
Abstract
In this study, we investigated morphologically and geochemically a skull from an open pit situated on the Călineasa-Şesu Gârzii Plateau, Bihor Mountains. The study presents the first 14C dating and stable isotope composition of bison skeletal remains from the Romanian Carpathians. The result of 14C determination yielded two relevant maxima: one is around AD 1550 and the second around AD 1645. The data indicate that the investigated bison died during the ‘Little Ice Age’ (LIA), approximately 200 years before the last ones were hunted to extinction. Stable isotope composition of carbon from bison tooth enamel suggests that the bison diet consisted mainly of C3 grasses, compatible with a high-altitude habitat, vegetation distribution and low mobility. Oxygen isotope composition of both phosphate and carbonate groups of tooth enamel indicates that the bison drank from stagnant water sources, such as lakes or puddles formed from rain water. This further suggests low mobility of the bison as well, as the presence of such small stagnant water sources is characteristic even today for the plateau. Calculated oxygen isotope compositions of rain and drinking water at the time the bison lived indicate a lower relative humidity, around 60–70% at that period of LIA when the bison lived, which is lower than today’s humidity of 80%.
Introduction
Bison bonasus (Linnaeus, 1758) is the largest mammal living at present in Europe. Regional evidence indicates that the bison has been a consistent member of the Holocene mammalian community, although climate and habitat changes as well as hunting influenced population numbers (Benecke, 2005; Krasinska and Krasinski, 2013; Kuemmerle et al., 2012). Both morphological (Geraads, 1992; Groves, 1983) and molecular (Buntjer et al., 2002; Pitra et al., 1997) phylogenies show that the European bison, Bison bonasus, has a close relative among the Bovidae, which is the North American bison, Bison bison (Linnaeus, 1758).
The European bison is a selective feeder (Pucek, 1987). Merceron et al. (2014) determined for the present living specimen in the Białowieza Primeval Forest, Poland, high involvement in browsing. However, for the Carpathian bison, Kuemmerle et al. (2010) indicate that the animals prefer managed grasslands.
Presently, Bison bonasus may be found in the Białowieża forest stretching between Poland and Belarus as well as in sectors of Romanian, Ukrainian, Polish and Slovakian Carpathians (Geacu, 2005; Krasinska et al., 2000; Ziółkowska et al., 2012). In Romania, Bison bonasus was reintroduced in natural reserves, such as Slivuţ-Haţeg (Hunedoara District), Vânători (Neamţ District) and Bucşani (Dâmboviţa District), as well as in two zoos (Bucharest and Târgovişte; Geacu, 2005). In May 2014, 17 European bisons were released in the Ţarcu Mountains at Armeniş (Caraş-Severin District; http://www.theguardian.com/environment/2014/may/21/-sp-european-bison-europe-romania-carpathian-mountains). After spending 4 months in a 0.15-km2 quarantine enclosure, the bisons were released in a 1.6-km2 enclosure, where they will be left to fend for themselves (http://www.rewildingeurope.com/news/the-bison-herd-we-brought-to-the-southern-carpathians-have-now-taken-a-new-step-towards-life-in-the-wild). Since 2014, captive bisons are kept also in Recea-Cristur, in the vicinity of Cluj-Napoca, where the herd is maintained for commercial purposes.
For the second millennium, for the present territory of Romania, there are only few published archaeozoological descriptions of Bison bonasus skeletal remains. For example, a skull of Bison bonasus was identified and described by Haimovici and Tărăbuţă (1968) in the Muşatini Fort of Moldavia (14–15th centuries).
Von Fichtel (1791) published in a short note that bones of Bison could be found in the forests of Căliman Mountains around 1760. Beginning with the 16th century, intense hunting in the Carpathians dramatically reduced the bison population, restricting it to the northern part of the East Carpathians (Nania, 1991; Nedici, 1940; Szalay, 1916) and at high altitudes (Săhleanu, 1933). According to Szalay (1916), in the 17th century, bisons populated only Rodna, Bistriţa, Căliman and Seletin Mountains, Romania. This region of the northern part of the Eastern Carpathians was belonging to the Habsburg Monarchy at that time. Szalay discusses over 50 documents regarding bison remains or hunting expeditions. He shows that organized export of the wild bison (‘planmässigen Bisonexport’) to Vienna or Pest (today part of Budapest town) for the so-called Hetztheaters was concomitant with bison disappearance. Such theatres, where the wild animals were killed by specially trained dogs and hunters in front of the public, functioned, for example, in Vienna between 1708 and 1796 (www.ki3.at/bezirksinfo_ge_bau_19.htm), as well as in Graz or Budapest. Szalay describes that due to poor conditions, such as dark and very narrow rooms where the animals were kept next to the theatres, the rate of mortality was very high among bisons even prior to shows, and accordingly, sustained import of animals was necessary. The author concluded his study concerning the last documented occurrences of Bison bonasus in the Carpathians that the last wild animal was shot in the Căliman Mountains (Kelemen Alpe) in 1790. Furthermore, a captive bison exemplar, Miska, was surviving in the Schönbrunn Zoo until 1809. In 1796, Miska was the only large mammal which succeeded to escape from the ‘Hetztheater’ during a devastating fire. Subsequently, as the theatre was completely burned, Miska was transferred to the Schönbrunn Zoo.
