
Abstract
Palaeoclimatic reconstructions from lake sediment biological records can be challenging, due to variation in non-climatic factors, which alter ecosystem responses. To consider this, it is important to replicate a study regionally, so as to gain information on spatial variability of ecosystem response and the influence of site-specific conditions. Previous pollen-based palaeoclimatic records from three well-dated Arctic lake sites highlight the response of regional Scots Pine (Pinus sylvestris) and Mountain Birch (Betula pubescens ssp. czerepanovii) forest-tundra transition to Holocene climatic variability and suggest the northernmost Peninsulas of Finnmark to be climatically sensitive. This study analysed dated sediment sequences between c. 3970 and c. 6200 cal. yr BP from these three previously published shallow lakes: Liten Čap’pesjav’ri (LCJ), over Gunnarsfjorden (OGF) and over Kobbkrokvatnet (OKV), for freshwater diatoms. Diatom assemblages showed an increase in the planktonic diatoms relative to benthic diatoms, with an onset towards higher abundances of small centric planktonic diatoms at OGF (between c. 5270 and 5350 cal. yr BP) and OKV (between c. 5280 and 5350 cal. yr BP). Additionally, a diatom compositional shift was detected at LCJ between c. 5180 and 5300 cal. yr BP. Trend analysis found the main diatom compositional change at all three sites to occur at 5300 cal. yr BP. However, this synchronous diatom shift during the mid-Holocene varies in magnitude within the three Finnmark lakes. The abrupt planktonic:benthic diatom ratio changes are independent of the Pinus:Betula ratios and are likely to be a result of lake ice-cover changes, allowing longer growing seasons, greater water column stability and higher nutrient concentrations from surrounding catchments. This study highlights the significant differences in aquatic ecosystem and terrestrial vegetation response to climatic changes, with diatom assemblages at these sites either responding before the regional tree-line shift or to a separate climatic event within the mid-Holocene.
Introduction
Lake ecosystems clearly respond to changes in meteorological factors such as temperature, wind and precipitation, and these changes are recorded in lake sediments over a range of timescales (Adrian et al., 2009; Battarbee, 2000). However, the challenge for palaeolimnologists remains to determine how, and to what degree, past climate is recorded in individual lake sediment records. In particular, the influences of non-climatic factors, and non-linear, and non-stationary responses mean that individual sites can produce quite different records resulting in spatial heterogeneity in the direction and timing of changes (Fritz, 2008). This problem is compounded by the scarcity of studies attempting to distinguish between local influences and regional patterns, and when such studies have been conducted, dating uncertainties can make comparisons difficult. Studies recommend assembling sites from a large spatial distribution to help disentangle localised from regional variability (Fritz, 2008). This study represents an attempt to examine lake response to regional climate by using three previously published and well-dated, pollen-based palaeoclimate records from Arctic sites to determine the influence of a well-documented climate change on the lake ecosystems.
Lakes in the Arctic, free from direct human influence, are usually seen as being sensitive to changes in weather and climate (Catalan et al., 2013; Rühland et al., 2008; Smol et al., 2005). The effect of temperature has been shown to be important in Arctic limnology where air temperature controls ice-cover (formation, duration and thickness) and affects the thermal stability within shallow lakes (Battarbee, 2000; Kilham et al., 1996; Post et al., 2009; Smol, 1988). Changes in temperature, wind and precipitation affect (directly and indirectly) aspects of lake ecosystem functioning, including growing season length, the extent of light penetration and lake water mixing, along with nutrient availability within the benthos and water column (Findlay et al., 2001; Karlsson et al., 2009; Post et al., 2009). Furthermore, climatic changes can also impact the complex lake-catchment feedbacks, changing lake metabolism, within-lake carbon processing and the export of catchment-derived dissolved organic carbon (Birks et al., 2000; Engstrom and Fritz, 2006; Heggen et al., 2010; Jones and Birks, 2004; Jones et al., 2011). As a result, the ecological consequences of these lake environmental changes on species composition and Arctic food-web structures are similarly complex (Jeppesen et al., 2003). Diatoms provide a useful proxy for assessing the impact of climatic change on Arctic lake ecosystems. Diatoms are unicellular siliceous algae, which can provide a unique insight into past lake environmental conditions through direct and indirect inferences of, for example, changes in lake water pH, salinity and nutrients (Bigler and Hall, 2003; Jones, 2013). Diatoms respond indirectly to climate changes through alterations in lake water properties associated with warming, such as longer ice-free periods, changes in thermal structure and changes in resource availability of light and nutrients.
The assumption is often made that higher proportions of planktonic diatoms in relation to benthic diatoms indicate longer periods of ice-free conditions, turbulent waters and more water column nutrients associated with warmer climatic conditions (Grönlund and Kauppila, 2002; Lotter and Bigler, 2000; Smol, 1988). In contrast, prolonged ice-cover is assumed strongly to inhibit planktonic growth and reduce lake productivity, with benthic diatoms (especially Fragilaria sensu lato spp.) dominating (Karlsson et al., 2009). In the Arctic, much attention has been given to the dynamics of small centric Cyclotella populations, which in some studies have been shown to be sensitive to effects of recent climatic warming (Catalan et al., 2002; Prowse et al., 2011; Rühland et al., 2008; Smol and Douglas, 2007; Winder et al., 2009), often displaying an increased abundance (at the expense of planktonic Aulacoseira and benthic Fragilaria populations) which has been related to their ability to thrive in lakes when the length of the growing season increases and there are longer periods of water column stability (Catalan et al., 2013). These may be related to less summer ice-cover (Prowse et al., 2011; Smol and Douglas, 2007). However, like other phytoplanktonic species, these diatoms respond directly to neither weather nor climate, but to proximal growing conditions (nutrients, light, temperature, mixing regimes and grazing), which are controlled by climate in a complex way (Adrian et al., 2009; Birks et al., 2000; Catalan et al., 2013; Jones and Birks, 2004; Pienitz et al., 1995; Rühland et al., 2008; Rühland et al., 2013; Saros et al., 2013; Smol, 1988; Smol et al., 2005). This has been the premise for interpreting diatom changes in response to climate reported in many limnological and palaeolimnological studies. Experiments have successfully shown that nutrient concentrations and incubation depth (light availability) play important roles in the growth of centric diatoms (Discostella stelligera, Cyclotella comensis and Cyclotella bodanica) within Arctic lakes (Saros et al., 2013). Nevertheless, fluctuations in past climate have induced changes similar to these recent Cyclotella–Aulacoseira–Fragilaria shifts (Ampel et al., 2010; Huber et al., 2010; Lami et al., 2010; Wang et al., 2008; Wilson et al., 2008), suggesting that these current taxon-specific changes are in response to an overriding effect of climate (Catalan et al., 2013) and that similar mechanisms may occur in the late mid-Holocene. However, at some Arctic and Alpine sites, Cyclotella spp. dominate throughout the Holocene (Perren et al., 2009), and at others, documented warming has not resulted in an increase in Cyclotella spp. (Koinig et al., 2002). Therefore, it is not expected that all Arctic lakes will show an increase in planktonic Cyclotella species in response to warming, or their first appearance, as some lake sites have long had Cyclotella species present. Therefore, it is important to consider the shift in the whole diatom assemblage and not just within the Cyclotella sensu lato (s.l.) taxa.
