
Abstract
Analyses of pollen, diatoms, XRF geochemistry, and pigments provide a unique window into how an insular ecosystem in Mauritius responded to an extreme drought event 4200 years ago. We provide a reconstruction of regional vegetation change and local wetland development under influence of sea level rise and inferred climate change between 4400 and 4100 cal. yr BP. Our multi-proxy data evidence a severe drought between 4190 and 4130 cal. yr BP, which ultimately led to mass mortality of larger vertebrates, including two species of giant tortoises and dodos in a <2-ha region. This prolonged drought around the Indian Ocean is recorded in many regions dependent on monsoon precipitation and is suggested to cause the collapse of human societies in East Africa and India. We demonstrate a direct relation between the mass mortality events in the Mare aux Songes (MAS) rock valley and the 4200 cal. yr BP drought. MAS represents a fresh water source that attracted and concentrated vertebrates. Abrupt increased aridity induced regional fires on Mauritius and caused decreased water levels, and a shrinking water surface resulting in further concentration of the animals in this coastal site. Upconing of the saline wedge underlying the fresh water source induced progressive salinization. The excrements of the animals produced hypertrophic conditions that, combined with salinization and high temperatures, created a suitable environment for potentially toxic cyanobacteria. These factors led to a deadly cocktail, resulting in the death of 100,000s of vertebrates by intoxication, dehydration, trampling, and miring, and promoted a unique conservation of fossils. The ‘4.2 ka megadrought’ likely induced similar bottlenecks elsewhere in the SW Indian Ocean region.
Keywords
Introduction
In 2005, a rich fossil bed was discovered at Mare aux Songes (MAS), a wetland in the southeast coastal lowlands of the oceanic island Mauritius (Rijsdijk et al., 2009). The fossil depository constitutes a Lagerstätten with more than 250 bone fragments/m2 with a diverse spectrum of plants and animals, including the extinct dodo (Raphus cucullatus) and extinct giant tortoises (Cylindraspis spp.). Radiocarbon dating of vertebrate bones, wood, and seeds indicates that fossils accumulated between 4235 and 4100 cal. yr BP (calibrated years before AD 1950). This suggests that hundred thousands of vertebrate individuals died within a time frame of less than 150 years (Rijsdijk et al., 2011), long before humans colonized the island in AD 1638.
The discovery of this fossil bed is important to improve our understanding of life boundary conditions on apparent paradisiac and climatically stable islands (Van der Plas et al., 2012). At a global scale, few records describe how vertebrate faunas have been affected by Holocene climatic extremes, and/or how such faunas have been able to withstand climatic variations on small islands (Rijsdijk et al., 2011). The unique setting at MAS provides an unprecedented opportunity to examine in detail the impact of a natural catastrophe on a rich insular ecosystem. It is still unclear under what conditions this great number of animals died within a wetland of a few hectares and what the ultimate cause was for this period of mass mortality.
Death scenarios were explored from dodo bone assemblages and hydrological data from the MAS rock valley (Meijer et al., 2012), and miring was considered among the most likely scenarios whereby animals were trapped in the sediment during repeated events. Rijsdijk et al. (2011) correlated the mass mortality with an extreme drought dated at around 4200 cal. yr BP in East Africa (e.g. Gasse, 2000; Marchant and Hooghiemstra, 2004; Thompson et al., 2002), North Africa (e.g. Williams, 2009), and India (e.g. Fleitmann et al., 2007; MacDonald, 2011; Staubwasser and Weiss, 2006). However, no direct link could be made with environmental changes in Mauritius, as paleoclimatic reconstructions from the Mascarenes were lacking (Rijsdijk et al., 2011).
A study by De Boer et al. (2014) documents a Holocene climatic reconstruction of the SW Indian Ocean from the Tatos wetland in the eastern lowlands of Mauritius. This study records a prolonged drought centered at 4300 cal. yr BP, representing the driest Holocene conditions in Mauritius. Because sediments in the MAS rock valley only cover the time period of roughly 4500–4000 cal. yr BP (Rijsdijk et al., 2011), the study by De Boer et al. (2014) provides a framework along which an environmental record from MAS can be assessed.
In this paper, we will reconstruct the development of the MAS wetland, and we will provide an environmental context that helps to understand the sequence of events that ultimately facilitated the formation of this rich fossil bed. Our main questions are as follows: (1) what were the environmental conditions that preceded and prevailed during the mass mortality events; (2) what factors induced mass mortality and what was the role of extreme drought? and (3) why is the evidence for vertebrate mass mortality exclusively documented at MAS and not elsewhere on other islands?
Setting
Geology and hydrology
Mauritius is a volcanic island located ~900 km east of Madagascar in the SW Indian Ocean (Figure 1). The island has a surface of 1865 km2, with several peaks reaching up to 828-m elevation and a central flat upland area between 500- and 650-m elevation that slopes gently toward the lowlands at the northern and eastern coastlines.

(a) Map showing the location of Mauritius in the Indian Ocean. (b) Map of the island indicating the approximate coastline during the ‘Last Glacial Maximum’ (−115 m), altitude lines (200-m steps), the distribution of main vegetation types before colonization (adapted from Cheke and Hume, 2008), and the extent of palm woodland according to our results (this paper). Lower montane forest includes mid-altitude forest, moist forest, wet forest, and upland vegetation types (e.g. heath). Coastal forest, mangroves, and wetlands are not shown. White dots indicate study areas discussed in the text: 1De Boer et al. (2014), 2Van der Plas et al. (2012), 3De Boer et al. (2013b), 4De Boer et al. (2013a). (c) Map of southeast Mauritius and the location of Mare aux Songes (MAS).
