
Abstract
The current study provides a fine-grained analysis of evidence for sustained pre-Columbian human occupation and socio-ecological interaction within Everglades National Park. Utilizing archaeological data on dietary and cultural patterns recovered from recent excavations at a prehistoric tree island site, we argue the role of ancient human populations in the formation or augmentation of tree islands should be incorporated into environmental models of the tree island lifecycle. High phosphorus levels in human waste, especially the largely organic waste of prehistoric populations, as well as other anthropogenic factors have not been adequately factored into current environmental models of tree island formation or the ecological evolution of the Everglades. More broadly, while socio-ecological modeling is at the core of current scholarly and restoration paradigms, expanded collaboration between environmental scientists and archaeologists will lead to more accurate identification of anthropogenic environmental impacts over time.
Introduction
As one of the largest and most costly environmental management projects in the history of the United States, the greater Everglades restoration program, which encompasses projects within many federal and state agencies including the Comprehensive Everglades Restoration Plan, has an estimated operating budget of more than US$20 billion dollars. The current restoration paradigm envisions the Everglades as a water management system. Restoration planning has relied upon mathematical models to reconstruct pre-drainage hydrology and interpret past vegetation change (Larsen et al., 2011; Ogden, 2008: 28; Willard and Bernhardt, 2011: 60). Natural and social scientists conduct research largely in isolation from one another on the issue of Everglades landscape change. As a result, evidence for prehistoric anthropogenic alteration of the Everglades environment has been under-utilized by environmental scientists, and some argue the extent of human modification of landscapes by pre-Columbian people has been exaggerated (Bernhardt, 2011; Bush et al., 2007). When anthropogenic factors are acknowledged, it is often in the absence of quantifiable data to demonstrate the nature of such influence (Graf, 2008; Wetzel et al., 2011). The current study provides examples of the kind of evidence that exists for prehistoric anthropogenic modification of tree islands in the Florida Everglades with the aim of fostering greater collaboration between natural and social scientists.
While human impact is at the core of current restoration models – both redressing misguided efforts to drain the wetlands for development as well as managing future human needs especially in terms of water usage and urban encroachment – the majority of scientific literature on Everglades restoration does not include the relatively abundant data on prehistoric modification of this area. Historic populations such as the 19th century Seminole and 20th century gladesmen have been acknowledged as human occupants of an environment most outsiders considered uninhabitable. The extent of their impact on the landscape, however, is poorly understood. The centuries of human occupation in the Everglades prior to the arrival of Europeans in Florida and the impact of these people on such key features of the landscape as tree islands, native fauna, and waterflow remain largely unexplored. The result is an attenuated understanding of the role of human modification of the Everglades that does not fully account for the ecological and heritage issues raised by restoration efforts.
To address these concerns, the current study augments models of past variability within the Florida Everglades by providing a fine-grained analysis of evidence for sustained pre-Columbian human occupation and socio-ecological interaction within the northeastern boundaries of Everglades National Park. Adopting a ‘socio-ecological systems approach’, we utilize empirical data ‘to identify key drivers, processes, and controls that regulate human behavior and interactions with the environment’ (Redman et al., 2004: 161). Humans are an integral part of virtually all ecosystems, and a socio-ecological systems approach holds the promise of increasing our understanding of human and environment interrelationships through the development of reinforcing and complementary models and frameworks. Archaeological excavations at a prehistoric tree island midden site during 2010 and the subsequent analysis of recovered faunal materials and artifacts adds to the growing body of evidence for long-term human occupation and modification of the Everglades landscape prior to the arrival of Europeans in the New World (Carr, 2002; Goggin, 1950; Griffin, 2002; Schwadron, 2006). The intention of the present study is to increase understanding of the reciprocal impacts of human and ecological systems in the Florida Everglades, particularly in relation to the phenomenon of tree islands. A more accurate understanding of the anthropogenic origin of tree islands could impact restoration and management strategies through greater attention to the demographic history of land use within the Everglades and an enhanced appreciation for how human choices affect the biophysical landscape. Future decisions about water level management, tree island conservation, and overall preservation strategies for cultural resources within the Park boundaries are implicated.
Tree Islands
Ecological studies of an important feature of the Everglades landscape, teardrop-shaped elevations of hardwood trees known as hammocks or more commonly in the natural science literature as ‘tree islands’, should incorporate available data on prehistoric human occupation of these features (Figure 1). The Everglades are frequently described as a patterned landscape that, like most large wetland ecosystems, is a mosaic of heterogeneous vegetation and nutrient composition (Larsen et al., 2011; Wang et al., 2011: 133, Willard and Bernhardt, 2011). Tree islands are patches of woody vegetation within a freshwater wetland environment dominated by non-woody species (Wetzel, 2002; Wetzel et al., 2011). Maintenance of the mosaic nature of the Everglades landscape has been identified as a conservation priority by many entities (National Research Council, 2003), and yet the origin and factors that affect the tree island landscape are poorly understood (Espinar et al., 2011: 90). Despite only partial socio-ecological knowledge, the Everglades patterned landscape is often described as ‘self-organizing’ or ‘oligotrophic’, both terms which imply a limited role for human activity in landscape formation (Larsen et al., 2011: 347, Wang et al., 2011: 134). Current conceptual models for tree island formation in the Everglades center on the roles of hydrology, erosion and soil deposition, nutrient availability, and climate variability with little consideration given to anthropogenic factors (Bernhardt, 2011; Espinar et al., 2011; Larsen et al., 2011; Sullivan et al., 2011; Wang et al., 2011). The most persistent theory posits that thin peat deposits collected downstream of high points in the underlying bedrock thus forming a gradual accumulation of sediment and topography (Graf, 2008; Sklar and Van der Valk, 2002).
