
Abstract
Holocene environmental change is poorly characterized throughout much of the High Plains. Playa wetlands are sites with appreciable potential for reconstructing environmental change because of their environmental sensitivity. Radiocarbon, litho- and magneto-stratigraphic, and stable carbon isotope (δ13C) data for four playas on the Kansas High Plains indicate significant change occurred throughout the Holocene, which had profound impacts on playa ecosystem functions (e.g. groundwater recharge, surface water storage, and habitat). Minimum δ13C values and buried soils observed during the Pleistocene–Holocene transition suggest sufficient moisture to support vegetative cover and promote pedogenesis. Low magnetic susceptibility, rapid increase in δ13C, and weak pedogenic alteration in overlying deposits suggest early Holocene aridity with increased landscape instability and sediment deposition. Early to middle Holocene stratigraphy transitioned from loess with little pedogenic modification into overlying buried soils; δ13C values indicate playa vegetation was composed of as much as 90% C4 plants. Maximum magnetic susceptibility and buried soils in late Holocene deposits indicate sufficient moisture to support vegetative cover and promote pedogenesis. The Pleistocene–Holocene transition was a cool, moist period in which sediment accumulation rates in playas were low and ecosystem functions were optimal. Throughout much of the early to early–middle Holocene, playas were subaerially exposed, sediment accumulation outpaced soil formation, and ecosystem functions were inhibited by lack of moisture. From the middle to late Holocene, the climate returned to more mesic conditions, sediment accumulation and soil formation were in near equilibrium, and the ability of playas to perform ecosystem functions increased.
Keywords
Introduction
Holocene environmental change has been poorly characterized throughout much of the High Plains because of a paucity of sites with dated high-resolution archives of environmental proxies. Playas, relatively small, shallow, depressional, ephemeral wetlands, are sites that are especially useful for reconstructing environmental change on the High Plains for several reasons. Because of their small size and ephemeral nature, playas are sensitive to environmental change and can preserve evidence of small-scale and large-scale environmental change. Playas are unique in that they are influenced by fluvial, lacustrine, and eolian processes, with the dominant geomorphic process at a given time being dependent upon prevailing environmental conditions (Bowen and Johnson, 2012; Holliday et al., 2008). Playas are also the lowest points within internally drained and geographically and hydrologically isolated watersheds (Haukos and Smith, 1994; Tiner, 2003), such that playas are the primary sink for sediment within these watersheds. As a result, playas contain some of the most comprehensive sedimentary records for the central and southern parts of the High Plains, including buried soils, loess, eolian sand, lacustrine muds, and alluvium accumulated under different climates and by distinct geomorphic processes. Additionally, playas are ubiquitous, with more than 80,000 playas distributed throughout the High Plains region of Texas, New Mexico, Oklahoma, Kansas (Figure 1), Colorado, and Nebraska (Burris and Skagen, 2013), providing records of environmental change for a nearly 500,000 km2 region.

Playa distribution on the High Plains of western Kansas with research playa locations.
Because of poor preservation of fossil pollen, stable carbon isotopes (δ13C) and environmental magnetics have been used to provide proxy records of environmental change on the High Plains. Stable carbon isotopes have been widely utilized as a paleoenvironmental proxy in the central Great Plains (Feggestad et al., 2004; Johnson and Willey, 2000; Johnson et al., 2007; Leavitt et al., 2007; Mason et al., 2008; Nordt et al., 2007, 2008) and to a limited extent within playas and associated lunette dunes on the High Plains (Arbogast, 1996a; Bowen and Johnson, 2012; Holliday, 1997; Holliday et al., 1996, 2008). Several studies within the central Great Plains have utilized the magnetic record preserved within loess units to aid in identification of buried soils and sedimentary units and to reconstruct paleoclimate (Geiss and Zanner, 2007; Geiss et al., 2008; Johnson and Willey, 2000; Johnson et al., 2007; Miao et al., 2007b; Rousseau and Kukla, 1994).
Although environmental change had dramatic impacts on the High Plains landscape throughout the Holocene, few paleoenvironmental studies have focused on playas. A better understanding of the timing and magnitude of environmental changes and their impacts on the geomorphic processes and ecosystem functions of playas is essential, given the importance and ubiquity of playas on the High Plains. Playas provide several critical ecosystem functions including groundwater recharge, surface water storage, and wetland habitat (Gurdak and Roe, 2009; Haukos and Smith, 1994; Smith, 2003). Determining how playas have responded to past environmental changes is crucial to understand how playas will respond to future environmental changes. We present new radiocarbon, litho- and magneto-stratigraphic, and stable carbon isotope data for four playa wetlands located on the central High Plains of Kansas. These data provide detailed records of environmental change throughout the Holocene and offer new insights into the role that environmental change has on the geomorphic processes occurring within and surrounding playas and playa ecosystem functions.
Regional surficial geology and stratigraphy
Playa research sites are located in Cheyenne, Finney, Scott, and Thomas counties, all on the High Plains region of western Kansas (Figures 1 and 2). Loess is the dominant surface deposit in these counties (Kansas Geological Survey, 2008), and bedrock outcrops include Niobrara chalk, Pierre shale, and the Ogallala Formation (Frye, 1945; Latta, 1944; Prescott, 1952; Waite, 1947). Bedrock units are only exposed along stream valleys; on the uplands, including within and surrounding playa research sites, bedrock is buried by several meters of loess deposits. Deep coring of two Kansas playas detected the Ogallala Formation more than 6 m below the playa surface (Bowen and Johnson, 2012). A backhoe trench excavated 3.6 m deep in a Kansas playa did not encounter bedrock (Arbogast, 1996a). Bedrock units were not encountered during coring of playas for this project, with cores extending to 6 m in two of the playas.