MAS (20°26′51.2″S; 57°41′23.5″E) is a 10-m-deep elongated basalt rock valley, formed after a phreatomagmatic explosion that occurred between 100,000 and 20,000 years ago (McDougall and Chamalaun, 1969; Figure 1c). The young basalts in the eastern lowlands have high porosity and water infiltration capacity because of poor soil development (Saddul, 2002); thus most of the precipitation, from highlands to coastal areas, infiltrates into subsoil, recharging the basaltic aquifers through which groundwater flows continuously toward the ocean. Groundwater from these aquifers reaches the surface at geological unconformities and at low-lying coastal rock valleys where the ocean hinders further transport of fresh groundwater. MAS is one of the most important coastal rock valleys where groundwater emerges, facilitating a constant supply of freshwater to this wetland (De Boer et al., 2014; Rijsdijk et al., 2011). These types of coastal wetlands, such as MAS, are often the only place in permeable basaltic areas where standing water is found. This water may be fresh, brackish, or saline depending on the mixing of the fresh groundwater with saline groundwater, originating from seawater intrusion from the nearby ocean. The observed increasing salinity of the groundwater of the sub-basins of MAS evidences that MAS is connected to the ocean (Rijsdijk et al., 2011). The measured tidal groundwater table fluctuates 100–700 mm within the four sub-basins, which also indicates that the sub-basins are connected with the ocean and with each other (Rijsdijk et al., 2009).
MAS is located less than 1 km from the ocean and consists of four sub-basins that are separated by rock thresholds (Hume et al., 2014). The present surface of these sub-basins is situated between mean sea level and 0.5 m above mean sea level. The N and S sides of the sub-basins are surrounded by 10-m-high bedrock with steep talus slopes (>60°). Gentler slopes are present on the E and W sides. Within the sub-basins, groundwater level is just below the present surface and the water is alkaline, with a pH ranging from 7.1 to 7.8 (Rijsdijk et al., 2009).
MAS sub-basin 1 is filled in with a sedimentary sequence. An up to 6-m-thick unit of coral sand is overlain by an up to a meter thick calcareous lake marl. The laminated lake marl is overlain by an up to 80-cm-thick fossil-rich peat. The basal 30 cm of this peat contains tree stems (<25 cm) and tree roots (5–15 cm). This grades into a 50-cm-thick bone-rich peat which in the top is rich in Tambalacoque (Sideroxylon grandiflorum) and Pandanus seeds. The sequence is capped by 30 cm of compressed peat and a layer of sedges. Thin sand laminae (from a few millimeters up to 1 cm) are interspersed in the organogenic sequence.
Climate
Precipitation in the western Indian Ocean is mainly controlled by the African and Indian monsoon systems, which can be separated into northern and southern hemisphere components. The southwest monsoon, which is divided into the Indian Summer Monsoon (ISM) and Asian Summer Monsoon (ASM), prevails during the boreal summer, whereas the northeast monsoon prevails during the austral summer (Van Rampelbergh et al., 2013). Mauritius receives most of its precipitation from November to April brought by the northeast monsoon. The lowlands experience a drier season from May to October under the influence of cool and dry easterly trade winds (Senapathi et al., 2010; Vaughan and Wiehe, 1937). Mean annual precipitation (MAP) depends on altitude and orientation of the slopes with respect to the prevailing wind direction. The western coastal lowlands are relatively dry with an MAP of 800 mm (Padya, 1989). The MAP is about 1200 mm in the eastern coastal lowlands, and increases to more than 4000 mm at the elevated central part of the island. The mean annual evapotranspiration (MAE) in the coastal regions (<50-m elevation) exceeds MAP. MAS is located at the windward position of the eastern coast, where the MAE is ~2200 mm and the MAP ~1300 mm resulting in a water deficit most of the year (Padya, 1989).
Coastal-lowland vegetation
At present-day, about 95% of Mauritius has been deforested and an outline of the natural vegetation distribution must therefore rely on early historical records and small remnants of degraded natural vegetation. The pristine island was fringed by a variety of coastal vegetation communities such as mangroves, coastal marshes, coastal forest and vegetation associations characteristic of basaltic cliffs and coralline sand dunes (Cheke and Hume, 2008). On the driest parts of the island (MAP < 1000 mm), palm woodland occurred behind a strip of coastal vegetation (Cheke and Hume, 2008). In lowland areas where MAP ranges between 1000 and 1500 mm, semi-dry forest, including ebony (Diospyros) forest occurred further inland behind the palm woodland or coastal vegetation (Cheke and Hume, 2008; Vaughan and Wiehe, 1937). Semi-dry forest was the dominant biome in the lowlands and covered over 40% of the island up to the start of human colonization in AD 1638 (Cheke and Hume, 2008). The island became rapidly deforested (Vaughan and Wiehe, 1937); the eastern lowland forests were largely destroyed in the 19th century. By AD 1872, palm-rich woodland had disappeared from the Mauritius main island (Cheke and Hume, 2008; Safford, 1997). The coastal lowlands around the MAS rock valley was transformed into sugarcane fields. The sub-basins of MAS itself proved unsuitable for agricultural practices and host remnants of semi-dry forest and wetland vegetation.
Materials and methods
Core description and chronology
The core described in this study was taken from Trench 4-2007, located in the center of sub-basin 1 (Hume et al., 2014; Rijsdijk et al., 2009, 2011). This trench was opened and excavated between 2007 and 2010, and the upper ~30 cm sampled for bones and other macrofossils. In August 2010, a 129-cm-long sediment core (MAS1-Tr4-C3; from here on labeled MAS1) was retrieved using a Russian Corer (Ø 50 mm) in 50-cm increments until the coral sand basement was reached at 225 cm below the water surface. Visual core description obtained directly after recovery revealed a gradual transition from coral sand to slightly laminated lake marl around 220-cm depth. At 137-cm depth, the marl deposition changes abruptly into peat. The top of the sediment column was at 96 cm below the water surface.