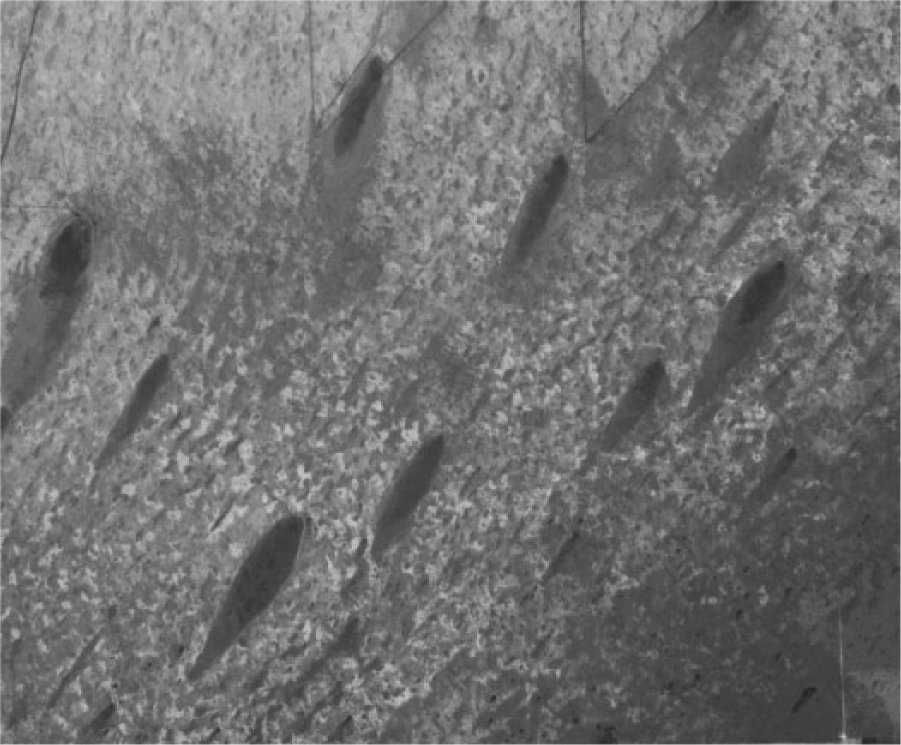
Aerial photo of teardrop-shaped tree islands, Everglades National Park, Florida.
Despite a flurry of research on the issue of tree island formation processes following congressional authorization of the US$7.8 billion Comprehensive Everglades Restoration Plan, there remains a lack of consensus on the importance of each of the factors listed above. The role of human activities in the augmentation of natural factors such as climate variability or nutrient availability has not been systematically examined despite evidence that prehistoric human occupation of tree islands was extensive, affected soil deposition, and altered nutrient availability (especially phosphorus levels which are critical to tree island formation given the overall P scarcity of surrounding marshland). Documentation of prehistoric occupation sites within the Everglades began in the 19th century, but systematic survey and excavation was not conducted until the 1960s. A recent comprehensive summary of tree island archaeology suggests that less than 1/10th of 1% of all tree island archaeological sites have been the subject of archaeological investigation (Carr, 2002: 203). Incorporation of data on the full range of human activities into models of tree island formation and conservation will produce a more accurate framework for ecosystem management.
In 2004, scientists associated with the Long-Term Ecological Research Network of the National Science Foundation issued a call for greater integration of social and environmental data in models of ecological change (Redman et al., 2004). The articulation of the importance of scientific research grounded in a framework that examines a single, complex social-ecological system rather than one that is separated into ‘natural’ and ‘human’ elements has been widely cited and highly influential in ecological research. In particular, recent ecosystem management programs have been designed with recognition of the complex interplay of ‘human and non-human associations’, a phrase employed by anthropologist Laura Ogden (2008) to convey how ecosystems encompass social actors as well as environmental processes. Examples of interactions which epitomize the linkages between human and ecological systems include land-use decisions, resource consumption patterns, and disposal habits (Redman et al., 2004: 164). The socio-ecological model is firmly established today as the best framework for understanding long-term environmental change and especially for the creation of models for future ecosystem management policies.
Methods and materials
In March 2010, the University of Miami in partnership with Everglades National Park conducted fieldwork at the Booth site, a tree island with evidence of prehistoric occupation (Figure 2). Field research was focused on obtaining simple data about the Booth site in order to better manage the conservation of cultural resources preserved at the site and document the potential time depth of human activity at this particular tree island. Basic information about the size, extent, and nature of the site was unknown and research was designed to provide the maximum amount of baseline data with minimal impact on cultural resources. Fieldwork was designed to provide enough information about the site components that guidelines for vegetation and debris removal could be drafted for the National Park staff and archaeological materials from the Booth site could be placed into a broad regional and temporal context.
