
Abstract
A Pliocene-age volcano on Penguin Island became active again in the Pleistocene/Holocene, forming the main cone of the island – Deacon Peak, and leaving late-Holocene phreatomagmatic craters, including ‘Petrel Crater’, about 200 m in diameter and filled currently by a maar lake with a flat bottom at 18 m water depth. Petrographic, geochemical, photosynthetic pigment, and diatom data from the 72-cm-long sediment core reveal that the crater was initially a marine lagoon with typical phytoplankton assemblages. Most probably, tectonic–volcanic activity about 1250 years ago, documented in tephra fallout, triggered an abrupt glacio-isostatic uplift that separated the lake from the sea. The horizon of tephra, probably from the Deception Island volcano, marks a sudden change in environment from brackish to lacustrine. The ecological evolution of the lake was initially constrained by an uplift, whereas the influence of marine water vanished with time, the lake became meromictic, and the freshwater mixolimnion layer expanded, while the monimolimnion became anoxic due to the influence of a penguin rookery situated on the shore. During the ‘Little Ice Age’ (LIA), the maar may have been covered by permanent ice. A discharge of mineralized guano from the possibly enlarged penguin rookery on the lake shore caused an expansion of the anoxic monimolimnion to the ice surface and an important reduction of autochthonous lacustrine biota of the maar, whereas a substantially increased participation of allochthonous biota passively supplied with guano. That record in the lake sediment core reaffirms the occurrence of a regional LIA event in the maritime Antarctic.
Introduction
Maar lakes are formed in volcanic craters, thus they typically have a well-determined watershed, often a flat floor, and a stratified water column, particularly in meromictic maars of phreatomagmatic origin. The stratigraphy of sediments at the bottom of meromictic lakes remains undisturbed because there are neither physical nor biological forces to stir them up. For this reason, sediment cores extracted from these lakes are valuable when reconstructing past environmental and climate evolution (Boehrer and Schulze, 2008; Gebhardt et al., 2011; Negendank and Zolitschka, 1993) and often provide environmental record of high resolution underlined by annual lamination that may sometimes cover very long period.
Petrel Crater maar lake is located on a little Antarctic island, beyond any influence of glacial and fluvioglacial activities that are usually leading factors responsible for the formation of lake environments in polar areas. Therefore, a direct response of sediment record to climate changes, regional glacio-isostatic uplift, and the activity of biota surrounding the lake may be expected, thereby rendering study of such issues is a key challenge in this study. The story of the lake began in a historical time (Angiel and Dąbski, 2012; Birkenmajer, 1980, 1982), after the last activity of the Penguin Island volcano, noticeable in the effects of the parasite volcanic eruption (Pańczyk and Nawrocki, 2011). A sea lagoon formed in the phreatomagmatic volcanic crater and was gradually transformed due to regional glacio-isostatic uplift into a meromictic maar lake with a well-defined stable freshwater upper layer (Rakusa-Suszczewski and Lipski, 1980).
Model of climate change with a distinctive record of ‘Little Ice Age’ (LIA) has already been well documented in the reconstruction of the temperature on the basis of ice core taken from the marginal zone of the Antarctic ice cap (Bertler et al., 2011; Domack and Mayewski, 1999; Hall, 2007; Rhodes et al., 2012; Schneider et al., 2006), in marine sediment cores (Domack et al., 2001; Hass et al., 2010; Khim et al., 2002; Leventer et al., 1996; Monien et al., 2011; Yoo et al., 2009), and as an ecological response to the LIA fixed in profiles of active and abandoned penguin rookeries (Baroni and Orombelli, 1994; Emslie and McDaniel, 2002; Emslie et al., 1998; Huang et al., 2011, 2016; Liu et al., 2005; Nie et al., 2015; Polito et al., 2002; Sun et al., 2000; Wang et al., 2007). In contrast, there is a lack of proof (Tavernier et al., 2014) or less convincing evidence (Verleyen et al., 2011) for LIA in the cores of lake sediment in the Antarctic margin zone.
The aim of this study is to reconstruct the past millennial climate development of the Western Antarctic Peninsula (WAP) region on the basis of sediment cores obtained from the maar lake located in the proximity to WAP with particular emphasis on LIA. It also refers to the question: how decadal to millennial past environmental changes (including the ‘Medieval Warm Period’ (MWP)–LIA and recent climate warming) constrained the genesis and development of ecosystems and how that history might influence the structure and function of contemporary ecosystem.
Site description
Geological setting
The Penguin Island volcano is located close to the SE coast of King George Island (Figure 1a), west of the rift axis of the Bransfield Strait. A 180-m-high basaltic stratocone built of Deacon Peak Formation (DPF) with a crater at the top occupies the central part of that volcanic island (Figure 1b). Moreover, there are at least two younger phreatomagmatic craters, one is submerged, and the other, Petrel Crater maar, is situated near the eastern shoreline of the island (Figure 1c). The geological structure and rock succession of the Penguin Island volcano have been investigated by several researchers. Birkenmajer (1982) and recently Pańczyk and Nawrocki (2011) summarized the existing data and ordered them in a new detailed stratigraphic sequence that is used in this paper, although we have expanded the succession and terminology of the youngest pyroclastic rocks on the basis of our own field observations.

Location and geological setting of Petrel Crater maar lake. (a) Sketch map of Antarctic Peninsula and the King George Island; (b) geological map of the Penguin Island volcano (after Birkenmajer, 1980, 1982; Pańczyk and Nawrocki, 2011, supplemented in the area of Petrel Crater maar lake). The location of basalt (MPF) sampling site is marked (Pańczyk and Nawrocki, 2011). It was used for geochemical comparison in the spider diagram (Figure 7); (c) Petrel Crater lake covered by ice on 1 January 2003, (d) outcrop at the SE rocky shore of Petrel Crater maar lake; (e) laminated yellow sediment of weathered basaltic tephra (DPF-WT); and (f) geologic cross section through Petrel Crater.
An outcrop on the east shore of the lake exposes a tephra covering basaltic lavas (Figure 1d–f). The lava flow belongs to the Maar Point Formation (MPF) and makes up the basement of Penguin Island. Combined whole rock 40Ar-39Ar isotopic dating yielded an age of 2.7 ± 0.2 Ma, and together with the paleomagnetic data, the middle Pliocene age (Piacenzian) is implied for the basaltic plateau (Pańczyk and Nawrocki, 2011). Loose black gravelly tephra covers the MPF. The tephra layer is up to 30-m thick and is divided in the middle by a yellow 5- to 10-m-thick layer of weathered tephra sediment of the DPF (DPF-WT). According to our field research, the lower tephra layer belongs to buried pyroclastics of the DPF (DPF-LT), and the upper tephra layer represents the youngest Petrel Crater Formation (PCF-UT). Elongated lens located between them and consisted of yellow lithified deposit with argillaceous matrix is defined as a runoff of weathered tephra suspension deposited probably in a local pool in time span between the main and phreatomagmatic eruptions. About 1000 regular laminae were counted (Figure 2D); if annual, they might indicate that this sedimentary environment lasted for centuries. Maar sediments constitute the youngest stratigraphic unit (PCF-MS).
Petrel Crater lake evolution during late-Holocene climate changes documented in environmental and ecological proxies. (A) Principal component analysis (PCA): biological and environmental factors controlling distribution of diatom groups: d1–d6. Red lines mark diatom ecological supergroups: LAG: sea-lagoon diatoms; AUTO: lacustrine autochthonous diatoms; ALLO: lacustrine allochthonous diatoms; (B) PCA: biological and environmental factors controlling pigments distribution: p1–p3; and (C) Petrel Crater lake evolution during late-Holocene climate changes documented in environmental and ecological proxies.
Radiocarbon analysis gave an apparent age of 1500 yr BP for the upper part of the DPF-WT deposit (Table 1). The correction for reservoir effect was not applied because the sedimentation proceeded in a freshwater (diatom frustules) and oligotrophic environment with a very low organic matter (OM) content (no manuring by birds). However, the lichenometric age for surface pyroclastics of DPF-LT yielded about 300 years and about 100 years for PCF-UT (Angiel and Dąbski, 2012; Birkenmajer, 1980). The inconsistency of those data with radiocarbon dating results probably from the recolonization of soil surfaces devoid of vegetation during the LIA.
Radiometric (AMS 14C) dates from maar lake sediment samples.

AMS: accelerator mass spectrometry; OM: organic matter; DPF-WT: weathered tephra sediment of the Deacon Peak Formation.