The age model of the sediment core is based on six accelerator mass spectrometry (AMS) radiocarbon ages from seeds of Pandanus screw pine (~1 cm3) and Latania palm (~0.5 cm3). These seeds originate from former vegetation in the direct vicinity of the basin and are likely not to have been transported or reworked. The southern hemisphere calibration curve (McCormac et al., 2004) was used to convert radiocarbon ages into calibrated calendar ages. The age calibration and the construction of an age–depth model was performed with the Bacon program (Blaauw and Christen, 2011). The Bacon program performs a Bayesian age–depth modeling that includes the dating probability distribution and excludes negative sedimentation rates.
To reconstruct the development of the MAS wetland we used an integrated multi-proxy approach. Pollen analysis was used to reconstruct regional vegetation change, while macrofossil analysis provided information on the local vegetation composition. Diatom analysis was used to reconstruct changes in the water conditions. The presence of microscopic charcoal indicates the occurrence of fire, while coprophilous fungi such as Sporormiella indicate the presence of larger vertebrates. XRF analysis provided details on the composition and origin of the sediment. Fossil pigment analysis was used to detect changes in the abundance of algae and cyanobacteria.
Pollen and diatom analysis
The MAS core was sampled every 4 cm for pollen and diatom analysis by extracting 1-cm-thick slices between 220- and 96-cm core depth. Diagrams were plotted with TILIA 1.5.12 (Grimm, 1993, 2004) software. Zonation was determined by CONISS cluster analysis, included in the TILIA program.
Pollen samples (~0.8 cm3) were prepared using standard pretreatment techniques including sodium pyrophosphate, acetolysis, and heavy liquid separation with a bromoform–ethanol mixture, specific gravity 2 (Faegri and Iversen, 1989). Additional details on sample preparation and pollen identification are described in Van der Plas et al. (2012). Pollen identifications in this study were predominantly based on modern pollen samples collected by E J de Boer, C Baider, and F B V Florens. Microscopic charcoal was classified into the size classes of 15–60 and 60–200 µm.
Approximately 0.8 cm3 of each sample was prepared for diatom analysis. Samples were immersed in 30 mL of H2O2 (30%) for 30 min at room temperature, after which a few drops of potassium permanganate were added. After the reaction stopped, 10 mL of HCl was added. Samples were then washed with distilled water, mounted in Naphrax on permanent slides, and analyzed with an Olympus microscope at 1000 magnification. A minimum of 400 valves was counted per sample (except for the sample at 140-cm depth for which 350 diatoms were counted). Diatom taxonomy and ecology were identified after Witkowski et al. (2000), Krammer and Lange-Bertalot (1991), Potapova (2009), and Morales et al. (2010).
Macrofossil analysis
During several fieldwork campaigns between 2005 and 2011, ~500 pieces of waterlogged wood – stems, branches, twigs, and roots – were sampled from the fossil bed in MAS (mainly trench 4), of which 350 pieces were selected for further analysis. In addition, seeds, fruits, and other plant parts were also collected. A reference collection of Mascarene dicot wood was created by sampling endemic, native, and pan-tropical species from local reserves, as well as from collections in Mauritius and Europe (Mauritius Herbarium; the Herbarium of Naturalis Biodiversity Center in Leiden; Museum for Central Africa, Tervuren, Belgium; Royal Botanic Gardens, Kew, UK). Transverse, tangential, and radial sections were prepared and analyzed light microscopically for scoring wood-anatomical features using the IAWA Hardwood List (Wheeler et al., 1989). Macrofossils were identified using the InsideWood (2004-onwards) Internet database, modern reference slides, and wood-anatomical publications of all genera occurring naturally in Mauritius.
XRF analysis
The bulk geochemical sediment composition was acquired nondestructively at the core surface using the Avaatech X-Ray Fluorescence (XRF) core scanner at the NIOZ (Texel, the Netherlands; Richter et al., 2006). XRF scanning was performed after the sampling for pollen and diatom analysis, resulting in a sampling resolution of 0.5 cm where no pollen and diatom samples were obtained. The measurements were conducted at 10 kV for 10 s, and at 30 kV for 20 s in combination with a Pd-tick filter, after the core surface was carefully cleaned and covered with a SPEXCerti Ultralene foil (Tjallingii et al., 2007). These two settings provided data of 13 elements (Al, Br, Ca, Cl, Fe, K, Mn, Rb, S, Si, Ti, Zn, and Zr). Elements that reveal the main compositional variations were selected as ratio pairs of element intensities. Relative compositional variations are presented as log ratios of element intensities, which are linearly related to log ratios of element concentrations (Weltje and Tjallingii, 2008).
Fossil pigment analysis
The analysis of sedimentary pigments was modified from the method described by Van der Staay et al. (1992). Pigments from approximately 100 mg of freeze-dried and homogenized sediment were extracted in a solution of 80% acetone, 15% methanol, and 5% MilliQ water after grinding the sediment in a Mini BeadBeater (BioSpec products) with 0.5-mm silica beads and incubation at 4°C for 12 h. After addition of an ion-pair mix (1.5% tributyl ammonium acetate and 7.7% ammonium acetate), the samples were mixed, centrifuged, and the supernatants were stored at −20°C until HPLC analysis. Pigments were separated on a Thermo Scientific ODS Hypersil column (250 mm × 3 mm, particle size 5 µm) in a Shimadzu HPLC and using a KONTRON SPD-M2OA diode array detector. The different pigments were identified based on their retention time and absorption spectrum and quantified by means of pigment standards.
Results
Chronology and lithology
The radiocarbon-based chronology of the sediment core is based on five selected macro-seeds (Table 1; Figure 2). The top-most sample at 102.5-cm core depth was not used, as its 14C-age of 1960 ± 35 yr BP, was beyond the expected range and plausibly contaminated with younger material. The presence of a stone from 96- to 100-cm depth also indicates that the top of the sediment sequence should be treated with caution. This stone results from the fill in of the marsh in the 1940s with dolerite blocks and gravels to eradicate malaria. Although the dolerite blocks and gravel affected mainly the top layers of the MAS-sediment sequence (Figure 3), some smaller stones were able to penetrate deeper into the sediment. Evidence that the dumping did not affect the layers below the fossil bone bed comes from the non-deformed lake marl and the preservation of sharp stratigraphical contacts between units (Rijsdijk et al., 2009, 2011).