Săhleanu (1933) recognizes that the estimation of Szalay (1916) concerning the last natural occurrences of Bison bonasus is close to that of Botezat (1931). According to Szalay, descriptions of bison occurrences later than 1809 are not reliable. Botezat (1931) describes a horn of Bison bonasus in a private collection, on which a fixed iron ring documents that the specimen was shot in 1808 in the Seletin Mountains from Bucovina, part of the Habsburg Monarchy at that time. For the moment, this is the most recent reliably documented bison hunt in the Carpathian area.
Besides being a target for hunting, a further reason for which the bison and Bos primigenius (Bojanus, 1827) are popular is that their figures are occurring on many types of weapons. The problem of the animal represented on the Moldavian or even Thracian weapons or coat of arms, specifically Bison bonasus versus Bos primigenius, is largely discussed by Botezat (1913), Cernovodeanu (1977) and Nania (1991), among others.
According to Săhleanu (1933), the skeletal remains of Bison bonasus are very rare in the Romanian Carpathians, even rarer to find than those of Bison priscus (Bojanus, 1827), its pre-Holocene counterpart. In this study, a skull of Bison bonasus found in a natural open pit, on the Călineasa Plateau, Bihor Mountains and Apuseni, is morphologically described. The main objectives are to date the skeleton and extract environmental and ecological information using stable isotopes in both phosphate and carbonate groups from tooth enamel. As comparative material for diet and climatic signal, we performed stable isotope analysis on a tooth belonging to a recent Ovis aries (Linnaeus, 1758), which lived at the same site.
Material and methods
Description of the location and present environment
The area where the Bison bonasus skull was found is a limestone plateau, characterized by the presence of dolines and open pits formed in Anisian limestones (Bleahu et al., 1980) and covered by vegetation consisting mainly of spruce stands and alpine herbs (Bodnariuc et al., 2002). The plateau is situated above 1000 m of sea level and is formed by three entities: Călineasa, Şesu Gârzii and Bătrâna with an approximate surface of 130 km2 (Figure 1a–c). For the plateau area, the present-day climate is predominantly influenced by west-north-west oceanic air masses and continental temperate climate. The MAT is 4°C, with an annual humidity of 81% (Birsan and Dumitrescu, 2014). The prevailingly western air flow in the Apuseni Mountains causes large precipitation amounts to fall on their western slopes with a mean of 1600 mm yr−1 at Stâna de Vale, around 30 km to the NW from the open. On the eastern slopes, the mean precipitation rates are reduced by half, below 850 mm yr−1 at Băişoara, around 35 km to the NE. At Scărişoara, 10 km distance from the investigated site, the mean annual precipitation amount varies around 1200 mm, with the highest monthly rates during the spring and early summer (Perşoiu et al., 2011).

(a) Călineasa, Bătrâna and Şesu Gârzii plateau in a Google Earth view, (b) Josephinic map (1:28:000) for the Şesu Gârzii sector of the plateau and (c, d) view of the Şesu Gârzii sector of the plateau with the location of the site.
In 1992, one of the authors (OG) was looking for new caves on the plateau area, also exploring a vertical pit. This pit is situated in an alpine pasture at 1200 m, belonging to plateau sector named Şesul Gârzii (Figure 1d). By exploring the pit, he reached a 3-m-high and 6-m-wide hall, with a smoothly descending floor. After 4 m, at the end of the hall, bones were found dispersed between rock fragments and a skull under a large boulder (Figure 2a). Comparison between the Google earth view of the plateau (Figure 1a and c) and the Josephine maps for Transylvania done between 1769 and 1773 (Hofstätter, 1989; Figure 1b) reveals that the present forest surface reduced in areal in respect to the time of mapping (Figure 1b and c).