Corrected value 1963 years = calculated age for horizon 0 cm in sediment core.
Geomorphological setting
The crown of maar lake caldera is inclined northward to the sea (Figure 1c). The lake–sea barrier is at an elevation of about 10 m; however, the inner compact dam consisted of loose blocks of rock keeps the current lake’s water level at about 2 m a.s.l. (Figure 1f), and no influence of tides on the lake’s water table was observed. The direct contact of the maar with marine water had to be restricted when the relative sea level (RSL) lowered during an uplift of the land to 10–8 m above the actual sea level, which might have happened according to regional glacio-isostatic land uplift models at least 1000–1800 years ago (Bentley et al., 2005; Simms et al., 2011; Watcham, 2011). Direct evidence of high sea level is marked on the slope of the maar–sea barrier. The age of the highest recognized raised beach (8 m a.s.l.) eroding the youngest tephra (PCF-UT) postdates the phreatomagmatic eruption that formed the maar.
However, Penguin Island is situated in the marginal zone of the ice extent at the Last Glacial Maximum (John and Sugden, 1971), and a reduced rate of glacio-isostatic rebound of land might be expected in that site when compared with the central part of King George Island (see Figure 11 in Watcham et al., 2011). These considerations raise the presumption that the RSL at an elevation of 10 m on Penguin Island could even be older than the age suggested from general assumptions. The influence of marine water on the lake diminished over time, with the RSL falling from 8–10 m to 2 m recently. Below 2 m, marine water level may only have occasionally fed the maar lake during heavy storms at syzygy tides.
Properties of water column
The Petrel Crater maar lake is round with a diameter of about 200 m and has a flat floor at 18 m water depth. The lake is meromictic with a permanent stratification (Boehrer and Schulze, 2008), divided into an upper brackish water layer (mixolimnion) and a lower, dense marine water layer (monimolimnion) inherited from its geological past as a sea water body trapped in the crater and lifted up with the land by isostasy (Figure 1f).
A permanent halocline in the maar is situated at the 8- to 12-m water depth zone. A rough hydrochemical survey of the lake was carried out once during core sampling on 1 January 2003. The chemical composition of the main ions in the monimolimnion (12–18 m) is similar to that of marine water, although it is free of oxygen and saturated with hydrogen sulfide. The relative depletion of sulfate down the water column results from the active bacterial reduction of sulfates and production of sulfides in sediments, as well as from the volatilization to the atmosphere of hydrogen sulfide partially dissolved in water.
The upper water layer (0–8 m) is brackish (5–10‰). The concentration of total nitrogen and phosphorus in the mixolimnion is low, and according to the phosphorus criteria by Carlson (1977), it may be considered as oligotrophic with 0.2 mg/L Ptotal, but the content of both elements in the monimolimnion reaches very high values: 2.5 mg Ptotal/dm3 and 25 mg Ntotal/dm3 near the bottom.
Methods
Fieldwork
A 72-cm-long sediment core was taken from the central part of Petrel Crater maar lake at 18 m water depth. The bottom of the maar is even in the central part (Rakusa-Suszczewski and Lipski, 1980). Drilling was performed from the ice platform using a Livingstone sampler modified by Więckowski (1970). The sampling stopped at the compact sandy mud that was partially sampled at the base of the core. The sediment, after cleaning and description, was immediately cut into 1-cm-thick samples and stored in cold dark containers.
Water analyses
Samples in the vertical profile of water column were investigated once in the summer. Temperature and oxygen were determined by dissolved oxygen meter YSI, model 64. Water samples were collected using a Bernatovic sampler, and after filtration through a glass fiber filter (GFF), cations were determined by atomic absorption spectroscopy (AAS). Total P and N, after digestion of water, were analyzed using standard spectrophotometric methods (Grasshoff et al., 1999).
Sediment analyses
A mixture of HCl and HNO3 (1:1) was used to extract sediment samples initially ashed at 450°C. Such treatment permitted a complete decomposition of phosphates (Belopolskij et al., 1974), as well as OM, easily soluble salts and oxides, and only partial dissolution of silicates and aluminosilicates. Standard AAS procedures in acetylene–air flame were used to determine Ca (in excess of La), Fe, Mn, Sr, Zn, Cu, and spectrophotometric molybdenum blue methods for P in the extract. A Carlo Erba gas chromatograph was used to conduct analyses of organic C and N. Traces of carbonates had earlier been removed by a stream of HCl vapors passing over sample. Sand consisted of grains left on a sieve with a diameter >0.63 mm. Opal silica was extracted from sediment at constant stirring for 60 min with 1.0 M NaOH, and dissolved Si was determined by molybdenum blue spectrophotometry (Müller and Schneider, 1993), with the results recalculated to SiO2,(opal), bearing 10% H2O (Mortlock and Froelich, 1989) and is defined in figures as biogenic silica (BSi). OM was evaluated as loss on ignition at 450°C. Ca-P component of sediment, identical to hypothetical formula of apatite, was calculated from P concentrations. Mud component of sediment represents part complementary to (100% − (OM + sand + BSi + Ca-P)). Observation under scanning electron microscopy (SEM) model JEOL JSM-6380 LA 20-kV WD 10 mm was used for petrologic investigations. X-ray analysis was applied for mineral identification.
Radiocarbon dating
The stratigraphy of sediment core is basically provided by three radiocarbon dating (Table 1). The age (years) of surface horizon at 0 cm sediment depth (x) was calculated from equation y = 0.2004x2 + 87,604x based on apparent radiocarbon dating, and the value was used (1963 years) as a modern reservoir effect to down-core age correction, according to a convention proposed by Andrews et al. (1999). According to the presented results, deposition of the lacustrine sediment started 1247 yr BP at a depth of 60 cm, and the rate of sedimentation (expressed in mm/yr) has steadily increased upward the sediment core due to higher water and OM content (lower density).
Diatoms
Approximately 1 g of wet sediment was dropped into a 50-mL glass with distilled water. After 24 h, the sediment was mixed, and after 10 s, the suspension containing diatom valves was taken by pipette from the middle part of the water column and then placed on a square glass 18 mm × 18 mm in size. Slides were prepared with a mounting medium of R.I. 1.69. The diatom valves were counted along vertical rows up to 300 specimens for each sample. The amount of the total diatom concentration was calculated by a quantitative method from Jouesè et al. (1969). The habitat preferences of diatoms (pH, salinity, trophy, and moisture) were based mainly on Van Dam et al. (1994). The taxonomy used is consistent with the papers of Krammer and Lange-Bertalot (1986, 1988, 1991), Kellogg et al. (1980), Schmidt et al. (1990), Bukthiyarova and Round (1996), Sabbe et al. (2003), and Cremer et al. (2004).
Photosynthetic pigments
For pigment analysis, samples were extracted ultrasonically using acetone (2 min at 10 W; Omni-Ruptor 250) while kept on ice (van Leeuwe et al., 2006). This operation was repeated until the supernatant was colorless (up to three times). The supernatants from the extractions were combined in a 6-mL flask and filled to a mark and then filtered using 0.2-mm Millipore syringe filters. Pigments were identified using reversed-phase high-performance liquid chromatography (Dionex liquid chromatograph provided with a ultraviolet–visible (UV–VIS) diode array detector). A highly selective Waters Spherisorb C18ODS2 chromatographic column was used to separate the pigments. The following carrier phases were used in the gradient: a mixture of solvent A (80:20 methanol:0.5 M ammonium acetate), B (90:10 acetonitrile:water), and C (ethyl acetate). The gradient elution was as follows: 0 min 100% A, 4 min 100% B, 18 min 20% B, 80% C, 21 min 100% B, and 24–29 min 100% A (Wright et al., 1991). Pigments were identified on the basis of their retention times and electronic spectra of standards (DHI products) and through literature data (Jeffrey et al., 1997; Roy et al., 2011). Calibration curves were made using external standards.
Statistical analyses
TiLIA software packages were used to construct diatom and pigment diagrams (Grimm, 1990). A chemical zonation of the sediment diatom and pigment spectra was facilitated using CONISS, which is included in the TiLIA software package. A principal component analysis (PCA) was used to relate biological data (diatom and pigments) to abiotic environmental factors (sand, OM, calcium phosphate, and biogenic silica), and a Spearman’s rank correlation coefficient was used to measure the statistical relationships between the observed data values (Statistica version 8).