List of radiocarbon dates and sample-specific data of the sediment core MAS1.


Age model of core MAS1 and identified periods, based on lithology, diatom and pollen zones. Upper panels indicate stability Markov chain Monte Carlo run, accumulation rate (green prior distribution, gray posterior), and memory (green prior distribution, gray posterior). Gray scales in lower panel indicate chronological uncertainty; darker gray indicates more likely ages. Dark blue shapes are the calibrated 14C dates; blue shape indicates the calendar date of the surface. Ages of the zone and period boundaries are taken from the weighted mean values.

Location of sediment core MAS1-Tr4-3 (MAS1) on a schematic side view of sub-basin 1 and a complete sedimentological sequence from the center of the basin (following Rijsdijk et al., 2009). Trench 4 was excavated in 2007, 2009, and 2010, during which the upper layers of the sediment sequence were removed until the top of the fossil bone bed was exposed. Subsequently, the sediment core MAS1 was collected in August 2010.
The calibrated ages show a large overlap because of a plateau in the calibration curve around ~4000 yr BP and the short distance between the samples. However, because of the clear stratigraphical sequence of the sediment core and rapid transitions in the pollen and diatom records, we assume a continuous sedimentation for the age-model reconstruction. The Bacon-based analysis indicates that the MAS-record covers the time span from 4370 cal. yr BP at 220-cm depth to 4110 cal. yr BP at 96-cm depth (Figure 2).
Based on the lithology, sediment composition, and the pollen and diatom zones, we recognized in the MAS1 record four periods (Figure 2). Periods MAS-1 (220- to 198-cm depth; 4370–4320 cal. yr BP) and MAS-2 (198- to 134-cm depth; 4320–4190 cal. yr BP) are contained within the lake marl section, while periods MAS-3 (134- to 106-cm depth; 4190–4130 cal. yr BP) and MAS-4 (106- to 96-cm depth; 4130–4110 cal. yr BP) are contained in the peat sediments. The top of the MAS1-core corresponds to the center of the fossil bone bed layer (Figure 3).
Pollen, wood, and macrofossils
Pollen preservation was poor in the calcareous marl sediments between 220- and 140-cm core depth, but better in the dark organic peat layers between 136- and 96-cm core depth. Therefore, pollen spectra are only shown for the upper core pollen samples; abundances below 140-cm core depth are shown as counts (Figure 4a). CONISS identified zone boundaries at 114-cm (4150 cal. yr BP) and 106-cm core depth (4130 cal. yr BP).

Pollen and diatom records, XRF geochemistry, and pigment occurrence, from sediment core MAS1 against depth. Individual pollen and diatom records show down-core changing proportions (%) related to the sum of pollen or diatoms; bar diagrams show counts. The pollen record (a) shows from left to right the identified periods and pollen zones, pollen concentration values (pollen/cm3 of sediment), and pollen sum values, pollen taxa arranged according to their ecological affinity (taxa with less than two occurrences are not shown), fern and fungal spores, charcoal, and the CONISS cluster dendrogram. The diatom record (b) shows from left to right the identified periods and diatom zones, the pigments zeaxanthin and diatoxanthin relative to β,β-carotene, diatom taxa arranged according to their preference for water salinity (taxa with less than two occurrences are not shown), unidentified diatoms, freshwater diatom spores, and the CONISS cluster dendrogram. Indicator values of the degree of organic pollution are given below the relevant taxa.
Pollen taxa reflect palm woodland, semi-dry lowland forest, and wetland vegetation (Figure 4a). Palm woodland is mainly represented by Latania, Dictyosperma, Acanthophoenix, and Pandanus (see also De Boer et al., 2014). Semi-dry forest is characterized by canopy trees Ficus, Eugenia, Sapotaceae spp., Terminalia bentzoë, Diospyros spp., Tabernaemontana, and Cassine orientalis; small trees of Gardenia type, Ixora, Zanthoxylum, Antidesma, Cordia/Hilsenbergia type, and Foetidia; and shrubs of Dodonaea and Dombeya spp. The palm genera Latania, Dicyosperma, and Acanthophoenix also occur in semi-dry forest, emerging several meters above the canopy (Vaughan and Wiehe, 1937). Although MAS is situated 1 km from the current coastline, no coastal vegetation elements are recorded.
The list of 28 identified wood taxa shows a large overlap with the pollen taxa (Table S1, available online). However, the wood samples are dominated by semi-dry forest taxa, while the pollen spectra have a better representation of palm woodland taxa. This indicates that the local standing vegetation was dominated by semi-dry forest taxa, while palm woodland pollen was mainly supplied by trade winds blowing land inwards.
Species of palm woodland are predominantly found in environments with an MAP of <1000 mm; semi-dry forest species are predominantly found in environments with MAP of 1000–1500 mm. Palm woodland of the driest areas of Mauritius are characterized by Latania and Dictyosperma palms, whereas Eugenia and Ixora are most common for the more humid semi-dry forest. All species are significantly present throughout the record. Therefore, log ratios of Latania + Dictyosperma and Eugenia + Ixora pollen counts were used to infer climate conditions.
So far, few reliable identifications could be made for the fossil seeds, fruits, and other plant parts because of the lack of a hands-on reference collection of Mauritian/Mascarene dried seeds and fruits. About 50–60 morphotypes of seeds could be distinguished. The most conspicuous, larger seeds and stones (>1 cm) were identified at an earlier stage (see Rijsdijk et al., 2009). However, these amount to not more than 20 taxa of dicot and monocot trees, leaving out all herbaceous species that are found in the finer sieve fractions (<4 mm).