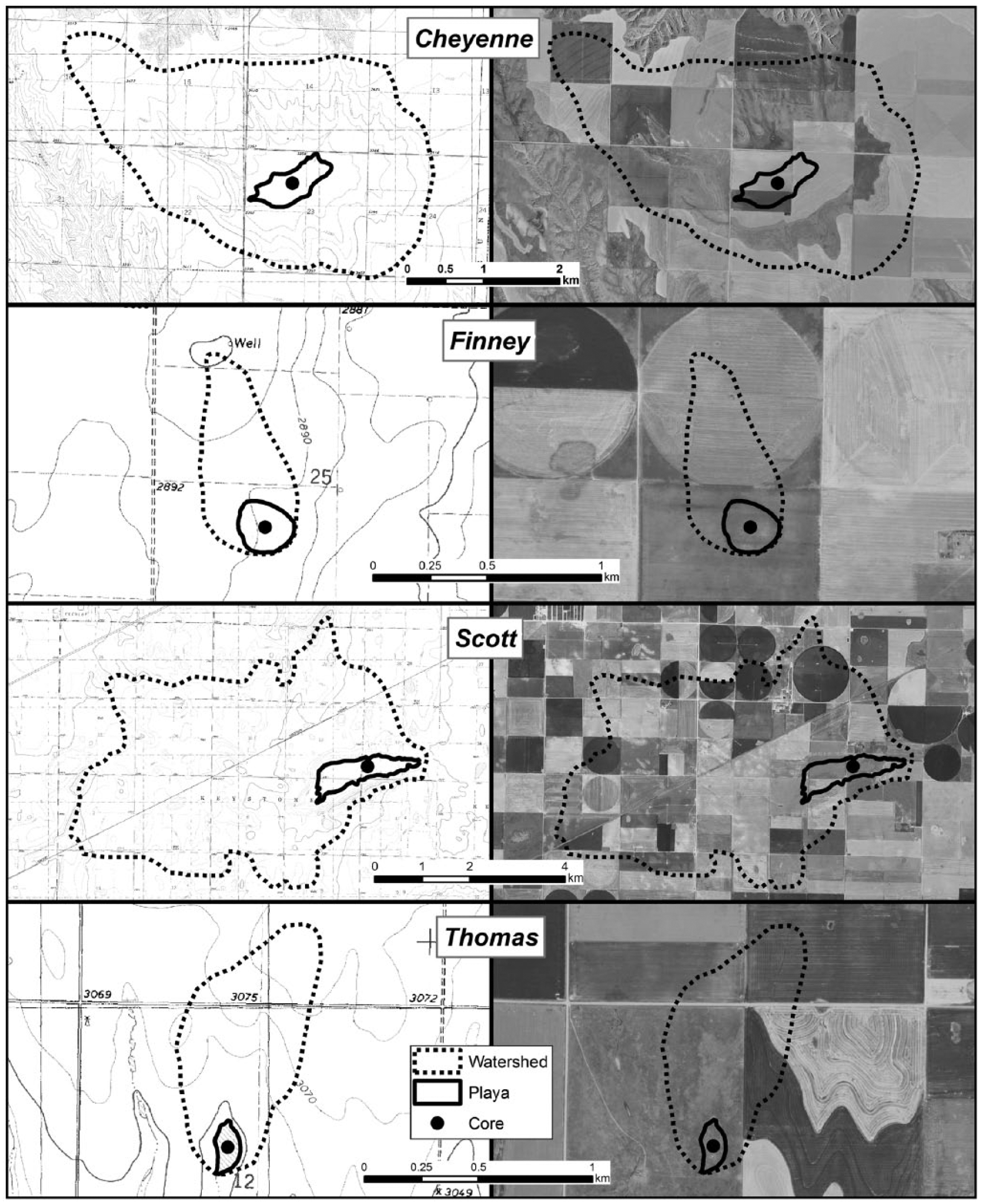
Digital raster graphics (DRGs) and 2 m aerial images of research playas. DRGs have a 1.5-m contour interval for Finney and Scott and 3.0 m for Cheyenne and Thomas.
Five loess units are commonly found on the central Great Plains and include Pre-Illinoisan loess, Loveland Loess (Illinoisan), Gilman Canyon Formation Loess (middle Wisconsin), Peoria Loess (late Wisconsin), and Bignell Loess (Holocene) (Frye and Leonard, 1951; Johnson et al., 2007). Buried soils are common throughout the loess sequence and often separate individual loess units. Loveland Loess accumulated during Marine Isotope Stage (MIS) 6, approximately 165–125 ka, and is capped by the Sangamon Soil which formed throughout MIS 5 and 4, approximately 125–60 ka (Forman and Pierson, 2002; Forman et al., 1992; Maat and Johnson, 1996). Gilman Canyon Formation Loess accumulated slowly throughout MIS 3, approximately 55–22 ka, and the unit is characterized by pedogenic alteration throughout (Johnson et al., 2007; Karlstrom et al., 2008; Maat and Johnson, 1996; Mason et al., 2007; Muhs et al., 1999b). Peoria Loess, deposited rapidly during MIS 2 from 25–12 ka, is the thickest and most extensive of the loess units (Mason et al., 2007; Roberts et al., 2003). The Brady Soil developed in the upper Peoria Loess beginning as early as 15 ka and pedogenesis continued until burial by Bignell Loess at the onset of the Holocene beginning 11–9 ka (Johnson and Willey, 2000; Mason et al., 2003, 2008; Muhs et al., 1999a). Bignell Loess deposition was highly variable spatially on the central Great Plains and temporally throughout the Holocene, with thickness ranging from 0 to 6+ m and multiple buried soils common throughout the unit (Mason and Kuzila, 2000; Mason et al., 2003; Miao et al., 2007a, 2007b).
Bignell and Peoria Loess and Holocene-aged and Brady-age-equivalent soils were the only units encountered during coring of playas (Table 1). Playa basins are inset into loess deposits, and their sedimentary fills consist predominantly of loess with a mixture of lacustrine mud, reworked fluvial deposits eroded from the surrounding uplands, and soil and sediment weathered in situ; uplands are dominated by regional loess deposits. US Department of Agriculture (USDA) Natural Resources Conservation Service SSURGO 2.2 Soil Survey and Spatial and Tabular Data (2009) mapped clay and silty clay loam soils within playas and silt loam soils on the uplands (Table 2).
Radiocarbon chronology for buried soils preserved within playa cores.
