. Zone I and II refer to the dinocyst assemblage zonation as Figure 3.
In Zone II, which is characterized by an increase in species diversity, the MAT-based reconstruction showed a gradual warming in surface waters. From ~7300 to ~7000 cal. yr BP, SST increased rapidly up to ~7°C, while salinities decreased to ~30.5 psu and sea-ice cover was reduced to 5–6 months/yr (Figure 4).Optimal conditions finally established at ~6000 cal. yr BP, with average summer SSTs of about 9–10°C. During this interval, SSS were low, around 28–30 psu, sea-ice cover was restricted to the winter season, and productivity slowly increased up to ~ 400 gC/m2/yr (Figure 4). Cooling pulses were recorded at ~4000–4200 and ~1500–1000 cal. yr BP. They correspond to increase in salinity (up to ~32 psu), which we interpret here as the result of reduced meltwater discharge.
Isotopic composition of benthic foraminifer shells
The isotopic analyzes allowed the establishment of a discontinuous record with large gaps before ~6300 cal. yr BP because of the rarity of specimens (Figure 5). After ~6300 cal. yr BP, the record is more continuous.

Isotopic compositions (δ13C and δ18O) of Islandiella norcrossi (solid diamonds) and Nonionellina labradorica (circles) from core MSM343300. Larger and smaller symbols correspond to the larger (>250 µm) and smaller (150–250 µm) size range of analyzed specimens. Zone I and II refer to the dinocyst assemblage zonation as Figure 3.
The taxon I. norcrossi usually records δ18O and δ13C values close to equilibrium with bottom sea-water, which makes it a useful paleoceanographic tracer (Ishimura et al., 2012). Zone I was characterized by high δ18O values (~4.0‰) from ~10,000 to ~9250 cal. yr BP, which reflect cold water conditions with a near standard mean ocean water salinity. It however depicted variable δ13C values, ranging between −1.91‰ and −0.53‰, with particularly low values (−4.72‰) around 8200 cal. yr BP. This light 13C excursion appeared significant since it was recorded in both taxa analyzed, I. norcrossi and N. labradorica. Zone II was marked by δ18O values ranging 3.56–3.1‰ from ~6100 to ~960 cal. yr BP and by δ13C ranging between −0.69‰ and −0.08‰.
The sporadic occurrence of N. labradorica could be related to episodes of high sea-surface productivity (e.g. Hald and Steinsund, 1992; Jennings et al., 2004; Mackensen et al., 2001; Mudie et al., 1984; Polyak and Solheim, 1994; Polyak et al., 2002; Rytter et al., 2002; Wollenburg and Mackensen, 1998). N. labradorica recorded δ18O values ranging from 3‰ to 3.57‰, which was very close to those observed in I. norcrossi. With respect to δ13C, N. labradorica usually yielded higher values than I. norcrossi, which may be related to the direct influence of high seasonal productivity of fresh phytodetritus (Knudsen et al., 2012; Murray, 2006; Polyak et al., 2002; Rytter et al., 2002). As mentioned above, in Zone I, the δ13C of N. labradorica was characterized by particularly low values at ~8200 cal. yr BP (−6.41‰ to −6.03‰), consistent with the isotopic excursion recorded by I. norcrossi. In Zone II, N. labradorica recorded larger variations than I. norcrossi, with values ranging from −2.93‰ to −0.74‰.