Results
Morphology of core
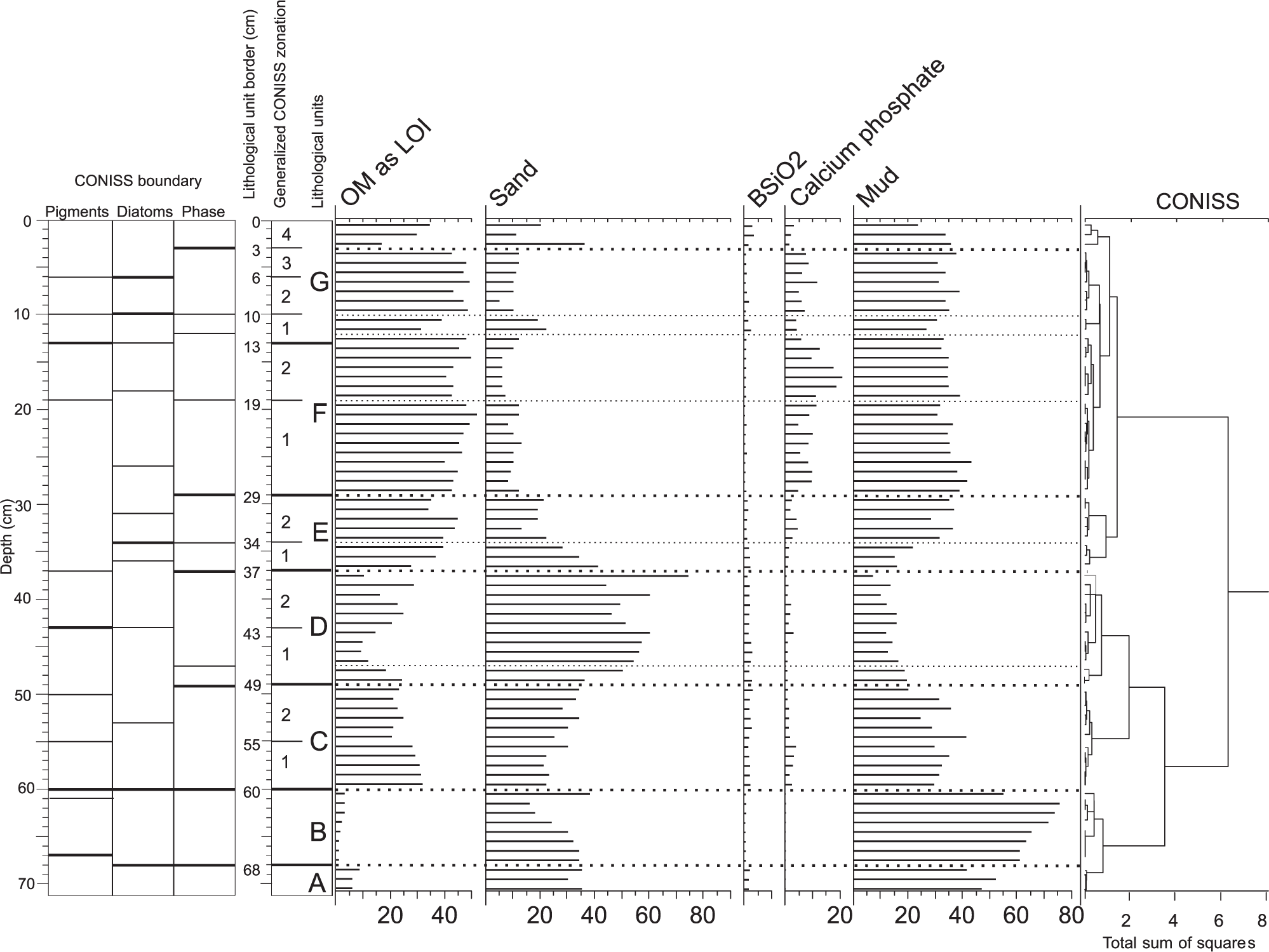
During an initial examination of fresh sediment in the core, only a lighter clastic (units A and B) and a black lacustrine muddy sapropel above (units from C to G) can be distinguished. A phase and chemical composition of the sediments, supported by diatom and pigment analyses, resulted in a more precise statistical subdivision of the core based on the CONISS program (Figure 2, Table 2). The boundaries between the units and subunits were not always perfectly synchronized among various analyses; however, generalization was necessary to make the description clearer.
Lithology and stratigraphy of Petrel Crater lake sediment core.

Lithology
The sediment core begins with sandy mud (unit A) deposited in a marine/estuarine environment (Figure 3). This loose sediment was hardly retained at the end of the corer. The real thickness of this sediment is unknown because the corer stopped due to the high resistance of the sediment. The marine/lacustrine boundary is represented by a well-consolidated 8-cm-thick silt layer (unit B). Lacustrine sediments (60 cm thick) grade from sandy (unit C) to muddy and finally to organic sapropel at the top (unit G), with enhanced concentration of penguin guano elements and the lowest C/N and Fe/Mn ratios.

Chemical composition of sediments (HCl plus HNO3 1:1 extract).
Calcium phosphate is the main component of mineralized guano in penguin rookeries. Enhanced concentrations of P and Ca occur also in the whole sediment core (Figure 3). Both elements are highly correlated (R2 = 0.8463), the Ca/P ratio equals >3 at the P content >1%, and >4 at the P content <1%. It is generally higher than in the theoretical formula of apatite (2.15), because certain amount of Ca belongs probably to Ca-plagioclases that are common in basaltic tephra and partially soluble during the extraction procedure. x-Ray sediment tests show that calcium phosphate is amorphous, unlike in ornithogenic soil where it transforms into fluorapatite after aging (Tatur and Keck, 1990).
Ornithogenic Ca-phosphate also contains elevated concentrations of several trace elements that share a geochemical affinity to Ca in fluorapatite, including Sr, Cu, and Zn determined during this study (Figure 3). We found that the correlation between Sr and P (R2 = 0.897) is even better than between Ca and P. Moreover, the linear regression of the Sr/P relationship crosses the axes at low level (19 ppm of Sr), which means that its content in the nonguano components of the sediment is relatively negligible. Thus, elevated concentrations of P and Sr in the sediment may be considered as a sensitive and specific proxy of ornithogenic ‘apatite’ content.
Phosphorus concentrations in sapropel of unit F2 ranged between 1% and 3% (Figure 3) that equals up to 20% of apatite indicating a high participation of penguin guano in sediment. Guano contains from 2.5% (fresh) to 12% (altered) of P (Tatur, 1989; Tatur and Myrcha, 1984; Tatur et al., 1997). The surface deposit of ornithogenic soils may be composed finally of almost pure calcium phosphate identified as fluorapatite containing (in theoretical formula) 14% of phosphorus (Tatur and Keck, 1990). Discharge of protein-rich guano to the lake reduces the C/N ratio of sediment (about 10, sections F and G), which is lower than in the rest of the core (between 13 and 20).
Clastics in the lacustrine sediments (units C–G, 0–60 cm) are composed of black porous tephra fragments that are identical to the local tephra of the Penguin Island volcano (DPF and/or PCF). The sand fraction in muddy sediments, bearing marine microfossils, and marine pigments at the base of the core (unit A, 68–72 cm), is lighter, usually devoid of porosity, and shows higher petrographic variability that is typical of sediments of the Bransfield Strait.
However, the most intriguing is the origin of the biologically barren loose silt from the lacustrine/marine boundary (unit B 60–68 cm). Silty sand in unit A is well sorted and consists of homogeneous vitro and litho pyroclasts, including single sharp-edged feldspar grains (Figure 4a). This sediment, identified as tephra, by its mineral and chemical composition, is clearly erratic to the Penguin Island volcano.

Scanning microscope images of Petrel Crater maar sediments: (a) tephra fallout horizon from the Deception Island volcano (unit B), (b) pyrite framboids scattered in sediments, particularly abundant in sapropel (unit F), and (c) presumed annual lamination, scanning microscope image (unit F). Dark (winter) lamina is rich in organic matter (c), while light (summer) lamina is enriched with higher calcium phosphate (Ca and P). Possible is iron sulfide and/or gypsum (high S content plus diffraction data). Surface efflorescence of sea salts (Cl) at the background in both laminae, pure halite at point 3. (d) Locally concentrated (denser) lamination due to thinner light laminae, scanning microscope image (unit F). Chemical composition of laminae comparable to that from Figure 6c. Minor differences: high Si content at a high Si/Al ratio in dark lamina suggests the presence of diatoms at the research point (material is free of quartz). Light lamina is poorest in organic matter. Both laminae contain traces of chlorides on the surface.