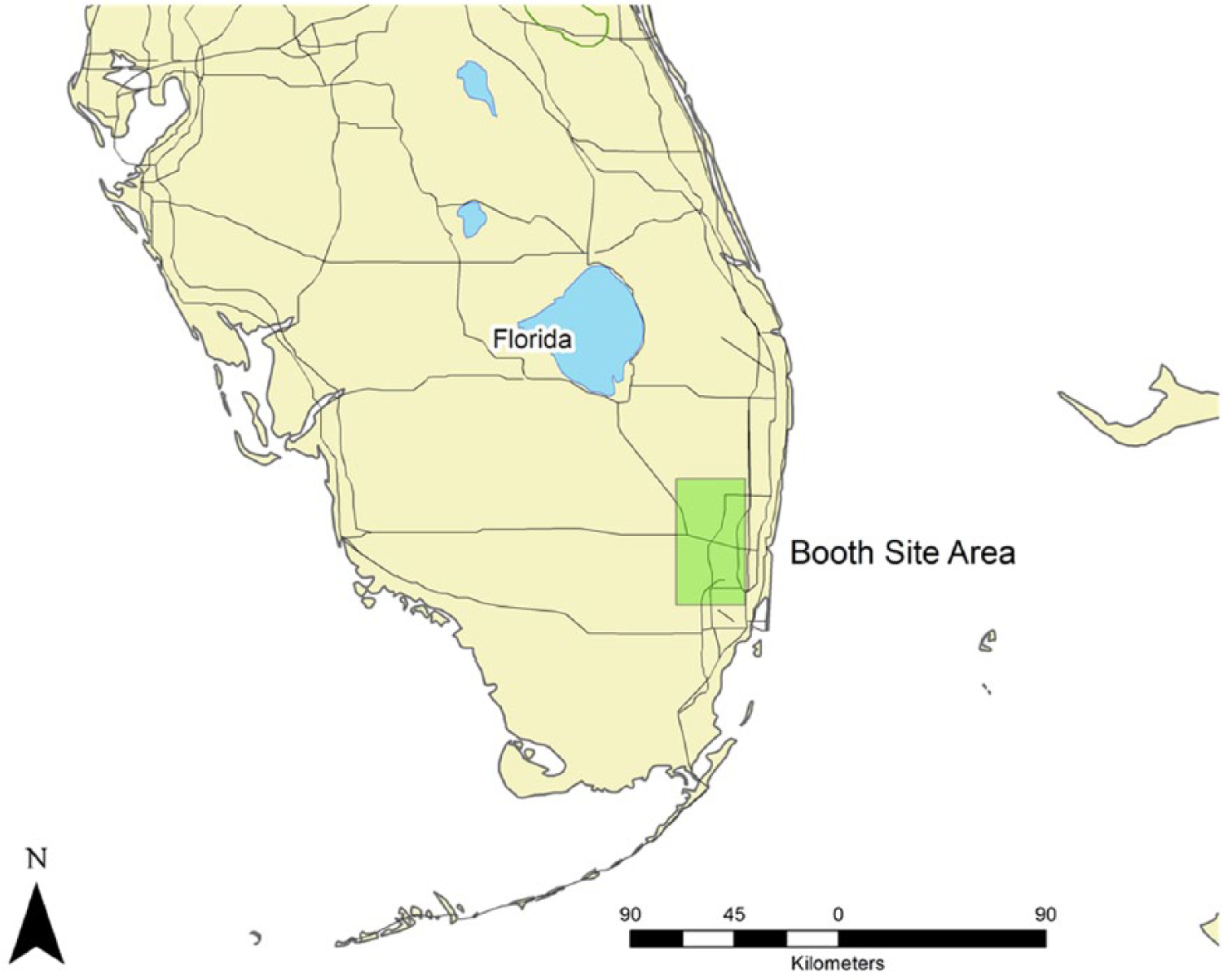
General location of the Booth site, Everglades National Park, Florida.
Site location and history
John Ehrenhard, Robert Carr, and R. C. Taylor (1978) created a predictive model for the location of prehistoric sites within the Everglades using infrared aerial photographs that revealed vegetation types and proximity to water. These models indicated prehistoric sites in the Everglades would most likely be found on the highest ground – hardwood tree islands. High-probability sites are described as prehistoric black earth middens, composed of dark brown to black organically stained soil. The soil contains a dense quantity of vertebrate faunal remains, some marine shell, and ceramics along with other midden debris. Because of the amount and type of debris found at these sites, some models suggest that they were seasonal or ‘special’ use sites (Griffin, 2002; Milanich, 1994: 310; Widmer, 1988). Ground reconnaissance proved the models to be extremely accurate, and 80–90% of Everglades tree islands contain archaeological materials that are the remains of prehistoric occupation (Ehrenhard, 1980: 110; Schwadron, 2009: 91). In an archaeological survey of Everglades National Park, the models proved successful and a total of 191 archaeological sites were identified within the Park boundaries (Schwadron, 2009: 91). A subsequent survey of Big Cypress National Preserve identified 395 additional archaeological sites.
In 2004, the Southeast Archaeological Center (SEAC) of the National Park Service began archaeological investigations of the Eastern Expansion Area (EEEA) of Everglades National Park. The EEEA is located on the eastern edge of the Shark River Slough, a 10-mile wide marshy river in the center of the Everglades. The Booth site, also known as the ‘Howard’ or ‘Coptic Camp’ site, and designated 8DA1085 in the Florida Master Site File, is an archaeological site located on a large hardwood hammock tree island in the extreme eastern edge of the EEEA. The site was first recorded with the state in 1979 by Robert Carr, who described it as a black earth midden located on the northeast end of an elongated tree island. In 1998, the site was revisited and a surface collection of artifacts was undertaken. An archaeological update form was submitted to the Master Site File giving the name of the site as the Coptic Camp site because of the occupation of the tree island as a homestead by members of the Ethiopian Zion Coptic Church during the 1970s–1980s.
Research methods
Investigation was undertaken in March 2010 to assess the aerial and chronological extent of the archaeological site as well as the relative state of preservation. Several structures constructed by the previous owner were slated for removal, and investigation was aimed at determining guidelines for National Park Service staff as they began the process of site maintenance and restoration.
Mapping
A 5000 m × 5000 m arbitrary site grid was established over the 30 m × 20 m area described in 1979 by Robert Carr, within which all topography, natural features, and archaeological features were recorded (Figure 3). The Booth site consists of a 2.5 m rise in elevation above the sawgrass floodplain of the surrounding eastern Everglades. This elevation is similar to other tree islands in the Everglades which do not accumulate significant sediment around the roots of trees and shrubs, even over long periods of time. Although the change in elevation is slight, the terrain varies quite dramatically. The floodplain is very flat, drainage is slow, and the areas off midden remain waterlogged much of the year. The soil at the highest points of the tree island is dark, aerated, filled with artifacts, and rarely inundated. Intermediate elevations are sandy and filled with rock. The topographic map shows the tree island is roughly kidney bean shaped and bisected by a modern gravel road. Two points of higher elevation exist on either end of the mound, which is orientated southwest to northeast, and an artificially enhanced pond exists on the northwestern side.