Discussion
The deglaciation phase from ~10,000 to ~7300 cal. yr BP
Important changes occurred in surface and bottom waters in the study area during the deglaciation and the Holocene. In the Disko Bugt area, the glacial retreat began after 13,800 yr BP (Ó Cofaigh et al., 2013). From the adjacent core MSM343340 (Figure 1), it was suggested that a fast ice retreat started at about 12,050 cal. yr BP, with a retreat rate of ~22–275 m/a (Ó Cofaigh et al., 2013). By 10,900 cal. yr BP, the ice margin had retreated at the mouth of Disko Bugt, where glacier ablation resulted in high terrigenous input from 10,140 to 8490 cal. yr BP (Jennings et al., 2014). The extremely high sedimentation rates recorded in our core at about 10,000 cal. yr BP also suggest a rapid sediment accumulation, which might be related with glaciomarine processes (Figure 2). According to Weidick and Bennike (2007), the deglaciation of Disko Bugt occurred rapidly during the early Holocene (~10,500 to ~10,000 yr BP). Long and Roberts (2003) also suggested that the Jakobshavns Isbrae recorded a rapid retreat after ~10,300 cal. yr BP with a velocity of about 4.8 km/yr, which is almost equivalent to the modern value of 6–7 km/yr. Atmospheric temperature significantly increased from ~10,000 to ~8000 yr BP as indicated from the NGRIP ice core data (Vinther et al., 2006). Whereas the GIS margin was retreating, surface waters were characterized by a dense sea-ice cover (>9 months/yr), low summer temperatures (0–3°C), and a low productivity (~100 gC/m2/yr), which is consistent with glaciomarine conditions. This is also compatible with the dominance of I. minutum that can live under pack ice (Potvin et al., 2013; Rochon et al., 1999). Moreover, high δ18O values (~4‰) in benthic foraminifers indicate low temperatures and a relatively high salinity in bottom waters.
Glaciomarine conditions were documented to have prevailed until 7800 cal. yr BP based on tephra shards analyses and the occurrence of the agglutinated benthic foraminifer Spiroplectammina biformis (Jennings et al., 2014), which was associated with glacial meltwater (Jennings and Helgadottir, 1994; Schafer and Cole, 1986). Hence, low surface salinity and stratification of the upper water column may have resulted from the occurrence of low salinity-cold surface waters, overlying warmer and more saline bottom waters linked to Atlantic water inputs. Glacier runoff probably slowed down as the glacier became land-based around 7700–7500 cal. yr BP (Seidenkrantz et al., 2013; Young et al., 2011). Widespread mollusc colonization and organic accumulation in coastal lakes began some 7500 years ago, after the retreat of the Jakobshavn Isbrae and/or GIS margin (Briner et al., 2010; Donner and Jungner, 1975; Fredskild, 2000). The postglacial warming along the southwest Greenland coastline was possibly delayed because of meltwater runoff along the Greenland margins, as documented from diatoms in the Ameralik fjord where the WGC reached the surface only by ~7800 cal. yr BP (Ren et al., 2009; Seidenkrantz et al., 2013) as well as from many coastal lake pollen records (cf. Fréchette and de Vernal, 2009; cf. Figure 6).