Diatom record and hydrology
In the diatom record, we recognized six zones with boundaries at 198-cm (4320 cal. yr BP), 154-cm (4230 cal. yr BP), 134-cm (4190 cal. yr BP), 126-cm (4170 cal. yr BP) and 118-cm core depth (4150 cal. yr BP; Figure 4b). The short time intervals per zone reflect rapidly changing water conditions mainly determined by organic pollution, depth of the water column, and salinity. Overall, the record is dominated by species tolerant to moderate to high organic pollution, alkaliphilous and freshwater but saline tolerant diatoms. However, the co-existence of species that inhabit brackish, saline, and freshwater water bodies indicates that MAS sub-basin 1 experienced frequent changes in salinity levels, evidencing the constant influence of fresh groundwater and brackish–saline groundwater.
Variations in water salinity throughout time are the result of the mixing of different amounts of fresh groundwater and saline groundwater which reflect meteorological variations in different ways: first, dry periods with high evapotranspiration rates lead regionally to lower water tables, and therefore decreased inflow of regional fresh groundwater into the wetland. Also, the wetland receives less rainfall during droughts and the freshwater in the wetland evaporates. Both lead to lower inputs of freshwater and therefore decreased dilution of saline groundwater, resulting in higher water salinities. Lower water tables in the wetland and reduced freshwater by evapotranspiration result in saltwater upconing (rising of the freshwater–saltwater interface) and, as a consequence, increased inflow of saline groundwater into the wetland. Therefore, higher water salinities indicated by salinity tolerant and marine diatom communities indicate drier periods.
Sediment composition and pigment analysis
Environmental reconstructions of the Tatos wetland based on comparing hydrological data and XRF core scanner measurements revealed that sediment compositions were linked to hydrological conditions. The hydrological conditions of the Tatos wetland, which is located in the northeastern coastal lowlands of Mauritius, are similar to the hydrology at MAS. Sediments at Tatos consist of detrital sediments, biogenically produced calcium carbonate (CaCO3), and organic matter that is mostly characterized by peat deposits (De Boer et al., 2014). Relative variations of detrital sediments are best indicated by the element Ti (Titanium), whereas the element Ca (Calcium) is associated with biogenic CaCO3. Therefore, relative variations of biogenic CaCO3 and detrital compositions can be indicated with ln(Ca/Ti) values.
Marine organic matter can be differentiated from terrestrial organic matter by elevated concentrations of the element Br (Bromine) and can be used to trace marine versus terrestrial organic carbon (Mayer et al., 2007; Ziegler et al., 2008). Br concentration will increase when saline groundwater infiltrates the MAS basin and the production of organic matter in the basin shifts from terrestrial to marine. Accordingly, relative saline water influence at the MAS basin can be indicated by ln(Br/Ti) values (De Boer et al., 2014).
Values of ln(Ca/Ti) are high around 180-cm core depth (4290 cal. yr BP), but rapidly decrease and reach minimum values around 160-cm core depth (4240 cal. yr BP). Subsequently, ln(Ca/Ti) values gradually rise. Values of ln(Br/Ti) are relatively stable until 160-cm core depth (4240 cal. yr BP), after which values rise exponentially; maximum ratios are reached around 117-cm core depth (4150 cal. yr BP).
Fossil pigment values were low in the calcareous marl sediments between 220- and 140-cm core depth but better in the dark organic peat layers between 136- and 96-cm core depth. This pattern is similar with the low pollen conservation levels in the lower part of the core, and is assumed to reflect a low preservation of organic material. Therefore, pigment results are only discussed for the upper part of the core. Peak concentrations of zeaxanthin/β,β-carotene and diatoxanthin/β,β-carotene ratios were observed at 117-cm depth (4150 cal. yr BP). Ratios were used instead of absolute concentrations in the sediment to avoid the effect of different degradation rates in the various layers of the core, as past studies have shown that most pigments are preserved more efficiently in anoxic than in oxic conditions (Bianchi et al., 1993; Sun et al., 1993). Zeaxanthin is a particularly stable pigment biomarker for cyanobacteria (Bianchi et al., 2000; Fernandez-Carazo et al., 2013). Diatoxanthin is an indicator for diatoms, while β,β-carotene is a very stable nonspecific pigment that is present in all classes of algae making it suitable as indicator for overall biomass (Bianchi et al., 2000; Hurley and Armstrong, 1991). The zeaxanthin/β,β-carotene ratio can thus be used as a measure of the relative abundance of cyanobacteria (Bianchi et al., 2000). Cyanobacteria occur often in high densities in eutrophic or hypertrophic conditions which likely occurred at times of the observed high zeaxanthin/β,β-carotene ratios.
Discussion – Environmental reconstruction
The MAS rock valley was formed after a phreatomagmatic explosion toward the end of the Pleistocene (Figure 5a). Tropical weathering of basalt led to the formation of a tropical clay (Rijsdijk et al., 2009; Figure 5b). Aeolian coral sands were deposited since the onset of the late glacial when sea levels started to rise and the coastline migrated land inwards (Figure 5c). The rising sea level caused the groundwater table to rise. The water table reached the coral sand surface of MAS sub-basin 1 during the final phase of sea level rise around 5000 years ago. Subsequently, a small and shallow lake started to emerge, reflecting a low energy environment where decomposed organic material started to accumulate below the water table, periodically interrupted by aeolian events that deposited the sand, forming lamina (Figure 5d).