Conditions of the last 6000 years are of particular interest to further our understanding of the Earth System because the boundary conditions of the climate system did not change dramatically (Wanner et al., 2008). How lake ecosystems responded to past changes in the late mid-Holocene may, therefore, give us clues on how they might respond in the future. The climate of the North Atlantic region has fluctuated on centennial to millennial timescales throughout the Holocene, as shown by palaeoclimatic studies from the Barents Sea (Duplessy et al., 2001; Risebrobakken et al., 2010; Sarnthein et al., 2003; Voronina et al., 2001) and Icelandic Sea (Bianchi and McCave, 1999; Rousse et al., 2006), as well as from Greenland (Bond et al., 1997, 2001; Hall et al., 2004; Mayewski et al., 2004) and Northern Fennoscandia (Korhola et al., 2000, 2002; Rosen et al., 2001). Terrestrial sites from the Barents Sea margin also show evidence of climatic changes during the Holocene (Allen et al., 2007; Huntley et al., 2013), where the role of the Norwegian and North Cape currents (NCaC) are of critical importance. These are branches of the North Atlantic current, which transport warm waters along the Norwegian coastline and around the North Cape region, into the Arctic basin of the southern Barents Sea (Figure 1) and control the regional temperatures around the southern Barents Sea, driving sea ice-cover and sea-surface temperatures (SST; Risebrobakken et al., 2010).
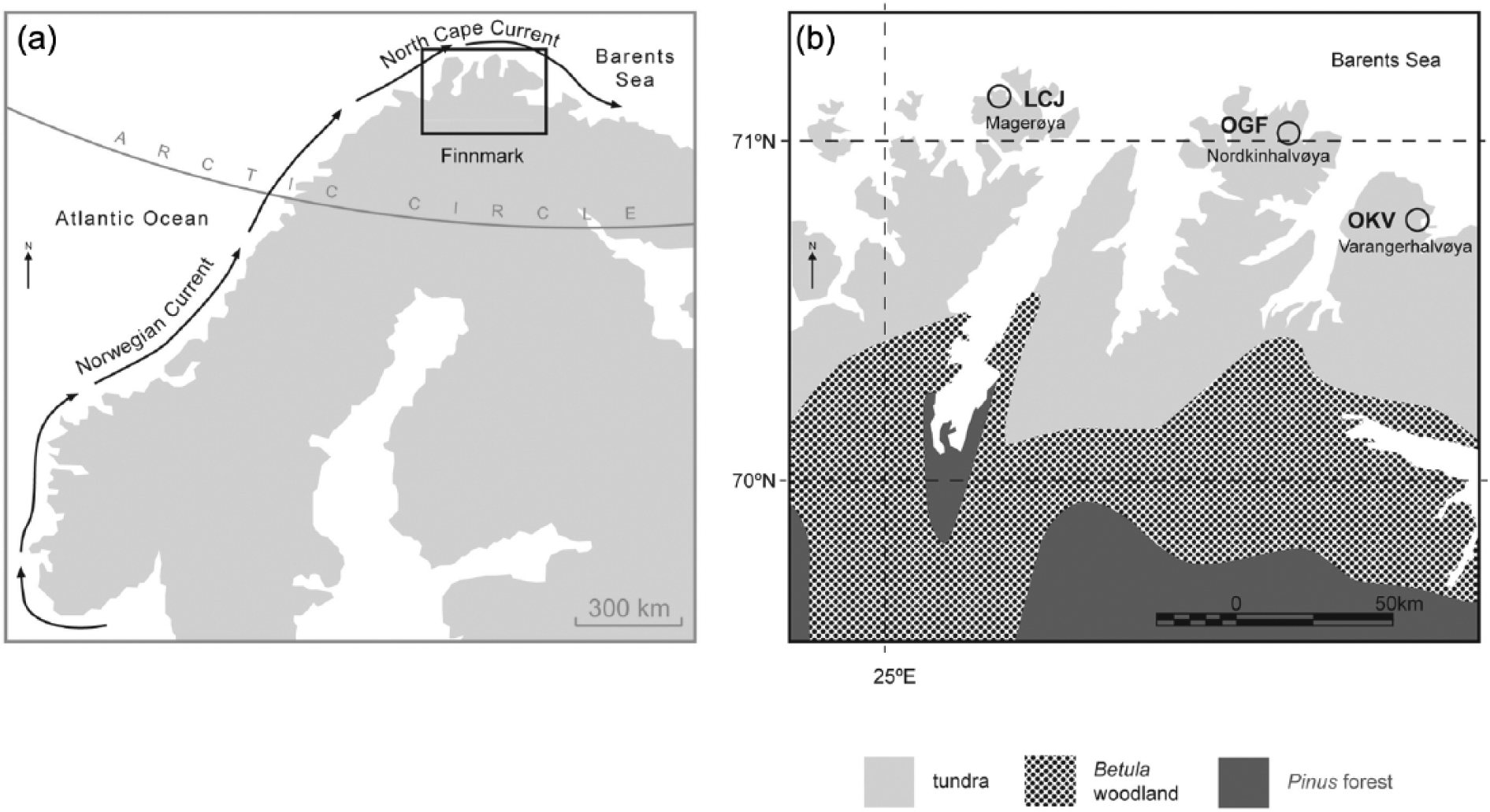
(a) Location of study area in Northern Scandinavia and route of major surface currents. (b) Map showing the location of the three lake sites in Northern Finnmark and the present-day tundra-Betula and Pinus:Betula ecotone positions.