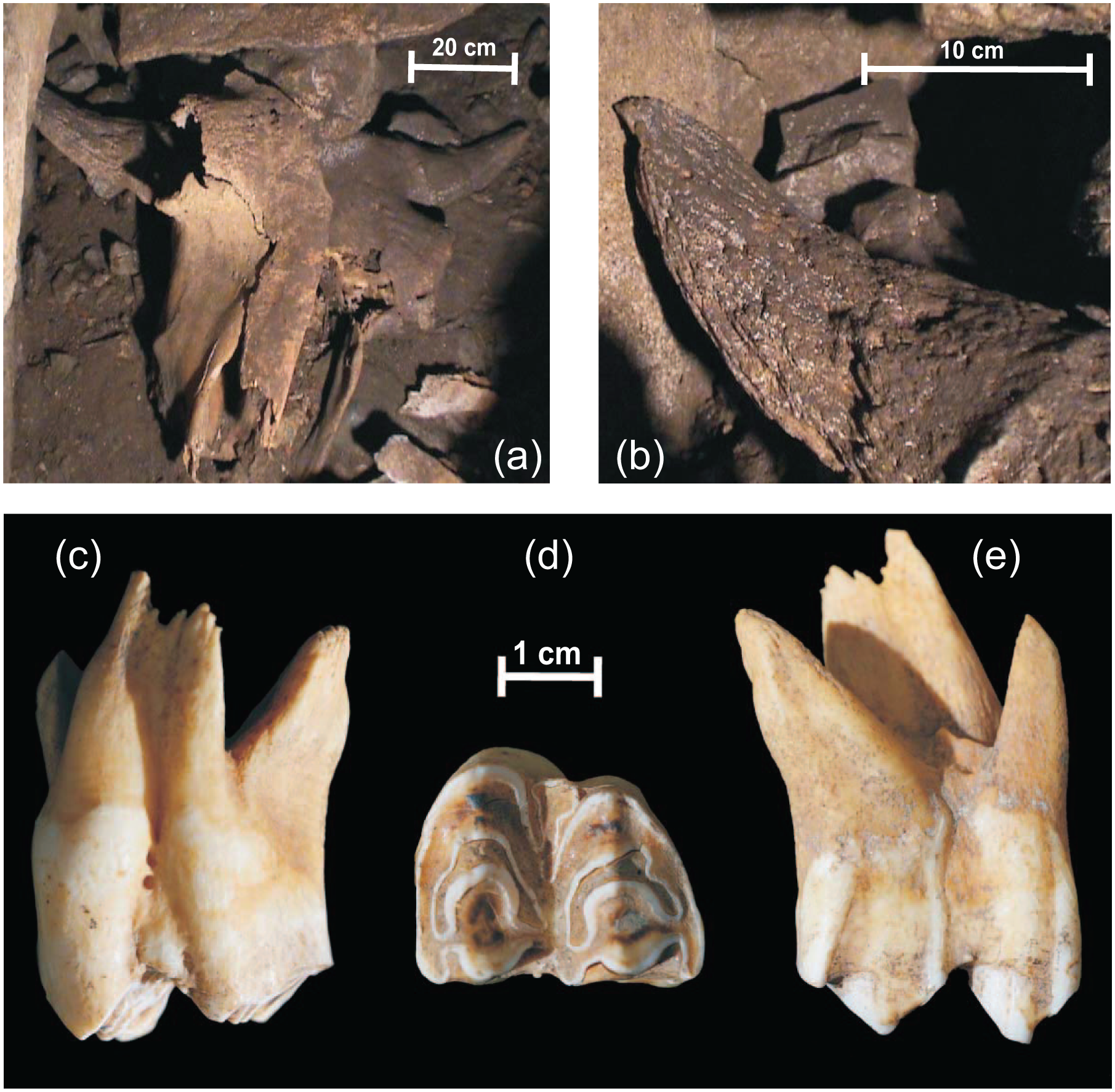
Skull and tooth of the Bison bonasus from Şesul Gârzii: (a) in situ anterodorsal view of the cranium; (b) anterodorsal view of the right horncore; and upper left third molar sampled for isotope analyses in (c) labial, (d) occlusal and (e) lingual views.
Skeleton elements
Although the nasals and the anterior part of the frontals are detached, the bovid skull from Şesul Gârzii is fairly complete, and it can be recomposed (Figure 2a). The dentition is also preserved, the upper left third molar supplying the samples employed for stable isotope analyses (Figure 2b).
The frontals are flat, without any sign of a medial protrusion. The orbits are tubular, strongly extending laterally from the skull. A deep postorbital constriction is present, clearly separating the orbits from the base of the horncores. The cornual processes (horncores), among the most relevant elements in the taxonomical assessment of fossil bovids, are fortunately well preserved, as are the frontals lying in between. The horncores expand laterally and only slightly posteriorly from the frontals. At the base, the horncores curve ventrally, and, at about mid-length, recurve dorsally, but only slightly, so that their tips remain just below the dorsal edge of the frontals. Numerous longitudinal furrows run along all sides of the horncores (Figure 2b). The cross section of the cornual processes is rounded throughout the apical and middle part of the bone and becomes slightly dorso-ventrally compressed towards the base.
Next to the morphology of the skull and, especially, that of the horncores, in order to compare the bovid from Şesul Gârzii to different Quaternary large bovids, the least distance between horncore tips (560 mm) was measured. Measurements were taken according to the procedure indicated by Von den Driesch (1976) for the Bos cranium.
The large bovids found across Europe along the Quaternary belong to a fairly small number of genera, the taxonomical diversity increasing only when various species and subspecies are taken into account. The most widespread European large bovids can be separated into three categories: (1) the early Pleistocene species of the genus Leptobos (Rütimeyer, 1868); (2) the middle and middle to upper Pleistocene Bison schoetensacki (Freundberg, 1910), Bison priscus, Ovibos moschatus (Zimmerman, 1780), Hemibos galerianus (Petronio and Sardella, 1998), Bubalus murrensis (Berckhemer, 1927) and Bos primigenius; and (3) the Holocene Bison bonasus and Bos primigenius extending from the late Pleistocene onto the Middle Ages (for the time ranges of the above-mentioned taxa, see Benecke, 2005; Kahlke, 1999; Kurtén, 2007; Masini et al., 2013; Van Vuure, 2002).
In Leptobos stenometopon (Rütimeyer, 1868), L. merlai (De Giuli, 1986) and L. furtivus (Duvernois, 1989), the horncores emerge posterolaterally and present a more complex twist around their axes: they twist laterally from the incipient emergence direction, then dorsally, and, finally medio-posteriorly (Masini et al., 2013) .In L. etruscus (Falconer, 1868) and L. vallisarni (Merla, 1949), the horncores are oriented postero-dorsally and remain roughly in the same plane with the frontals (Duvernois and Guérin, 1989; Masini et al., 2013; Merla, 1949). None of the horncore morphologies known for the early Pleistocene species of Leptobos fit the shape and the orientation seen in the bovid from Şesul Gârzii, the latter bearing horncores oriented laterally and contained in a plane almost perpendicular to the frontals.
