
Abstract
The distribution pattern of relict and specialised species in calcareous fens was revealed to be non-stochastic, with ancient fens harbouring more of these species than younger ones. This phenomenon could be caused by long-lasting in situ survivals over millennia, but direct palaeoecological evidence is lacking. We addressed the question whether at least some ancient calcareous fens indeed retained open-fen patches throughout the Holocene, using a palaeoecological approach involving proxies with different taphonomies (pollen, vascular plants, bryophytes, molluscs). We identified three old fens in the Western Carpathians, where several postglacial relict species have recently been found, and we reconstructed their histories with respect to sedimentary processes, vegetation structure and dynamics of relict species. The development at all the sites started with a (semi)-open fen community dominated by sedges and brown mosses. The site with the highest recent number of relict species was reconstructed to harbour open patches continually since the late Glacial to the present, including the middle Holocene when open-fen patches were restricted. By contrast, at the site with the lowest recent number of relict species, a large sedimentary hiatus suggested peat mineralisation or erosion that prevented the survival of light-demanding species. At all the sites, characteristic snails of European Glacial periods occurred during fen initiation, but disappeared around the early/middle Holocene transition. The probability of a relict species being present in a modern fen community increases with fen age, but it also depends on the continual existence of open-fen patches and peat accumulation throughout the middle Holocene.
Introduction
Understanding and predicting biodiversity loss is one of the most serious problems in current ecology. Palaeoecology may contribute to this question by providing data about the rate and spatial dynamics of extinctions, survivals and coexistences by studying the fossil record containing evidence of rapid climate changes over long time scales (Seddon et al., 2014). Unfortunately, ecology and Quaternary palaeoecology have largely developed as parallel disciplines, and only a few palaeoecological studies have addressed ecological questions or focussed on dynamics of non-woody species. Closing the gap between ecology and Quaternary palaeoecology is an important and topical challenge for ecologists and palaeoecologists (Reitalu et al., 2014; Seddon et al., 2014). One long-standing question at the interface between ecology and Quaternary palaeoecology is, ‘Why are some boreal and continental plant and animal species, many of which are highly endangered, so unpredictably scattered across temperate fens?’.
Ancient calcareous fens are fascinating ecosystems acting as refugia for many highly specialised species (Hájek et al., 2011; Wassen et al., 2005). In the temperate zone, they harbour a suite of species that display an island-like scattered distribution and that are recently critically endangered (e.g. Hampe and Jump, 2011; Schenková and Horsák, 2013). Since the beginning of biogeography in Central Europe, these species have been considered relicts of the cold past and were therefore called glacial relicts (Holmquist, 1962; Rybníček, 1966); the term corresponds to the terms postglacial relicts, relicts since postglacial times and cold-stage relicts, which have been used more recently (Hájek et al., 2011). The relict concept (Holmquist, 1962) was very popular in the descriptive phase of Central and Southern European field botany and zoology, but later, it was abandoned by mainstream science because of the difficulties of obtaining unequivocal evidence of relict distribution patterns of some species. However, this concept has been persisting, mostly in the world of local studies, local nature conservancy and enthusiastic nature lovers. The revisiting of the relict concept was triggered by the development of phylogeographic methods (e.g. Reisch et al., 2003; Vogler and Reisch, 2013), although even in this field, the cold-loving postglacial relicts still stay in the shadow of true glacial relicts (from the phylogeographic perspective), i.e., the warmth-demanding species that survived the Full Glacial period in small pockets with favourable mesoclimate conditions (e.g. Stewart et al., 2010).
In our previous study (Hájek et al., 2011), we started to revitalise the concept of postglacial relicts for the archipelago of calcareous fens in the Western Carpathians (Central Eastern Europe, Slovakia). In that study, we dated 47 recently well-preserved calcareous fens and demonstrated statistically that a suite of plant and mollusc species was significantly linked to old fens at the millennial scale, independent of the effect of recent fen area. High age of a fen site determines the occurrence of those species that are indeed rare, endangered and unevenly distributed in temperate Europe (Horsák et al., 2012). Although this result brought certain evidence for the validity of the relict concept for the archipelago of Central European calcareous fens, some questions remained unresolved. We observed certain differences in stratigraphy among particular ancient fens. Some of them contained phases rich in wood remains, while others seemed to be continuously open, but detailed sequential palaeoecological analyses were not performed (Hájek et al., 2011). Particular sequences contained different species at their bottom, and some of them contained snail indicators of glacial landscapes (Hájková et al., 2012b).
In addition, our other studies have suggested that the recent plant and snail communities of young fens have been assembled predominantly during the Wallachian colonisation taking place from 14th to 17th centuries (Hájková et al., 2012b; Jamrichová et al., 2013). It was a large colonisation wave from the eastern part of the Carpathians to the west, which completely changed so far poorly settled mountain landscapes. The large areas were deforested for the pastoral purposes, forest springs were opened as well, and many new spring-fens have originated on the landslides. Thus, we cannot reject the assumption that such a huge landscape transformation also altered somehow the ancient fen biota. Hence, it is unrealistic to suppose that relict assemblages of ancient fens are completely the same as those that occurred in the late Glacial or early Holocene. Furthermore, we still have not obtained a coherent set of evidence for continuous in situ survival of light-demanding fen species during the middle Holocene forest optimum in the Western Carpathian Mountains. Scattered data came from some undated snail sequences (Ložek, 1964) and for Cladium mariscus in one hot spring lowland fen (Hájková et al., 2013).
For a more thorough understanding of the dynamics of rare, relict fen species in temperate landscapes, it is therefore necessary to focus in more detail on the most representative fen peat sequences and to reconstruct their history using identifications of their fossils. In minerotrophic soligeneous fens, however, this approach does not always bring sufficient evidence supporting or rejecting the hypotheses about continual Holocene survival. Fen sequences often contain intervals where either pollen microfossils or plant macrofossils (determinable to species level) were not preserved in sufficient numbers. Nevertheless, our previous studies (Hájková et al., 2012b; Jamrichová et al., 2013) have demonstrated that the combination of fossils with different taphonomic modes (pollen, snail shells and plant macrofossils) leads to more direct and more interpretable evidence of environmental changes in the Holocene. We therefore applied this multi-proxy approach to all three recently known well-preserved fens in the intermountain basins of the Inner Western Carpathians, which have at least four important site characteristics in common: (1) they are old, with peat initiation in either the late Glacial or at the onset of the Holocene; (2) they favour fossilisation of both snail and plant remains; (3) they harbour postglacial relicts (tested successfully by Hájek et al., 2011) in recent assemblages; and (4) they contain characteristic species of glacial landscapes in the bottom layers (data from Hájková et al., 2012b).
From these three fens, we took sequences of their deposits in order to address the following questions: (1) did open, treeless fen patches persist in these fens during the entire Holocene? If there are differences in their extent and continuity in particular profiles, do they correspond to the level of extinction at a site and to a present-day representation of relict species? (2) Is it possible to find a threshold when the species characteristic of glacial landscapes became extinct? (3) To what extent do the present-day fen assemblages correspond to ancient ones and how did the late Holocene Wallachian colonisation affect them? (4) If the late Glacial assemblages differ, are they at least partially analogous to the present-day assemblages in boreal or continental landscapes? and (5) Which macroclimatic conditions could be reconstructed for the late Glacial stadial in the Inner Carpathian basins based on the past occurrences of snail species?