Palaeovegetation research at three Arctic sites in Northern Finnmark situated along a West-East transect, adjacent to the southern shore of the Barents Sea, shows a regional response of the ecotone between Pinus forests and Betula woodlands to Holocene climatic changes (Allen et al., 2007; Huntley et al., 2013). Pollen analysis from tightly chronologically constrained lake sediment cores was used to reconstruct spatial and temporal patterns in the Holocene position of this ecotone (Huntley et al., 2013). In a c. 2000-year period between c. 3970 and 6200 cal. yr BP, the largest shifts in the Pinus:Betula ratio of the entire Holocene period were found at the three sites lying along a c. 170-km West-East transect (Figure 1). The Pinus:Betula ratios show two significant peaks at all three sites (Figure 2), which are short-lived at c. 5675 cal. yr BP (named peak (d) in Figure 9 of Huntley et al., 2013; 95% age ranges at the sites ordered West-East 5450–5850, 5475–5875 and 5600–5850 cal. yr BP) and at c. 4525 cal. yr BP (peak (e) in Huntley et al., 2013; 95% age ranges at c. 4300–4675, 4350–4750 and 4100–4750 cal. yr BP). These suggest episodes of regional climatic warming, resulting in northerly shifts of the Betula tree-line and Pinus–Betula ecotone, as inferred from higher ratio values associated with increased pollen accumulation rates (Allen et al., 2007; Huntley et al., 2013). Palaeoclimatic reconstructions suggest that in these warmer periods, there was a higher mean annual thermal sum than currently experienced in Finnmark at present, and mean temperatures of the warmest month were 0.7–2.2°C warmer than the present day (Huntley et al., 2013).
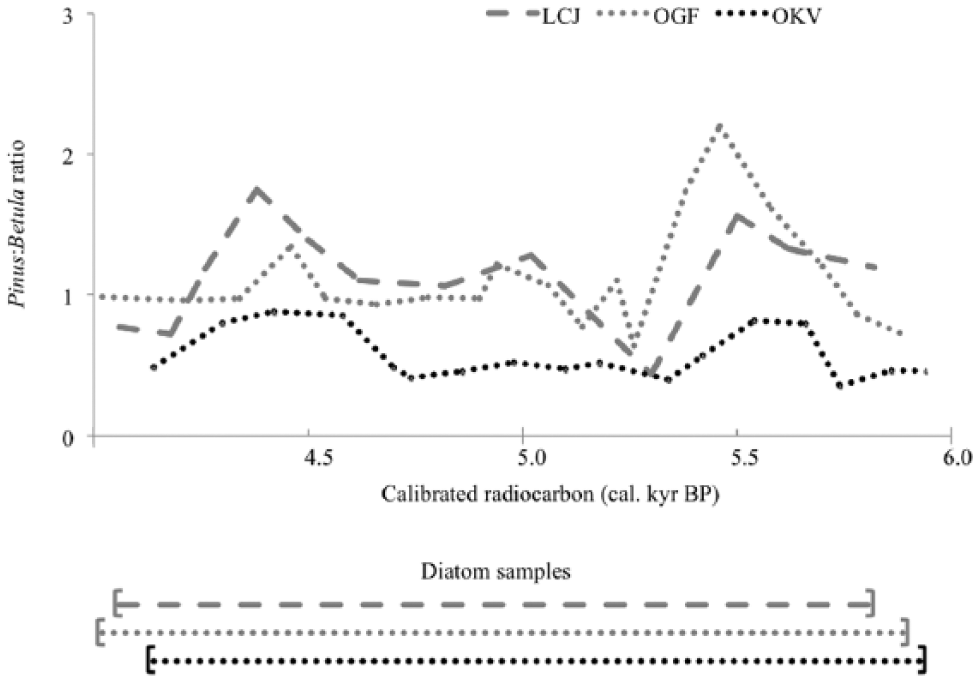
Pinus:Betula ratios at all three lake sites between c. 3970 and c. 6200 cal. yr BP. The time intervals analysed for diatoms at each lake site are displayed underneath.
This study aims to establish the effect of these Holocene climatic fluctuations between c. 3970 and c. 6200 cal. yr BP on diatom community composition within the three Arctic lakes: Liten Čap’pesjav’ri (LCJ), over Gunnarsfjorden (OGF) and over Kobbkrokvatnet (OKV). We hypothesise that if lake ecosystems are sensitive to climate, and if diatom assemblages reflect climatic changes, then diatom community composition should be driven by fluctuations in the strength and eastward penetration of the NCaC and respond with the climatically driven Pinus–Betula ecotone shifts. Although lake biota can respond to catchment vegetation shifts (Jones et al., 2011), in this case, there have been no major shifts in catchment vegetation (Allen et al., 2007). As a result, the aim is to examine biotic response to climatic changes in the absence of major catchment vegetation shifts. This study will investigate whether there are any synchronous diatom compositional changes in terms of planktonic:benthic diatom ratio shifts, between the three lakes located across a West-East gradient spanning a distance of ~170 km. The West-East gradient will allow us to address potential spatial variability and site-specific differences in diatom responses to mid-Holocene climate change. We expect that higher planktonic:benthic diatom ratios will be observed during the Pinus:Betula ratio peaks (at c. 4525 and c. 5675 cal. yr BP), when regionally warmer climatic conditions are inferred.