In both Hemibos galerianus and Bubalus murrensis, the horncores are directed posteriorly (Masini et al., 2013; Schreiber and Munk, 2002), orientation that is, again, different than the one seen in the bovid from Şesul Gârzii. More than that, the cross section of the horncore base in Bubalus murrensis is more complex in shape than the subcircular outline seen in the bovid from Şesul Gârzii (e.g. Berckhemer, 1927; Schreiber and Munk, 2002). A different shape of the horncores is also seen in Ovibos moschatus, where the bases of the horncores are massive, covering most of the postorbital region of the cranium, from which they deflect ventrally and slightly anteriorly, recurving dorsally only towards the tips (Allen, 1913; Lent, 1988).
Bos primigenius, commonly known as the aurochs, one of the most common bovid in late Pleistocene fossil assemblages and also in early Holocene archaeozoological assemblages from Europe (for a review of Romanian occurrences, see Bejenaru et al., 2012) also has a very distinct shape of the horncores, markedly different from the one seen in the bovid from Şesul Gârzii: in the aurochs, the horncore bases are oriented dorso-laterally at the base, then curving antero-medially and, later on, medio-dorsally at the tips. More than that, the base of the horncores is placed more posteriorly on the skull of Bos primigenius, overhanging the occipital area (Martínez-Navarro et al., 2007), morphology not present in the bovid from Şesul Gârzii.
The horncore morphology seen in the bovid from Şesul Gârzii is consistent with the one present in various species of the genus Bison (Hamilton Smith, 1827). Indeed, in all the Eurasian species of Bison, the horncores are oriented laterally, then ventrally and, finally, dorsally. Sometimes, the tip of the horncores slightly twists anteriorly. Longitudinal furrows are always present on the surface of the horncores, and their cross section is circular or oval in the basal part, the latter shape being interpreted as a sexual dimorphic feature (e.g. Skinner and Kaisen, 1947). The general morphology of the skull is similar to the three Bison species mentioned above, but they can be differentiated by a series of morphometric parameters. When it comes to the distance measured between the tips of the horncores, the largest values are seen in the various subspecies of the steppe wisent, Bison priscus: 1126 mm for the lectotype (data from Skinner and Kaisen, 1947); 614–1120 mm, range of average values for three different subspecies from Siberia (Vasiliev, 2008). Intermediate values are recorded for the woodland bison, Bison schoetensacki: 650–870 mm (Skinner and Kaisen, 1947) and 750–870 mm (Flerov, 1969). The lowest values are recorded for the European bison, Bison bonasus: 608 mm (average of 16 measurements ranging 525–689 mm (Skinner and Kaisen, 1947); 313–552 mm for females and 450–770 mm for males (Empel, 1962; Table 5)). The value measured for the bison from Şesul Gârzii (560 mm) fits best in the range recorded for the European bison, Bison bonasus.
The third upper molar (M3) appears as roughly quadrate shaped and robust in occlusal view (Figure 2d). It measures 30 mm in length and 22 mm in width. These proportions are consistent with the molars of Leptobos, Bos and Bison, which are not as elongated antero-posteriorly as those of Ovibos or Hemibos (Gentry, 1992).
In lingual and labial views, the molar appears as less hypsodont than those of Bos primigenius or even Bos taurus (Linnaeus, 1758), a feature typical of Eurasian bisons (Martínez-Navarro et al., 2007). This condition may also be apparent in the molar from Şesul Gârzii as a consequence of a rather advance state of wear. The general morphology of the molar from Şesul Gârzii resembles that of Bubalus, but the lingually and labially protruding elements (i.e. the paracone, metacone, parastyle, mesostyle and metastyle, respectively, the protocone, metaconule and entostyle – named according to the terminology proposed by Bärmann and Rössner, 2011) are not as salient as in the latter; the labial side in particular appears as more flattened in the molar from Şesul Gârzii (Figure 2c and d). The entostyle is well developed in the molar from Şesul Gârzii. It is placed more posteriorly, closer to the metacone. The fossae are equally narrow and deep (i.e. elongated labio-lingually). The anterior fossa is not folded, whereas the posterior is constricted postero-lingually by the presence of the metaconule fold. The paracone and metacone are both well developed, slightly more in the case of the former.