Materials and methods
Characteristic of the study sites and field sampling
The three preserved fen sites that fulfilled our pre-selection criteria are located close to each other, in the intermountain basins of north-eastern Slovakia (Figure 1), namely in the Poprad Basin (Hozelec – HOZ), at the margin of the Ľubovňa Basin (Valalská voda – VAL) and in the Liptov Basin (Brezové – BRE). The study sites are located in the montane belt in altitudes of 677 m.a.s.l. (HOZ), 714 m.a.s.l. (VAL) and 894 m.a.s.l. (BRE). All these sites can be classified as calcareous spring-fens supplied by an alkaline mineral-rich groundwater (pH 7–7.8; water conductivity varied from 360 µS cm−1 in BRE to 918 µS cm−1 in HOZ). Vegetation growing in all study sites belongs to the association Caricetum davallianae Dutoit 1924 (for more information about floristic composition, see Dítě and Pukajová, 2004 and Hájek et al., 2006). Snail assemblages are species-rich and include several threatened wetland specialists and relict species (for a complete species composition of recent assemblages, see Appendix S3, available online). The sites differ in their recent size. The Brezové is the largest fen out of the study sites today (about 11.500 m2), Valalská voda is smaller (ca. 1900 m2) and Hozelec is the smallest one with only ca. 60 m2; however, this site was a part of a large fen complex in the past being destroyed by human activities. All sites represent remnants of large fen complexes, and therefore they are protected and managed by nature conservancy. The regular mowing prevents overgrowing by trees and supports well-developed bryophyte layer with many relict species. Without this management, probably only the wettest places with spring pools would remain open.
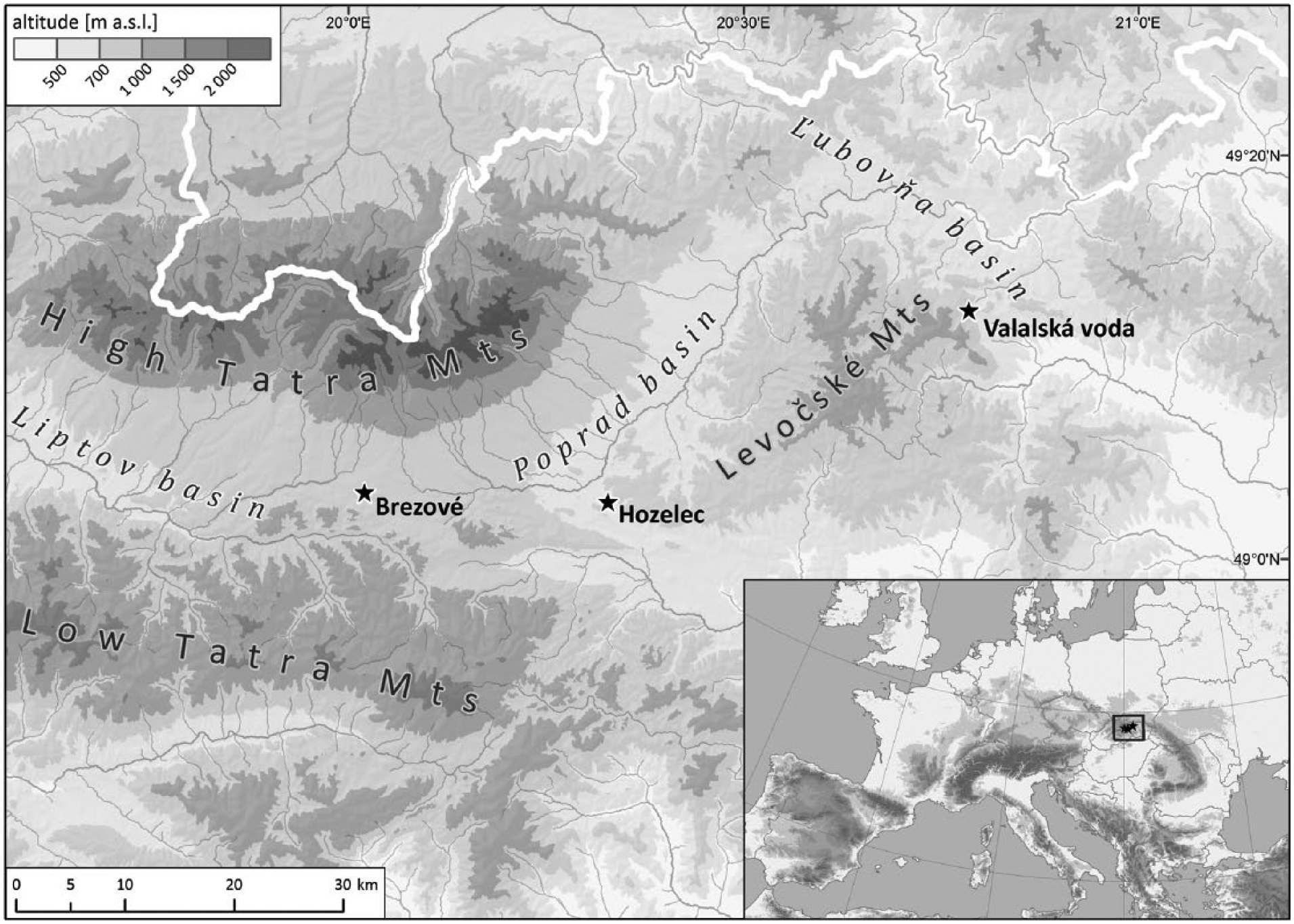
Map of the study area.
These sites differ in numbers of postglacial relict species (Table 1), with the lowest in the Valalská voda fen and the highest in the Hozelec fen. There are mostly species strongly specialised to calcareous fens with a preserved hydrological regime, i.e., stable water level around the surface for almost the whole year (e.g. plants Campylium elodes, Carex dioica, Primula farinosa and Triglochin maritima; snails Euconulus praticola, Pupilla alpicola and Vertigo geyeri). Some relict species display wider ecological niche occurring also in the moderately rich fens with lower amount of calcium in groundwater (Drepanocladus trifarius, Carex diandra and Hamatocaulis vernicosus). All relict species are related to open habitats and are not able to survive in a close canopy. Also the increase in nutrient supply can be detrimental for these species because they are competitively poor and enhanced amount of nutrients promotes competitively strong tall-herb vascular plants and ubiquitous grassland bryophytes such as Calliergonella cuspidata (Hájek et al., 2014).
Characteristics of studied calcareous fens: Hozelec (49.045278N/20.334167E; 677 m.a.s.l.), Brezové (49.050833N/20.028333E; 891 m.a.s.l.) and Valalská voda (49.205N/20.786389E; 714 m.a.s.l.) and the number of relict species recently occurring in a site. The first number refers to the number of calcareous fen relicts predicted by null model (Hájek et al. 2011), and the second one to the number of arbitrary assessed relicts (the species, which are not obligate calcareous-fen specialists and hence they were not tested in Hájek et al. 2011).

Bryo: bryophytes; vasc: vascular plants.
At Valalská voda and Hozelec, we sampled a single peat profile from an open excavation. Peat for mollusc analyses was sampled separately to obtain sufficient volume of material. Peat for macrofossil and pollen analyses was stored in metal boxes (50 cm × 10 cm × 10 cm). At the Brezové fen, we sampled two profiles: one in a place with extremely high calcium carbonate precipitation today and another in a place with shallow pools with less calcium carbonate precipitated. The first profile was obtained using a percussion drilling set because of difficulties with manual drilling. In order to obtain enough material for mollusc analysis, we took three parallel cores located close to each other (BRE-A1, BRE-A2 and BRE-A3). The second peat profile was sampled from an open excavation to metal boxes (BRE-box). For more details about peat composition, see Appendix S1, available online.