Pyrite framboids
Pyrite framboids commonly occur in lacustrine sediments of the investigated meromictic maar (Figure 4b), proving the presence of an anoxic environment at the bottom since the beginning of the lake formation. The highest concentration of framboids in sapropel results from a higher population of sulfate-reductive bacteria feeding on abundant guano residues. The distinctive anoxic environment is confirmed by the low Mn/Fe ratio (Figure 3). The size of the framboids is similar along the core, not exceeding 5 µm in diameter.
Annual lamination
Traces of regular lamination are visible in a few sections along the core, mainly in the upper part (units F and G), where the rate of sedimentation measured in the field ranged from 1.0 to 1.5 mm for a pair of laminae per year. This rate of sedimentation is slightly higher than the rate of annual sedimentation calculated from radiocarbon data (from 0.8 mm/yr at 29 cm sediment depth to 1.2 mm/yr at the sediment surface).
Regular lamination is well visible under the electron microscope (Figure 4c), and several microprobe chemical analyses explain its nature. Wider and lighter laminae contain more amorphous calcium phosphate and usually also clastics but less OM. Thinner and darker laminae contain more OM and a lesser amount of other components. However, dark laminae can often be combined in a densely laminated cluster with a highly reduced number of light laminae (Figure 4d). This phenomenon has to increase the rate of sedimentation calculated from lamination. Clastics and calcium phosphate–rich light laminae may be composed largely of allochthonous material due to erosion processes during the breeding period in summer. OM-rich laminae are probably supported by blooming of algae during the end of summer or possibly in winter under limited erosion process.
Diatoms
The diatom assemblages identified in the sediment core of Petrel Crater lake consist of low-diversity benthic diatoms belonging to 30 species. Planktonic diatoms have not been found. Marine species were rare and usually appeared as fragments of broken valves, indicating sediment reworking or eolian transport. The benthic diatom flora of Petrel Crater lake is represented by brackish, brackish-freshwater, and freshwater-brackish taxa (Figure 5).

Diatom stratigraphy of the more frequent species in the lake sediment core with a zonation based on the CONISS cluster analysis (right side).
At the base of the core (unit A), mainly brackish water diatoms are noted: Craspedostauros laevissimus West & West, Tryblionella marginulata (Grunow) D.G.Mann, and Achnanthes brevipes (Kützing) Cleve, and brackish freshwater ones: Nitzschia frustulum (Kützing) Grunow, Craticula molesta (Krasske) Lange-Bertalot & Willmann, and Hantzschia virgata (Roper) Grunow. Diatoms were completely absent in unit B. In unit C, typical brackish diatoms disappeared, and the freshwater diatoms became more abundant. This sediment section is dominated by Planothidium delicatulum, N. frustulum, Diadesmis contenta (Grunow) D.G.Mann, Stauroneis anceps Ehrenberg, and Planothidium renei Lange-Bertalot & Schmidt. Units C and D are characterized by the largest amount of diatoms. In units E and F, the total abundance of diatoms was low and ranged between 63 and 200 million/1 g of sediment. P. delicatulum dominated with accompanying Psammothidium metakryophilum Lange-Bertalot & Schmidt) Sabbe, Pinnularia microstauron (Ehrenberg) Cleve, and Luticola muticopsis (Van Heurck) D.G.Mann. In the interval of 10–16 cm, abrupt changes in diatom quantity associated with a high increase in diatom frequency (up to 720 million/1 g of sediment) were observed. The diatom assemblage was dominated by P. delicatulum (up to 92%). Decreasing diatom abundance was found at a depth of 6–10 cm. The lake’s freshwater stage is marked by the dominance of freshwater species Psammothidium metakryophilum (up to 79%). Great concentrations of diatoms were observed at the top of the core (0–6 cm).
Pigments
The results revealed a vertical fluctuation of pigment distribution caused by changes in the composition of algal communities in the lake evolution. A total of 18 photosynthetic pigments have been identified along the sediment core, which belong to chlorophyll (chlorophyll a, b, and c and bacteriochlorophyll a and b), along with their derivatives (pheophytin a, pheophorbide a, and pyropheophorbide a) and diverse carotenoids (Figure 6). The presence of chlorophyll and bacteriochlorophyll indicates that the community of primary producers comprised both oxygenic and anoxygenic photoautotrophs (Squier et al., 2002, 2005).
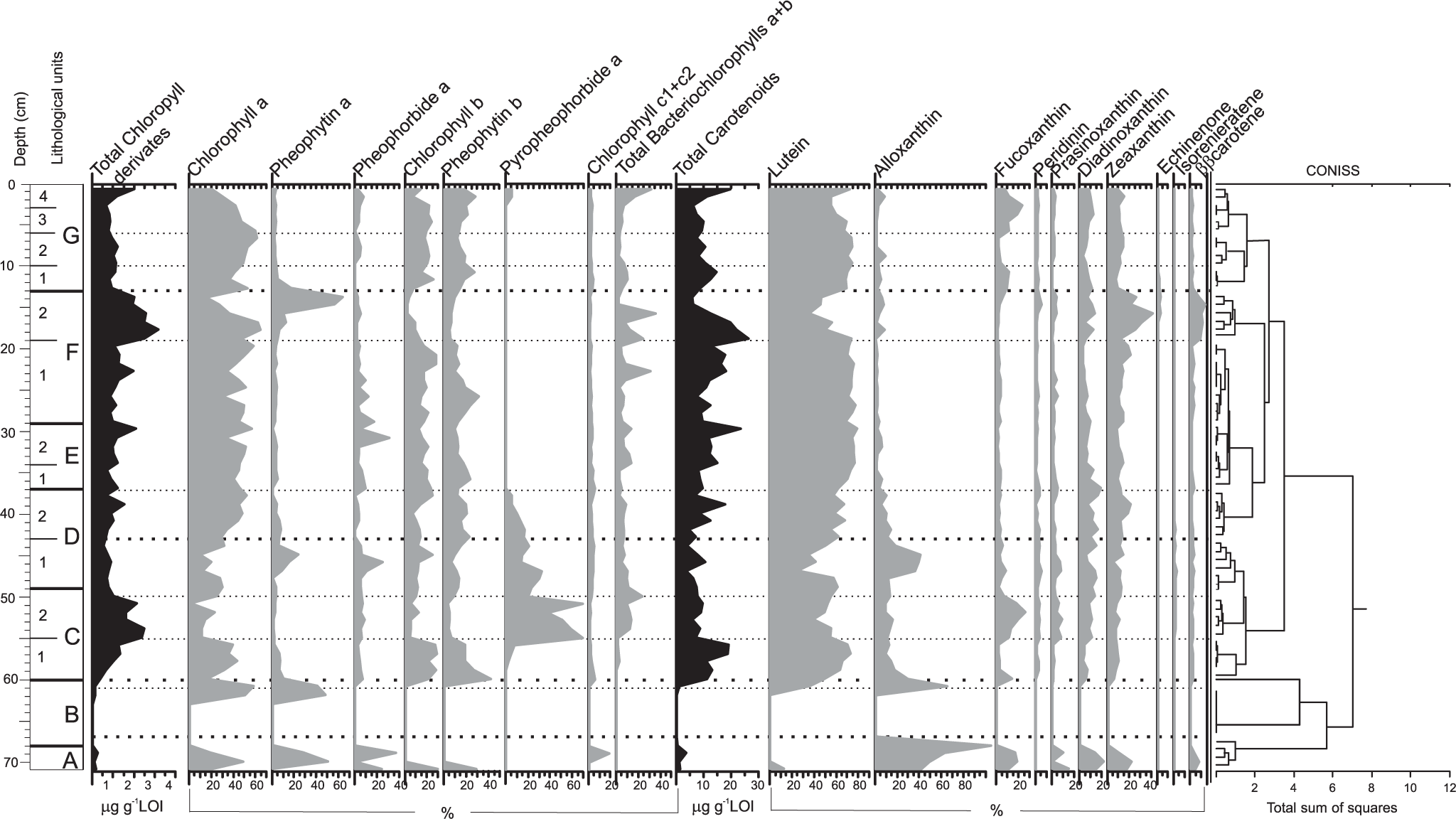
Pigment stratigraphy in the lake sediment core, with a zonation based on the CONISS cluster analysis (right side).