The timing of tooth eruption and apparition of enamel folds on the occlusal surface as a result of tooth wear in Bison bonasus are discussed by Wegrzyn and Serwatka (1984). The M3s are shown to erupt between the 30th and 35th month of life (they are, therefore, fully erupted when the animal is 3 years old), whereas the folds in the wear figures on the occlusal surface form between the 53rd and 84th month of life (fully formed when the animal is 7 years old). The M3 from Şesul Gârzii shows an advanced degree of wear: the entostyle is extensively worn, to such an extent that only its basal part is preserved (Figure 2c), and seen in occlusal view, its apical part forms an enamel loop that merges to the labial side of the tooth, between the paracone and the metacone (Figure 2d). The remaining tooth crown measures 30 mm in height, about half of what it might have measured if it were not worn. Extrapolating from ageing techniques based on mandibular tooth wear (Frison and Reher, 1970; Haynes, 1984; mirroring the wear stage and ratio of the third upper molar described above to that of its lower counterpart), we estimate the bison from Şesul Gârzii died around the age of 12 years – the beginning of old age, considering that free-ranging European bisons passing the mark of 20 are quite rare, even among the longer-lived females (Krasinska and Krasinski, 2013). As in other bovids, before erupting, the molars of the European bison form in the crypts inside the dentary and maxillary bones, starting with the tooth apex and ending with its cervix and the roots (Brown et al., 1960; Hillson, 2005). For bison, the crown of the third molars forms in about 15 months, roughly between the 9th and the 24th month of age (Gadbury et al., 2000; Higgins and MacFadden, 2004; Niven et al., 2004). Since most births recorded for the last free-ranging population of European bison took place between April and June (Krasinska and Krasinski, 2013; Pucek, 1987), it results that their M3 molar forms between January–February in the first year of life and April–May at the end of the second year of life. Given that the unworn portion represents the basal half of the tooth, the bulk enamel sample collected from along the length of the M3 accounts for the last 7–8 months of tooth mineralization, estimated to have occurred between September–October and April–May. For most bison calves, born in April–May, weaning occurs at the beginning of or during their first winter, before the mineralization of the M3s (Gadbury et al., 2000; Higgins and MacFadden, 2004; Krasinska and Krasinski, 2013). The chemical elements that build up the M3 are, therefore, assimilated from vegetal food ingested exclusively by the young bisons, feeding by themselves.
Radiocarbon dating
The material used for radiocarbon dating was a rib from the Bison bonasus. Radiocarbon dating was performed in the Gliwice Radiocarbon Laboratory. The collagen from the bone sample was prepared according to the method described by Piotrowska and Goslar (2002). Briefly, the inorganic fraction was removed by 0.5 M HCl and the collagen extracted by heating the acidified (pH = 3.5) sample for 12 h at 80°C. The supernatant was separated by centrifugation and dried at 65°C in oven. Following steps included combustion in continuous flow of oxygen, purification and graphitization of obtained CO2 according to the protocol described by Piotrowska (2013). The accelerator mass spectrometry (AMS) measurements were performed for the sample and relevant standards at Rafter Radiocarbon Laboratory, New Zealand (Zondervan et al., 1997). The raw 14C age of the sample was calibrated with the use of OxCal v4.2.4 software, utilizing the Bayesian analysis (Bronk Ramsey, 2009) and calibration curve IntCal13 (Reimer et al., 2013).
Stable isotopes
For stable isotope investigations, two teeth, represented by a Bison bonasus M3 and a sheep molar, were prepared. The sheep remains come from a specimen, which lived recently in the same region.
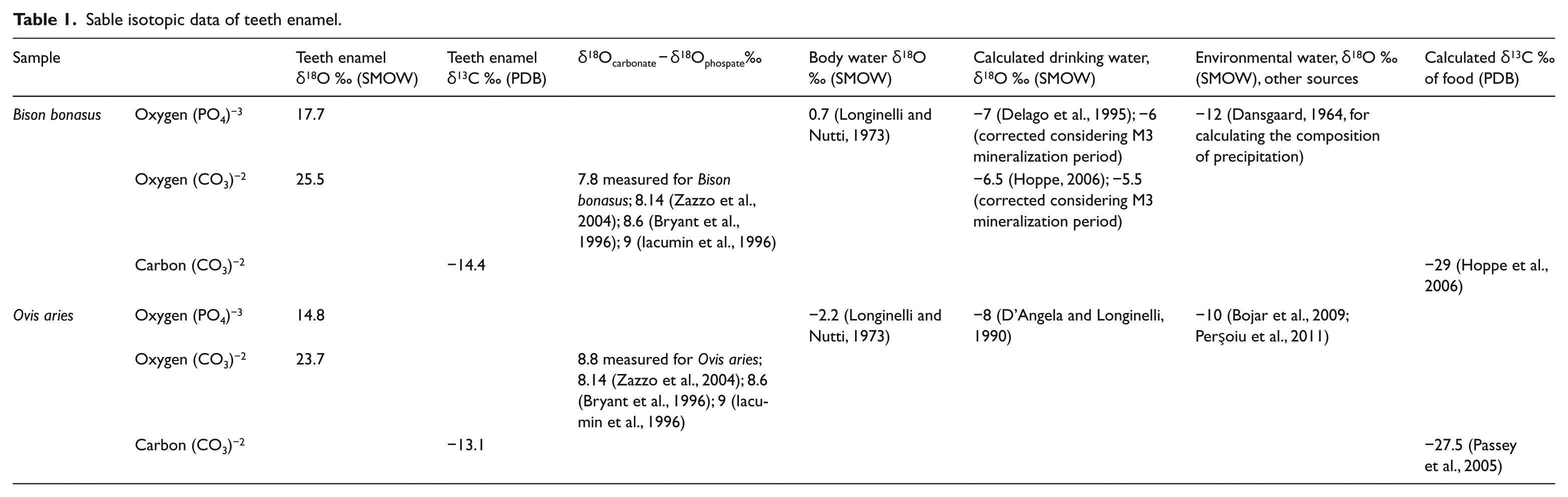
For both teeth, enamel was sampled from the entire length of the teeth. The isotopic data are given in Table 1. The stable isotope analyses were performed in the Lublin Mass Spectrometry Laboratory.