Radiocarbon dating
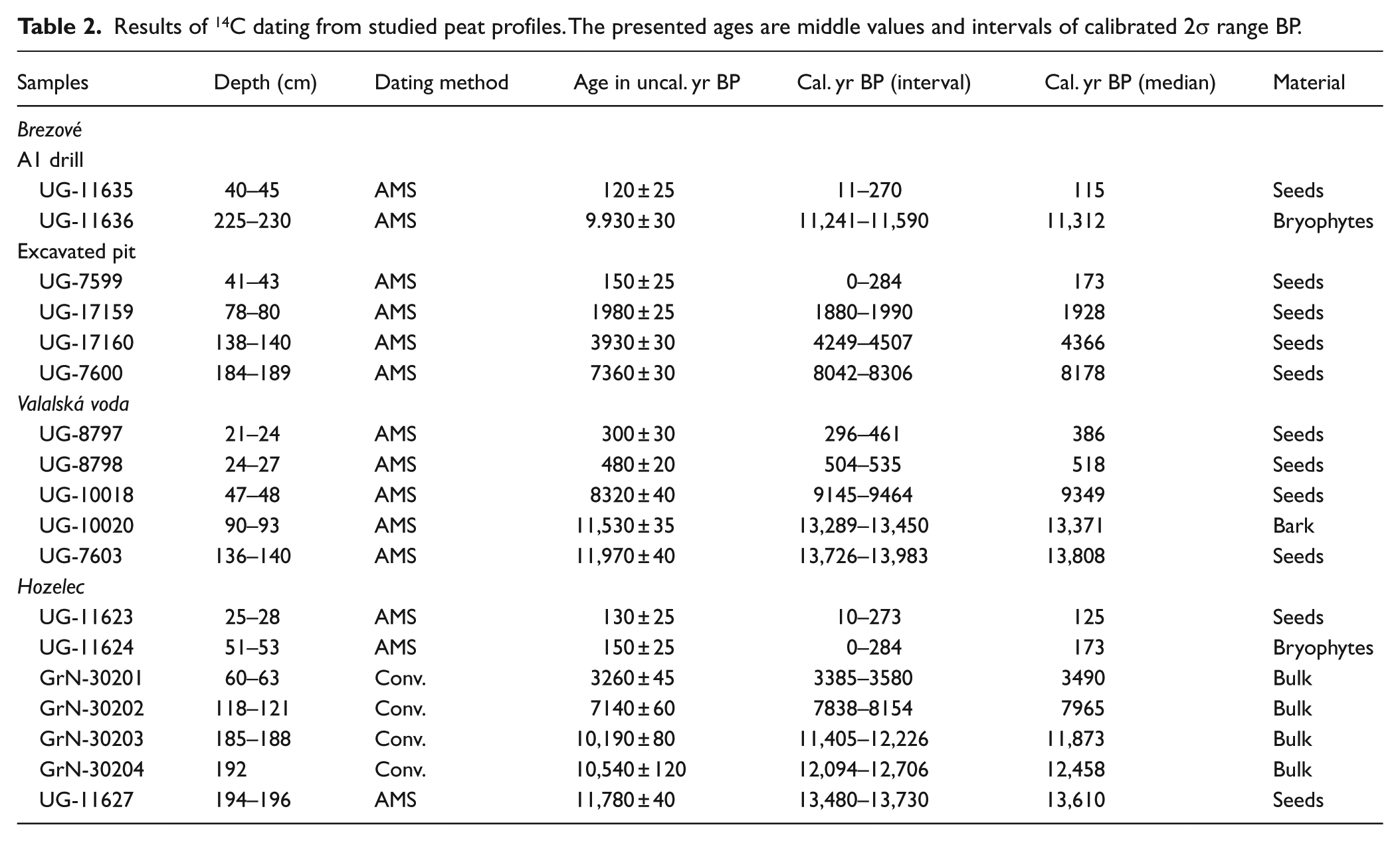
Macrofossils of chosen samples were radiocarbon-dated at the Center for Applied Isotope Studies, University of Georgia, Athens, USA (for details, see Table 2). We have strictly chosen seeds of terrestrial plants (mostly Carex seeds) and terrestrial bryophytes (Tomentypnum nitens and Bryum pseudotriquetrum) to avoid the hard-water error (Grimm et al., 2009). At the Hozelec site, some layers did not contain suitable macrofossils for dating; therefore, we had to use the bulk for the dating. The bulk dates appear reliable as they fit existing knowledge on vegetation development in the region (Jankovská, 1988). Possible biases caused by the bulk dating cannot alter our main result on vegetation succession at the Holocene scale. Bulk samples were analysed at the Centre for Isotope Research, University of Groningen, the Netherlands. Data were calibrated using the IntCal13 calibration curve (Reimer et al., 2013) with OxCal4 software version 4.2 (Bronk Ramsey, 2009), and the middle value of the calibrated 2σ range (before the year 1950) was used.
Results of 14C dating from studied peat profiles. The presented ages are middle values and intervals of calibrated 2σ range BP.
Laboratory analyses
Each sample for plant macrofossil analysis (volume of 100 cm3) was sieved into three fractions (>1 mm, 1–0.63 mm and 0.63–0.20 mm). The larger fractions were analysed using a stereomicroscope. A quantity of 1 mL of the finest fraction was examined at 100×–400× magnification in order to identify small seeds (e.g. Juncus spp.) or oogonia of Characeae. The material for pollen analyses was subsampled (volume of 1 cm3) at 10 cm (VAL, BRE-box) or 5 cm (HOZ) intervals along the profile and processed using standard techniques (Fægri and Iversen, 1989). Samples containing clastic material were pre-treated with cold concentrated hydrogen fluoride for 24 h and then potassium hydroxide solution before acetolysis. At least 300 pollen grains of terrestrial plants per sample were identified (HOZ profile). Because of low quantities of palynomorphs in VAL and BRE profiles, at least three microscopic slides were counted for each sample. Fossil mollusc shells in BRE-A1 and BRE-box were extracted from the same samples (volume of 100 cm3) as macrofossils, and in the VAL and HOZ, the molluscs were sampled separately (the approximate volume 500 cm3). Mollusc shells were obtained from recent surface samples by carefully washing each sample (12 L of bryophytes and litter) in a bowl-shaped sieve (mesh size 0.5 mm) to remove fine soil (see Horsák, 2003). After drying, the shells were separated from the remaining material. See Appendices S2–S4, available online, in Supporting Material for the list of all identification literature. All presented diagrams were created using C2 software (Juggins, 2003). The nomenclature follows Beug (2004) for pollen, Marhold and Hindák (1998) for vascular plants, Kučera et al. (2012) for bryophytes and Horsák et al. (2013) for molluscs. In the paper, we use the modern simplified division of the Holocene: early Holocene (11,700–8200 cal. yr BP), middle Holocene (8200–4200 cal. yr BP) and late Holocene (4200 cal. yr BP–AD 2000). P Hájková identified macrofossils in all profiles, M Horsák snails in Hozelec and Valalská voda profiles, J Moutelíková snails in profiles from the Brezové fen, E Jamrichová pollen in Hozelec profile and V Jankovská pollen in Valalská voda and Brezové profiles.