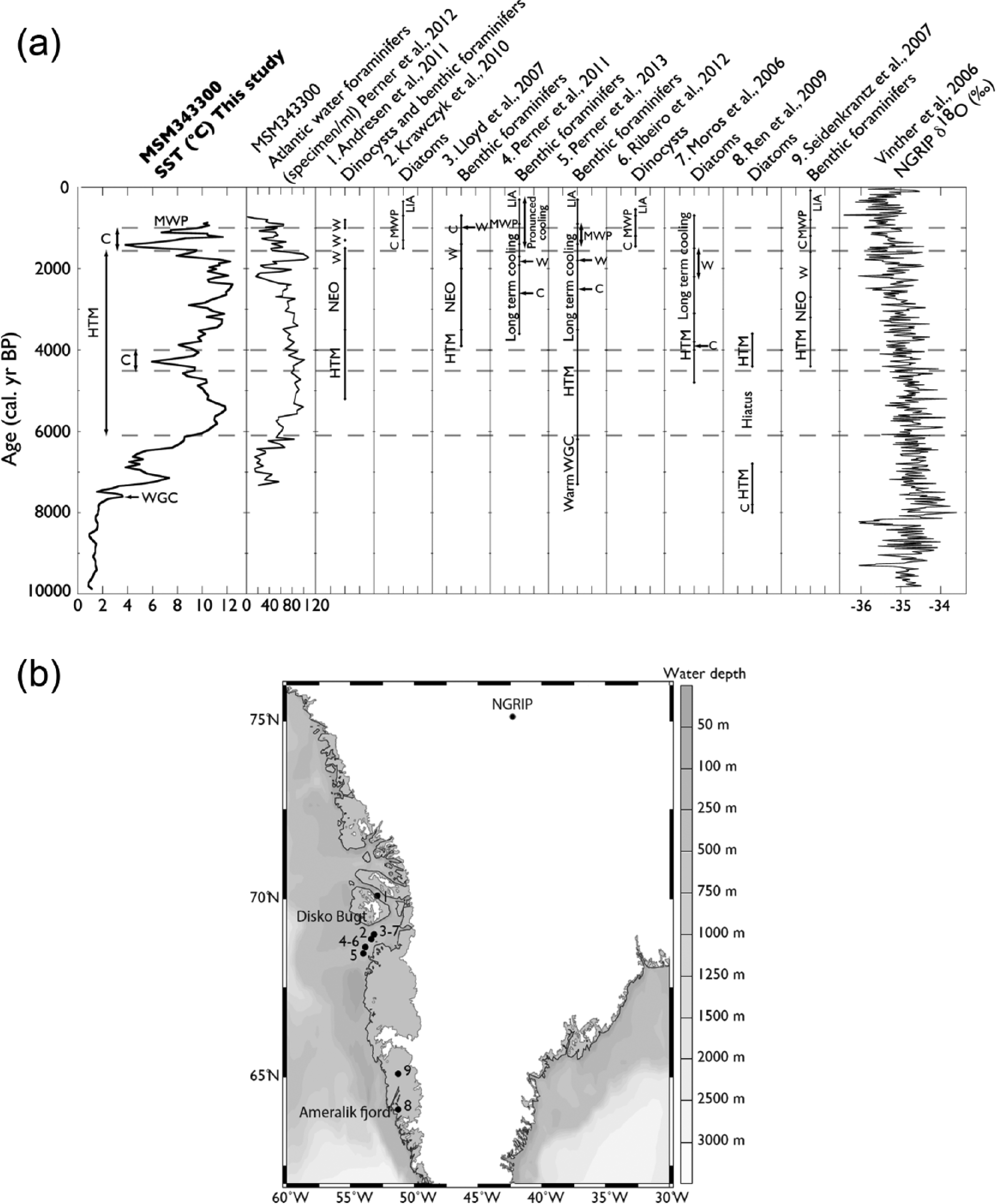
(a) Comparison of sea-surface temperature from core MSM343300 (this study) with reconstruction from adjacent marine cores of Southwest and West Greenland, and with the NGRIP δ18O record (Vinther et al., 2006; graphic representation). (b) Map of the sites referred to the sketch of Figure 6a.
The light 13C excursion recorded by both benthic foraminifer species at ~8200 cal. yr BP deserves some attention. Based on the micropaleontological data presented here, neither surface conditions nor benthic production or bottom water properties have changed significantly during the interval spanning from ~8200 to ~7000 cal. yr BP. We are thus tempted to link this excursion to a methane seepage event on the seafloor (Kaneko et al., 2010). Further geochemical analyses are underway to document this hypothesis.
First WGC influence ~7300 cal. yr BP
In surface waters of the Disko Bugt, the northward penetration of the WGC likely occurred at about 7300 cal. yr BP as shown by dinocyst-based reconstruction of SSTs and sea-ice cover. This transition was marked by an increase in dinocyst species diversity, with the significant occurrence of P. dalei, O. centrocarpum, and S. ramosus, in addition to S. quanta. From ~7300 to ~6000 cal. yr BP, large amplitude warming of surface waters was recorded. The change in dinocyst assemblages led to reconstruct SSTs increase from 3°C to 10°C in summer, which was accompanied by a decrease in sea-ice cover from >8 to <3 months/yr. This change was also marked by an increase of annual productivity, which reached up to 350 gC/m2/yr. The transition from ~7300 to ~6200 cal. yr BP is also recorded in bottom waters as shown by the synchronous increase of benthic foraminifer species associated with Atlantic waters, such as I. norcrossi and Cassidulina reniforme (Figure 6; Perner et al., 2013).
It is notable that the SST increase at ~7300 cal. yr BP was accompanied with a salinity decrease in surface waters, which suggests that warmer condition may have led to an accelerated glacier melting, resulting in enhanced freshwater discharges from the Jakobshavn Isbrae and/or the Greenland margins. Therefore, our data support the hypothesis of a delayed influence of WGC in the surface waters as compared with seafloor conditions, because of high meltwater supplies (Lloyd et al., 2005).