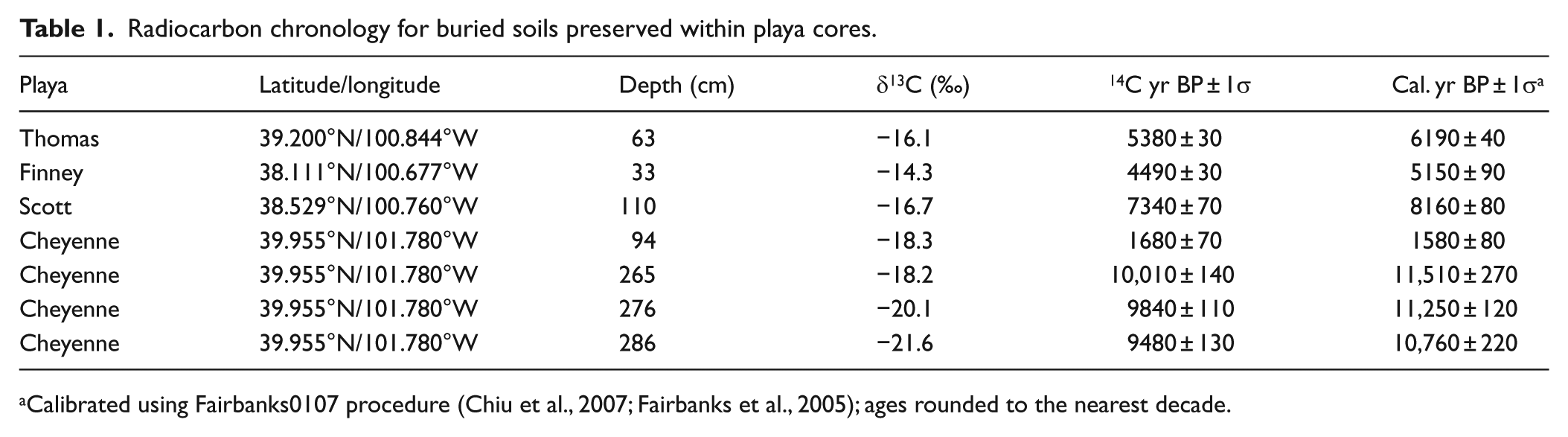
Calibrated using Fairbanks0107 procedure (Chiu et al., 2007; Fairbanks et al., 2005); ages rounded to the nearest decade.
Mapped soils in research playas and on surrounding uplands (USDA Natural Resources Conservation Service, 2009).

Regional climate and vegetation
The High Plains region of Kansas currently experiences a semi-arid climate (Peel et al., 2007). Based on weather station records proximal to each playa, average annual temperature and precipitation were approximately 11°C and 50 cm, respectively, with lowest temperatures of ~−2°C in January, highest temperatures of ~25°C in July, and the bulk of precipitation falling from May through August (High Plains Regional Climate Center, 2015). Native vegetation of the region consisted of short grass prairie dominated by Northern grama-buffalograss (Bouteloua–Buchloe) prairie (Kuchler, 1974). Today, little native vegetation remains because most of the prairie has been converted to agriculture.
Based on the 2006 USGS National Land Cover Dataset, Thomas playa was mapped as grassland within and surrounding the playa and cultivated cropland for the remainder of the watershed. The entire Finney playa watershed was mapped as cultivated cropland in the dataset. In 2008, Finney playa and the surrounding area was enrolled in the USDA Conservation Reserve Program (CRP) and planted to a mix of non-native grasses. The majority of Scott playa watershed was mapped as cultivated cropland in the dataset, with only a small portion of grassland remaining. Approximately the western half of Scott playa has been converted to wheat production, while the remaining half is grassland. Cheyenne playa and watershed have also been cultivated except for a small portion of grassland adjacent to the western and southern edges of the playa.
Playa and watershed size
Thomas playa is relatively small with a surface area of 1.8 ha and watershed area of 44.5 ha (Figure 2). Based on USGS 1:24,000 topographic maps, playa depth is between 1.5 and 3.0 m. Finney playa is relatively small, with a surface area of 4.3 ha and a watershed of 26.1 ha. Given that the playa is not detectable on the 1:24,000 topographic map, depth is less than 1.5 m. Scott playa is relatively large with a surface area of 89 ha and watershed area of 2480 ha and, based on USGS 1:24,000 topographic maps, a depth between 1.5 and 3.0 m. Cheyenne playa is relatively large with a surface area of 34.4 ha and is ~6 m deep, while the watershed encompasses 1008 ha.
Methods
A single soil core was collected near the center of playas to a depth of 119 cm at Thomas playa, 121 cm at Finney playa, and 600 cm at Cheyenne playa (only the upper 365 cm was analyzed). Two cores were collected from eastern grassed portion of Scott playa to depths of 597 cm (core S1) and 385 cm (core S2). All cores were collected in specially designed plastic liners using a heavy-duty Giddings® hydraulic coring machine. Thomas and Finney playa cores were selected for in-depth analysis, while Scott and Cheyenne playa cores were used to provide corroborating data.
To that end, Thomas and Finney playa cores were described in detail using standard methods of the USDA National Soil Survey Center (Schoeneberger et al., 2012), that is, soil horizonation, Munsell color, soil structure, redoximorphic features, carbonate occurrence, root and root-trace size and density, and pore size and density (Tables 3 and 4). These two cores were analyzed at the National Lacustrine Core Facility (LacCore) at the University of Minnesota (Minneapolis/St Paul) using a variety of Geotek multi-sensor core logging systems that enabled a suite of measurements to be obtained at 0.5 cm intervals without damaging or disturbing core stratigraphy. Bulk density was measured for whole cores using a Geotek gamma density sensor utilizing the gamma ray attenuation and porosity evaluator (GRAPE) method (Anderson et al., 1990). Bulk density was used to aid in identification of distinct soil and sediment units. Magnetic susceptibility of Thomas and Finney playa cores was measured using a Geotek-mounted Bartington MS2C core sensor. Cores were then split in half length-wise using a band saw, and core faces were cleaned by hand. Magnetic susceptibility was again measured using a Geotek-mounted Bartington MS2E point sensor. Geotek GeoScan-III and DMT CoreScan systems were used to collect continuous digital images (0.1 mm intervals) of cores (Figures 3 and 4). Mass-dependent magnetic susceptibility was measured in 5 cm intervals in the Scott playa cores and 2.5 cm intervals in the Cheyenne playa core using a Bartington MS2B sensor at the University of Kansas Soil Geomorphology Research Laboratory.
Description of soil stratigraphy at Thomas playa.

Description of soil stratigraphy at Finney playa.