Study sites
The vegetation of the northernmost peninsulas of Finnmark is especially sensitive to changes in the Atlantic meriodional overturning circulation (AMOC), and in particular to variations within the strength and eastward penetration of the NCaC (Huntley et al., 2013). Huntley et al. (2013) propose that a stronger AMOC and a greater eastward penetration of the NCaC into the southern Barents Sea resulted in higher temperatures and a northerly shift of the Pinus–Betula ecotone, or increased pollen productivity within the Pinus forest, with higher Pinus:Betula ratios. In contrast, a weaker NCaC resulted in southward retreat or decreased pollen productivity of Pinus, with lower Pinus:Betula ratios, due to colder conditions. Today, Scots Pine (Pinus sylvestris) forms the northernmost boreal forests within Fennoscandia, being situated south of the Mountain Birch (Betula pubescens ssp. czerepanovii) dominated sub-Arctic woodlands that extend to the Arctic tree-line (Huntley et al., 2013).
Only brief site details are given here, further information is given in Huntley et al. (2013). The three small oligotrophic and shallow Arctic lake sites are situated beyond the Pinus and Betula limits, north of 70°N and adjacent to the southern Barents Sea shore (Figure 1; Table 1). They form a West-East transect (25°–29°E) across the northernmost Peninsulas of Finnmark, spanning a total distance of c. 170 km. LCJ has a combined ice and water depth of 7 m and is the westernmost site on the transect, situated on Magerøya. Situated in Nordkinhalvøya, the central site is OGF with a combined ice and water depth of 4.8 m, while the easternmost site is OKV, situated in Varangerhalvøya with a combined ice and water depth of 2.3 m (Figure 1). All sites lie outside the Younger Dryas ice limits of northern Scandinavian and are c. 100 and 70 km north of the present-day Pinus tree-line and Betula forest-tundra boundary, respectively (Table 1; Allen et al., 2007).
Northern Finnmark lake sites.

Source: Allen et al. (2007) and Huntley et al. (2013).
LCJ: Liten Čap’pesjav’ri; OGF: over Gunnarsfjorden; OKV: over Kobbkrokvatnet.
Northernmost Finnmark (70–71°N) experiences a relatively mild and wet climate compared to other Arctic regions at this latitude, as a result of the influence of the AMOC (Risebrobakken et al., 2010; Romundset et al., 2010). Warm oceanic waters around the Magerøya, Nordkinhalvøya and Varangerhalvøya regions from the NCaC are the cause of mild snowy winters and wet cool summers (Romundset et al., 2010). Two atmospheric pressure systems, the North Atlantic and Polar Arctic Fronts, control the regional climate. The coldest mean month temperatures at the three sites are −3.6°C (LCJ), −5.4°C (OGF) and −6.5°C (OKV) in winter (October–March; Huntley et al., 2013). In summer (April–September), the warmest month mean temperatures are 11.2°C (LCJ), 9.5°C (OGF) and 10.5°C (OKV; Huntley et al., 2013). Present-day catchment vegetation around the three lake sites consists predominantly of Arctic tundra dominated by dwarf-shrub species, including Crowberry (Empetrum nigrum ssp. hermaphroditum) and Dwarf Birch (Betula nana; Huntley et al., 2013).
Methods
Sediment coring, sub-sampling and chronology
A 7.5-cm diameter square-rod piston corer was used from lake ice in the late winter to collect sediment core sections from the deepest points of the three lake basins (Huntley et al., 2013).
OGF was sampled in April 2000 and LCJ and OKV in April 2001. Two sets of consecutive 1-m core segments were collected from each lake with the second set covering the joints in the first. Only one set was used in subsequent analyses as no significant changes were found in the analyses across the joints. The cores were subsequently stored at 4°C at Durham University, UK. Sediment cores comprised mainly of homogenous algal gyttja. AMS 14C radiocarbon dates were obtained by 14C measurements of terrestrial macrofossils. Radiocarbon ages were calibrated, and age–depth models were constructed using a Bayesian method, Bchron (Parnell et al., 2008). This resulted in tightly constrained chronologies at all three sites with 95% uncertainties of c. 200 years for the modelled dates for the majority of the Holocene (Huntley et al., 2013).
Diatom analysis
Wet sediment samples were prepared by conventional methods (H2O2, HCl) with a water bath (Battarbee, 1986; Renberg, 1990). Diatoms were identified to species level at 1000× magnification using a Zeiss light microscope and phase contrast under oil immersion. Published keys and guides (Camburn and Charles, 2000; Krammer and Lange-Bertalot, 1986, 1988, 1991a, 1991b) aided diatom taxonomy. Small centric diatoms (Cyclotella aff. comensis (Grunow) and Cyclotella rossii (H. Hakansson)) dominated the plankton; these had very variable valve morphologies and, because they were difficult to separate consistently, were grouped into a Cyclotella rossii-comensis complex (Cremer and Wagner, 2004; Hausmann et al., 2001). Diatom samples were taken at the same stratigraphic levels as the pollen samples, to enable sample-by-sample comparison between the diatom and Pinus:Betula ratio records for the chosen time interval.
Data analyses
Stratigraphic plots of diatom percentage abundances were drawn using C2 (Juggins, 2007). Discrete diatom assemblage zones were determined using Optimal Partitioning within the program ZONE (version 1.2; Juggins, 1992). The broken stick model (Bennett, 1996) was applied to establish the number of statistically significant zones. Rate-of-change analysis (Birks et al., 2000; Grimm and Jacobson, 1992) was performed on the diatom species data to establish the amount of assemblage change in the stratigraphical profile. The statistical package, Bchron, was applied to test the synchronicity of events in the data sets (Haslett and Parnell, 2008).
Results
The diatom floras from the three Finnmark cores (Figure 3) were diverse with a total of 164 diatom species identified from the late mid-Holocene. The dominant benthic taxa were Stauroforma exiguiformis, Brachysira vitrea, Achnanthidium minutissimum and Nitzschia fonticola. At all three lakes, benthic diatoms dominated the diatom assemblages, but planktonic diatoms were still an important component. The highest relative abundances of planktonic diatoms (16%) were found at OGF, and the other two lakes had notable abundances up to 7–10%. There was a sparse planktonic flora in OGF and OKV, contributing less than 20% of the total diatom assemblage at either site. The diatom valves were well preserved, with minimal amounts of valve breakage and dissolution.