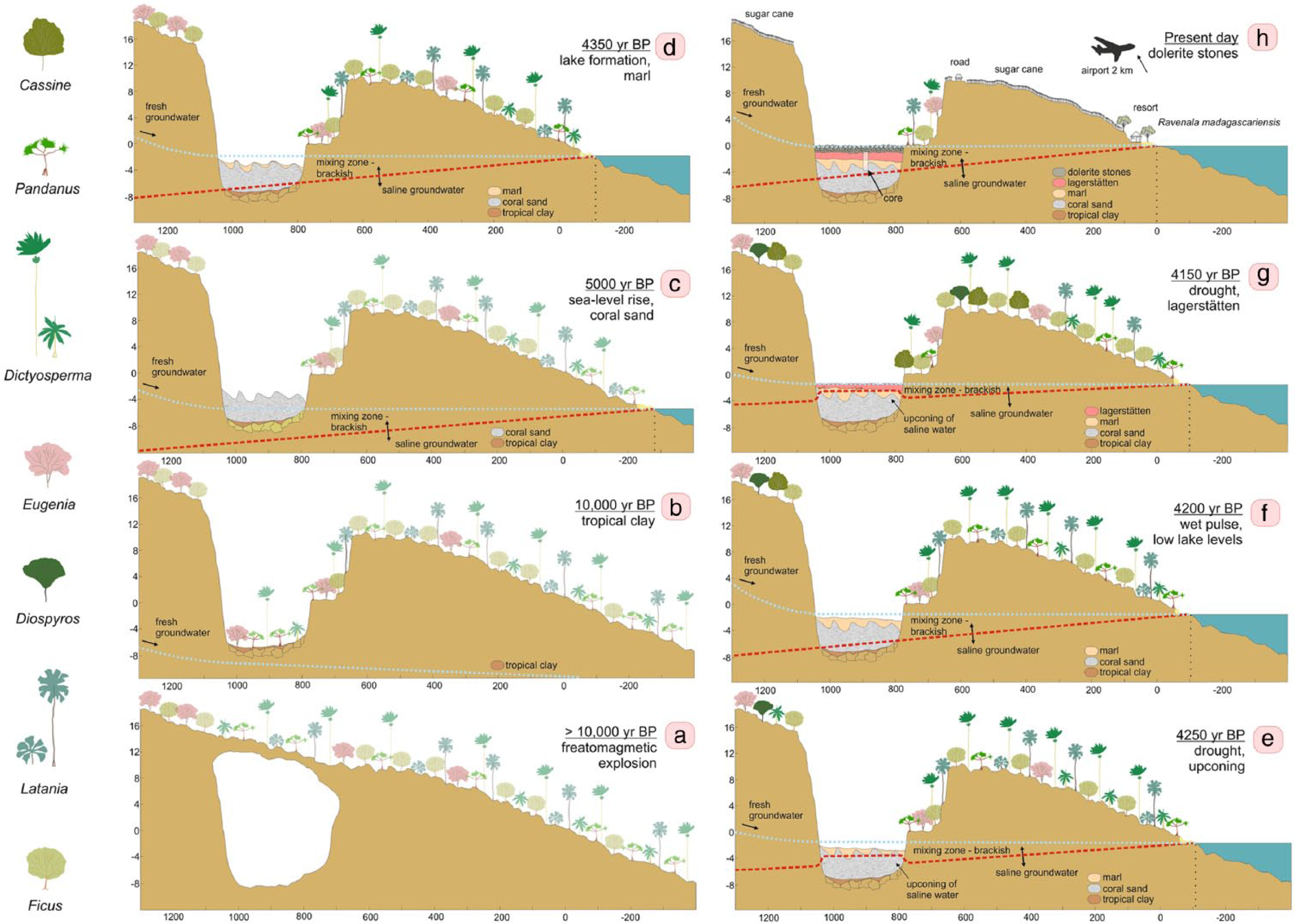
Schematic environmental reconstruction of Mare aux Songes (MAS) sub-basin 1 in relation to the changing position of the coastline and the surrounding coastal lowlands at different snapshots in time ((a)–(h)). The accumulation of sediment in MAS sub-basin 1 is driven by sea level rise. Droughts and wet pulses mark changes in the hydrology of sub-basin 1 caused by shifting boundaries of saline and fresh groundwater. Trends of vegetation change are shown by the following seven main plant genera: Pandanus, Dictyosperma, and Latania (reflecting palm woodland); Eugenia, Cassine, and Diospyros (reflecting semi-dry evergreen forest); and Ficus (reflecting palm woodland and semi-dry evergreen forest). Vegetation is shown transparent during snapshots (a) to (c) when no pollen data are available.
Under present-day conditions, water levels at MAS follow relative groundwater level variations which are controlled by the dynamic balance between tidal fluctuations and rainfall. Levels increase when groundwater discharge from higher ground increases (Rijsdijk et al., 2011) and by lowering of the interface between fresh and saline groundwater. Around 4500 cal. yr BP, sea levels were approximately 175 cm lower than present (Camoin et al., 2004) resulting in a lake at MAS that was about 1 m deep.
Period MAS-1: 4370–4320 cal. yr BP; 220- to 198-cm core depth
The diatom record is almost completely dominated by the freshwater but saline tolerant species Fragilaria brevistriata, indicating that water conditions are mainly controlled by the inflow of fresh groundwater from the surrounding area, but also influenced by more saline groundwater from below. The presence of several brackish–saline Amphora species also suggests an increased influence of saline groundwater, i.e., rising of the interface between fresh and saline groundwater.
Vertebrates such as the now extinct giant tortoises and dodo would have been attracted to the MAS rock valley as it provided an important source of fresh water in the southeastern lowlands. The presence of spores of Sporormiella and other coprophilous fungi indicates that these larger vertebrates were visiting the rock valley, as these spores have a limited dispersal distance (Wood et al., 2011). Stagnant freshwater is rare in this part of the lowlands because of rocky basaltic soils with poor water retention capacity and high evaporation rates (Rijsdijk et al., 2009; Figure 1c). The dominance of F. brevistriata also indicates that water levels were relatively high and were relatively rich in suspended organic matter. While organic matter is partially derived by the decomposition of plant debris produced by the vegetation fringing MAS, the presence of several N-tolerant diatom species suggests that sub-basin 1 received a significant input of excrements from the larger vertebrate species.