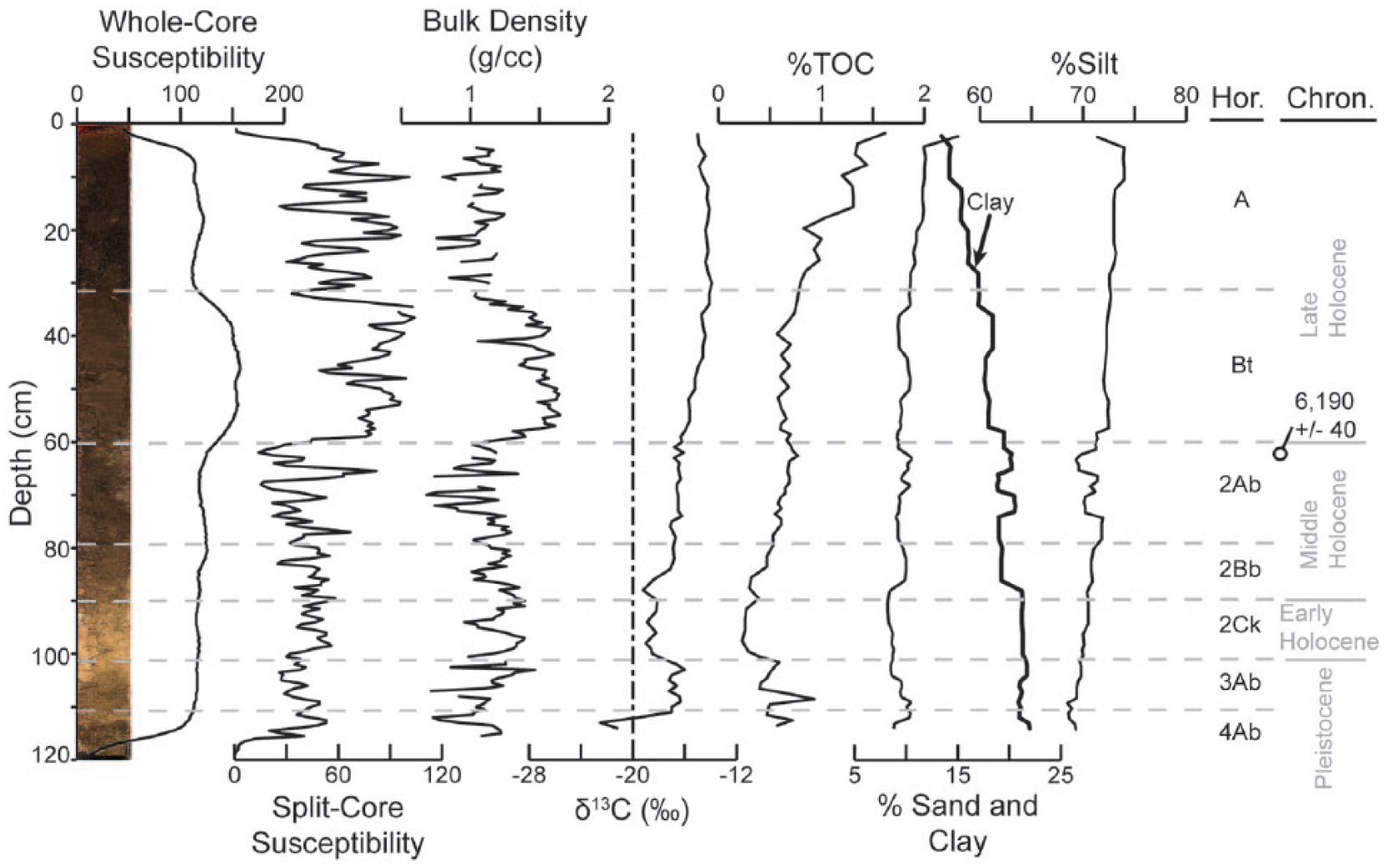
Whole-core and split-core magnetic susceptibility, bulk density, stable carbon isotope, total organic carbon, and particle size distribution data for Thomas playa core.

Whole-core and split-core magnetic susceptibility, bulk density, stable carbon isotope, and total organic carbon data for Finney playa core.
Magnetic susceptibility is typically related to degree of pedogenesis; however, local factors such as hydrology and variations in sediment source also influence magnetic enhancement (Evans and Heller, 2003). For example, cycling between saturated, anaerobic conditions and aerated, aerobic conditions eliminates much of the magnetic signal. Additionally, when sediment accumulation outpaces soil formation, the magnetic signal is depleted. As such, magnetic susceptibility can be used to infer changes in flood frequency and duration and sediment accumulation rates related to environmental changes. Magnetic susceptibility should be greatest during mesic periods in which there is sufficient moisture to support dense plant growth, stabilize the landscape, reduce sediment accumulation rates, and promote pedogenesis. During more arid conditions, plant density decreases, pedogenesis is inhibited, eolian processes dominate, and sediment accumulation rates increase, reducing the magnetic signal. During more aquic conditions, greater runoff delivers more water and sediment to the playa, and fluvial and lacustrine processes dominate. Playa soils remain saturated for prolonged periods, which inhibits pedogenesis, and anoxic-reducing conditions deplete the magnetic signal. Arid and aquic conditions were differentiated based on the absence/presence of redoximorphic features and stable carbon isotope data.
Samples were collected in 1 cm intervals from Thomas playa soil core to determine particle size distribution using a Malvern Mastersizer 2000 laser diffraction particle size analyzer. The Malvern Mastersizer consistently underestimated the clay-size fraction within playa samples (Bowen, unpublished data), which is common using laser diffractometer techniques to measure particle size (Beuselinck et al., 1998; Buurman et al., 2001; Eshel et al., 2004), so the clay fraction was adjusted to include all particles smaller than 8 µm (Bowen and Johnson, 2012).
Cores from all four playas were sampled for stable carbon isotope analysis. Samples were collected from Thomas and Finney playa cores in 1 cm intervals within buried soils and 2 cm intervals from the remainder of the core. Scott playa core S1 was sampled in 10 cm intervals within the upper 130 cm of the soil core and 10–20 cm intervals within the lower 110 cm; only six unequally distributed samples were collected from the upper 2 m of core S2. Cheyenne playa was sampled every 2–9 cm within the upper 90 cm of the soil core and every 5–21 cm in the lower 270 cm.
Stable carbon isotope data were used to reconstruct past vegetation and, by inference, associated climatic conditions. The carbon isotopic ratio (δ13C) of plant material can be used to distinguish between C3 and C4 photosynthetic pathways: C3, cool, moist-season adapted vegetation has an average δ13C value of −27‰, while warm, dry-season adapted C4 vegetation has an average δ13C value of −13‰ (Tieszen and Boutton, 1989). Samples collected from Thomas, Finney, and Scott playa cores were pretreated with 1 M HCl and analyzed with a Thermo Finnigan MAT 253 mass spectrometer at the Keck Paleoenvironmental and Environmental Stable Isotope Laboratory (KPESIL) at the University of Kansas. Similar procedures were conducted to analyze samples collected from Cheyenne playa core at the Stable Isotope Mass Spectrometry Laboratory (SIMSL) at Kansas State University. In addition to δ13C, resulting laboratory data included percent total soil organic carbon (SOC). To estimate the proportion of total SOC contributed by C3 and C4 plants, the following formula was applied (Ludlow et al., 1976; Nordt et al., 1994):

where δ13CSOC is the δ13C value of SOC relative to the Pee Dee belemnite (PDB) standard, δ13CC3 is the mean value for C3 plants (−27‰), and δ13CC4 is the mean value for C4 plants (−13‰).