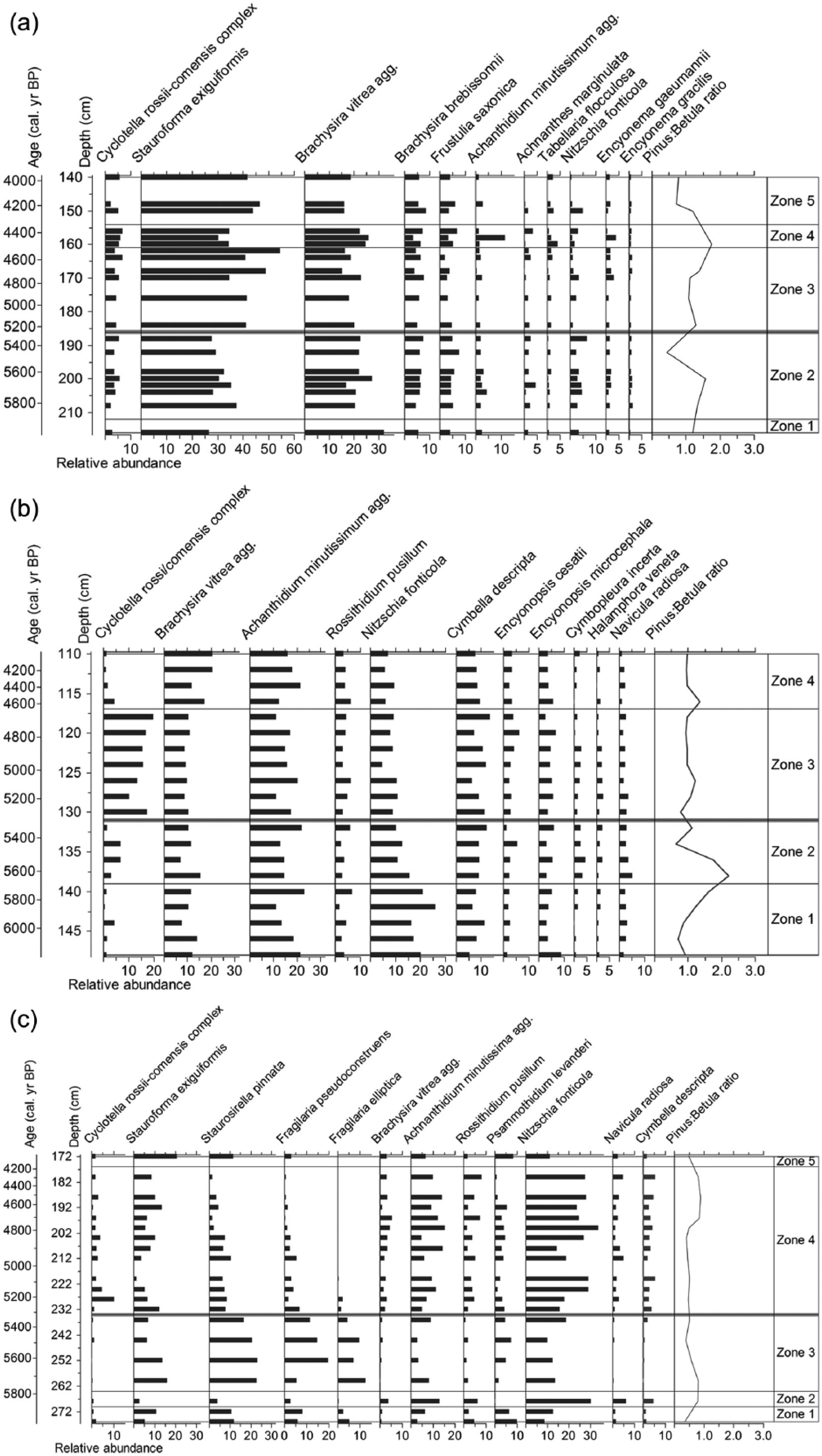
Summary stratigraphic profile of diatom percentage abundance of the most abundant diatom species (>2% relative abundance). Thick grey line highlights onset of diatom community change. (a) LCJ, (b) OGF and (c) OKV.
LCJ
A summary diagram for LCJ is presented in Figure 3a. The Cyclotella rossii–comensis complex is present at low abundance throughout the section with no abrupt changes. Stauroforma exiguiformis and Brachysira vitrea dominate throughout the core sequence. Subtle changes in S. exiguiformis delineate the zones, along with declines in small benthic Achnanthes (s.l.), Tabellaria, Nitzschia fonticola and Encyonema taxa at the onset of Zone 3.
OGF
A summary diagram for OGF is presented in Figure 3b. Cyclotella rossii–comensis complex reaches highest relative abundances in Zone 3. In Zones 1 and 2 (6200–5690 and 5690–5310 cal. yr BP), planktonic taxa are sparse, and there are high abundances of Nitzschia fonticola (11–26%) and Achnanthes sensu lato spp., mainly Achnanthidium minutissimum (11–23%). In Zone 3 (5310–4580 cal. yr BP), there are significant diatom floristic changes, as the Cyclotella rossii–comensis complex increases to relative abundances of 10–16% and Nitzschia fonticola declines to relative abundances of 5–11%. This results in a peak in planktonic:benthic ratios, between c. 5310 and 4580–4700 cal. yr BP (Figure 4). In Zone 4 (4580–4040 cal. yr BP), Cyclotella spp. decline to 7–3%, Brachysira vitrea increases to 12–21% abundance and planktonic:benthic ratios remain low.