Other analyses
Several recent studies have shown that southern Siberian mountain ranges harbour biotas that act as the best modern analogies of those reconstructed for full-glacial Central Europe (e.g. Kuneš et al., 2008; Pavelková Řičánková et al., 2014). This was also confirmed by our malacological research in the Altai Mountains (e.g. Horsák et al., 2010). Here, we used macroclimate characteristics of 34 Altaian sites with the presence of four characteristic glacial snail species that were found in the bottom layers of the studied fen deposits. For details about the sampling of modern snails in 2005–2006 and 2011 and the obtaining of climatic data, see Horsák et al. (2010).
Results
For the complete information about species composition of fossil assemblages, see Figures 2–5 (showing synoptic palaeoecological diagrams with the selected taxa and ecological groups of taxa) and Appendices S2 (pollen), S3 (molluscs), S4 (macrofossils) and S5 (ecological groups), available online for full data.
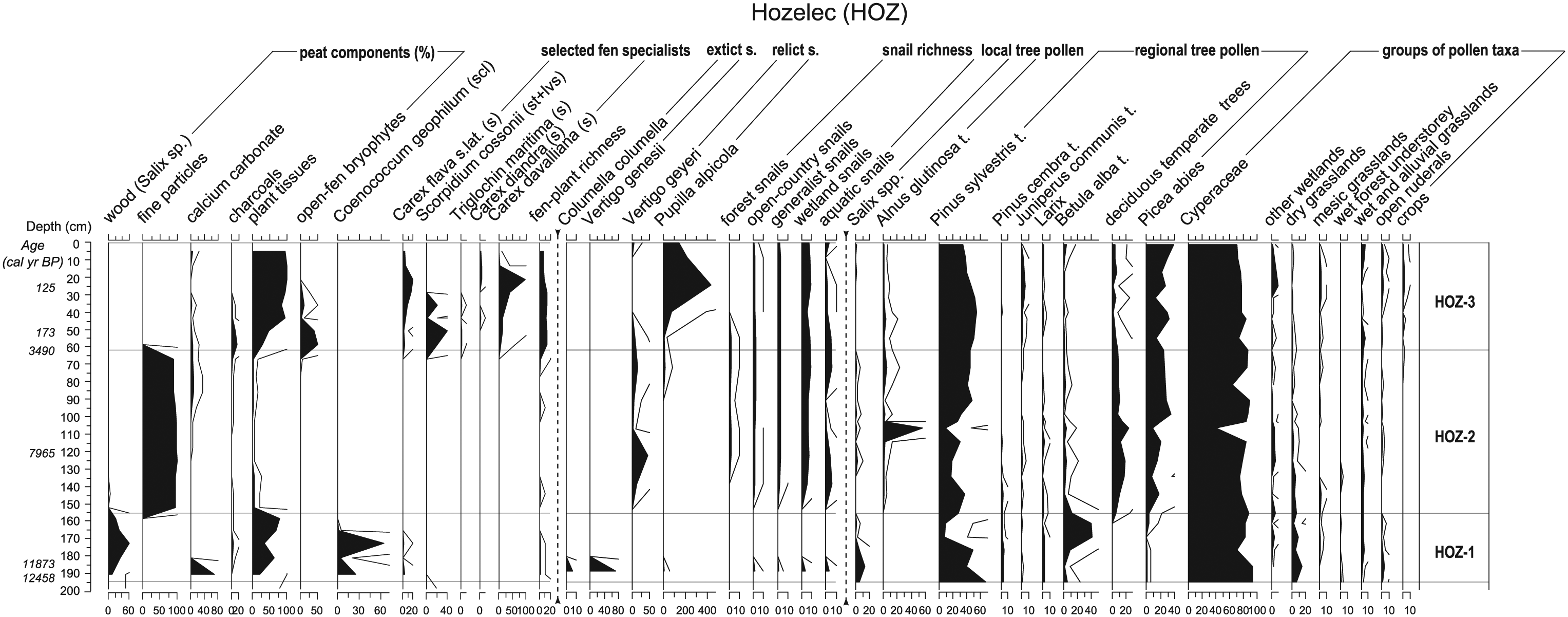
Diagram of particular peat components (in percentages), selected taxa of bryophytes, vascular plants, molluscs (in absolute values) and pollen taxa (in percentages) for Hozelec profile (HOZ). Furthermore, some ecological groups of taxa are shown (for included taxa, see Appendices S3, S5, available online). The total pollen sum (TS = AP (arboreal plants) + NAP (non-arboreal plants) = 100%) was calculated based on the terrestrial pollen sum; (semi)aquatic plants, Pteridophyta, algae, fungi, and the other non-pollen palynomorphs (NPP) were excluded. The percentages of (semi)aquatic taxa, spores and NPP were related to the extended sum (AP + NAP + (semi)aquatic + spores + NPP = 100%).
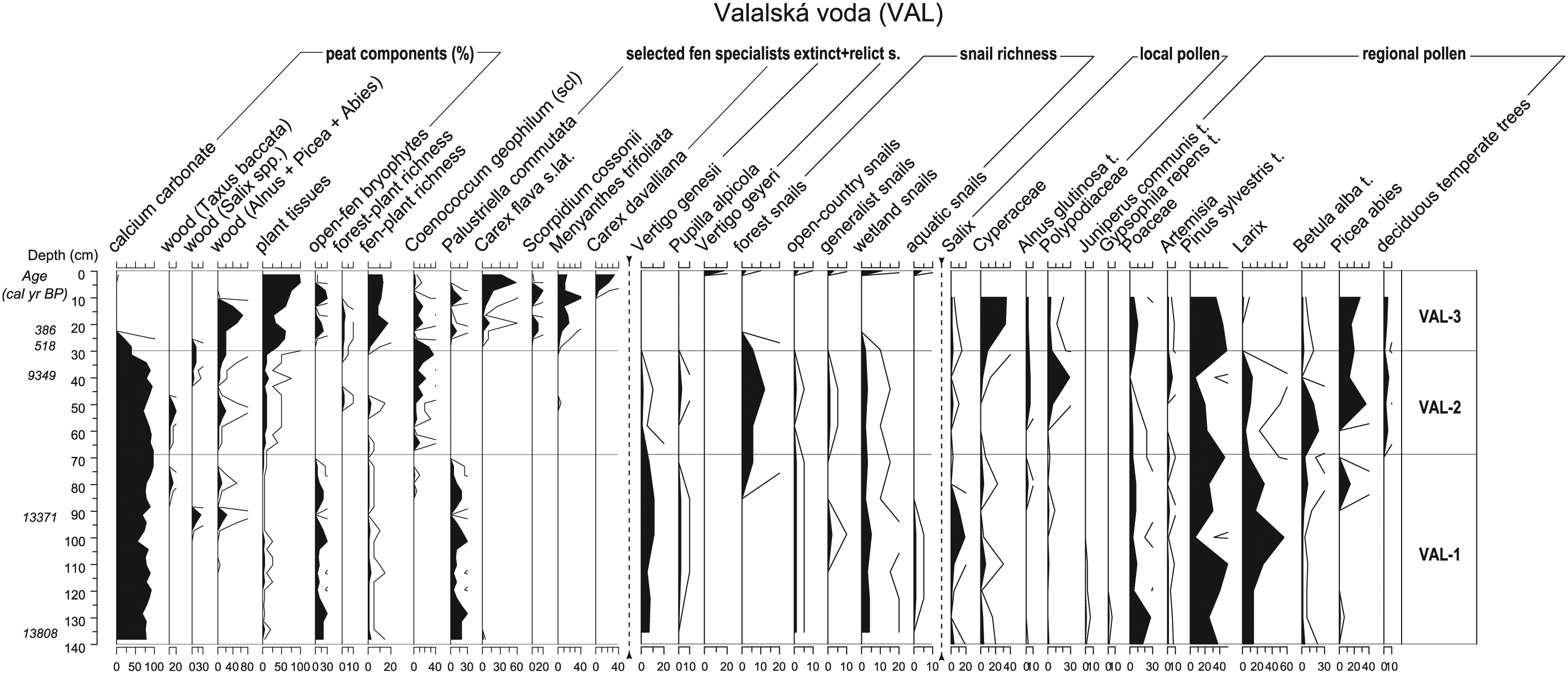
Diagram of particular peat components (in percentages), selected taxa of bryophytes, vascular plants, molluscs (in absolute values) and pollen taxa (in percentages; for details, see the legend of Figure 2) for Valalská voda profile (VAL). Furthermore, some ecological groups of taxa are shown (for included taxa, see Appendices S3, S5, available online).