Alloxanthin is a dominant pigment in unit A, indicating a great contribution of cryptophytes to the algal community, complemented by diatoms and/or chrysophytes (fucoxanthin), prasinophytes (prasinoxanthin), and cyanobacteria (zeaxanthin). This unit is characterized by low concentration of chlorophyll a but relatively high concentration of its derivatives. The pigment signals were not detected in unit B, but in unit C, the total concentration of chlorophyll is high, indicating the increase in primary production. The great proportion of lutein along with chlorophyll b and pheophytin b suggests the dominance of OM derived from green algae. The increasing proportion of bacteriochlorophyll (sulfur bacteria), pyropheophorbide (grazing pressure), fucoxanthin (diatoms and chrysophytes), and peridinin (dinoflagellates) was noted in the upper part of this unit (C2 53–49 cm). The same composition of algal assemblages was observed in unit D, but the total algal biomass was lower than that in unit C. The low total algal biomass was also found in units E and F. The higher concentration of bacteriochlorophyll a and b in this part of the core suggests anoxic condition. The carotenoid composition revealed predominance of chlorophytes (lutein) in deposited OM and low proportion of other algal groups. In the upper part of unit F (19–13 cm), the chlorophyll concentration rapidly increased, and at a depth of 14–13 cm, great amount of pheophytin a was detected. Green algae were still the dominant group but with conspicuous subdominance of cyanobacteria. Unit G represents the modern part of lake history characterized by contribution of green algae (as a dominant component), diatoms and/or chrysophytes, cyanobacteria, dinoflagellates, cryptophytes, prasinophytes, and sulfur bacteria in the autotroph assemblage.
Discussion
Stratigraphy and chronological evidence
There are several explanations for frequently noted erroneous old 14C dating of modern Antarctic materials (e.g. Björck et al., 1991a, 1991b; Ingólfsson, 2004). Surface sediments in Antarctica commonly yield ages >2000 years (Hall, 2009, and references therein), that is at least twice more than the correction value regularly applied as a marine reservoir effect ranged from 750 to 1200 years (after Ingólfsson, 2004). The age calculated for surface sediments of the investigated maar lake equals 1997 years and is similar to the modern reservoir effect adopted, for example, by Melles et al. (1997) at an epishelf lake in the Bunger Hills, East Antarctica, and to the correction value for marine sediments of the Bransfield Strait, applied by Kyu-Cheul Yoo et al. (2009).
Some properties of the lake explain the relatively high value for radiocarbon correction. The stable stratification of water column in meromictic maar maintains relic marine water that was trapped in the monimolimnion at the beginning of the lake formation. Moreover, there is a regular supply of OM from current and relic ornithogenic soils that occur along the north shore of the maar. Thus, primary production in the maar may have used old carbon sources. The recycling of old carbon in the closed environment of the stratified lake made it increasingly older (Hendy and Hall, 2006; Squyres et al., 1991), particularly if initial carbon sources originated during pre-bomb time with importantly higher correction values (Berkman and Forman, 1996).
The accepted radiocarbon correction contains a risky and simplified assumption that biogeochemical relations in lake remained constant through time (Hall, 2009). Therefore, the use of supplementary checking methods of dating are recommended (Doran et al., 1999). Unfortunately, the concentrations of 210Pb and 137Cs most suitable for that operation occurred at very low levels in the sediments and do not appear to be viable dating techniques in the investigated case. Doran et al. (1999) also came to similar negative results. Radiocarbon stratigraphy of the maar lake after tephra fallout about 1247 yr BP may be roughly verified by the other chronological proofs:
Lamination (most probably annual) preserved in some sections of the core is well documented under the scanning microscope. The rate of sedimentation estimated from the lamination remains roughly consistent with that calculated from radiocarbon data.
Geomorphological setting constrained by glacio-isostatic uplift of land indicates that direct contact of the marine lagoon with the ocean was blocked and lacustrine sedimentation in the maar started when the RSL lowered during uplift to 10–8 m a.s.l. This may have happened according to regional uplift models about 1000–1800 years ago.
The tephra horizon at the base of the lacustrine sediments correlates with an important horizon in the regional tephrochronological pattern (horizon A-5 of Björck et al., 1991c) dated to 1250–1400 yr BP.
Geological setting of DPF-WT with an age of 1500 yr BP (uncorrected) predated the Petrel Crater lake formation.
Unfortunately, presented dates show a high degree of uncertainty; however, taken together, they form a coherent geological scenario that seems to support the reliability of radiocarbon stratigraphy.
Origin of the lacustrine environment following the tectonic/volcanic event about 1247 years ago
Geochemical composition of the tephra horizon (unit B) at the base of lacustrine sediments has a geochemical affinity to rocks and pyroclastics occurring on the Deception Island volcano about 160 km distant (Figure 7a and b). The Deception Island shield volcano is situated close to the spreading axis of the Bransfield Strait rift and is known as the largest and most active in the region during historic times. The Penguin Island volcano lies off the axis of the Bransfield Strait rift and was supported by a different magma than the Deception Island volcano. The geochemical discrepancy between the Deception and Penguin volcanoes is particularly pronounced not only in the concentration of heavy rare earth elements (HREEs; see Eu-Lu) but also in the much lower content of Ni, Cr, and Mg in the rock of the Deception Island volcano (Tatur et al., 1999). Moreover, using a ‘Decision Tree’ constructed on the basis of geochemical ratios of trace and rare elements by Kraus et al. (2013), the origin of the considered maar tephra horizon can be distinguished from possible volcanic sources other than Deception Island. The tephra horizon in the maar lake overlies the marine sediments. The chemical composition of the silty sediments of the Bransfield Strait is also similar to both that of the tephra from Deception Island (Jae Il Lee et al., 2005) and of the maar tephra horizon, except for the clearly higher content of large-ion lithophile elements (LILE; see Rb, Ba, Th, and U), which is typical of the sedimentary environment.
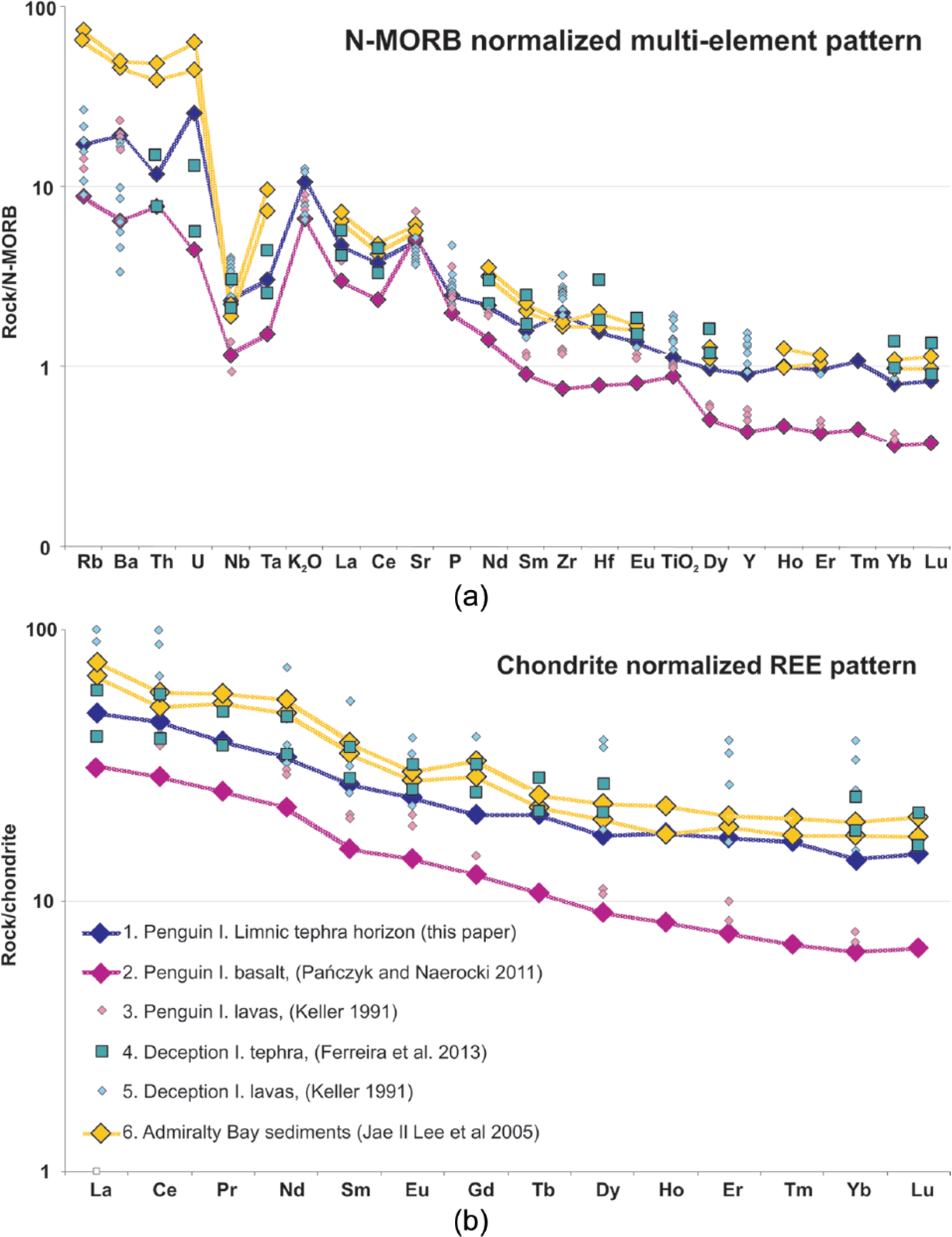
Spider diagram. The maar tephra horizon 60–68 against the tephra from Penguin Island, Deception Island, and Admiralty Bay sediments: (a) N-MORB-normalized multi-element patterns and (b) chondrite-normalized REE patterns. The normalization values for MORB are taken from Sun and McDonough (1989) and for CHONDRITE from McDonough and Sun (1995). Detailed description of compared samples.