Sable isotopic data of teeth enamel.
For the investigation of the oxygen isotopic composition of the enamel phosphate, the protocol of Stephan (2000) is used. According to this procedure, the teeth phosphates are converted to the silver orthophosphate (Ag3PO4) from which oxygen is subsequently extracted for δ18O analysis. At least 40 mg of the tooth material is needed for analysis. From the powdered teeth, organic and humic substances are removed by treatment with NaOCl and NaOH followed by residues dissolution with HF. After separation of the insoluble part of the sample, the remaining solution with the phosphate fraction is neutralized with KOH solution and subsequently buffered with water solution of AgNO3, NH4NO3 and NH4OH. The solution is then gradually warmed up to 70°C. This temperature is held for 3 h and then the solution is cooled down slowly at the room temperature. During heating, due to evaporation of ammonia, the pH of the solution decreases leading to the precipitation of Ag3PO4 crystals. The obtained silver orthophosphate is then washed several times with distilled water and dried. The extraction of the oxygen from the Ag3PO4 is done using the recently developed graphite reduction method of Pelc and Hałas (2010). In the first step of oxygen extraction process, both reagents, the Ag3PO4 and graphite (Sigma Aldrich, Poznań, Poland), were roasted in vacuum at 500°C and 900°C, respectively. Then, Ag3PO4 was mixed thoroughly in agate mortar with graphite in 2:1 ratio. The quantity of about 40 mg of this mixture is loaded into a platinum boat. The boat is then installed into a glass reaction chamber and heated. To avoid a sample spillage, the reaction chamber is slowly evacuated to attain a pressure below 10−3 mbar. Then, degassing of the sample is done at 200°C over about 20 min in order to remove any volatile impurities. After degassing, the boat temperature is raised gradually to 800°C. At these conditions, the reduction reaction of silver orthophosphate with graphite proceeds for about 30 min. In the reduction reaction, the CO2 and CO gases are produced, with efficiency depending on the boat temperature. Inasmuch as CO2 gas is more convenient for the mass spectrometric analysis than CO, the CO fraction is converted to CO2 by the glow discharge enhanced by applying a pair of external neodymium magnets (Hałas et al., 2007). The CO2 collected is subsequently cryogenically transferred to the sample ampoule, which is then connected to the dual inlet system of the isotope ratio mass spectrometer with triple collector for the determination of the oxygen isotope ratio. Since the oxygen extraction in the graphite reduction of silver orthophosphate is incomplete, the obtained isotope composition has to be corrected by +0.7‰. This correction was precisely determined by the analysis of standards determined with the fluorination method that assures the complete oxygen extraction from the Ag3PO4 (Hałas et al., 2011).
Carbonate phase in the enamel was analysed simultaneously for both carbon and oxygen isotope ratios using the same as above triple Faraday cups assembly in dual inlet isotope ratio mass spectrometer. The CO2 gas was quantitatively generated from teeth samples using a manifold with McCrea (1950) apparatus connected to the inlet system. The precision of both delta values δ18O and δ13C was better than ±0.05‰.
Results and discussions
14C dating
This study presents the first 14C dating of bison skeletal remains from the Romanian Carpathians, which gives the result of 275 ± 20 BP. Due to the wiggles in calibration curve, the result of 14C determination yielded two relevant maxima of the probability distribution of calendar age (Figure 3). Even when analysing the 95.4% ranges, the maxima can be distinguished, although both are quite narrow: one is around AD 1550 (between AD 1521 and 1575, 39.5%) and the second around 1645 (between AD 1626 and 1665, 55.43%; Figure 3). This age indicates that the bison investigated in this study died during the ‘Little Ice Age’ (LIA), approximately 200 years before the last ones were hunted to extinction. Between the 15th and 19th centuries, the so-called ‘The Little Ice Age’ was documented for the Northern Hemisphere from multiple records, which indicate slightly lower regional MAT than today (Taricco et al., 2014).
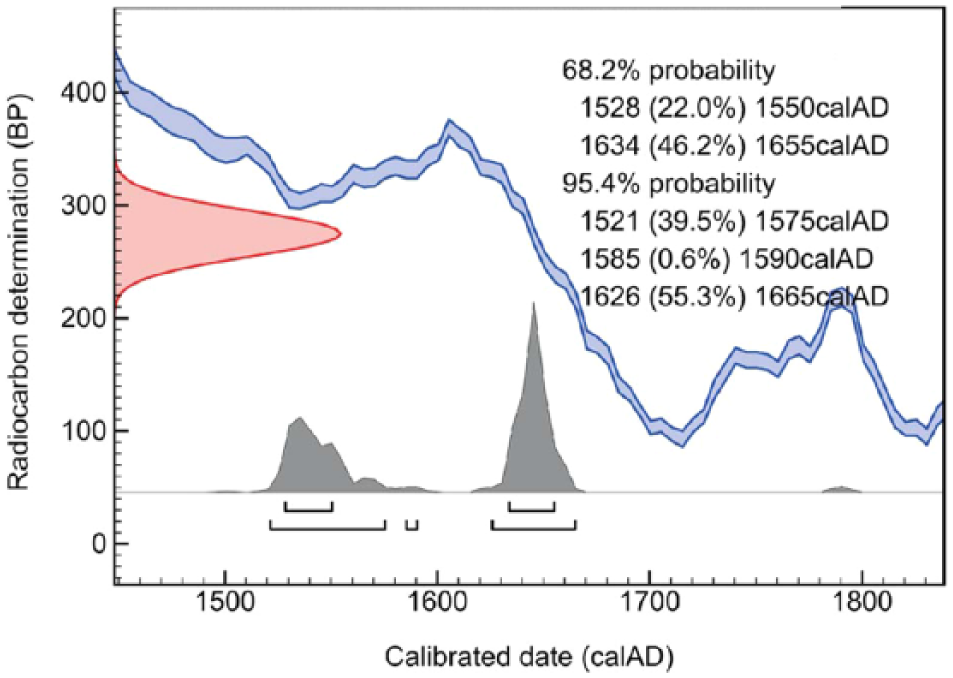
Radiocarbon dating of the Bison bonasus skeletal remains from Şesul Gârzii.