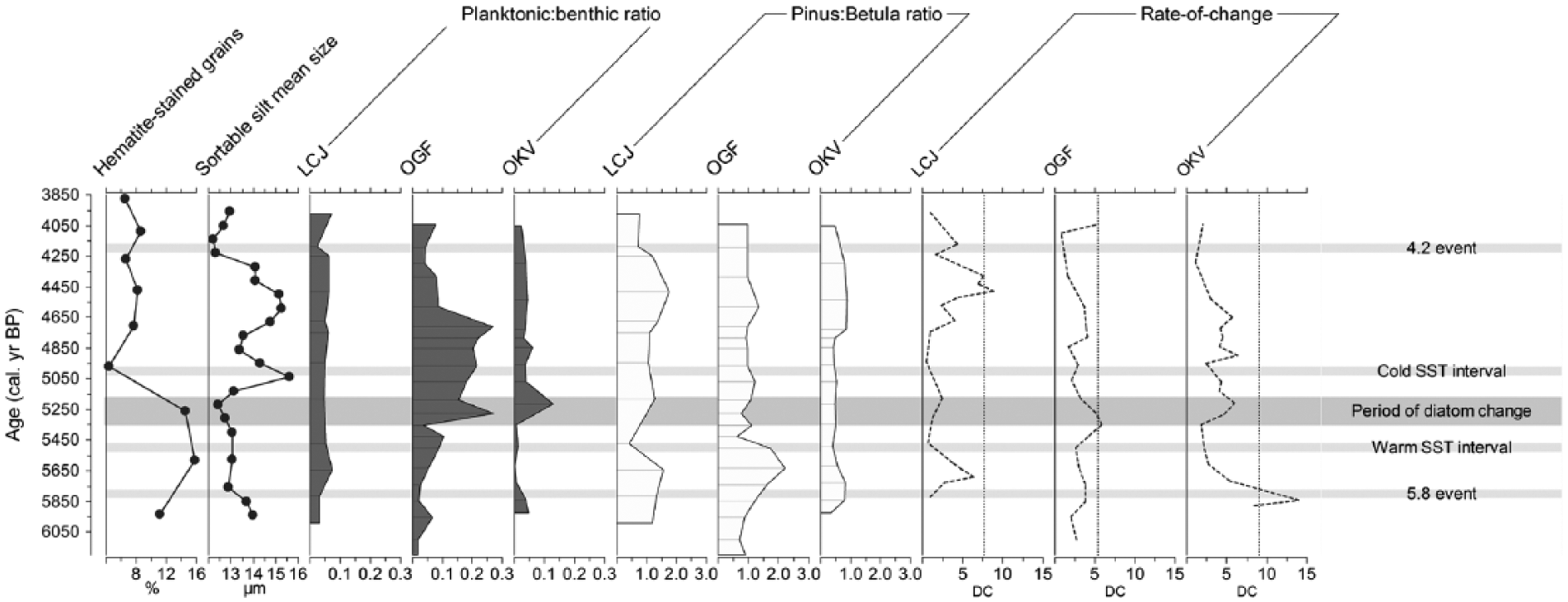
Summary diagram showing North Atlantic climatic proxy data; Haematite-stained grains (HSG) and sortable silt (SS) mean size (Bianchi and McCave, 1999), planktonic:benthic ratios, Pinus:Betula ratios and diatom assemblage rate-of-change data, including significant rate of change lines (95% confidence level). This figure includes the cold SST interval at 5000 cal. yr. BP in SE Barents Sea region (Voronina et al., 2001), warm SST interval at 5500 cal. yr. BP in western Barents Sea region (Sarnthein et al., 2003), IRD event 3 (4.2 event) and IRD event 4 (5.8 event; Bond et al., 1997, 2001) and period of diatom change between c. 5180 and 5350 cal. yr BP.
OKV
A summary diagram for OKV is presented in Figure 3c. Zones 1 and 2 (5920–5840 and 5840–5790 cal. yr BP) mainly consist of benthic Fragilaria sensu lato spp. (33–37%), Achnanthes sensu lato spp. (22–23%) and Nitzschia fonticola (9–13%), with a rich diversity of pennate diatoms, including Navicula radiosa, Brachysira vitrea and Cymbella descripta. Planktonic taxa are sparse. In Zone 3 (5790–5320 cal. yr BP), there are increases in the abundances of Fragilaria sensu lato spp. (Stauroforma exiguiformis, Staurosirella pinnata, F. pseudoconstruens and F. elliptica) and decreases in acidophilous B. vitrea, Achnanthes sensu lato and Nitzschia spp. Planktonic taxa are not abundant. The most notable diatom assemblage changes occur at the onset of Zone 4 (5320 cal. yr BP). In Zone 4 (5320–4300 cal. yr BP), significant diatom floristic changes occur, with a rise in Cyclotella rossii–comensis complex species and large pennate diatoms, such as N. radiosa. Fragilaria sensu lato spp. subsequently decline from 64% to 11%. The plankton:benthic diatom ratio peaks within this zone (Figure 4). In Zone 5 (4300–4050 cal. yr BP), there is a return to higher Fragilaria sensu lato spp. abundances.
Rate-of-change analyses highlight the significant changes (95% confidence levels) in the three diatom records to be at 4480 (LCJ), 5350 (OGF) and 5840 cal. yr BP (OKV; Figure 4). The results from the Bchron statistical analyses suggest that a significant event in the diatom records at all three sites occurred synchronously and at 5300 cal. yr BP. This diatom shift observed across the three sites occurs after the first Pinus:Betula peak at c. 5675 cal. yr BP (LCJ: 5450–5850 cal. yr BP, OGF: 5475–5875 cal. yr BP and OKV: 5600–5850 cal. yr BP) and before the second Pinus:Betula peak at c. 4525 cal. yr BP (LCJ: 4300–4675 cal. yr BP, OGF: 4350–4750 cal. yr BP and OKV: 4100–4750 cal. yr BP; Figure 3).
Discussion
The diatom compositions within the three northern Finnmark lakes during the mid-Holocene show remarkable commonality, with a similar period of diatom change between c. 5270 and 5350 cal. yr BP. However, the synchronous diatom assemblage shifts in the mid-Holocene differ in their magnitude across the three sites. OGF and OKV show higher Cyclotella abundances within the late mid-Holocene, with changes centred around 5300 cal. yr BP. At the central site, OGF, an abrupt taxonomic shift occurred after c. 5350 cal. yr BP, with higher abundances of planktonic taxa (mainly Cyclotella rossii–comensis complex) together with a larger number of motile, large periphytic (attached to plants/mosses) diatoms, such as Cymbella spp., and fewer attached benthic forms. At the easternmost site, OKV, the diatom record also exhibits this trend towards higher planktonic:benthic diatom ratios at c. 5350 cal. yr BP and more motile, larger periphytic pennate diatoms, such as Navicula radiosa. At LCJ, the westernmost site, the diatom composition changes are more subtle, but there is an assemblage shift around c. 5180 and 5300 cal. yr BP (Zones 2–3), which matches the timing of diatom changes observed at the other two sites. Therefore, the diatom communities at all three sites react at a similar time, presumably due to regional climatic events.