Period MAS-2: 4320–4190 cal. yr BP; 198- to 134-cm core depth
Decreasing values of ln(Ca/Ti) indicate a reduced influence of marine water. This is also reflected in the diatom record, where the freshwater but saline tolerant diatoms Fragilaria brevistriata, F. construens, Navicula cuspidate, and Anomoeoeneis sphaerophora var. sculpta are dominant. Increasing values of freshwater diatoms, brackish–saline diatoms, and freshwater diatom spores indicate more frequent changes in salinity reflecting meteorological variations. The presence of aerophilic diatoms and gradually increasing input of charcoal suggest that MAS dried out and fires were frequent in the surrounding lowlands, indicating that dry conditions prevailed. Likely, seasonal contrasts became more pronounced (Marchant and Hooghiemstra, 2004). Peaks of charcoal particles around 4220 cal. yr BP (152-cm depth) coincide with peak values of Sporormiella. The large input of spores from coprophilous fungi points to a higher concentration of larger vertebrates in the rock valley, suggesting that these animals were more dependent on freshwater availability from this particular wetland during times of drought.
After 4230 cal. yr BP (156-cm depth), the disappearance of freshwater spores and aerophilic diatoms, decreasing numbers of brackish–saline diatom species, and gradually decreasing proportions of Fragilaria construens suggest a short interval of less dry climate conditions. The charcoal content after 4220 cal. yr BP almost disappears, also suggesting less arid conditions and/or a reduction of the potential burnable biomass.
Period MAS-3: 4190–4130 yr BP; 134- to 106-cm core depth
The transition from period MAS-2 to MAS-3 coincides with the transition from lake marl to peat. This environmental change is reflected by several shifts in the composition of the diatom community and an improved preservation of pollen grains and pigments (Figure 3). Coral reconstructions by Camoin et al. (2004) show that sea levels were approximately 135 cm lower than present-day, which suggests that at the start of this period sediment infill in MAS sub-basin 1 equated the sea level controlled groundwater level. The transition from calcareous lake marl to peat therefore represents a terrestrialization of MAS sub-basin 1 from a shallow lake to a swampy peat land. A temporarily increased proportion of Poaceae pollen suggests that vegetation encroached in the wetland ecosystem. This is also reflected in the lower part of the peat layer, which consists of a woody peat containing roots (Rijsdijk et al., 2009). A decrease in brackish–saline diatoms also indicates a reduced marine influence in sub-basin 1. However, conditions at MAS remained wet as fresh groundwater continued to flow into the rock valley from surrounding higher grounds, most notably during the wet season. Cyclotella meneghiniana rapidly takes over the entire diatom community for a brief period from approximately 4190–4170 cal. yr BP (134- to 126-cm depth). This planktonic diatom species indicates organic-polluted conditions (Moro and Furstenberger, 1997) and grows optimally in nitrogen-rich waters (Gasse, 1986).
The woody peat layer is overlain by an organic fossil layer containing bone material from more than 20 different vertebrates, including two extinct species of giant tortoise and the dodo (Rijsdijk et al., 2009). Because of its high concentration of fossil material, this layer is categorized as a vertebrate-bone concentration–Lagerstätten; sediments comprise large woody plant remains (up to 50 cm in length) and bone fragments (up to 40 cm in length) and smaller elements such as twigs and seeds (Table S1, available online; Rijsdijk et al., 2009). The onset of bone accumulation coincides with a shift in the diatom record where Nitzschia capitella and Navicula cuspidata take over the assemblage, previously dominated by Cyclotella meneghiniana. This shift is accompanied by peak values of diatoxanthin and zeaxanthin pigments relative to β,β-carotene, indicating an increase in suspended organic matter reaching hypertrophic levels, which corresponds to the concentrated fossil bone bed reflecting an environment where 100,000s of vertebrates died during a short time interval of less than 150 years (Rijsdijk et al., 2011). Higher numbers of spores of Sporormiella suggest that a larger number of vertebrates congregated in the rock valley, plausibly because their dependence on freshwater in MAS sub-basin 1 was greater during times of aridification. Rising values of Br/Ti indicate that climate conditions were increasingly drier.
The Tatos rock valley in the eastern Mauritian lowlands (Figure 1b), which records a dynamic environmental history during the last 8000 years, shows an unstable climatic period between approximately 4500 and 4000 cal. yr BP, during which decadal to centennial droughts alternate with decadal wet pulses (De Boer et al., 2014). Unequivocal indications for driest Holocene conditions are recorded at Tatos around 4300 cal. yr BP, which correspond to a global-scale period of monsoon weakening and a series of dust-peak events (Davis and Thompson, 2006; Fleitmann et al., 2007; Thompson et al., 2002; Figure 6). This aridity event, also referred to as the ‘4.2 ka Megadrought’ has been recorded in other sites around the Indian Ocean, and was considered a driving factor in civilization collapses in Pakistan, Mesopotamia, and Eastern Africa (Cullen et al., 2000; MacDonald, 2011; Staubwasser et al., 2003; Thompson et al., 2002; Wang et al., 2005). The complex anatomy of this climatic drought period is reflected in the δ18O curve from Fleitmann et al. (2007), which records a prolonged period of monsoon weakening superimposed by short intervals of enhanced precipitation (Figure 7). At MAS, this prolonged drought period is recorded between 4320 and 4130 cal. yr BP, punctuated by two wetter intervals at ~4290 and ~4200 cal. yr BP (Figure 7).

Selected pollen and diatom records of Mauritian climate during the last 8000 years compared with Holocene monsoon records from circum Indian Ocean locations. The horizontal bar indicates the time window of the MAS1 record. During this 500-year time interval, driest Holocene conditions are recorded in the eastern Mauritian lowlands (De Boer et al., 2014), coinciding with a reduction in the strength of northern hemisphere monsoons (Oman: Fleitmann et al., 2007; China: Wang et al., 2005), dust peaks (Oman: Cullen et al., 2000; Huascaran: Davis and Thompson, 2006; Kilimanjaro: Thompson et al., 2002), and wettest Holocene conditions in western tropical Australia (Denniston et al., 2013).

Summary diagram of environmental change in Mare aux Songes (MAS) sub-basin 1, Mauritius. Various proxy records from the MAS1 record are shown on a linear timescale. A comparison with environmental change in Oman as inferred from the Qunf stalagmite record (Fleitmann et al., 2007) is shown.