Diagram of particular peat components (in percentages), selected taxa of bryophytes, vascular plants, molluscs (in absolute values) and pollen (in percentages; for details, see the legend of Figure 2) for Brezové profile (BRE-box). Furthermore, some ecological groups of taxa are shown (for included taxa, see Appendices S3, S5, available online).

Diagram of particular peat components (in percentages), selected taxa of bryophytes, vascular plants and molluscs (in absolute values) for Brezové profile (BRE-A1). Furthermore, some ecological groups of taxa are shown (for included taxa, see Appendices S3, S5, available online).
Development of the landscape context
On the basis of pollen analysis (see Figures 2–4 and Appendix S2, available online), the late Glacial landscape was reconstructed to be covered by hemiboreal forests in the mosaic of tundra and steppe habitats. Forests were mainly composed of Larix (dominant), Pinus cembra, P. sylvestris/mugo (dominant), Betula pendula/pubescens and Picea abies, which probably occurred in humid and microclimatically favourable places. However, the studied sites differed in the abundance of individual tree taxa. Pollen of Larix decidua/sibirica (Larix type) showed the highest abundance around the Valalská voda fen, whereas the representation of P. cembra was the highest at the Hozelec site and the species was almost absent at the Valalská voda site. Sorbus aucuparia was locally present with a low abundance (VAL). The understorey was composed of, for example, Polygonatum verticillatum, Valeriana cf. tripteris (pollen type V. montana) and cf. Scilla bifolia/kladnii (the Scilla type). The two former species nowadays occur in mountain coniferous forests, but the latter species occurs in either deciduous or alluvial forests (its occurrence in open hemiboreal forests of the late Glacial cannot be excluded). Treeless vegetation consisted of steppe–tundra species such as Artemisia sp., Chenopodiaceae, Botrychium sp., Selaginella selaginoides, cf. Gypsophilla repens (pollen type of the same name), and Ephedra sp. (Ephedra fragilis type), with scattered shrubs of Juniperus communis and Betula nana/humilis (few pollen grains of Betula nana type). Pollen of other, perhaps heliophilous species such as Ranunculus sp. (R. acris type), Silene sp. (S. vulgaris type) and Potentilla sp. were recorded at the Hozelec site that provided the best preserved pollen record.
The rise in temperature in the early Holocene was at first reflected by a decline of P. sylvestris/mugo and steppe and tundra elements, along with the increase of Picea abies and Betula pendula/pubescens. Later, some deciduous temperate tree species (e.g. Corylus avellana, Quercus spp., Tilia spp. and Ulmus spp.) started to spread, but the pattern was locally different. The late Holocene period was, in the study region, characterised by the expansion of Abies alba, whereas Fagus was only sporadic compared with the records from other Central European mountains. Larix continually occurred from the late Glacial to present times.
Development of fen communities from the late Glacial to middle Holocene
The late Glacial and early Holocene calcareous fens were characterised by calcium carbonate precipitation and by light-demanding fen plant and mollusc communities (for molluscs, see Appendix S3, available online and for plant macrofossil, Appendix S4, available online). In the bryophyte layer, we recorded light-demanding species such as Scorpidium cossonii (BRE-A1, HOZ), Tomentypnum nitens (BRE-A1), Palustriella commutata (syn. Cratoneuron commutatum, VAL, HOZ), Campylium stellatum and Ditrichum crispatissimum (syn. D. gracile, HOZ). Mollusc communities were composed of common open-country species such as Vallonia pulchella (BRE-A1, HOZ), and species of humid or marshy habitats Succinea putris (BRE-A1), Nesovitrea hammonis (HOZ) and Euconulus praticola (BRE-A1), but also contained several characteristic glacial species such as Columella columella (BRE-A1, HOZ), Vertigo genesii (all sites), V. modesta and V. parcedentata (BRE-A1). The bottom layers of all three sequences contained Pupilla alpicola, a calcareous-fen specialist that has survived to the recent time in HOZ, but is extinct in VAL as well as in BRE. The presence of small calcareous pools was indicated by oogonia of Chara vulgaris (HOZ) and Potamogeton seeds and pollen (HOZ). Fen vegetation was represented by sedges Carex flava agg. (HOZ, VAL), C. nigra (VAL), C. rostrata (HOZ) and Trichophorum pumilum (BRE-A1). The presence of Salix shrubs at fen sites was documented by both wood remains in the fen deposits (BRE-box, VAL and HOZ) and also by pollen (all localities). Also, trees or shrubs of Taxus baccata might occur in the studied calcareous fens during the late Glacial and early Holocene, as evidenced by wood remains found in the Valalská voda deposit (see Hájková et al., 2012b for other fossil records of Taxus in fens). Pollen assemblages further suggested the existence of wet meadows and tall-herb vegetation at the fen margins, as evidenced by pollen of Scorzonera cf. humilis (pollen type of the same name), Filipendula sp., Polygonum cf. bistorta (pollen type of the same name) and Thalictrum sp.
During the middle Holocene, all glacial land-snail species except P. alpicola disappeared definitely from all three study fens. Nevertheless, the successional development differed among these fens. At the Hozelec site, the shallow water basin (or possibly a small pool) had developed. Although the plant macrofossil record was very poor (Valeriana cf. simplicifolia, Cirsium sp. and Equisetum sp.), the other proxies, i.e., aquatic molluscs (e.g. Valvata cristata and Anisus leucostoma), algae (Botryococcus sp.) and pollen (Oenanthe cf. aquatica and Potamogeton natans type.) clearly indicated a high and stable water level. Besides aquatic communities, semi-terrestrial wetland habitats, including calcareous fens were developed, as indicated by pollen (Cyperaceae, Cirsium sp., Equisetum sp., Peucedanum cf. palustre, Mentha sp., Lysimachia cf. vulgaris, Valeriana cf. dioica, V. cf. officinalis and Iris sibirica; Appendix S2, available online) and molluscs (Euconulus praticola, Oxyloma elegans and Vertigo angustior; Appendix S3, available online). Species-rich open-fen mollusc communities were evidently present during the entire middle Holocene and contained, among others, highly specialised fen snail Vertigo geyeri, which started to spread after the late Glacial/Holocene transition and which replaced here V. genesii, a species characteristic of the late Glacial. Based on pollen data, the site was surrounded by a spruce–alder carr, and a high water level had most likely been a crucial factor in keeping at least the central part of the site open.