Of interest is that higher planktonic:benthic ratios in the diatoms occurred between the timing of two significant peaks in Pinus:Betula ratio. This could be a result of leads and lags in the palaeoecological records not being fully accounted for, namely, the time taken for Pinus sylvestris to migrate northwards, closer to the lake sites on the Barents Sea coast. In comparison to vegetation, diatoms respond rapidly to climate-driven changes of lake ecosystems, and their community assemblage is composed of many different species. Within this diverse assemblage, some species exhibit narrower ecological niches compared to terrestrial vegetation types (Birks and Birks, 2008). Freshwater diatoms are sensitive to both lake water and catchment changes, in particular temperature-associated changes in the water column, ice-cover dynamics and water chemistry changes. Thus, diatoms inherently respond indirectly to regional changes in wind patterns, air temperatures and precipitation fluctuations at a different rate and magnitude to terrestrial vegetation.
The maximum extent of the past Pinus–Betula forest ecotone for this region has been suggested to lie at the present-day Betula-tundra ecotone boundary (Seppä et al., 2002). Therefore, the three lake sites on the northernmost Peninsulas of Finnmark would always have been significantly beyond the Pinus sylvestris limit, although at times close to or within the zone of Betula pubescens ssp. czerepanovii sub-Arctic woodlands (Allen et al., 2007). The catchments of these tundra lakes remained treeless throughout the Holocene. As a result, no significant lake-catchment vegetation changes are likely to have affected the lake ecosystem directly through terrestrial inwash, as seen in Kharinei Lake in North Eastern European Russia where organic inwash occurs (Jones et al., 2011). Pinus sylvestris pollen grains preserved within-lake sediments are wind transported over large distances from distant Pinus sylvestris–dominated forests; for example, for lakes of similar size to those we studied, MacDonald and Cwynar (1991) estimated that 20% of Pinus pollen was derived from beyond a radius of 100–200 km.
Cyclotella increase and climatic change
The periods of higher Cyclotella abundances within OGF and OKV are likely to be a result of decreased summer ice-cover duration and increased growing season length. These climate-driven changes are inter-related to the fundamental lake processes which affect algal production and composition, thermal stability and vertical mixing patterns and availability of resources (light and nutrients; Adrian et al., 2009; Korhola et al., 2000; Rühland et al., 2008, 2013; Saros et al., 2013; Smol et al., 2005; Sorvari et al., 2002). Many remote Arctic lakes have provided evidence for global climatic change from palaeolimnological studies (Catalan et al., 2013; Prowse et al., 2011; Rühland et al., 2008; Smol and Douglas, 2007). These studies have reported increases in small planktonic Cyclotella spp. (Cyclotella comensis and Discostella stelligera) with climatic warming, due to onset of longer growing seasons with higher water column stability, resulting in increased nutrient availability. Studies have shown that climatic warming can result in abrupt changes within the diatom community and increased primary production, with appearances of small and more competitive Cyclotella spp. (Cyclotella comensis and Discostella stelligera) and of large benthic pennate diatoms (Catalan et al., 2013). Increases in the abundance of large pennate diatoms (Navicula radiosa and Cymbella descripta) are observed alongside the Cyclotella rise at OKV. Large complex pennate species require a longer growing season for development in harsh environments, and with warming the diatom assemblage, richness can increase due to the availability of diverse aquatic habitats in near-shore environments, which large pennate diatoms exploit (Keatley et al., 2008; Michelutti et al., 2003; Paul et al., 2010). Therefore, higher large pennate diatom abundances within high-latitude lake ecosystems indicate longer ice-free periods and an increase in the availability of diverse littoral aquatic habitats. Within OKV (Zone 4), the abundances of the less motile benthic diatoms, which inhabit rock surfaces, such as Fragilaria spp., are significantly reduced. This substantial decrease in small benthic Fragilaria taxa and the concurrent increase in a more diverse planktonic and benthic assemblage suggest a change to greater littoral habitat availability with reduced ice-cover extent.
Spatial variability across the transect
The timing of the main diatom compositional change, inferred from the planktonic:benthic diatom ratios, appears to occur synchronously among the three lakes along the West-East transect, and this is supported by trend analysis. Diatoms preserved within OGF sediments record the greatest assemblage change, with an abrupt shift in the planktonic:benthic ratios. This lake may have responded more to climatic change due to site-specific conditions, as OGF has the largest surface area (c. 5 ha) and no discrete inflows in comparison to the other sites (LCJ: c. 2.5 ha and OKV: c. 1.7 ha). OGF may be more sensitive to ice-cover changes, which drive changes in the diatom community, and wind may be playing a stronger role at this lake by making the ice-free period longer than at the other sites. Furthermore, the position of OGF along the coast may have influenced lake response; the site would have been affected by changes within the NCaC strength.
OKV, the easternmost site, exhibits notable changes in diatom community composition but has less Cyclotella species in comparison to OGF. This could possibly be a result of past ocean circulation dynamics around the Barents Sea region, as only a strong NCaC can transport warm waters as far eastwards as the Varangerhalvøya region. The diatom flora composition preserved within OKV suggests a weaker ocean circulation around Varangerhalvøya, as higher abundances of small benthic Fragilaria species were prevalent in Zone 3 of the diatom record. Then, low abundances of large complex periphytic diatoms such as Navicula radiosa and Cymbella descripta appear in the warm period (Zone 4) when benthic Fragilaria species decline substantially.
Surprisingly, the ecosystem response examined at the deepest lake in our study (LCJ) is more muted in comparison to the other two lakes and shows the least amount of diatom change over the period examined. LCJ, the westernmost site, has the highest abundances of Stauroforma exiguiformis and no significant changes within the Cyclotella rossii-comensis complex, which might be a result of site-specific differences. The lack of substantial change in the LCJ diatom profile might be resulting from thermal inertia within this deeper lake, increasing the time taken for the onset of ice-free conditions. LCJ could possibly be less affected by wind-driven ice dynamics and, therefore, takes longer to become ice-free and freezes up more readily at the end of the summer (Keatley et al., 2008).