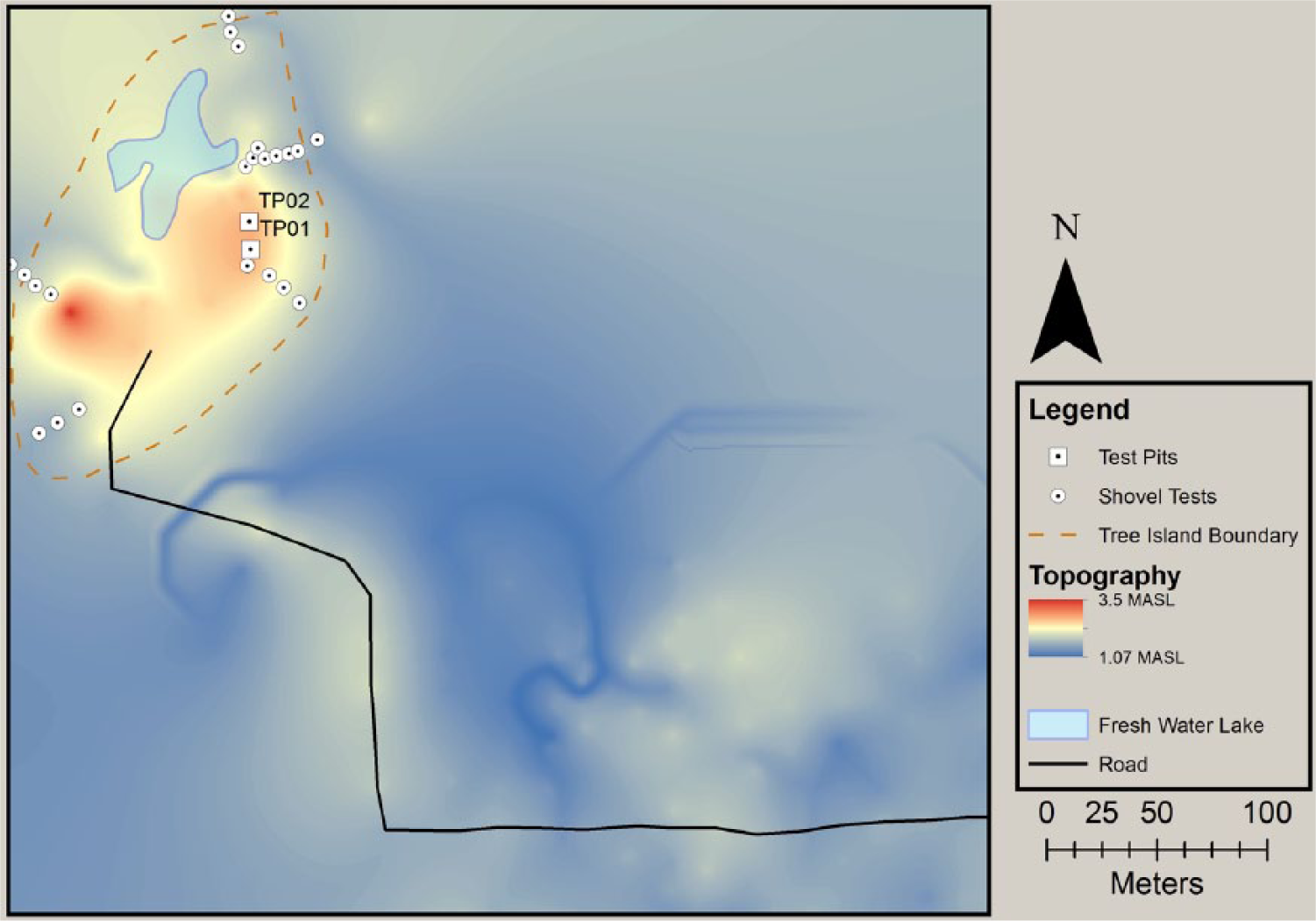
Topographic map of the Booth site indicating locations of shovel tests and test pit excavations.
Excavations
Shovel testing was utilized to determine the extent of the midden. Cultural material from the recovered matrix diminished quickly along transects, as testing proceeded away from visible midden toward the low-lying areas where only culturally sterile waterlogged humic debris was recovered. The small amounts of ceramic and bone material recovered from these tests were fully analyzed in the field and reburied in situ prior to the end of fieldwork at the request of the Miccosukee Tribe of Indians of Florida.
In order to obtain a more representative sample of materials from the midden itself, two 1 m × 1 m test units were placed on relatively undisturbed areas of the midden. Test Pit 1 reached a depth of 40 cm, and Test Pit 2 reached a depth of 47 cm before encountering the calcretion layer, a distinctive feature of tree island archaeological sites that often impedes further excavation (Schwadron, 2006). Both units produced a great quantity of cultural material, especially vertebrate remains and ceramic sherds well known from other black earth middens in South Florida (Carr, 2002, 2012; Goggin, 1950; Griffin, 2002; Schwadron, 2009). After analysis, these artifacts were also reburied on site at the request of the Miccosukee Tribe of Indians of Florida.
Results
Artifact analyses
Ceramics
The Booth Site contained a wide array of ceramic materials that could be firmly dated to individual periods within the Glades regional chronology. The Glades Region Ceramic Chronology was initially developed by John Goggin (n.d.) and has been refined through further excavation and chronometric dating by Griffin (2002) and Russo et al. (1991). Plain ceramic wares appear as early as 500 BCE and continue throughout the entire sequence while various decorated wares are separated into three major phases spanning the period from 500 BCE to 1513 CE and European arrival. Table 1 lists the ceramic types found in each test pit, their relative percentages, and overall numerical counts. The presence of both Glades Plain as well as ceramic styles representative of all Glades time periods indicates a potential habitation of the site from Glades I Late (650–750 CE) all the way through Glades IIIb (1400–1513 CE) with one exception. Ceramics from the Glades IIc period (1100–1200 CE) are not explicitly represented in this assemblage. Our ceramic evidence confirms earlier assessments that this is the most elusive period within the Glades tradition (Griffin, 1988: 140; Schwadron, 2009: 56).