Sediment samples were collected from buried soils preserved within soil cores for radiocarbon analysis (Table 1). Samples collected from Thomas, Finney, and Cheyenne playas were submitted to the National Ocean Sciences Accelerator Mass Spectrometry Facility (NOSAMS); a sample collected from Scott playa core S2 was submitted to Geochron Laboratories. Samples were first dried, pulverized, and rootlets and other materials were removed under a microscope, and then samples were acid treated to remove inorganic carbon. Radiocarbon ages were determined on bulk, de-calcified samples and converted to calendar ages using the Fairbanks radiocarbon calibration program (Chiu et al., 2007; Fairbanks et al., 2005). Ages were classified as Pleistocene (>~11,700 cal. yr BP), early Holocene (11,700–8200 cal. yr BP), middle Holocene (8200–4200 cal. yr BP), or late Holocene (<4200 cal. yr BP) (Walker et al., 2012).
Results
Playa stratigraphy
Thomas playa contained ~6 cm of historical sediment overlying the modern surface soil (Table 3 and Figure 3). Sediment was differentiated from the underlying soil by lack of structure and color differences. The entire core had been modified by pedogenesis, with thin slightly weathered loess units near the base of the core. The modern surface soil exhibited A-B horizonation and was welded to a buried soil with Ab-Bb-Ck horizonation. A radiocarbon age, determined on a sample collected near the top of the buried soil, returned a middle Holocene age of 6190 ± 40 cal. yr BP (Table 1). Underlying this buried soil, a second weakly developed buried soil was identified, which consisted of an Ab or AC horizon. This weakly developed buried soil was separated from an underlying well-developed buried soil by a thin loess unit; only the upper 6 cm of the Ab horizon of this buried soil was captured during soil coring. It is believed that this lowermost buried soil is of equivalent age as the Brady Soil found in upland loess deposits (hereafter referred to as Brady-equivalent soil).
Finney playa contained ~4 cm of historical sediment overlying a modern surface soil consisting of A-Bt horizonation (Table 4 and Figure 4). The surface soil was welded to a buried soil with Ab-Btb-BCkb-Ck horizonation. A radiocarbon age collected near the top of the buried soil returned a middle Holocene age of 5150 ± 90 cal. yr BP (Table 1). The lower 50 cm of the core consisted of slightly weathered loess with carbonate nodules and diffuse carbonate throughout.
Detailed stratigraphic descriptions were not conducted for Scott and Cheyenne playas; however, horizons were identified, boundary depths were recorded, and hand-texture was determined in Cheyenne playa (Table 5 and Figures 5–7). Scott playa core S1 had 12 cm of historical sediment overlying a well-developed modern surface soil with A-AB-B horizonation welded to a buried soil with Ab-Bb-C horizonation; the upper ~2 m of the core had been heavily modified by pedogenesis (Table 5 and Figure 5). The lower 4 m of core consisted of weakly modified loess. Scott playa core S2 had similar stratigraphy as core S1 (Table 5 and Figure 6). The modern surface soil consisted of an Ap-A-AB-B profile and was welded to a buried soil with an Ab-AC-C profile with weakly modified loess from the base of the Ab horizon to the base of the core. A radiocarbon age collected near the top of the Ab horizon in core S2 returned an age of 8160 ± 80 cal. yr BP (Table 1). Pedogenesis in Scott playa likely began during the later portion of the early Holocene and continued into the middle Holocene.
Soil stratigraphy of Scott playa cores S1 and S2 and Cheyenne playa.


Mass-dependent magnetic susceptibility, stable carbon isotope, and total organic carbon data for Scott playa core S1.

Mass-dependent magnetic susceptibility and stable carbon isotope data for Scott playa core S2.

Whole-core and mass-dependent magnetic susceptibility, stable carbon isotope, and total organic carbon data for Cheyenne playa core.
At Cheyenne playa, the modern surface soil had been degraded because of prolonged cultivation, with an Ap surface horizon or Ap-Bw horizons overlying weakly modified C horizon loess (Table 5 and Figure 7). This upper portion of the profile consisted of 6–20 cm thick silt-rich units separated by 4–8 cm thick sand units; below the surface C horizon, the core was primarily composed of silt-rich deposits with little variation in soil texture. A buried soil underlying the surface soil C horizon consisted of Ab-ABb-Btb-C horizonation. A radiocarbon sample collected near the top of the Ab horizon returned a late Holocene age of 1580 ± 80 cal. yr BP (Table 1). A second buried soil, separated from the base of the upper buried soil by more than 1 m of weakly modified, loess-rich deposits, had an Ab-Bb-C profile. Three radiocarbon samples collected from the Ab horizon of this buried soil produced Pleistocene–Holocene (P-H) transition to early Holocene ages of 11,510 ± 270 cal. yr BP at 265 cm, 11,250 ± 120 cal. yr BP at 276 cm, and 10,760 ± 220 at 276 cm (Table 1). These ages are inverted but within approximately 1 standard deviation of each other.
Bulk density
Bulk density data were generated from Thomas and Finney playa cores. At Thomas playa, bulk density ranged from 0.68 to 1.65 g/cm3 (Figure 3) and averaged 1.2 g/cm3 for the entire core. Bulk density was greatest within the surface soil B horizon, and above and below the B horizon, bulk density was lower but oscillatory. Bulk density was also greater in the 2Bb horizon compared with the 2Ab horizon. At Finney playa, bulk density ranged from 0.76 to 1.79 g/cm3 (Figure 4) and averaged 1.45 g/cm3. Bulk density was greatest near the surface and exhibited a generally declining trend to the base of the core, with a slight increase in the 2Btb horizon.
Particle size distribution
Particle size distribution data were derived for only Thomas playa (Figure 3). The playa was dominated by silt-sized particles, with silt content ranging from 68.5% to 74%, clay content ranging from 13.4% to 22%, and sand content ranging from 8.2% to 15.1%. Silt and sand contents were greatest near the surface and exhibited declining trends with depth. Clay exhibited the opposite trend with minimum values near the surface, and clay content generally increased with depth. Based on the Shepard sediment classification system, the soil core consisted entirely of clayey silt. Given the small size and low relief of the watershed restricting the amount of sediment delivered via fluvial processes, lack of distinct trends in clay associated with most soil horizons, and generally increasing silt content from the base to the top of the core, influx of loess via eolian deposition must have remained near equilibrium with soil formation throughout the Holocene since the entire profile has been pedogenically modified.