Comparisons between diatom response and North Atlantic climatic proxies
Records of haematite-stained grain (HSG) percentages and sortable silt (SS) mean sizes (Bianchi and McCave, 1999) from North Atlantic marine sediments provide proxies for sea-ice rafting and the strength of the thermohaline circulation (THC), respectively. Comparing the diatom and Pinus:Betula ratio profiles to the HSG and SS records, over the period of interest, enables this study to examine whether the three lake ecosystems and regional tree-line in Finnmark are responding to changes within the North Atlantic and Arctic air mass circulation. A shift to lower HSG percentages and higher SS mean sizes indicates less ice rafting and a stronger THC. Such a shift occurs at c. 5130 cal. yr BP (Figure 4). Although the onset of higher planktonic:benthic values occurs earlier at c. 5270–5350 cal. yr BP at OGF and OKV, and thus may appear not to correlate with lower HSG percentages or higher SS mean sizes, dating constraints must be considered when making such comparisons. Even our well-dated terrestrial records generally have 95% uncertainty ranges of c. 200 yr, while age uncertainties in the marine record are considerably greater. Moreover, the relationship between the strength of the warm NCaC into the southern Barents Sea and North Atlantic deep-water current strength is complex (Allen et al., 2007). Episodes of warmer and cooler SSTs detected from Barents Sea sediment records do not correspond to the diatom compositional changes (Figure 4). Although similar climatic patterns between 8.5 and 4.3 cal. kyr BP inferred from the Pinus–Betula ratio fluctuations are not consistently seen within the marine records (Hald et al., 2007; Huntley et al., 2013), the vegetation changes may nonetheless be reflecting episodic shut downs in the Barents Sea inflow (Semenov et al., 2009). The diatom responses observed at the three sites apparently reflect independent signals of climatic changes and do not provide evidence of coupled ocean atmosphere forcing through linkages with the North Atlantic Bond Cycles (ice-rafted debris (IRD) events 3 and 4).
Conclusion
Diatom flora shifts have been observed at three small Arctic lake sites located close to the Barents Sea margin in northern Finnmark. The onset of the diatom assemblage shift is synchronous (within the errors of dating) between the three lake sites. It is likely that the species changes observed are indicative of fluctuations of the lakes’ ice-cover duration and thickness, and thus microhabitat availability, lake water nutrient concentrations and light availability, all driven by climatic changes. However, the diatom community shifts differ in magnitude across the West-East transect of study sites, due to site-specific environmental conditions. Small regional climatic changes, in particular temperature and wind, will exert a significant impact on these high-latitude lake ecosystems, as they only experience ice-free conditions for a short period each summer season. These climatic factors will influence lake ice-cover dynamics, and in turn affect the lake water column properties which are fundamental for algal production, thermal stability and vertical mixing, along with resource availability from light and nutrient distributions within the lake. Climate has played an important role in diatom compositional changes within shallow Arctic lakes throughout the Holocene, resulting in shifts within the planktonic and benthic diatom communities as lake systems change between ice-cover and ice-free conditions.
In comparison to regional North Atlantic palaeoclimatic data, the three lake sites on the Barents Sea margin do not appear sensitive to shifts between the North Atlantic and Arctic air masses within the mid–late Holocene (specifically between c. 3970 and 6200 cal. yr BP). This is because the synchronous period of diatom change at these three lake sites are not coeval with the decline in HSG percentages and rise in SS mean sizes at c. 5130 cal. yr BP. However, there are problems with correlating these events, as sample resolution, for example, can impact record comparison. The freshwater diatom assemblage changes can be assumed to be driven by climate-modulated changes in habitat and availabilities of nutrients and light, which are controlled by the length of the growing season, aquatic macrophyte coverage and extent of seasonal ice or snow cover (Pienitz et al., 1995). OGF and OKV are the most sensitive to climate-driven ecosystem change, displaying the largest abundance shifts within both lake plankton and benthos. A more subdued diatom response is detected within LCJ sediments.
The diatom floristic shifts at c. 5270 and 5350 cal. yr BP within these three lake sites occur between two notable short-lived peaks in Pinus:Betula ratios. Pollen records from OGF and LCJ have higher Pinus:Betula ratios during the period examined, in comparison to OKV where the tree-line response to climate appears more muted. The results highlight significant differences in the responses to climatic changes between aquatic ecosystems (freshwater diatoms) and terrestrial vegetation (tree limit fluctuations). There are clear similarities between the diatom records; however, site-specific differences in the diatom response can make it more difficult to interpret a clear climate signal, as diatoms respond to a variety of environmental factors. Climatic signals from aquatic systems can be problematic, due to the complexity of climate-driven changes within high-latitude lake ecosystem functioning, with many factors affecting the diatom community composition. The diatom assemblage records from these lake sites appear to be responding to regional climatic changes and are modulated by their catchment and lake site conditions. The abrupt diatom assemblage shifts found within these lakes between c. 5270 and 5350 cal. yr BP are indicative of the onset of warm conditions. This is where Cyclotella species and large pennate diatoms (Navicula radiosa and Cymbella descripta) increase in OGF and OKV. However, pollen data suggest that the warming trend did not occur until c. 4525 cal. yr BP. This could be a result of lags in the response of trees to climatic changes, or it is possible that the diatoms within these three Arctic lakes are responding to a separate climatic event to which the vegetation was not sensitive. Thus, our results did not find synchronous responses of tree limits and diatom assemblages. Given, however, that these biological systems are experiencing and responding to different climatic or climate-related variables, such a result should perhaps not be a surprise but should encourage further research that aims to clarify the climatic variables to which each proxy are responding (Huntley, 2012).
Footnotes
Acknowledgements
We are very grateful to Dr K Rühland and one anonymous reviewer for their helpful comments and suggestions on the manuscript. We would like to thank Dr N Solovieva from ECRC, UCL, for her help with the rate-of-change analyses and Dr A Parnell from University College Dublin for his help with assessing synchronicity using Bchron. We also thank Dr S McGowan and Dr GEA Swann from University of Nottingham for their useful comments on the manuscript.
Funding
This study was undertaken as part of the MSc Quaternary Science degree at Royal Holloway University of London and University College London, with 0.5 NERC Studentship funding.