Prehistoric ceramic materials.

Test Pit 2 had more than twice the amount of ceramics as Test Pit 1. This same pattern can also be observed in the frequency of faunal bones recovered from the test pits as seen in Table 2. In order to determine site ceramic density, the volume of the removed layers of soil was calculated in cubic meters for each test pit. The first test pit yielded 0.4 m3 of soil (1 × 1 × 0.4). The total number of ceramic sherds (n = 384) divided by the cubic meters of soil returns a value of 960 sherds per cubic meter. The density for Test Pit 1 is consistent with that of the previous archaeological excavation on site (980 per m3) (Schwadron, 2009: 105). The second test pit yielded 0.3 m3 of soil (1 × 1 × 0.3). The total number of ceramic sherds (n = 796) divided by the cubic meters of soil returns a value of 2653 sherds per cubic meter. According to a scale of densities constructed by Schwadron, the original site density (980 per m3) and the density of Test Pit 1 both rank as ‘light’ (1–999 per m3). In contrast, Test Pit 2 is much greater, measuring 2653 per m3, and, as a result, ranks as ‘dense’ (>2000 per m3). This major discrepancy of densities between the two test pits may indicate that the central point of the midden is in close proximity to Test Pit 2, or that earth moving activities by prior residents of the homestead artificially enhanced the midden density in this area. In either case, prehistoric ceramic materials were left on the tree island after discard, resulting in an enhancement of the land mass and elevation.
Vertebrate materials.
Glades IIIb and IIIc ceramics are correlated with the pre-Columbian occupation of South Florida by a culture group known as the Tequesta in ethnohistoric documents. Given the presence of Glades IIIb and IIIc materials, we conclude that while a Tequesta occupation of the Booth site is possible for the final period of the Glades tradition based on the presence of Glades Tooled ceramics in every level, it is unlikely that they were present during its entire usage. It should also be noted that the limited presence of pottery styles associated with the Ten Thousand Islands, such as Gordon’s Pass and Sanibel Incised, supports Carr and Beriault’s (1984) theory that the inhabitants of the east Everglades were ‘a separate tribal group’ (p. 3) from those of the west.
Bone artifacts
As noted by Karen Walker (1992) and others, points are the most common bone artifacts found at prehistoric sites of South Florida (p. 230). Three bone points were recovered from the two test excavations at the Booth site. One example may represent what Walker (1992) has called the ‘bi-pointed symmetrical variety’ that she suggests were throat gorges, used to catch fish (p. 230). We also recovered two beads made of fish vertebrae. In addition to a central perforation, both beads have a groove carved around the outer edge of the vertebrae indicating human modification. Examples of shark teeth drilled for use as pendants were recovered from a collection of artifacts in the possession of the previous landowner, and although it is likely these materials originated from the 8DA1085 midden, their exact provenience is uncertain. Multiple unworked shark teeth, including tiger shark and lemon shark (see below), were recovered from excavation, although none of these exhibited evidence of drilling or human modification. The accidental or deliberate discard of these tools and ornaments contributed to the accumulation of tree island mass.
Concretion
As previously mentioned, excavation in each test pit was concluded when excavators encountered concreted sediment. The concretion was present at a depth of approximately 40 cm below ground surface in the first test pit, and approximately 47 cm below ground surface in the second test pit. The material was difficult to penetrate and further excavation was impossible without mechanical assistance. The concretion is most likely a calcrete duricrust layer as evidenced by the fact that it was found beneath a black earth midden on a hardwood tree island surrounded by seasonally flooded sawgrass, all of which are characteristics shared by other sites with suspected calcrete layers (Schwadron, 2006). Calcrete layers normally form in drier areas; however, it is possible for limestone to undergo in situ replacement to calcrete where annual rainfall is between 1000 and 1500 mm/yr. At 1200–1700 mm, the Everglades fall within this range. This seems especially clear because of the presence of a layer of gray ‘calcite mud’ sediment encountered above the calcrete layer in our test pits (Gleason et al., 1984). Calcite mud sediment is a low-magnesium calcisiltite found in some freshwater environments of South Florida such as the Shark River Slough. It is believed to be formed by precipitation of calcite by blue-green algae in submerged algal mats (Gleason et al., 1984: 306; Gleason and Stone, 1994: 150). Graf has observed the distinctly wet/dry climate of Florida with seasonal moisture deficits and high evaporation rates can be conducive to the formation of such calcrete layers (Graf, 2008).
The blocks of calcrete recovered from the site were carefully picked apart with a chisel and hammer. A cross section of the block revealed a harder, darker calcrete layer .8 cm in thickness. It was significantly less porous than the adjacent layers and contained a great deal more shell material. However, during the subsequent dissection of the entire block, the sample was found to be surprisingly uniform, containing no material other than calcrete and miniscule fragments of shell. It is significant that no artifacts or other cultural materials were present in the concretion, although artifacts have been discovered well below similar concretion layers at other Everglades tree island sites (Graf, 2008; Schwadron, 2006). Based on these results, it seems likely that the calcrete layer resulted from local environmental factors rather than anthropogenic factors as suggested by some authors. Additional research on the formation process of calcrete layers on tree islands with evidence of archaeological occupation is needed to determine whether such concretions contributed to the permanence of tree islands or their overall lifecycle.