At the Brezové site, we analysed two profiles of different age. The older profile (BRE-A1) captured the patch that was overgrown by spruce–alder carr already in the early Holocene, as indicated by a high amount of wood remains. Only small pools with Chara vulgaris and their banks could remain at least partially open in the middle Holocene. The younger profile (BRE-box) captured a patch where peat initiation occurred later, at the beginning of the middle Holocene (about 8200 cal. yr BP). At that time, ancient fen vegetation was composed of boreal bryophytes Calliergon giganteum, Meesia triquetra and Scorpidium scorpioides (the two latter species are recently extinct at the site).
The sediment of the basal 30 cm (BRE-box) contained calcium carbonate, which allowed fossilisation of mollusc shells that were missing in the rest of the profile, with the exception of sub-recent layers (0–19 cm). Three relict species still occurred at the study site (Columella columella, Pupilla alpicola and Vertigo genesii; Figure 4) at the beginning of the middle Holocene (ca. 8200 cal. yr BP). The presence of shallow pools was indicated by oogonia of Chara vulgaris, algae Pediastrum boryanum and seeds of Potamogeton filiformis. Among sedges, Carex flava s.lat., C. paniculata and Blysmus compressus occurred (Appendix S4, available online). This part of the fen was also overgrown by a rather dense spruce–alder carr later in the middle Holocene (from 140 cm upwards; about 4400 cal. yr BP). This woodland phase was characterised by forest spring vegetation with Carex remota and Glyceria nemoralis. At the Valalská voda site, the middle Holocene record was completely missing, which could be caused either by strong mineralisation under the dense alder carr or by repeated water erosion by a nearby brook.
Late Holocene development
The calcareous sedge–moss vegetation resembling the modern vegetation was largely established as late as in the late Holocene, about 500 cal. yr BP at the Valalská voda site, about 140 cal. yr BP at the Brezové site (consistent result from both profiles), and between 3500 and 150 cal. yr BP at the Hozelec site; exact dating was impossible because of a high rate of mineralisation or presence of sediment hiatus in this zone. The modern calcareous fen vegetation was partially composed of the same vascular plants (e.g. Carex flava agg., C. paniculata, C. nigra, Linum catharticum and Menyanthes trifoliata) and bryophyte species (Campylium stellatum, Palustriella commutata and Scorpidium cossonii) that occurred previously in the older periods (for details, see Appendix S4, available online). On the other hand, an important group of species characteristic of present-day communities was found only in layers dated to the period after modern deforestation (Carex davalliana, C. dioica and Eriophorum latifolium).
Similar to the plant assemblages, the modern mollusc assemblages established rather recently (BRE-A1 and VAL) except for the Hozelec fen, where a high number of recently occurring species already occurred during the middle Holocene (about 8000 cal. yr BP) and may thus represent continually persisting relicts. Species composition during the middle Holocene was very similar to that found recently and differed only by the presence of aquatic species. On the contrary, most of the recently occurring land-snail species appeared not long ago at the Valalská voda fen, but this result may be partially confounded by the low species richness of fossil samples. An intermediate situation could be reconstructed at the Brezové site.
Ecology of relict snail species
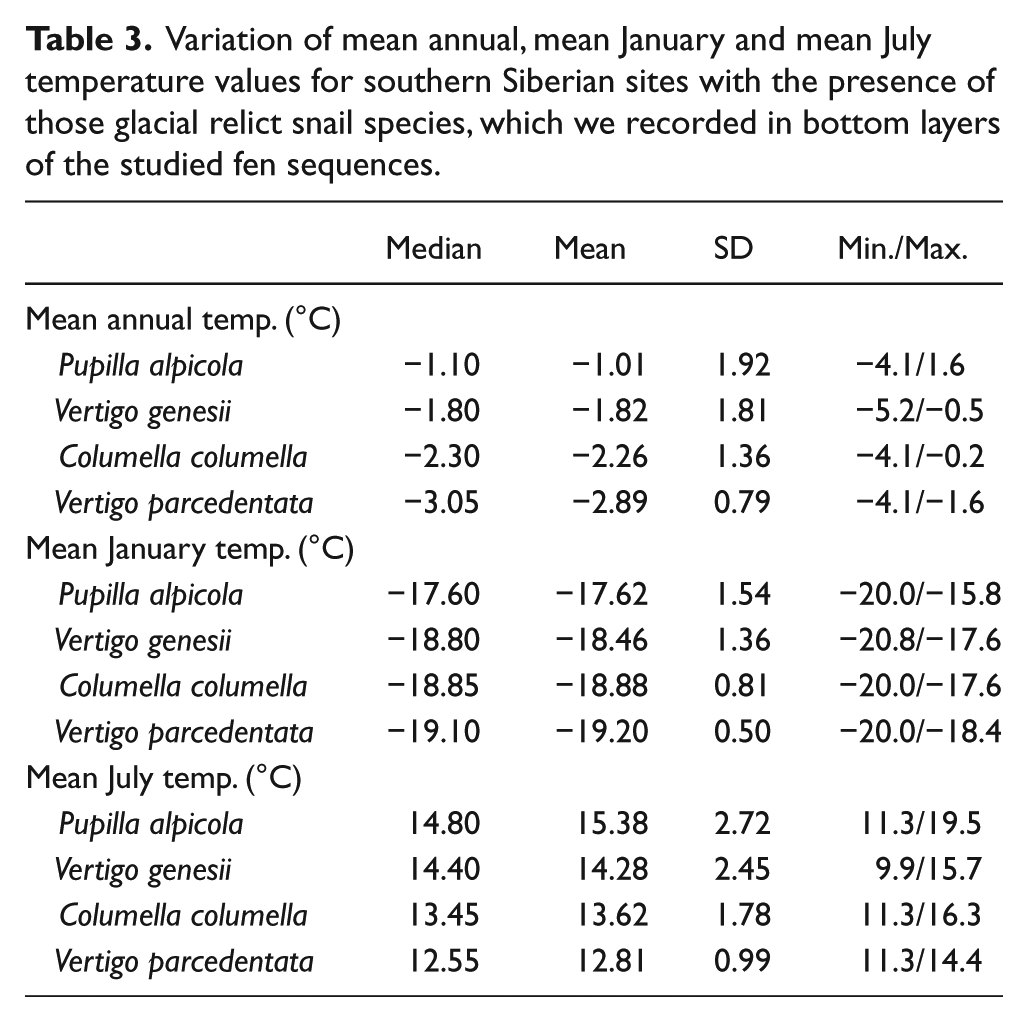
Using 66 extant Altaian populations of four characteristic Full Glacial snail species of temperate Europe, we reconstructed mean annual, January and July temperatures for the studied fens at the Pleistocene/Holocene transition (Table 3). Although there were differences among the optima of particular species, the main variation was found between preferences of Pupilla alpicola (a species that survived in fens of the study area to recent times) and three other species that became extinct approximately at the transition between early and middle Holocene. Southern Siberian populations of these three latter species expressed very consistent temperature preferences, with the mean annual temperature varying between −3.07 and −1.15°C. P. alpicola preferred climatically milder sites, with the optimum temperature of 1.95°C.
Variation of mean annual, mean January and mean July temperature values for southern Siberian sites with the presence of those glacial relict snail species, which we recorded in bottom layers of the studied fen sequences.