The tephra horizon in the Petrel Crater maar (1247 years old) might eventually be correlated with the relatively pronounced 29-cm horizon recognized in lake sediments in Fildes Peninsula, King George Island (Tatur et al., 1999, 2004) and possibly the same horizon named A5 found in the sediments in other lakes of the South Shetlands by Björck et al. (1991c), Hodgson et al. (1998), and Molén (2012) and dated at 1250–1400 yr BP (at a calibration value 700 years). A possibly comparable tephra horizon was recognized in the sediment core of the Bransfield Strait, level 1, by Fretzdorff and Smellie (2002) and signalized earlier by Tatur and Del Valle (1986) and Mäusbacher et al. (1989) in lakes and by Matthies et al. (1990) in marine sediments.
The tephra horizon of Petrel Crater maar sediments sharply separates the lower marine-brackish environment from the upper lacustrine one. This abrupt change resulted most probably from a sudden uplift and suggests that the period of volcanic activity in the region might have resulted from tectonic correction of the isostatic process, the rate of which varied along the Antarctic Peninsula (compare the data of Bentley et al., 2005; Roberts et al., 2009; Watcham et al., 2011). The rate of post-glacier rebound of land in the marginal zones of the South Shetland Plate was slower than in the central part, which creates tension released by discontinuous deformations (Fretwell et al., 2010).
Relationship between diatom composition and core lithology
A significant relationship between the composition of diatoms and the lithology of the sediment was documented by PCA which has identified six groups of diatoms (Figure 8a).
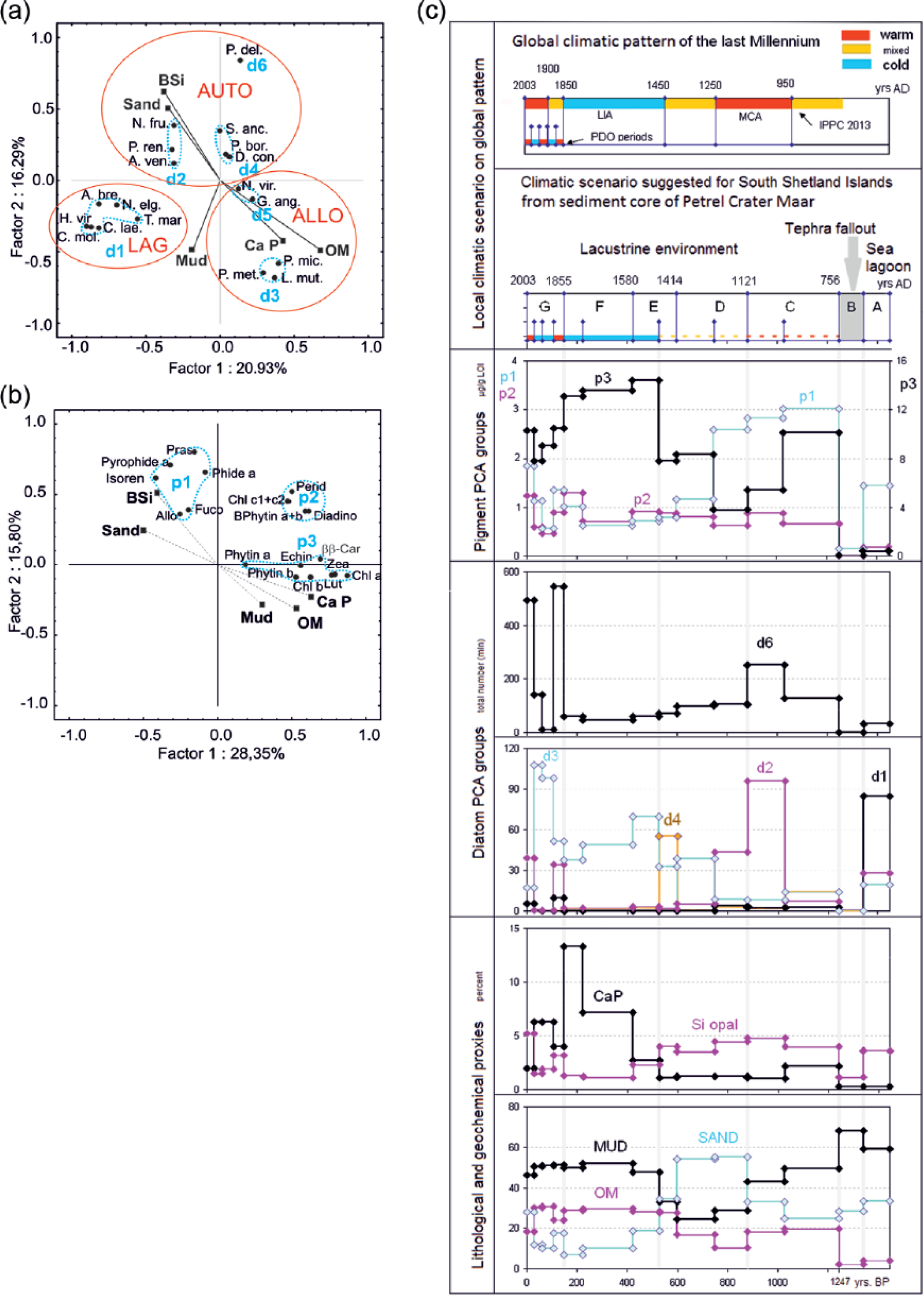
Petrel Crater lake evolution during late-Holocene climate changes documented in environmental and ecological proxies. (a) Principal component analysis (PCA): biological and environmental factors controlling distribution of diatom groups: d1–d6. Red lines mark diatom ecological supergroups: LAG: sea-lagoon diatoms; AUTO: lacustrine autochthonous diatoms; ALLO: lacustrine allochthonous diatoms. (b) PCA: biological and environmental factors controlling pigments distribution: p1–p3. (c) Petrel Crater lake evolution during late-Holocene climate changes documented in environmental and ecological proxies.
Sea-lagoon diatoms (PCA group d1)
It seems to be the most distinct and coherent group in the species composition. It is affiliated to mud sediments which are poor in OM and phosphorus and comparable with open-sea sediments (Figure 8a). The assemblage is dominated by brackish-water and brackish-freshwater diatoms. C. laevissimus requires a salinity range of 10–15‰ for optimal growth and does not tolerate freshwater or marine conditions (Watanuki, 1979). The cosmopolitan species T. marginulata occurs along the sea coast (Krammer and Lange-Bertalot, 1986), and it is considered by Verleyen et al. (2004) to be a taxon typical of the transition zones between marine and lacustrine environments. H. virgata is also a cosmopolitan littoral species of sea coasts of a wide range of tolerance to the salinity 11.5–130.8‰ (Roberts and McMinn, 1998) and high resistance to changes in osmotic pressure (Cholnoky, 1968). Craticula molesta is reported by Hodgson et al. (1998) only from slightly brackish lakes, but according to Sabbe et al. (2003), it is common in both freshwater and brackish lakes in the Larsemann Hills.
This assemblage is specific to unit A of the core. It seems that Petrel Crater lake was initially a lagoon fed by marine water and open-sea sediments. The sea-lagoon environment is separated from a younger lacustrine ecosystem by a barren tephra horizon (unit B).