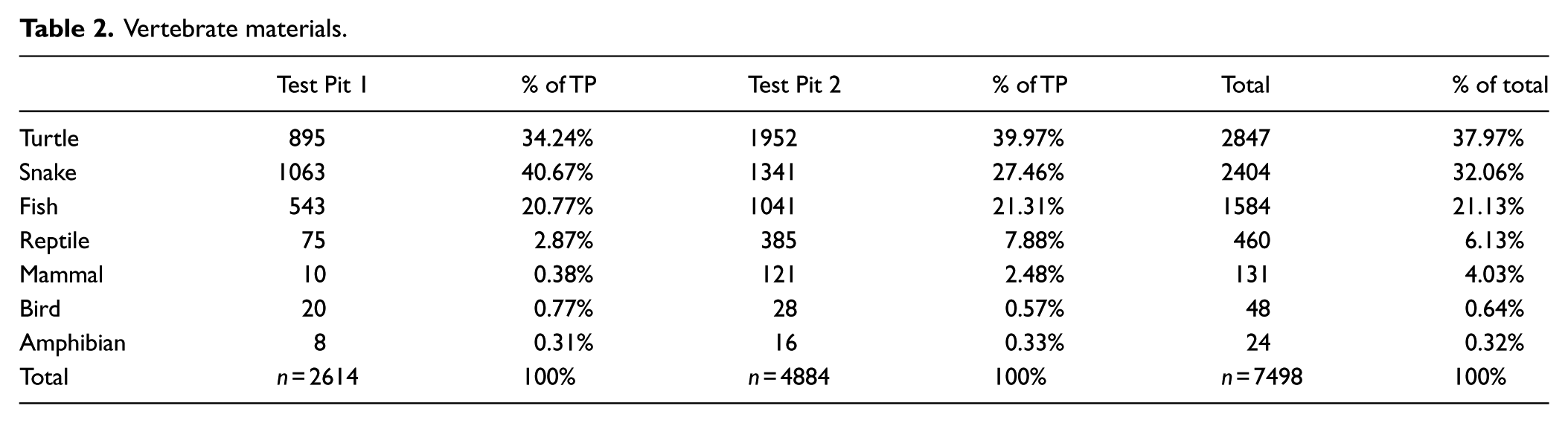
Faunal remains
Table 2 lists the results of analysis of 7498 fragments of vertebrate faunal bone that were positively identified at the class level. As a whole, snake and turtle remains (including Colubridae and Vipimidae) were the most common materials represented in the sample. Table 3 provides the list of most common taxa such as Apalone ferox, otherwise known as Florida Softshell Turtle, discovered in every level of both test pits. Fish and alligator remains were common, including two species of shark, Lemon (Negaprion brevirostris) and Tiger (Galeocerdo cuvier), represented in the form of four unworked teeth from Test Pit 1. These might represent food remains and/or raw material used for cutting tools or personal ornaments. Mammals, amphibians, and bird remains were the least common, although these bone elements are the most susceptible to degradation in the acidic soils of the tropics and their preservation may not be representative of their importance to prehistoric occupants of the site.
Most common faunal taxa.

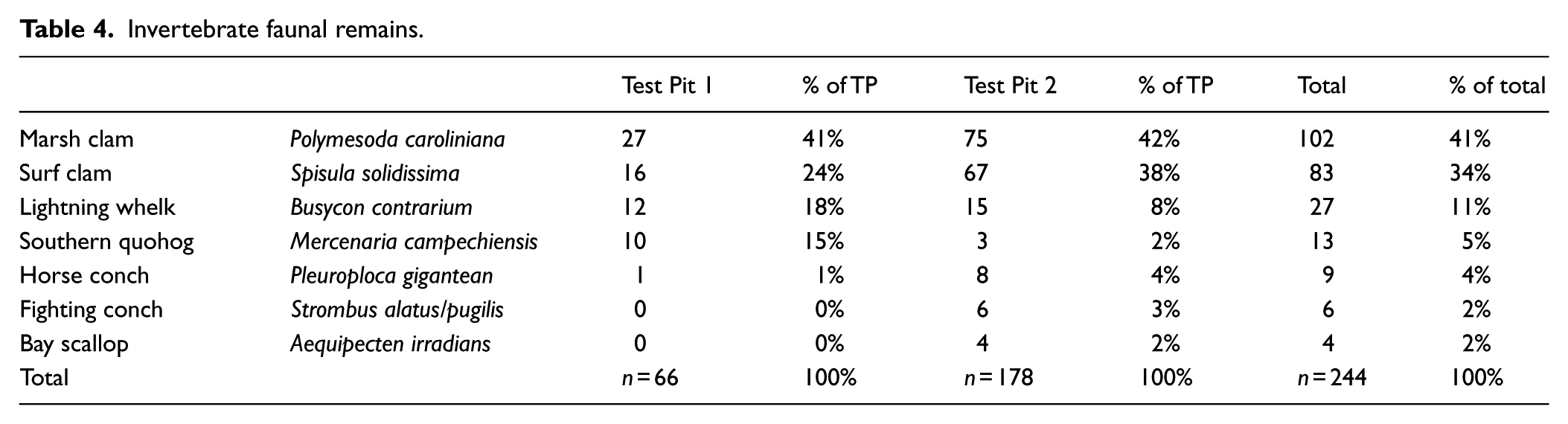
Table 4 provides a list of the 244 whole or partial invertebrate faunal remains that were positively identified at the class and species level, all of them Mollusca. The most common invertebrate was the marsh clam (Polymesoda caroliniana), an edible mollusk endemic to estuarine environments in the Everglades. Marsh clams have been found to nest in the prop root structures of mangrove forests (Rhizophora mangle) and are tolerant of a wide range of salinity conditions (Murray and Wingard, 2009). However, the overall majority of invertebrates were marine species, such as the surf clam (Spisula solidissima), the Lightning whelk (Busycon contrarium), Southern quohog (Mercenaria campechiensis), Horse conch (Pleuroploca gigantea), Fighting conch (Strombus alatus/pugilis), and Bay scallop (Aequipecten irradians). These are edible species on which the diets of prehistoric coastal populations in Florida were based. Whelk, clam, and conch shells were used for a variety of tools, and some of the elements present in this sample exhibited signs of human modification for tool use and adornment. A single very small shell bead was recovered, although the species of shell was not determined. Like the discard of exhausted bone tools, the deliberate disposal of faunal remains on the island contributed significantly to the accumulation of tree island mass and nutrient availability.
Invertebrate faunal remains.