Mid-Holocene optimal conditions from ~6000 cal. yr BP
Optimal conditions were established in surface waters from ~6000 cal. yr BP. The thermal optimum was locally characterized by sea-ice cover restricted to the winter season (~3 months/yr), high primary productivity (up to 300–400 gC/m2/yr), and summer SST reaching up to 10–12°C. The general salinity decrease to about 29 psu was possibly related with meltwater discharge from the ice margin when Jakobshavn Isbrae recorded its maximal retreat, which occurred by ~4000 cal. yr BP (Weidick, 1992; Weidick et al., 1990). Many paleoclimate studies of the southwest and western Greenland are consistent with the reconstructions of sea-surface conditions presented herein. Most of them suggest that optimum conditions were attained relatively late, from ~6000 to ~3500–3000 cal. yr BP (cf. Figure 6). In particular, Perner et al. (2013) have described an episode of high phytodetritus supply, coherent with the high surface primary productivity we reconstructed based on dinocyst assemblages. Moreover, St-Onge and St-Onge (2014) documented the ice margin retreat to the present position from 7000 to 6000 cal. yr BP based on magnetic susceptibility and density.
The dinocyst-based reconstruction shows cooling pulses at ~4200–4000 (down to 6°C) and ~1500–1000 cal. yr BP (down to 3–4°C). They could be associated with the Neoglacial cooling phase (e.g. Dahl-Jensen, 1998; Kaufman, 2004; Kelly, 1980; Miller et al., 2005). Hence, these cooling pulses seem to represent a consistent feature on regional scale. Diatom data from Disko Bugt also show a temperature decrease between ~4000 and ~3800 cal. yr BP (Moros et al., 2006). Similarly, in Ameralik Fjord, an episode of extensive sea-ice cover and colder surface water was observed from ~4400 to ~3600 cal. yr BP (Ren et al., 2009). The Neoglacial cooling period recorded from ~3500 to ~2000–1800 yr BP in many regional reconstructions of climate changes (Andresen et al., 2010; Krawczyk et al., 2010; Lloyd et al., 2007; Perner et al., 2011, 2013; Figure 6) can be correlated with a Jakobshavn Isbrae glacier readvance (St-Onge and St-Onge, 2014). Moreover, the second cooling phase we reconstruct at ~1500 cal. yr BP was also reported from microfossil data in the Disko Bugt area as the so-called Dark ages period. Krawczyk et al. (2010) and Seidenkrantz et al. (2008) reconstructed a cooling from ~1500 to ~1300 cal. yr BP based on diatom and dinocyst data. Seidenkrantz et al. (2007) suggested a cooling period from ~1600 to ~1200 based on benthic foraminifers analyses. Jensen et al. (2004) and Ribeiro et al. (2012) documented a cooling event from ~1450 to ~1250 yr BP (cal.
In addition to SST changes, the dinocyst data permitted to reconstruct surface salinity changes, which ranged from 29 to 32 psu during the interval. A striking feature is the almost systematic opposite trend of surface-water salinity and summer temperature: warm episodes correspond to low salinity conditions, and vice-versa, the coldest phase being marked by the highest salinity. The salinity versus temperature variations in the study core tend to support the hypothesis of accelerated Jakobshavn Isbrae melting and freshwater discharges under warm conditions in relation to the strength and/or the temperature of the WGC.
In bottom sediments, benthic foraminifers became abundant enough around 6000 cal. yr BP to establish a continuous isotopic record. The δ18O values in I. norcrossi slightly decreased from ~6000 to ~5000 cal. yr BP (from ~3.45‰ to 3.2‰), which might correspond to a temperature rise (≤1°C) in bottom waters. This is consistent with the increase of I. norcrossi relative to Arctic species such as Elphidium excavatum and Cassidulina arctica, which together suggest a stronger Atlantic component in the WGC or reduced meltwater supplies through the EGC (Lloyd, 2006a; Perner et al., 2013). Overall benthic foraminifer assemblages also showed a thermal maximum from ~6000 to ~5000 cal. yr BP, followed by a gradual cooling until ~3900 cal. yr BP (Lloyd et al., 2007; McCarthy, 2011; Perner et al., 2011, 2013).