Lacustrine autochthonous diatoms (PCA groups d2, d4, and d6)
P. delicatulum forming a solo d6 PCA group (Figure 8a) is a dominant species among diatoms in the lacustrine section of the sediment core. P. delicatulum is classified as a mesohalobous, alkaliphilous species preferring water with a high nutrient concentration (Saunders et al., 2009; Schmidt et al., 1990) and according to Sterken et al. (2012) associated with sea spray. P. delicatulum is a dominant also in PCA group d2 (typical for the sandy sections C2, D1, G1, and G4) but with an important co-occurrence of other diatoms (Figure 8a). These diatoms show diversified ecological preferences, although all of them may be classified as brackish freshwater. N. frustulum and P. renei usually appear in high abundance in freshwater found mainly in circumneutral habitats (Björck et al., 1993; Reimer, 1954; Schmidt et al., 1990). Amphora veneta has been found in eutrophic and freshwater environments (the optimal water salinity for this species is 6.7‰; Roberts and McMinn, 1998). Diatoms of PCA group d4 stand out alternatively with those of group d2 in the middle part of the core (units C1 and E1). They comprise freshwater-brackish or brackish-freshwater species: D. contenta, S. anceps, and Pinnularia borealis. These are alkalibiontic diatoms classified as cosmopolitan and littoral, usually found in eutrophic conditions (Cleve-Euler, 1951, 1955; Krammer and Lange-Bertalot, 1986). S. anceps is a brackish-freshwater species that thrives in water with a salinity range of 0.2–9.7‰ (Roberts and McMinn, 1998). The enhanced content of S. anceps observed in the lower part of unit C may be related to the underlying tephra horizon since this species was reported from the Midge Lake (Antarctica Peninsula) sediments, from the tephra layer (Björck et al., 1991a). S. anceps and D. contenta are also known to be aerophilic species.
Lacustrine allochthonous diatoms (PCA groups d3 and d5)
The number of lacustrine autochthonous diatoms is reduced by an order of magnitude in horizons of sediments with a high phosphorus concentration (up to 13% of calcium phosphate), which is equal to over 50% of guano (Tatur, 1989). Phosphates in sediment come from the penguin rookery located on the lake shore (about 451 breeding pairs in season 1980/1981; Jabłoński, 1984); these numbers were confirmed in the 2007/2008 season by Krzysztof Banach (field report; Figure 1b). The concentration of phosphate proves that in recent centuries, the lake water was periodically enriched by guano leachates and suspensions washed from the rookery. Mineralized guano constituted one of the main components changing the trophy of the lake and the chemical properties of sediment: low C/N ratio because of its high protein content or low Fe/Mn ratio due to intense microbial reduction (Figure 3). The supply of guano to the lake promotes diatoms of PCA group d3. The absolute frequency of group d3 diatoms increased several times and became relatively subdominant or dominant in units E2, F1, F2, G2, and G3. Group d3 includes Psammothidium metakryophilum, Pinnularia microstauron, and L. muticopsis (Figure 8a). L. muticopsis and Pinnularia microstauron are common in Antarctic terrestrial habitats (Broady, 1996; Kawecka and Olech, 1993) and have been recorded in the marginal zone of ornithogenic soils (Garraza et al., 2011; Mataloni and Tell, 2002). Psammothidium metakryophilum also origins from the soil in the proximity of the lake, although it usually occurs in lakes with very low salinity <1‰ (Gibson et al., 2006). Similarly, L. muticopsis is known as an aerophilic species typically found in freshwater lakes with increased salinity (up to 5–10‰) caused by sea spray (Karasawa and Fukushima, 1977; Van de Vijver and Beyens, 1997; Watanuki, 1979). The examples given above strongly suggest that these diatoms usually occur in freshwater environments and form a low-frequency background along the entire core. However, passive transport with guano enhances their numbers in phosphate-rich horizons. Moreover, species from group d5 Navicula viridula (Kützing) Ehrenberg and Gomphonema angustatum (Kützing) Rabenhorst (Figure 5) are reported as being associated with nutrient-rich waters in Antarctica (e.g. Jones, 1996; Jones et al., 1993).
Relationship between pigment composition and core lithology
A PCA separates three groups of pigments and simultaneously brings together algal species with shared pigment composition. Moreover, there are statistically significant correlations between some pigments and lithological changes in the history of sedimentation (Figure 8b).
Sea-lagoon PCA group p1
This group contains pigment proxies (alloxanthin, fucoxanthin, and prasinoxanthin) typically found in cryptophytes, diatoms, or chrysophytes and prasinophytes, respectively (Figure 8b). Such pigments are dominant in sea-lagoon clastic sediments with elevated biogenic silica content (unit A). Cryptophytes and prasinophytes are commonly found in the sea waters of the Antarctic region, and their enhanced biomass in this unit is probably associated with the inflow of marine water into the lagoon basin. Cryptophytes are also reported as an important element of phytoplankton assemblages of oligotrophic and ultra-oligotrophic lakes in continental and maritime Antarctica, which inherited marine water trapped during an uplift, and have a wide spectrum of salinity (from hypersaline to freshwater) (Laybourn-Parry and Wadham, 2014; Laybourn-Parry et al., 2005; Roberts and Laybourn-Parry, 1999). Concentrations of chlorophyll a in unit A are low, but the relatively high concentration of chlorophyll a derivatives suggests an environment favoring decomposition of that pigment (well-illuminated and oxidized conditions). Additionally, the high contribution of pheophorbide among chlorophyll a derivatives might be the effect of intensive grazing pressure (e.g. Camacho, 2006). The pigment pattern of the sea-lagoon sediments (unit A) are masked by other pigments typical of the lake sediments in units C and D. Moreover, a decreasing upward trend in the abundance of pigments of group p1 in the lacustrine core section is probably related to a gradual reduction in the influence of marine water on the increasingly isolated maar affected by progressive isostatic uplift.
Lacustrine eutrophic PCA group p3
It comprises common pigments that are proxies of the total phytoplankton biomass (chlorophyll a and ββ-carotene), pigments specific for green algae (lutein, chlorophyll b, and pheophytin b), cyanobacteria (zeaxanthin and echinenon), purple sulfur bacteria pigments (bacteriochlorophyll a and b), and traces of pyropheophorbide (evidence of grazing pressure). Pigments of this group are important along the entire length of the core except of sea-lagoon sediments (unit A). The highest concentrations occur in sapropel sediments rich in ornithogenic phosphates (units E1, F1, and F2). It is possible that in these units, terrestrial green algae and cyanobacteria were washed from the surface of ornithogenic soil to the lake, so they are allochthonous in origin. This assumption is supported by evidence indicating that chlorophytes and cyanobacteria are the main components of algal communities developing in the soils of penguin rookeries (Garraza et al., 2011; Izaguirre et al., 1993; Mataloni and Tell, 2002). This seems to be also confirmed by the PCA showing the correlation between lutein, chlorophyll b, zeaxanthin, and ornithogenic apatite (Figure 8b). However, there is not convincing relationship between the high content of p3 pigments in the lower part of the lacustrine section (unit C1) and the phosphates level, so the autochthonous production of these algae in a eutrophic environment is also possible.
Nonspecific PCA p2 group
Pigments belonging to group p2 are quantitatively less important. Moreover, they are evenly distributed in the sediment core, irrespective of the lithological composition (e.g. peridinin characteristic of dinoflagellates; Figure 8b).
Pressure of penguin manuring on the lake’s ecosystem
Antarctic lakes that developed along seashores are usually affected by marine animals: seals and birds, forming large rookeries. Nutrients derived from mineralized guano washes down from the breeding area enhance lake production and shape the structure of the lake ecosystem. A suspension in leachates carries some particulate biotic elements, for example, undigested diatoms, algae from food or microbial mats formed in situ on the surface of ornithogenic soil, as well as calcium phosphate that is abundant in guano of birds feeding on krill, crustaceans with chitinous exoskeleton. Guano deposits (Baroni and Orombelli, 1994; Emslie, 2001) or calcium phosphate, released from mineralized chitin of krill (Tatur et al., 1997), is commonly used in paleolimnological and paleopedological studies as a marker of penguin activity and even as an indicator of population size changes over time (Huang et al., 2011, 2016; Liu et al., 2005; Sun et al., 2000; Wang et al., 2007).