Radiocarbon analyses
Once faunal analysis was complete, eight bone samples were selected for radiocarbon analysis with Beta Analytic. Insufficient carbon was preserved in the collagen of six of the samples, but two samples proved successful as shown in Table 5. The dates fall within the Glades IIc–III periods of prehistoric occupation, and while this is a very limited sample, it documents an occupation of at least 500 years at the Booth site prior to European arrival. Mammal bone was selected because of its relative buffering from the effects of carbonate contamination. This form of contamination can bias the bone collagen of marine organisms and those terrestrial organisms that feed on marine species, such as birds, to yield dates that are too recent. Furthermore, the highly acidic conditions of tree island middens may cause radiocarbon dates of bone collagen carbons to be skewed in favor of more recent dates because of the highly negative C12/C13 ratio. For these reasons, results should be interpreted as a ‘minimum age’ until confirmed through a larger sample size.
Radiocarbon dates.
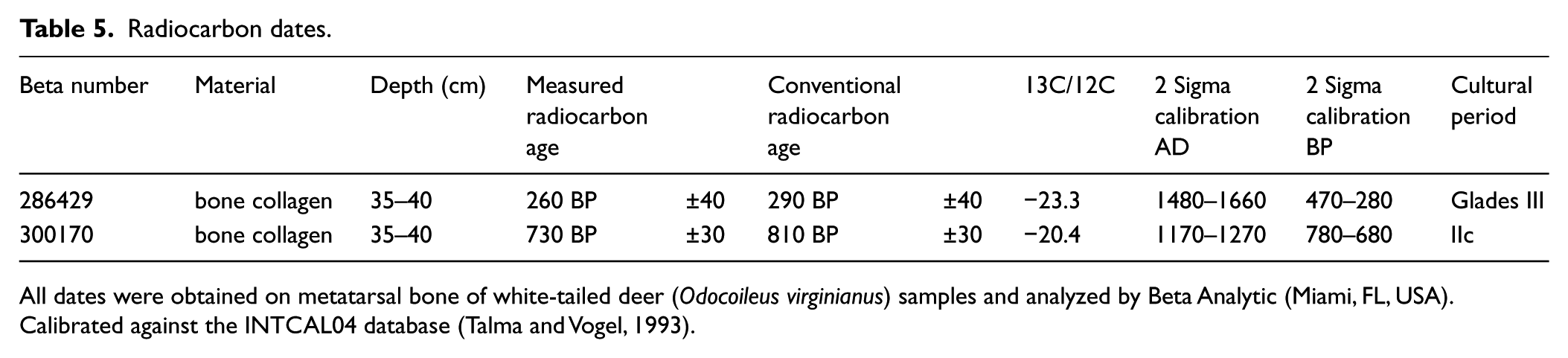
All dates were obtained on metatarsal bone of white-tailed deer (Odocoileus virginianus) samples and analyzed by Beta Analytic (Miami, FL, USA).
Calibrated against the INTCAL04 database (Talma and Vogel, 1993).
Discussion
Everglades Tree Island archaeology
Although the time available for recovery of archaeological materials from the Booth site was very limited, the data are consistent with patterns of material evidence for prehistoric occupation and modification of tree islands well documented by prior research. Starting in the 19th century, naturalists encountered ceramic sherds and other artifacts on tree islands within the Florida Everglades (Schoolcraft, 1848). In the 1930s, John Goggin made the first systematic exploration of tree island sites and was the first scientist to document the long occupational history of human habitation within the Everglades. Later investigations by a variety of researchers have confirmed the presence of black earth middens on the majority of tree islands within the greater Everglades, as well as a range of activities and periods of occupation at these sites. At three tree islands in Hollywood and Weston, Robert Carr (2002) recovered lithic artifacts dated to the Late Paleoindian period (12,000–7000 BCE) which suggest the much cooler and drier pre-Everglades habitat supported a migratory human presence (p. 195). Margo Schwadron and others have identified extensive occupation of the Everglades during the Archaic period (7000–3500 BCE) when modern environmental conditions emerged in lower South Florida (Carr, 2002: 196; Schwadron, 2006). Populations grew dramatically during the Glades Period (500 BCE–1513 CE) when native people throughout South Florida traded materials such as chert and decorated pottery, settled permanently in villages, and negotiated political arrangements with neighboring complex chiefdoms such as the Tequesta, centered at the mouth of the Miami river. Although population estimates for archaeological cultures are always of limited reliability, historians have paid special attention to the population of the New World prior to European contact. Scholars estimate the indigenous population of South Florida ranged between 20,000 and 100,000 prior to the arrival of the Spanish in the early 16th century (Carr, 2002: 198; Dobyns, 1983; Griffin, 2002; Grunwald, 2006: 22; Milanich, 1994). Even the low end of those estimations indicates humans would have had a substantial impact on the environment of South Florida over a very long period of time.
Indigenous populations settled on tree islands during the Glades period because the higher elevation provided by increased humic soils from woody vegetation provided suitable living conditions within a wetland environment rich in natural resources. There is very little evidence for the kinds of living structures that were built at these settlements, although perishable raised wooden platforms may have been used because of their documentation at contemporary archaeological sites on the southwestern coast and inland near Lake Okeechobee (Carr, 2002; Milanich, 1994; Widmer, 1988). The black dirt middens found on tree islands accumulated because of long-term human discard of mostly organic subsistence remains such as turtle and reptile bones, native fruits and grains, broken bone or shell tools, charcoal, and other waste associated with human occupation. Like the Booth site, most tree islands demonstrate a pattern of densest midden deposits on the north end of the island where a higher elevation results from natural bedrock undulations, waterflow, and the accumulation of humic soils (Carr, 2002, 2012; Griffin, 2002). Middens are also typically found toward the southern edge of an island (Carr, 2002; Griffin, 2002). These patterns suggest ancient people settled on the highest ground of the tree island, and discarded trash on the opposite end of the island, perhaps moving their occupation site periodically from one end of the island to the other, in order to allow the midden debris to degrade into usable soil. Given the routine presence of charcoal in these midden deposits, it is likely that large areas of tree islands were burned periodically in the past, perhaps to augment natural plant growth, or prepare the ground surface for habitation. A recent paleoecological analysis conducted on sediment cores supports that early human occupation of tree islands coincided with a change in plant ecology (Bernhardt, 2011). In addition to daily living, tree islands were used by pre-Columbian people for mortuary purposes, and there is historical documentation that Seminole people used specific tree islands for ceremonial purposes as well (Grandage, 2013; Simmons and Ogden, 1998; Tiger and Kersey, 2002). Each of these occupational patterns shows intentional modification of tree islands and surrounding landscapes that clearly affected the presence, distribution, and overall number of tree islands within the Everglades.