Stable carbon isotopes
Stable carbon isotope values and trends were generally similar at all playas (Figures 3–7). Values of δ13C obtained from Thomas playa core ranged from −22.5‰ to −13.9‰ (Figure 3): minimum δ13C values were recorded near the base of the core with an overall minimum value of −22.5‰ (117 cm), increased to −16.3‰ at 113 cm, fluctuated between −16.0‰ and −17.2‰ between depths of 113–107 cm, and progressively decreased to −19.0‰ at 102 cm. Isotope values oscillated between −19.2‰ and −18.1‰ from depths of 102–92 cm, and above a depth of 92 cm, δ13C values exhibited a generally increasing trend to a maximum value of −13.9‰ at 34 cm. Isotope values then exhibited a decreasing trend to −15‰ in the uppermost sample collected at a depth of 6 cm.
At Finney playa, δ13C ranged from a minimum value of −21.0‰ at a depth of 120 cm, at the base of the core, to −13.6‰ at a depth of 24 cm (Figure 4). Values exhibited a generally increasing trend between these depths, although δ13C oscillated between −17.1‰ and −14.6‰ from 66 to 43 cm. Two distinct increases occurred from −20.8‰ at 105 cm to −18.2‰ at 100 cm and from −17.1‰ at 68 cm to −14.7‰ at 66 cm, while δ13C values declined sharply from −17.8‰ at 95 cm to −19.2‰ at 90 cm, from −14.7‰ at 60 cm to −16.4‰ at 58 cm, and from −13.6‰ at 24 cm to −15.8‰ at 22 cm. From 24 cm to the surface, δ13C values generally declined to −16.6‰ in the uppermost sample collected at a depth of 6 cm.
In Scott playa core S1, δ13C values ranged from −11.2‰ to −22.5‰ (Figure 5). The maximum δ13C value of −11.2‰ was recorded at 575 cm, 22 cm above the base of the core, and exhibited a declining trend to a minimum value of −22.5‰ at 325 cm. Values of δ13C increased to −18.4‰ at 300 cm, exhibited minor variability, and then increased from −18.8‰ at 260 cm to −16.8‰ at 242 cm before rapidly decreasing to −22.5‰ at 232 cm. Values then rapidly increased to −18.1‰ at a depth of 212 cm. From 212 to 30 cm, δ13C exhibited a generally increasing trend, with a maximum value of −12.8‰ at 30 cm. Isotope values then progressively declined to −18.9‰ at the surface. While only a limited number of samples were analyzed from core S2, δ13C had a generally similar trend as the upper 1.9 m of core S1, although values were slightly more negative (Figure 6).
The greatest variability and range in δ13C were recorded in Cheyenne playa, ranging from −24.6‰ to −13.7‰ (Figure 6). Minimum values were recorded near the base, with values declining from −23.7‰ at 365 cm to −24.6‰ at 318 cm. δ13C values then progressively increased to −17.9‰ at 266 cm. Values of δ13C decreased to −18.4‰ at 248 cm and then increased to −17.4‰ at 229 cm before progressively declining to −20.0‰ at 185 cm. From 185 to 143 cm, δ13C oscillated by less than 1‰ and then progressively increased to −18.0‰ at 91 cm. Values were constant until sharply increasing from −18.0‰ at 82 cm to −14.0‰ at 66 cm. From 66 cm to the surface, δ13C oscillated between −15.8‰ and −13.7‰.
Magnetic susceptibility
Whole-core magnetic susceptibility at Thomas playa was relatively high and uniform, with low values obtained near the upper and lower portions of the soil core and increased magnetic susceptibility within the surface soil Bt horizon (Figure 3). Whole-core magnetic susceptibility values of underlying buried soil Ab, Bb, and Ck horizons were similar to values of the A horizon of the surface soil. Split-core magnetic susceptibility values from Thomas playa were lower and exhibited greater variability than whole-core magnetic susceptibility but exhibited similar trends (Figure 3). Highest split-core magnetic susceptibility was recorded within the surface soil B horizon, but the surface soil A horizon had greater magnetic susceptibility compared with underlying buried soils. Several individual peaks and troughs were detected in the split-core analysis. The MS2E sensor is highly sensitive to surface characteristics, including undulations of less than 1 mm (Dearing, 1999). Given a lack of distinct changes in stratigraphy and particle size coincident with changes in magnetic susceptibility, individual peaks and troughs are likely because of slight undulations and cracks on the face of the core as a result of splitting and handling the core.
Whole-core magnetic susceptibility at Finney playa exhibited a declining trend from near the surface to the bottom of the core, with slight magnetic enhancement within the Ab and Btb horizons of the buried soil welded to the Bt horizon of the surface soil (Figure 4). Split-core magnetic susceptibility was similar in the upper and lower portions of the playa core but exhibited considerable variability in between (Figure 4). Split-core magnetic susceptibility was enhanced in the lower portion of the A horizon and upper portion of the B horizon for both the surface soil and buried soil and within the transitional BCk horizon of the buried soil. Similar to Thomas playa, individual peaks and troughs in magnetic susceptibility are likely related to variations in the surface of the core during measurement, rather than a result of pedogenic, hydrologic, or depositional processes.
Mass-dependent magnetic susceptibility in Scott cores was nearly identical for cores S1 and S2 (Figures 5 and 6). Magnetic susceptibility was greatest within the surface soils. Susceptibility values declined within the lower portion of surface B horizons and remained relatively low and uniform to the base of the cores. Magnetic susceptibility was slightly enhanced in the upper portion of Ab horizons of the buried soil compared with underlying horizons, but values were much lower compared with surface soils.
The greatest mass-dependent magnetic susceptibility values at Cheyenne playa were recorded within the surface soil A horizon, and values declined through the C horizon of the surface soil and Ab horizon of the underlying buried soil, although values were highly oscillatory (Figure 7). Three distinct peaks in the upper ~75 cm occur within silt units and troughs occur within interlayered sandier units. Magnetic enhancement occurred within the ABb and Btb horizons and the upper portion of the C horizon of the uppermost buried soil. Magnetic susceptibility values were also enhanced within the lowermost buried soil, and values were only slightly lower compared with the surface soil. Two distinct peaks were recorded near the base of the second C horizon and near the top of the 3Ab horizon. The cause of these peaks is unclear, but since they are both the result of single samples and there were no distinct changes in stratigraphy, they are not likely the result of changes in pedogenic, hydrologic, or depositional processes.