Also, habitat features of the analysed sites in the Altai refugium can provide further important data for reconstructions of conditions at the study fens. While half of these Siberian sites represented typically treeless fens and fen meadows (17 sites), the other half consisted of fen shrubs (10 sites) and sparse fen woodlands (7 sites). These shrubby and woodland fens were even more frequent habitat types when looking only at sites with the presence of more than two of these relict species. According to the fossil samples from the analysed profiles, the glacial snails were living in all three of these contrasting microhabitats – in shrubby fens (HOZ and VAL), a treeless calcareous fen (BRE-box) and also fen woodlands (BRE-A1 and VAL). Simultaneous presence of fossils of light-demanding fen plants suggested the existence of open patches in shrubby and woodland vegetation.
Discussion
In this study, we provide evidence that the application of the relict concept to explain recent distributional, metacommunity and diversity patterns of temperate fens is meaningful. We have clearly documented that the whole-Holocene local continuity of a calcareous fen is possible, and that populations of specialised fen species can persist locally for millennia.
Long-term continuity of the habitat
Low-productive calcareous fens are dynamic ecosystems whose character is determined by multiple factors including water chemistry, macroclimate (affecting the hydrological regime), autogenic succession and disturbances, and nutrient enrichment by wild animals or man. Stratigraphical hiatuses and reversal developments may frequently occur in fen sediments (Ammann et al., 2013; Hájková et al., 2012a). Such high dynamics are probably the reason for poor palaeoecological evidence of continual in situ persistence of calcareous fens and their specialised plants.
Grootjans et al. (2006) reviewed the knowledge on the long-term stability of calcareous fens and similar habitats. The longest stability they reviewed concerned percolation mires in Germany spanning about 3000 years (3000–6000 BP; Michaelis, 2002). Our results from the Hozelec site provide evidence of much longer in situ stability of open-fen patches, from the late Glacial to present times. We thus confirmed the hypothesis that a great recent concentration of relict fen species is a result of a long, continual Holocene history of calcareous fen vegetation (Hájek et al., 2011). However, this hypothesis could not be verified based on a single proxy, because of some hiatuses in the fossil record, and only the combination of all three independent proxies (pollen, macrofossils and snails) provided unequivocal evidence. Pollen documented unspecified treeless wet habitats during the late Glacial, while macrofossils documented calcareous fen vegetation around the late Glacial/early Holocene transition and in the late Holocene, and specialised snail species indicated the continuous existence of calcareous fen habitats since the early/middle Holocene transition.
The mollusc succession at the Hozelec site showed the lowest level of compositional turnover and species richness changes throughout the whole profile (Appendix S3, available online). Fossil snail records, in which more complete assemblages compared with plants are usually preserved, hence appear to be crucial for completing the evidence of long-term fen stability. The reason for exceptional conservatism of some fens in the Western Carpathians, such as the Hozelec fen, probably lies in the specific hydrological systems with deep circulation of extremely mineral-rich groundwater (Hájková et al., 2012a; Madaras et al., 2012) that alleviate macroclimatic fluctuations and may block or even reverse autogenic succession (Hájková et al., 2012a, 2013).
Sequences from the two other fens did not yield such an unequivocal evidence of long-term local presence of calcareous fen habitat or particular species. During the Holocene, these fens showed extensive changes in species composition and richness of snails, as well as plants. We expected the long-term persistence of open-fen patches at the Brezové fen, where a high number of relict fen species occur at present (Hájek et al., 2011). In all cores taken from this fen, however, the forested phase was virtually without any light-demanding species (see Figures 4 and 5, Appendix S4, available online). The most critical bottleneck for survival of light-demanding species started after about 4400 cal. yr BP. The only sign of possible persistence of small open-fen patches was the presence of oogonia of Chara vulgaris that indicated the presence of small calcareous pools, with the open-fen species probably surviving on their shores. Our results confirm those of Madaras et al. (2012), who drilled a series of cores throughout the Brezové fen (not analysed by palaeoecological methods) and reported on the existence of a layer of compact, very decomposed dark peat with wood remains that completely overlay basal carbonate-rich peat. They concluded that the site was forested in the past. It is impossible to determine from our data whether the fen species that were found in the bottom layers and then after the late Holocene deforestation have persisted locally in small populations in a few open patches, or whether they re-colonised the site by long-distance dispersal.
The third site, Valalská voda, was probably forested densely for a long time, as indicated by both the long stratigraphical hiatus and the poor representation of relict and specialised fen species in the modern assemblages. The few relict species that occur today at this site had to re-colonise or colonise the fen de novo after the Wallachian colonisation. For example, shells of Vertigo geyeri, the recently occurring relict snail, were found only in the youngest layers of this profile.
Persistence and extinction of fen species
One of the aims of this study was to test the hypothesis about whole-Holocene local survival of the species whose recent distributional patterns suggest their relictualism from late Glacial times with direct fossil evidence. So far, the evaluation of species relictualism has been mostly based on indirect evidence such as phylogeography of modern populations (Ehrich et al., 2008; Lambracht et al., 2007), specific recent distribution (Dítě et al., 2013), wider distribution of a species in the past (Jankovská and Rybníček, 1988; Magyari et al., 1999), and correlations between the fen age and fen specialists (Hájek et al., 2011). In contrast, continual occurrences in peat profiles have been poorly documented.
We were successful in verifying whole-Holocene persistence of an open calcareous fen (at the Hozelec site, Figure 2), but we failed to confirm such continuity at the level of individual species. A high water level at the Hozelec site during the middle Holocene probably enabled the existence of treeless patches, but unfortunately caused a hiatus in the macrofossil record. The longest local continuity we found thus concerns the late Glacial relict snail species Vertigo geyeri, which undoubtedly occurred continually over more than 8000 years at this site. However, this species was not recorded in the late Glacial samples in any profile studied, which can be explained by its intolerance to extremely harsh climates. Recently, it was found to avoid sites at high altitudes and latitudes where it is replaced by Vertigo genesii. These two late Glacial sibling Vertigo species rarely co-occur at one site even in the Alps and Scandinavia where they are frequent today (Schenková et al., in press). A comparatively long in situ persistence was further found for other open-country yet less tightly fen-associated snail species such as Vallonia pulchella, Vertigo angustior and Nesovitrea hammonis (Appendix S3, available online).
We found a set of calcareous plant species that occurred in both the old and the young deposits (e.g. Carex flava agg., C. nigra, Bryum pseudotriquetrum, Campylium stellatum, Juncus articulatus and Scorpidium cossonii; see Appendix S4, available online), but they are rather common today in the Western Carpathians and are therefore not generally considered to be relicts from the cold past. In addition, they did not show a statistically significant affinity to ancient fens in the study of Hájek et al. (2011). Nevertheless, the ancient fens could act as a source of these species after the Wallachian colonisation. Most of the species with clear relict distributional pattern (see Hájek et al., 2011) were not found in our profiles. It is difficult to determine whether their absence from fen deposits was caused by taphonomic reasons (e.g. transient seed bank and seed predation in the case of Primula farinosa) or whether they truly did not occur in the late Glacial because of different environmental conditions. The environmental changes from the late Glacial to recent times evidently caused certain species exchange, as evidenced by the late Glacial and early Holocene occurrence of fen species that are recently absent in the study fens such as Trichophorum pumilum (BRE-A1), Potamogeton filiformis (HOZ, BRE-box), or boreal bryophyte species Meesia triquetra and Scorpidium scorpioides (BRE-box). These species are very rare today in Central Europe and they are traditionally considered to be relicts from the cold phases (Dítě and Šoltés, 2010; Rybníček, 1966).