Among archaeologists, opinions on the role of these settlements within the greater pre-Columbian South Florida environmental and cultural mosaic differ. Widmer and others have suggested tree island sites represent the remains of temporary camps used for hunting in the interior of the state, but that these were not year-round settlements (Milanich, 1994; Widmer, 1988). Carr (2002) and others have argued the large size of some tree island sites (200 m in diameter) and the large number of mortuary sites suggest the presence of small villages that may have been occupied year-round or seasonally over a long period of time (p. 192). While the Booth site is classified as a smaller tree island settlement, the time depth demonstrated through ceramic and radiocarbon dating indicates human presence for a minimum of at least 500 years. The variety of settlement patterns known for tree islands, including domestic settlements, mortuary deposits, and ceremonial sites likewise suggests a full range of human activities were conducted within the interior of the peninsula, not just freshwater resource harvesting. The ubiquitous presence of marine resources such as large Mollusca used for tools, shark teeth used for ornamentation or cutting tools, and dietary remains such as surf and marsh clams at the Booth site midden and other similar tree island sites argues for a population that traveled from the EEEA in the northeastern area of the greater Everglades down to the coast and Florida Bay on a regular basis. The clams in particular would not have been edible more than a week post-harvesting, even if they were kept in containers of saltwater. Waterborne travel in wooden canoes would have been possible nearly year-round in the pre-drainage Everglades and the wide variety of faunal remains identified in the Booth site midden shows the population was well adapted to a diet of predominantly freshwater with some saltwater culinary resources such as softshell turtle and alligator from the sawgrass and marsh clam and shark from the sea (Tiger and Kersey, 2002). The well-documented marine resources of Calusa period shell middens, especially Mollusca, and the easy calories they represented for the indigenous populations of coastal southwestern Florida may have biased archaeologists against the possibilities for sustained village life in the Everglades based on a subsistence strategy of primarily interior wetland flora and fauna.
Environmental research into the formation of tree islands often emphasizes the low phosphorus available within the freshwater marshes of the Everglades and the relatively high phosphorus concentrations found on tree islands. A wide range of nutrient inputs have been tested, from sources such as climate change, bedrock mineralization, bird guano deposition, precipitation, and so on (Wetzel et al., 2005). The two main hypotheses from this line of research emphasize the role of bird guano (Givnish et al., 2008) or chemohydrodynamics (Wetzel et al., 2005, 2011) in the higher phosphorus concentrations of tree islands. Despite the well-documented presence of high phosphorus levels in human waste, especially the largely organic waste of prehistoric populations, the role of ancient human populations in the formation or augmentation of tree island phosphorus levels has not been sufficiently incorporated into environmental models of the tree island lifecycle. Given the long history of human occupation within South Florida, even prior to, and coeval with, the emergence of the Everglades, as well as the strong evidence for intentional human modification of tree island landscapes, pre-Columbian occupation added to the elevation of some tree islands which then affected sheet flow and microhydrology (Carr, 2002: 200). The discarded organic materials that eventually formed black earth middens also provided advantageous habitats for vegetation such as gumbo-limbo and hackberry, two species that are known to opportunistically flourish on archaeological sites within South Florida. The presence of these trees provided further habitat for waterbirds and other fauna that deposit guano. Humans may not have been fully responsible for the initial formation of tree islands as variations in underlying limestone geology are also correlated with the presence of tree islands, but the role of intentional burns, nutrient augmentation, and other forms of prehistoric landscape modification should be more thoroughly investigated for their potential contribution to environmental models of tree island formation.
Conclusion
The greater Everglades restoration program, one of the most costly environmental management projects in history, is guided by the principles of socio-ecological modeling. Yet, while modern human impact is at the core of current restoration models, data on prehistoric human modification of the area remain poorly understood and thus under-utilized in current environmental explanations of the earlier periods of Everglades evolution.
Within pre-Columbian New World archaeology, the full extent of human presence in the tropics has recently been rewritten completely after decades of underestimation (Bush et al., 2007; Heckenberger et al., 2003). Archaeological excavations at a prehistoric tree island midden site during 2010 and the subsequent analysis of recovered faunal and artifactual materials adds to the growing body of evidence for long-term human occupation and modification of the Everglades landscape prior to the arrival of Europeans in the New World. The role of ancient human populations in the formation or augmentation of tree islands may be under-appreciated in current models of tree island lifecycles. The research presented here suggests that high phosphorus levels in human waste, especially the largely organic waste of prehistoric populations, as well as other anthropogenic factors such as intentional burns, nutrient augmentation, and prehistoric landscape modification have not been adequately factored into current environmental models of tree island formation or the ecological evolution of the Florida Everglades. While evidence from the Booth site supports a largely anthropogenic role in tree island formation, other agents cannot be negated entirely with this study alone. Only further collaboration between environmental and social scientists dedicated to Everglades restoration and preservation will provide confirmation of the role of anthropogenic alterations of the landscape.
Footnotes
Acknowledgements
We gratefully acknowledge comments on an earlier version of this paper from Jonathan Grandage, Maureen Mahoney, Leonel Sternberg, and two anonymous reviewers. Any errors remain the responsibility of the authors.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.