Discussion
The Holocene stratigraphic record at all research playas consists of weakly modified Bignell Loess overlain by a single buried soil welded to the modern soil (Figure 8). Soils are silt- and clay-rich with few sand laminations, and there is no evidence to suggest that playas experienced prolonged saturation during any time within the Holocene. Radiocarbon and stratigraphic data confirm that the Cheyenne and Scott playa soil cores preserved the entire Holocene record, although a Brady-equivalent soil was not encountered in Scott playa deposits. At Thomas playa, a radiocarbon age of 6190 ± 40 cal. yr BP at 63 cm and a poorly developed A horizon overlying a well-developed A horizon at the base of the core indicate the entire Holocene record was likely captured because a well-developed A horizon overlain by a poorly developed A or AC horizon is common for the Brady Soil throughout the Great Plains (Jacobs and Mason, 2004). Based on a radiocarbon age of 5150 ± 90 cal. yr BP near the top of the Finney playa core, C3-dominated stable carbon isotope values at the bottom of the core, and only slight pedogenic modification of the lower 50 cm of the core, the entire Holocene record was likely acquired although a Brady-equivalent soil was not encountered.

Stratigraphy of playa soil cores with associated radiocarbon ages. To improve readability, only the upper 350 cm is included for Cheyenne and Scott cores because the lower profiles consisted of only unweathered loess. Supporting data for radiocarbon ages are listed in Table 1, and detailed stratigraphic descriptions are included in Tables 3–5.
Stable carbon isotope data from all playas indicate the P-H transition was sufficiently moist to support the expansion of wetland vegetation within playas. However, the Brady-equivalent soil in Thomas and Cheyenne playas and lack of redoximorphic features in Finney and Scott playas indicate they did not store water for prolonged periods during the P-H transition. Eolian and fluvial deposition was relatively slow and lacustrine processes would have been temporary. Playa ecosystem functions were likely at their peak during the P-H transition compared with any other time in the Holocene though, with playas providing at least seasonal groundwater recharge and water sources and wetland habitat for wildlife.
The transition from the Brady-equivalent soil into overlying sediment during the early Holocene within Thomas and Cheyenne playas indicates that sediment deposition rates increased and eventually sedimentation outpaced pedogenesis in the early Holocene. The increase in δ13C values from Pleistocene to early Holocene deposits indicates that C4 plants expanded as the climate became warmer and/or drier during the initial phases of Bignell Loess deposition. However, a 3‰ decline in δ13C in Thomas playa, a 5‰ decline in δ13C in Scott playa, and slight δ13C declines in the other two playas near the P-H boundary indicate that C3 plant abundance increased in the early Holocene. Similar δ13C trends have been observed elsewhere in the central Great Plains in early Holocene Bignell Loess deposits (Feggestad et al., 2004; Johnson and Willey, 2000; Mason et al., 2003; Miao et al., 2007a). This decline in δ13C was likely because of increases in drought-tolerant C3 weeds in response to increased aridity and landscape instability. Similarly, Feggestad et al. (2004) attributed a decline in δ13C in the early Holocene to increases in herbaceous C3 annuals better suited to disturbance and high rates of sediment accumulation. Increases in C3 herbaceous weeds were also coincident with drought conditions in northern Great Plains lake cores (Clark et al., 2002) and throughout the High Plains during the Dust Bowl of the 1930s (Weaver and Albertson, 1943).
Relatively low magnetic susceptibility and weak pedogenic alterations observed at Cheyenne, Finney, and Thomas playas support the interpretation of early Holocene aridity with low plant productivity and high rates of sediment deposition inhibiting pedogenesis. Loess deposits are not laminated and are similar in color and texture to surrounding upland deposits, suggesting playas were predominantly influenced by regional eolian processes with relatively high rates of loess inputs via airfall on a subaerially exposed playa floor. While the early Holocene began moist enough to allow playas to provide ecosystem functions, the ability to do so was inhibited and declined as playas remained dry for prolonged periods.
Alternatively, the decline in δ13C and low magnetic susceptibility could have been the result of a cooler and wetter climate in the early Holocene. If the climate was cooler and wetter, playas would have stored water for longer periods, promoting expansion of C3 wetland vegetation and depleting the magnetic signal. However, playa deposits are similar to regional loess deposits on uplands and do not exhibit gleying or any other evidence of prolonged saturation to support expansion of wetland vegetation and depletion of the magnetic signal by hydrologic processes.
Stratigraphic and stable carbon isotope data collected from a lunette dune associated with a playa on the east-central edge of the Kansas High Plains indicate that the early Holocene in Kansas was characterized by periodic droughts that destabilized the landscape and resulted in high sediment accumulation rates (Arbogast, 1996a). Playa deposits excavated during an archaeological investigation in southwest Kansas consisted of well-developed P-H-transition-aged soils buried by nearly 1 m of early Holocene deposits (Mandel and Hofman, 2003). An analysis of playas and associated lunettes on the Southern High Plains revealed that the P-H transition and early Holocene was a period alternating between prolonged phases of sediment deposition and brief phases of non-deposition and soil formation, with relatively rapid fluctuations between wet and dry conditions within playas (Holliday, 1997).
Scott playa was the only playa with an early Holocene buried soil preserved (Figure 8). Scott playa is a large playa with a much larger watershed compared with the other playas. Because of its large size, the playa would have been less susceptible to hydrologic fluctuations, receiving more runoff to enhance soil moisture and promote pedogenesis for longer periods compared with smaller playas.
From the early to middle Holocene, the stratigraphic record at all playas transitioned from light-colored loess with little pedogenic modification into overlying buried soils (Figure 8). However, δ13C data indicate the proportion of C4 vegetation in playas increased and playas were likely dry for prolonged periods in most years. Early to middle Holocene C4 plant expansion has been documented throughout the Great Plains (Leavitt et al., 2007; Nordt et al., 2007).
Dated middle Holocene–aged buried soils in Thomas and Finney playas and enhanced magnetic susceptibility in buried soils at all playas indicate that despite regional aridity, sediment accumulation within playas remained relatively low. While arid conditions dominated the early Holocene and early to middle Holocene transition, the middle Holocene was a period with sufficient moisture to support pedogenesis within playas. Precipitation must have still been relatively low because playa floors had to be primarily subaerially exposed in most years to allow pedogenesis without developing hydric soil characteristics. Middle Holocene buried soils have been observed at several sites throughout western Kansas and Nebraska (Miao et al., 2007a, 2007b; Olson and Porter, 2002; Olson et al., 1997), indicating a regional increase in landscape stability and pedogenesis despite regional aridity.