Small islands are geo-hydrologically very sensitive to changes in water supply and amplify small climatic perturbations because of the short response times of small catchments and aquifers. As MAS represented a fresh water hole, it attracted and concentrated vertebrate animals (Figure 8). During the drought between 4320 and 4130 cal. yr BP less freshwater entering at MAS and the upconing of the fresh–saline groundwater interface resulted in a salinization of the wetland. In addition, water levels decreased and the shallow lake shrank in size, resulting in a further concentration of animals. The increased input of excrements in the wetland as recorded by coprophilous fungi and N-tolerant diatoms led to hypertrophic conditions. A mix of organic pollution, high temperatures, and intense sunlight provide optimum conditions for the growth of cyanobacteria and microalgae (Briand et al., 2003; Pitois et al., 2000). Peak values of pigments of cyanobacteria in the MAS-record indicate the occurrence of potentially harmful algal blooms during this period. Many cyanobacteria produce toxins, which can cause serious and sometimes fatal liver, digestive, and neurological diseases (Carmichael et al., 2001; Codd et al., 2005; Dittmann and Wiegand, 2006). Poisoning of vertebrate animals at MAS by drinking toxic water could have resulted in immediate death, explaining the presence of animals at the center of the wetland. Many incidents of cattle or wild animal intoxication have been reported from all parts of the world (see list in Kotut et al., 2006 and Pitois et al., 2000). A Pleistocene example is represented by the Neumark-Nord Lake, where seasonal blooms of cyanobacteria were associated with the mass death of large mammals (Braun and Pfeiffer, 2002). Seasonal blooming at MAS between 4190 and 4130 explains the high concentrations of fossils. Less mobile vertebrates dependent on the availability of freshwater at MAS would be intoxicated in the swampy wetland; the carcasses would be distributed over the swampy wetland during the wet season, when water levels increased and improved the water quality.

Artist impression of pristine Mare aux Songes: ‘Mauritius palm-rich Dodo habitat ca. 4000 years ago, based on pollen and fossil wood analysis. Forest dominated by Latania and Dictyosperma palms, and with Tambalacoque Sideroxylon grandiflorum’ (painting by Julian P Hume, 2014; artwork commissioned by TV and EJdB).
Period MAS-4: 4130–4110 cal. yr BP; 106- to 96-cm core depth
This period corresponds to the top of the MAS1 record, which was disturbed by small stones that penetrated deeper into the sediments after the wetland was filled in with dolerite rocks in the 1940s. Nonetheless, the microfossil records show little signs of human contamination. The diatom record remains stable except for the return of Fragilaria brevistriata, indicating relatively deeper water conditions. The pollen record shows an increase of semi-dry forest with increasing proportions of Diospyros (ebony), Zanthoxylum, Eugenia, and Ixora. Lower numbers of charcoal, freshwater diatom spores, and coprophilous fungi indicate that water levels got higher and MAS turned back into a shallow lake.
Conclusion
Boundaries of island life
This study describes the direct result of an extreme drought 4200 years ago on a local system. But how unique was this event? No similar climatic events during the Holocene were recorded in the Mauritian uplands, related to the long-term high precipitation levels facilitated by elevated topographies (De Boer et al., 2013b). Compared with the uplands, the coastal lowlands are much more sensitive to seasonal variation and consequently to droughts.
A unique combination of factors was responsible for the deposition of the rich fossil bed at MAS. First, the 4200 cal. yr BP drought led to decreased water levels and salinization. Second, at the same time the lake became terrestrialized as the sediment accumulation reached the lake level; the shallow lake turned into a muddy wetland allowing the animals to encroach to the center of the wetland. Third, high alkalinity and seasonally changing water levels facilitated an excellent preservation of bones and other fossil material (Hume et al., 2014; Rijsdijk et al., 2009; Van der Sluis et al., 2014). Climatic and hydrological conditions were similar at Tatos (De Boer et al., 2014). However, as the Tatos basin was deeper than MAS, Tatos was still a too deep lake during the 4200 cal. yr BP drought, preventing non-volant vertebrates entering the basin.
The ‘4200 cal. yr BP megadrought’ was an extreme event in the Indian Ocean and surrounding areas (e.g. Booth et al., 2005; Gasse, 2000; MacDonald, 2011; Marchant and Hooghiemstra, 2004; Staubwasser and Weiss, 2006; Thompson et al., 2002; Williams, 2009), but it was not unique; likely more similar extreme aridity events occurred that may have affected insular biota (Fleitmann et al., 2007; Wang et al., 2005). These events and biotic response may have been registered in other sites with a rich integrated fossil preservation on islands (e.g. Burney et al., 2001, 2004). Therefore, we assume that population collapse at MAS itself was not a unique event; rather, the sedimentary sequence and the preservation of the fossils are distinctive here. Life on islands was not always a paradise for those species that relied on a stable source of freshwater. During times of climatic instability and drought, nonterritorial volant species, such as flamingos, ducks, and pigeons, could move to other parts of the island with a different micro-habitat. However, non-volant and territorial freshwater-dependent species such as the dodo and giant tortoise were tied to the local water holes and may have experienced severe climatic bottlenecks (e.g. Olivieri et al., 2008). It is significant that many of these insular species went extinct shortly after human arrival, although they had persisted dynamic climate conditions throughout the Quaternary.
Footnotes
Acknowledgements
We thank Annemarie Philip for preparing pollen and diatom samples and Pieter Slot for pigment analyses. Pieter Baas is thanked for his valuable comments on the manuscript. We thank all the members of the International Dodo Research Project for their help in the field (2010 fieldwork) and the many stimulating discussions. The sugarcane company OMNICANE (Mauritius) is greatly acknowledged for logistic support in the field.
Funding
We thank the Netherlands Organization for Scientific Research (project number ALW 819.01.009) for financial support.