δ18O values in I. norcrossi slightly increased (up to ~3.32‰) from ~2100 to ~1100 cal. yr BP, thus suggesting a temperature decrease of the bottom waters matching that reconstructed at the sea-surface. The corresponding change in benthic foraminifer assemblages might be associated with enhanced influence of the EGC (Jennings et al., 2011; Perner et al., 2011, 2013) relative to a decrease influence of the IC (Lloyd et al., 2007).
Finally, one should highlight the fact that the productivity variations reconstructed from dinocysts are consistent with the occurrence of the phytodetritus feeder N. labradorica from ~6000 to ~5100 and ~4000 to ~2800 cal. yr BP (Jennings et al., 2004; Lloyd, 2006b; McCarthy, 2011).
Medieval warming (~1000–800 cal. yr BP)
From ~1000 to ~800 cal. yr BP, summer SSTs increased to about 10°C, which is much higher than the present day summer SST of ~4.4°C ± 1.24°C at the coring site (NODC, 2001). In the dinocyst assemblage, I. minutum percentages decreased, whereas the S. elongatus percentages increase. We suggest that this event corresponds to a warming of the WGC. Benthic foraminiferal analyses also led to suggest enhanced contribution of IC to the WGC from ~1400 to ~900 cal. yr BP (Perner et al., 2013). This event can be associated with a regional signature of the ‘Medieval Warm Period’ as described, for example, by Dahl-Jensen (1998) from Greenland ice cores and based on many other paleoceanographical proxies of the Disko Bugt and western Greenland areas (Andresen et al., 2010; Krawczyk et al., 2010; Lloyd et al., 2007; Perner et al., 2011; Ribeiro et al., 2012; Seidenkrantz et al., 2007; Figure 6). Correlation with records from southern and eastern Greenland (Anderson and Leng, 2004; Jensen et al., 2004; Kaplan, 2002; Lassen et al., 2004) and with Icelandic shelf (Eiríksson et al., 2000; Jiang et al., 2002) records might also be proposed.
Conclusion
We have attempted here to reconstruct paleoceanographical changes along the West Greenland margin during the Holocene from a multi-proxy approach with a better than centennial time resolution for some proxies. Dinocyst-based reconstructions provided information on sea-surface conditions and permitted to distinguish both temperature and salinity signals as a complex response to the northward flow of the WGC and freshwater discharge from the GIS. Data indicate glaciomarine conditions until ~10,000 cal. yr BP, while the earliest evidence of some Atlantic influence through the WGC appeared in deep water, when benthic foraminifer I. norcrossi was first recorded in this core. The WGC influence was recorded much later in surface waters, which were characterized locally by cold conditions with a dense ice cover until ~7300 cal. yr BP, likely because of important discharge of ice and meltwater from the GIS. After a gradual transition marked by an increasing influence of Atlantic waters, optimal conditions finally settled at ~6000 cal. yr BP. Some cooling pulses were recorded later on, at ~4200–4000 and ~1500–1000 cal. yr BP, alternating with warm phases at ~6000–4200, ~4000–1500, and ~1000–800 cal. yr BP. Throughout the postglacial, there is an opposition between SSTs and surface salinities, with warmer intervals being characterized by more diluted surface water. We thus associate the warming linked to increasing influence of Atlantic waters through the WGC, to phases of enhanced melt, thus resulting in higher freshwater discharge from Greenland ice margins and notably from the Jakobshavn Isbrae. The Holocene paleoceanographical record of Disko Bugt presented herein tends to demonstrate close linkage between the ocean and ice dynamics.
Footnotes
Acknowledgements
This study is a contribution to the project Past4Future of the Framework 7 Framework Program. We are grateful to Maryse Henry, Audrey Limoges, and Nicolas Van Nieuwenhove for their precious help with dinocyst identification and interpretation. We also thank Jean-François Hélie for his help with stable isotope analyses.
Funding
The authors thank the Ministère du Développement Économique, Innovation et Exportation (MDEIE) and Fonds Québécois de Recherche sur la Nature et les Technologies (FQRNT), and the Natural Sciences and Engineering Research Council of Canada (NSERC).