Except for enhancement within the buried soils, magnetic susceptibility did not exhibit clear trends from the early to middle Holocene. Miao et al. (2007a) proposed that a lack of any obvious trends in magnetic susceptibility during the middle Holocene was because of variations in sediment source masking climate-controlled pedogenic enhancement. Given that playas are depositional settings, similar processes may be masking the climatic signal at these locations.
Rather than an increase in amount of precipitation, variability decreased or timing may have shifted during the middle Holocene such that more precipitation was delivered during the peak summer growing season for grasses. This would have increased plant productivity that stabilized the landscape, reduced sediment deposition, and allowed C4 grasses to expand relative to C3 vegetation. Skinner et al. (2002) observed that reduced rainfall variability during the summer growing season resulted in greater C4 productivity, while C3 productivity was relatively unaffected. Changes in rainfall variability within grassland ecosystems in Kansas initiated a vegetation response, particularly for some C4 grasses, equivalent to a 30% change in rainfall amount (Fay et al., 2003).
Alternatively, a shift to more humid conditions may have begun during the middle Holocene, in which lacustrine and fluvial processes would have been more important. Based on pollen, plant macrofossils, phytoliths, carbon isotopes, and alluvial stratigraphic records, Baker et al. (2000) concluded that beginning 5800 yr BP, intermittent periods of drought fluctuated with more moist phases sufficient to stabilize the landscape and promote pedogenesis. The age of buried soils preserved within playas suggests this shift to more humid conditions would have had to occur much earlier in the playas. Given the paucity of evidence on the central High Plains supporting a more humid middle Holocene, and lack of additional evidence within the playas to support more humid conditions, enhanced pedogenesis within playas during the middle Holocene was likely because of decreased precipitation variability rather than increased precipitation.
From the early to middle Holocene, eolian processes became less effective as a shift in the timing of precipitation stabilized the landscape. Playas were persistently seasonally inundated, enhancing soil moisture, stabilizing the playa floor, and promoting pedogenesis. Lack of gleying, enhanced magnetic susceptibility in middle Holocene soils, and lack of laminations or sand units suggest that lacustrine and fluvial processes were minimal, and playas were dominated by pedogenic processes with low rates of eolian sediment deposition. The late middle Holocene would have been a time of increased ecosystem functions within playas, at least seasonally.
Although plant communities were predominantly composed of C4 vegetation during the late Holocene (Feggestad et al., 2004; Leavitt et al., 2007; Miao et al., 2007a; Olson and Porter, 2002), evidence throughout the central Great Plains suggests a more humid climate with lower sediment accumulation rates and greater pedogenic imprinting compared with the middle Holocene (Jacobs and Mason, 2004; Mason et al., 2003). As much as 80–95% of the vegetative cover at all four playas consisted of C4 plants during the late Holocene. However, maximum magnetic susceptibility and well-developed soils in playas provide evidence that the late Holocene was dominated by slower sediment deposition and more effective pedogenesis. Late Holocene deposits are thicker in the two larger playas (Scott and Cheyenne), and distinct sand layers were deposited in Cheyenne playa, suggesting playas were influenced by fluvial processes more during the late Holocene while eolian and lacustrine processes were minimal.
Several small-scale fluctuations in climate throughout the late Holocene may have impacted playa ecosystem functions, yet playas likely functioned similar to today. Anecdotal evidence from land owners indicate that currently several centimeters of precipitation within 1–2 days are required for water to accumulate within playas, but playas rarely store water for more than a few days. Today, ecosystem functions of most playas, even those in near-natural condition, are greatly reduced because of recurrent drought conditions and lowering of the water table, although these conditions have been exacerbated by agricultural activities (Johnson et al., 2012; Smith et al., 2011). Several centimeters of recent sediment blanketing the modern soil in playas highlight the role of increased agricultural activities within the watersheds on increased sediment accumulation within playas, a factor that reduces water storage capacity and hydroperiod and inhibits playa ecosystem functions.
Conclusion
Records of Holocene environmental change preserved within High Plains playas are in close agreement with environmental records collected from a variety of other High Plains settings including upland loess deposits (Feggestad et al., 2004; Jacobs and Mason, 2004; Mason et al., 2003; Miao et al., 2007a), alluvial fans and terraces (Faulkner, 2002; Mandel, 2008), other playa-lunette systems (Arbogast, 1996a; Bowen and Johnson, 2012; Holliday, 1997; Holliday et al., 2008; Rich, 2013), and eolian sand dunes (Arbogast, 1996b; Forman et al., 2001, 2008; Halfen and Johnson, 2013; Miao et al., 2007a; Muhs, 1985).
Lack of hydric soil characteristics throughout soil cores suggests that the entire Holocene was dominated by arid to mesic conditions in which the playas were subaerially exposed most of the year in most years. Eolian processes dominated during the more arid early Holocene, while fluvial processes were enhanced during the late Holocene. Pedogenesis within seasonally inundated playas characterized the P-H transition and much of the middle and late Holocene. Playas did not store water for prolonged periods at any point in the Holocene, so lacustrine processes were minimal.
Playas provided the greatest ecosystem functions during the P-H transition and the earliest early Holocene, when moisture availability was greatest. From the early Holocene to early–middle Holocene, playa ecosystem functions were greatly inhibited by prolonged arid conditions that resulted in high sediment accumulation rates on subaerially exposed playa floors. A return to more mesic conditions during the middle Holocene, and continuing into the late Holocene, delivered more water to playas and enhanced playa ecosystem functions, at least seasonally.
Footnotes
Acknowledgements
Special thanks to undergraduate assistants Erin De Lee, Tanner Popp, Scott Klopfenstein, and Chelsy Herring for their help in collecting and analyzing soil cores. The authors would like to thank Vance Holliday and Joe Mason for providing constructive reviews that greatly improved this manuscript. We would also like to express our gratitude to the land owners for allowing access to their playas.
Funding
This work was supported by the University of Wisconsin Oshkosh Faculty Development Fund (grant number FDR716); the University of Kansas General Research Fund, the Playa Lakes Joint Venture (grant number 574); the US Environmental Protection Agency (grant number 98780001); and the US Fish and Wildlife Service (grant number F13AP00338).