Trichophorum pumilum is common in continental Asia, showing a more compact European distribution range during the Full Glacial as well as late Glacial and surviving recently in travertine fens in the Western Carpathians (Dítě et al., 2013). Meesia triquetra and Scorpidium scorpioides are moss species that are recently very rare and endangered across Central Europe (Kučera et al., 2012), but they are common in various mire habitats in boreal and arctic Eurasia (Hedenäs, 2003). During the late Glacial and early Holocene, they had a broader distribution in Central Europe (Dobrowolski et al., 2012; Hájková et al., 2012c; Jankovská, 1970; Magyari et al., 1999; Rybníček and Rybníčková, 1968). Potamogeton filiformis is regularly present in cold-stage sediments (e.g. Gałka et al., 2014; Stachowicz-Rybka et al., 2009; Wohlfarth et al., 2006) and today, it occurs widely in Northern Europe and in the Alps (e.g. Lauber and Wagner, 2012; Rørslett, 1991).
Based on these findings, we can state that the late-Glacial assemblages are at least partially analogous to the present-day assemblages known from boreal and continental landscapes, at least in terms of the presence of these boreal and continental elements, which are extremely rare in the Western Carpathians today. However, it is unfortunately impossible to reconstruct the complete species assemblages using macrofossils, because of taphonomic constraints. We therefore cannot exclude that these boreal and continental species co-existed with temperate fen species such as Carex davalliana, C. dioica or Primula farinosa, the latter being found in pollen in two late Glacial/early Holocene deposits at lower altitudes in the Western Carpathians (Hájková et al., 2013; Jamrichová, unpublished).
Possible causes of snail extinctions in temperate Europe
The land-snail species characteristic of the European Full Glacial period, which we surprisingly discovered in basal samples of all studied profiles, enabled us to trace the history of their populations and Holocene extinction events in the Western Carpathians. Generally, these species became extinct across the whole of temperate Europe around the Pleistocene/Holocene transition (Horsák et al., 2010; Ložek, 1964, 2001), but some of them retreated to high mountain refugia in the Alps and Carpathians (Kerney et al., 1983; Schenková and Horsák, 2013). For Columella columella, a shift in habitat preferences was documented at the end of the Glacial, when this originally xerophilous and characteristic loess-dwelling snail species (Ložek, 1964) established large populations in calcareous wetlands (Ložek, 2007 and our results). In recent times, it commonly inhabits fens in cold regions of Norway (Pokryszko, 1993) and the Alps (Kerney et al., 1983). Our records suggest that during the late-Glacial climate oscillations, calcareous fens could act as refugia also for other characteristic species of loess snail fauna such as Vertigo parcedentata. Some of these species have managed to survive in calcareous fens up to recent times (e.g. Pupilla alpicola), but most of them became extinct during the early Holocene (Ložek, 1964 and our results) across the entire temperate Europe, except for some high mountain regions. The reasons for these extinctions remain unclear and only poorly resolved. At least three possible causes can be suggested: (1) the pure ecophysiological effect of climate change, (2) habitat change and (3) biotic interactions. As the level of interspecific competition among land snails is believed to be low or negligible (e.g. Hylander et al., 2005; Waldén, 1981), which seems to be true particularly for minute fen species (e.g. Horsák, 2006; Horsák et al., 2014), we did not consider competition as the leading force of these extinctions. To disentangle the pure effects of climate and habitat change is more challenging because the postglacial habitat changes are driven mainly by the changing macroclimate (e.g. Williams and Shuman, 2013).
Our fossil evidence suggests rather dynamic habitat conditions at the study fens during the Holocene with respect to the water regime and tree canopy. The question is whether these snail species, adapted to a glacial landscape, could cope with these changes. Analogous assemblages of these snails from their continental refugium in the Altai Mountains in southern Siberia (Horsák et al., 2010; Meng, 2009) may bring a possible answer to this question. In the Altai Mountains, these species dwell in alkaline open-woodland fens (Horsák et al., 2010), most likely because scattered trees and shrubs provide suitable shelter against the harsh climate during the coldest months. Structurally, these sites clearly resemble those we reconstructed based on our records from the studied fen sequences. If these species can survive in forested fens, the pure effect of climate changes might be more important than subsequent changes in habitat structure towards woodland stages.
The crucial role of climate change in extinction of glacial snails in temperate Europe is further supported by the fact that only Pupilla alpicola survived largely in the Western Carpathian fens to recent times. Modern climatic demands of refugial populations of glacial snail species in the Altai Mountains indeed showed that P. alpicola displayed the highest heat tolerance and even certain preference for a warmer macroclimate (Table 3). Only this snail thus managed to survive all habitat and temperature fluctuations throughout the Holocene in the Western Carpathian fens. In contrast, Vertigo parcedentata, which became extinct throughout Europe except for a single small area in Norway (Pokryszko, 1993), showed preference for the coldest areas. The other two species, Vertigo genesii and Columella columella, were intermediate and indeed, they became extinct in the lowlands but survived in the high altitudes of the Alps and Carpathians.
Conclusion
In this study, we provide evidence that the application of the relict concept to explain recent distributional, metacommunity and diversity patterns of temperate fens is meaningful. Based on the data from the Hozelec fen in the Western Carpathians, we have clearly documented that the local continuity of a calcareous fen is possible for the whole Holocene, and that populations of specialised fen species persisted locally for millennia (maximum about 8000 years in this study). Moreover, because the habitat maintained stable abiotic conditions (calcium richness, light and high water table) for 12,000 years, we can suppose the possibility of even longer in situ survival of the relict fen species, but direct evidence for such a long time span is still missing. Nevertheless, fen habitats fed by abundant, mineral-rich groundwater from deep circulation appear to be good postglacial refugia, comparable to other stable habitats such as rocks, screes or alpine tundra. This finding is rather surprising, because fen habitats are known to be rather prone to natural successional changes. Indeed, for the other two fens for which we could anticipate similar continuity, considerable dynamics, including forested phases were found, and there was no clear evidence of long-term local survival.
Despite unequivocal evidence of whole-Holocene persistence of a calcareous fen habitat at least at one site, we generally found rather low similarity between modern and the oldest fen assemblages. The species typical of European glacial periods disappeared from the studied Carpathian fens during the middle Holocene, with the last record of recently extinct glacial snail species at about 8200 cal. yr BP (the Brezové fen), most probably because of climate change. Boreal bryophytes Meesia triquetra and Scorpidium scorpioides, which are recently rare in the Carpathians, disappeared from the same fen around 4400 cal. yr BP after canopy closing. The modern composition of both plant and mollusc fen communities seems to be largely a product of the late Holocene assembly in most of the Western Carpathian fens. The intense deforestation during the late Middle Ages (e.g. the Wallachian colonisation) has led to the development of calcareous fen assemblages that are rather similar across the Western Carpathians. In some old fens, however, the relicts from the late Glacial stages, persisting over the middle Holocene habitat bottleneck, enriched newly assembled communities and made these old fens inestimable and irreplaceable in terms of habitat and species conservation. The Hozelec and partially also the Brezové fen belong among these unique sites, which underlines the need to make their protection and conservation management a priority.
Footnotes
Acknowledgements
We are grateful to Ondřej Hájek for preparing the map, Stanislav Němejc and Jan Myšák for help with the profile sampling, and Zuzana Formánková for laboratory work.
Funding
Our palaeoecological research was supported by projects of the Czech Science Foundation P504/11/0429, institutional support of Masaryk University, and long-term research development project no. RVO 67985939. Research in the Altai Mountains was supported by the project P504/11/0454.
Supplementary Material
Additional information may be found in the online version of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.