Stable isotope data
Tooth enamel, dentine and bones have inorganic or organic components in variable proportions between the different tissues. The organic component is present as collagen, a fibrous protein. The inorganic component is found as hydroxylapatite Ca5(PO4)3(OH), which also contains a small amount of carbonate substituting for phosphate and hydroxyl ions. The carbon and oxygen isotopic compositions of phosphate have long been considered a potentially useful record of climate, diet, trophic level and migration patterns. It has been demonstrated that tooth enamel is less porous than bone, contains less than 5% organic matter and has larger apatite crystals (LeGeros, 1981). For this reason, enamel is more resistant to diagenesis and suitable for isotopic investigations and, therefore, reconstructions of ancient conditions (Sharp et al., 2000).
Luz et al. (1984) developed a mass balance equation based on the fact that the isotopic composition of body water depends on the total oxygen flux through the body for which (1) the principal oxygen inputs are atmospheric O2, liquid water and food and (2) the principal outputs are oxygen in water (liquid and vapour) and CO2. So, body water is a mixture between drinking water and metabolic water produced during the oxidation of food with oxygen from the atmosphere. The equation predicts a linear relationship between the composition of body water and that of ingested water for mammals that do not obtain the majority of the ingested water from plant leaves or other highly fractionated sources. Such relationships have been demonstrated for some groups of mammals, including Bovidae (D’Angela and Longinelli, 1990; Luz and Kolodny, 1985) and also particularly for bison (Hoppe, 2006; Widga et al., 2010). Furthermore, since rain and drinking water are related to mean annual temperature (Dansgaard, 1964; Fricke and O’Neil, 1999), then the δ18O value of body water, via drinking water, is also related to the ambient temperature. Seasonal temperature variations were recorded, for example, for bison by Fricke and O’Neil (1996), Bernard et al. (2009), Widga et al. (2010) and Julien et al. (2012); seasonal amount variations by Higgins and MacFadden (2003).
Carbon isotopic composition found in mammal phosphates is mainly controlled by the proportion of C3–C4 in their diet, trophic level or spatial niche partitioning (Lee-Thorp and Van der Merwe, 1987; Longinelli and Nutti, 1973; Thorp and Sponheimer, 2005; Wang et al., 1994). Specifically and of particular interest for this study, δ13C values of enamel were determined in order to reconstruct the palaeoecology of Bison bonasus as, for example, grassland vegetation, temperatures, migratory pattern or niche partitioning (Feranec et al., 2009; Hoppe et al., 2006; Julien et al., 2012; Widga et al., 2010) or to estimate variations related to timing of apparition of M1–M3 molars and feeding patterns (Gadbury et al., 2000).
For the investigated case, the plateau area where the bison was found is characterized by the presence of alpine grasses. For the bison tooth, the measured δ13C value of −14.4‰ is much lower than, for example, these measured for Bison priscus from the southern steppe of the Eastern Europe (Julien et al., 2012) for which mean values higher than −10.3‰ were determined. Bovid enamel δ13C values are fractionated with ±14‰ relative to diet (Cerling and Harris, 1999; Passey et al., 2005; Zazzo et al., 2010). Thus, the δ13C value of the diet for the investigated bison was −29‰, in accordance with plants using a C3 type of photosynthesis (Tieszen and Boutton, 1989). The δ13C value for the M3 molar of the sheep from the same area is −13.1‰, close to the value measured for bison. The value is indicating also a C3-rich diet with a similar δ13C value of −27.5‰ (Passey et al., 2005).
Oxygen from the carbonate and phosphate groups of mammalian enamel is in isotopic equilibrium, the fractionation between δ18O values of non-altered tooth carbonate and phosphate varying from 8.1‰ (Zazzo et al., 2004) to 8.6‰ (Bryant et al., 1996) and 9‰ (Iacumin et al., 1996). For data reliability, we measured the isotopic composition of both enamel carbonate and phosphate for the M3 molar from both bison and sheep. The data indicate that the measured values are in the range of determined fractionation factors between the two anionic groups of apatite, considering the standard deviation of the present measurements as well (Table 1).