Penguins and giant petrels have been present in the vicinity of the maar lake since the very beginning of its formation. This is proven by enhanced phosphorus content in the lowest lacustrine sediment (unit C1). However, a very significant increase in phosphorus concentration was noted only in the LIA (unit E2/F1, about 400–530 years ago). This time span fits to the LIA expansion of glaciers on King George Island, which caused exodus of the penguins from a large rookery at Low Head situated about 10 km away on King George Island. The exodus is documented by an abandoned cover of ornithogenic soils bearing penguin bones that are partially included into the most external band of lateral moraines of LIA and can be seen in an outcrop cut at 4 m a.s.l. by heavy storm waves (Tatur, 1989; Tatur and Myrcha, 1989). The youngest penguin bones from the sediment in a local pool were dated by Tatur et al. (1997) and Emslie (2001) to 700–750 yr BP (end of Medieval Climate Anomaly (MCA)). Thus, the expansion of LIA glaciers demolished the rookery annihilated easy access from the sea to breeding sites and forcing the population of Adélie penguins to look for a new breeding site. The penguins could be attracted by the nearest accessible breeding site on the open-sea coast of Penguin Island since fast ice may have been absent here from early summer.
An increase in the penguin population during the LIA explains the higher concentration of phosphorus and allochthonous biotic elements in the lake sediments (as observed by Hu et al., 2013; Huang et al., 2016); however, it does not explain the lack of erosion at that time, nor the suppression of autochthonous diatom numbers observed in that time.
The maar lake is situated in a deep crater in the shade of Deacon Peak, 200 m high. The lake is relatively deep (16 m), and the winter ice cover on the lake lasts longer here than on any of the other lakes on King George Island. The core was sampled from the ice on the first day of January 2003 (the middle of summer) during warm austral summer, but in colder summers of the LIA, when the Antarctica was generally cooler and stormier (Bertler et al., 2011), permanent ice may have covered this lake. Permanent ice cover on the lake and frozen soils in the watershed would reduce erosion processes, and crucially, the suspension of guano with attached flora would be washed down from the always warmer rookery to the lake through the near shoreline water band around the ice cap covering the lake (formed commonly due to the higher insolation of the land). The ice and snow cover reduced the light penetration and eliminated autochthonous algae from the lake ecosystem, protects the lake water column against mixing, and allows upward expansion of the anoxic monimolimnion that may have occupied a shallower habitat suitable for the development of autochthonous benthic diatom, green algae, and cyanobacteria.
Anoxic environment at bottom sediments of maar
Pyrite framboids are minute raspberry-shaped spherical aggregates, up to 5 µm in diameter, consisted of assemblages of tiny crystallites. They develop from the most labile sources of iron in saline microenvironments experiencing high rates of bacterial sulfate reduction (Canfield et al., 1992; Folk, 2005). In euxinic environments, framboid nucleation and growth occurs within anoxic water columns and/or within anoxic sediment pore waters. Nucleation takes place very early during anoxic diagenesis, generally within the top few centimeters of burial, and once formed, framboids are stable over geological time (Wilkin et al., 1996).
The uniform size of framboids along the sediment core indicates that the sedimentary environment was geochemically relatively stable during the lake’s history (Wilkin et al., 1996). Pyrite framboids are usually formed in water columns on the oxic-anoxic water interface (Muramoto et al., 1991) or are eventually largely completed at or above the sediment–water interface (Lyons and Berner, 1992). In the investigated case, the formation of framboids seems to be limited by availability of reactive iron, as in comparable environments described by Suits and Wilkin (1998). Hydrogen sulfide occurs abundantly in the anoxic monimolimnion of the maar. However, the supply of available iron with oxidized surface water from the watershed seems to be less probable than the release of divalent iron from sediments weathered in anaerobic conditions. Therefore, we suppose that the main site of framboid formation in our case is the sediment–water interface.
Considering the concentrations and atomic ratios of iron and sulfur in microprobe analysis in lighter and darker laminae (Figure 4c and d), we have found that besides clear pyrite framboids, a large part of the iron and sulfur of both laminae might be bound in amorphous iron sulfides hidden in the matrix. Iron sulfides may constitute an important component of the sediment, at least in the sapropel unit (up to 5%).
Origin and evolution of the Petrel Crater maar
The filling of this crater by sediment produced the youngest sediment sequence (PCF-MS) proving the maar evolution from a marine-brackish lagoon to a lacustrine environment (units from A to G in Table 2). The history of the maar is described on the basis of ecological proxies evaluated in discussion: geochemical data, PCA groups of diatoms (d), and pigments (p) presented on the background of global climate changes (Figure 8c).
Initial environment changes due to the glacio-isostatic Holocene regional uplift
The muddy sediment of marine lagoon at the base (unit A) contains benthic littoral diatoms typical of the brackish environment (d1) and comprises an important participation of photosynthetic pigments (p1) that are distinctive for marine water.
Most probably, tectonic–volcanic activity about 1250 years ago documented in silty tephra fallout (unit B) triggered an abrupt glacio-isostatic uplift that separated the lake from the sea. The tephra horizon (probably from the Deception Island volcano) marks a sudden change in the sea lagoon to a lacustrine environment.
The ecological evolution of the maar lake was initially (units C and D1) constrained by an uplift; the maar water level decreased and shore erosion (sand) intensified. The influence of marine water biota vanished with time (d1 and p1), the lake became meromictic, and the freshwater mixolimnion layer expanded (d2, d6, p2, and p3).
Influence of climate on maar paleoenvironment in the last centuries
Regional uplift of the land was no longer important as an environmental driver in the physically stabilized lake environment about 750 years ago (D1/D2 unit), with a constant lake table determined by a threshold about 2 m a.s.l. The lake’s ecosystem evolved onward mainly to the rhythm of climatic changes. Since that time, two completely different ecological patterns of the lake have occurred.
The first pattern (units: D2, E1, G1, and G4) is comparable with the current status of the maar: a lake with muddy sedimentation and the anoxic monimolimnion consisted of marine water, and the distinctly formed freshwater-brackish, well-oxygenated mixolimnion. Phytoplankton consists of diverse algae species documented in pigments and diatoms (relatively abundant p2 and dominant d6), which are common in the lakes of the area and, therefore, identified as autochthonous.
The second pattern (units: E2, F, G2, and G3) is characterized by sapropel sedimentation rich in penguin guano (a very high phosphate CaP content, a low C/N ratio, and highly reductive conditions expressed by abundant pyrite framboids and a low Mn/Fe ratio). Formerly important phytoplankton assemblages were highly reduced and green algae plus cyanobacteria (p3), and a few diatoms (forming d3 PCA group: Psammothidium metakryophilum, Pinnularia microstauron, and L. muticopsis) became absolute dominants, probably mostly of allochthonous origin supplied to the lake with guano washed from the penguin rookery.
Last Millennium climate scenario from sediment of Petrel Crater lake
The sequence of maar paleoenvironmental patterns (modes) in the last millennium is convergent with rhythm of climate changes in global (Figure 8c) and regional (synthesized in introduction) ranges. The second mode, related to intense pressure of penguin manuring (units E2, F1, and F2), fits to the climate cooling during the LIA (AD 1250–1650, according to IPPC 2013) and units G2 and G3 might tentatively be correlated with periods of colder Pacific Decadal Oscillation (PDO) over the last century – 1890–1915 and 1945–1977 (Easterbrook, 2011, and references therein) – marked on the general warming trend (Thomas et al., 2009).
Conclusion
Regional tectonic–volcanic activity about 1250 years ago triggered glacio-isostatic lifting up of submerged phreatomagmatic crater that changed into meromictic lake. Ecological evolution of this lake was initially controlled by vertical movement of land; however, impact of climate changes becomes important in the last centuries.
During the LIA, the maar might have been covered by permanent ice. The intensity of erosion processes in the frozen catchment area ceased. A supply of mineralized protein-rich guano from the possibly increased penguin rookery on the lake shore caused lack of oxygen due to intense OM mineralization and an expansion of the anoxic monimolimnion to the ice surface and contributed to a significant reduction of autochthonous lacustrine biota of the maar. While substantially increased discharge of allochthonous biota delivered passively with guano. The compiled results imply for the first time the record of LIA in lake sediments of the maritime Antarctic.
Footnotes
Acknowledgements
We thank our friends who helped us with the fieldwork: Andrzej Przybycin, Anna Dolimat, and Mikołaj Golachowski. We are grateful to Kontekst Transations for English language revision. Finally, we want to sincerely thank anonymous reviewers for their constructive comments.
Funding
This work was carried out as part of the IMCOAST Project (PolarCLIMATE-PP-001) European Partnership in Polar Climate Science and supported by the project UMO – 2012/05/B/ST10/01130 financed by National Science Center, Poland.