Hoppe (2006) shows that for bison, between the mean δ18O value of drinking water and the δ18O value of enamel carbonate, the fractionation equation is similar to that one derived by Huertas et al. (1995) for drinking water and enamel phosphate of feral and wild equid. The equation of Hoppe (2006) is δ18Ocarbonate = 30.06 + 0.7 × δ18Owater, whereas of Huertas et al. (1995) is δ18Ophosphate = 22.6 + 0.71 × δ18Owater. In the present case, using the equations of Huertas et al. (1995) and Hoppe (2006) and the oxygen isotope composition of phosphate in enamel, the calculated drinking water for bison is −7‰ and −6.5‰, respectively (Table 1). The preserved mineralized sector spans for less than 1 year, namely, it represents mineralization from October to May of the following. For the given latitude, this may be related to an underestimation of maximum 1‰ for the oxygen isotopic value of drinking water calculated from the phosphate and carbonate. Accordingly, the corrected values of drinking water are higher, −6‰ and −5.5‰, respectively (Table 1).
Data available up to now for the present rain water in the neighbouring Scărişoara village, situated at a similar altitude of 1165 m, have a mean δ18O values of approximately −10‰ (Bojar et al., 2009; Perşoiu et al., 2011). From literature information, the mean annual temperature differences between AD 1600 and AD 2000 can be estimated as 2°C (Climate Change: The Physical Science Basis, 2013; Taricco et al., 2014); thus, the MAT around AD 1600 at the investigated site can be approximated as 2°C. Using the relationship between the mean isotopic composition of oxygen from precipitation and MAT (Dansgaard, 1964) for AD 1600, the isotopic composition of precipitation at the time the bison lived is calculated as −12‰. This value for precipitation is much lighter than the estimated drinking water using the δ18O isotopic composition of carbonate and phosphate from bison enamel, the data indicating an enrichment of about 6‰ between the measured isotopic composition of drinking source and meteoric water (Table 1). This suggests that the bison was drinking from stagnant water sources, with inflow related to water from precipitation and subject to evaporation, such as small lakes or puddles. Using a simplified graphical approach (Gat and Bowser, 1991; Mook, 2001) and considering that between lake water and inflow water (precipitation), the calculated enrichment is of 6‰, we can estimate a total inflow to evaporation ratio of 2–3 and a palaeo-humidity between 60% and 70%. The estimated relative humidity is below the present one measured for the region of 81%, this supporting a lower relative humidity for AD 1600, at the time when the bison lived. The data suggest as well that the bison habitat was restricted to the Călineasa, Şesu Gârzii and Bătrâna plateau area, where the bison drank water from small lakes and puddles principally fed by precipitation and subject to evaporation. The formation of such water sources is enhanced on the plateau area by a general thin layer of clayey soil covering the limestones.
Using the equation of D’Angela and Longinelli (1990) for sheep, we calculate the isotopic composition from which the recent sheep drank as −8%, which matches closer to the isotopic composition of the present-day precipitation which is −10‰, suggesting lower evaporation and higher humidity in comparison with the LIA.
A further issue is why the bison skeletal remains are found today in the open pit. Bisons were herd animals and a target for human hunters. A possibility is that the skeleton of Bison bonasus found in the open pit from the Şesu Gârzii documents a procedure of hunting. For example, Canon (2004) refers to such trap strategies for northern Utah, south-eastern Idaho and south-western Wyoming, where Bison bison remains have been found. A more recent article by Carlson and Bement (2013) investigates the migration patterns of bison and associated hunting for Wyoming, Montana, the boundary to Canada and Oklahoma. In both cases, one of the practices used by a small group of hunters was to stalk an animal and bring it to a bluff edge or marsh area. Accordingly, in the present investigated case, one of the possibility of finding the bison remains in the open pit may be related to hunting practices in which the animal was brought to the edge of a pit which was not so deep, in order to recuperate later the hunted animal. The second possibility is related to accidental fall of the animal in the open pit.
Conclusion
The bison from the Şesu Gârzii open pit, Bihor Mountains, radiometrically dated around AD 1600, belongs to one of the last known free living bison from the Carpathians. About 200 years later, in the Carpathians, bisons were hunted and exported to extinction.
Stable isotope composition of carbon from bison enamel indicates that the bison’s diet consisted mainly of C3 grasses, compatible with a plateau of high-altitude habitat and low mobility. Oxygen isotope composition suggests that the bison drank from lakes subjected to evaporation. This indicates low mobility of the bison as well, as the presence of such lakes is characteristic even today for the Călineasa, Şesu Gârzii and Bătrâna plateau. Calculated oxygen isotope composition of rain and drinking water at the time the bison lived supports a lower relative humidity, around 60–70% around AD 1600, lower than today.
Footnotes
Acknowledgements
Hans-Peter Bojar, Graz, Austria, is thanked for interesting discussions regarding historical issues. Magda and Călin Drăgan, Cluj-Napoca, Romania, are thanked for local support and discussions. Valentin Dumitraşcu and Adrian Bălăşescu (Bucharest, Romania) as well as Gottfried Tichy (Salzburg, Austria) are thanked for helping with useful literature and/or various discussions concerning the topic. Two anonymous reviewers are thanked for their comments and suggestions that helped improving our article.
Funding
This work was partially financially supported by CEEPUS freemover grants 1314-73 and 1415-80539.