
Abstract
Integration of geomorphological, stratigraphic, malacological, sedimentological and micropalaeontological techniques and 14C dating allows us to characterise the processes and evolution of the coastal barrier–lagoon system of Valencia (Spain), from the middle Holocene to the historical epoch, as well as the responses to global climate events. Four stages are recognised. Phase 1: around 8240 ± 80 cal. yr BP, a brackish lagoon of moderate energy and in restricted environment was formed, with an energy peak that could correspond to the maximum Holocene marine transgression. Dating (8240 ± 80 cal. yr BP) carried out in peat corresponds to a cold cycle and low water levels in inland lakes of the western Mediterranean. Phase 2: from 6450 cal. yr BP to 3710 ± 130 cal. yr BP, a lagoon remained, in restricted environment and connected with the sea, but with a notable energy decrease and recurrent saturation processes similar to those described in other Mediterranean continental lakes. This phase is contemporaneous with a period of increase in the aridity trend and global cold cycles. Phase 3: from 3710 ± 130 cal. yr BP, a brackish lagoon without marine connection was formed. Towards 820 ± 90 cal. yr BP, a shift to a totally isolated lagoon environment took place (changing from brackish lagoon to freshwater). This process is coeval with a palaeohydrological phase of high flooding frequency in the river flood plains of Spain and Southern France. Phase 4: freshwater lagoon environment becomes a widespread flood plain. During a phase of high frequency and magnitude of floods (‘Little Ice Age’), the flood plain is formed on the top level of the sequence. Phases and processes recorded in sedimentation could be placed in relation with global mid-to-late Holocene events.
Keywords
Introduction
The Mediterranean coastal lagoon–barrier systems were formed after the last glacial episode, with the global rise in Holocene sea level and formation of morphologically indented shorelines and estuaries along the low-lying alluvial coasts. Due to the significant tectonic activity, the Mediterranean region shows considerable variation in the altimetric position of sea level during the Holocene. According to Pirazzoli (1991, 2005), the altimetric maximum (about 2 m a.s.l.) was reached around 6000–5000 yr BP, subsequently followed by a gradual descent to the current level. When the sea level stabilised, the sediment deposits from rivers (ordinary flows and floods) formed gravel beach barriers and spits around river mouths. Behind these barriers, enormous volumes of clay and silt draped and buried the drowned glacial coastline (Stewart and Morhange, 2009). Complex associations of flood plain, coastal lagoons and deltas like those of the rivers Rhône (Arnaud-Fasseta, 1998; Boyer et al., 2005; Bruneton et al., 2001), Tiber (Amorosi and Milli, 2001; Bellotti et al., 1994), Ombrone (Bellotti et al., 2004), Arno (Amorosi et al., 2008; Rossi et al., 2011), Turia–Júcar (Carmona and Ruiz, 2011), among others, were formed during this period. Although the stabilisation of sea level and availability of sediment are important variables in this process, interdisciplinary research has brought to light the importance of other fundamental factors, such as the palaeogeography of the lagoonal environment and the specific fluvial and marine sedimentary dynamics in each case study. In historical times, there were still many wetland environments on the low-lying alluvial Mediterranean coasts, which were only completely filled in quite recently, during the Middle Ages and Modern Period (Grove and Rackham, 2001).
In addition to works related with eustatic sea level curves and coastal geomorphological dynamics, recent research has revealed the climatic variability of the Holocene. These variations have been very frequent (multidecadal to multicentennial timescales) and their fluctuations between warm and cold and humid and arid cycles are well documented in the scientific literature (Bond et al., 1997, 2001; Denton and Karlén, 1973; Mayewski et al., 2004; Wanner et al., 2008, 2011). In the North Atlantic region, Holocene climate events recorded in different palaeoclimate records have been shown to be correlated in time and closely related with glacial advance and retreat phases (Denton and Karlén, 1973). Moreover, several studies indicate that they are also synchronous with changes recorded in sea surface temperature in lower latitudes (Cacho et al., 2001; DeMenocal et al., 2000).
Specifically, in the Mediterranean basin extreme hydrological events have been found to be an inherent component of past (Holocene) and present hydrology. For this reason, an important part of palaeoenvironmental Holocene research has focused on identifying phases of high frequency and magnitude of floods related with climatic variability (Benito et al., 2008, 2015; Macklin and Woodward, 2009; Macklin et al., 2006; Thorndycraft and Benito, 2006a, 2006b). In the Mediterranean basin, several research works document a centennial to multicentennial see-saw pattern in flooding, indicating bipolar hydroclimatic conditions during the Holocene. In the western Mediterranean region, periods with more frequent floods coincide with transitions to cooler and wetter climates, while flood frequency in Northern Africa is linked with drier climate and in the eastern Mediterranean with wetter conditions (Benito et al., 2015).
On the other hand, in the western Mediterranean area, sedimentary records on valleys and flood plains have provided ample data on geomorphologic responses to Holocene hydroclimatic oscillations. In alluvial environments from the NE of Spain, different phases of incision and accumulation are identified on slopes and valley bottoms (Constante et al., 2011; Gutiérrez and Peña, 1998; Peña et al., 1993, 1996; Sancho et al., 2008). Palaeohydrological research in the Medjerda valley (Northern Tunisia) has identified alternating phases of aggradation and soil formation (Fletcher and Zielhofer, 2013; Zielhofer and Faust, 2008; Zielhofer et al., 2004). Likewise, Holocene fluvial research in the Italian Peninsula has provided large amounts of data on great floods in the Tiber river (Camuffo and Enzi, 1995) and Holocene geomorphic activity related to climate change (Giraudi et al., 2011; Piccarretta et al., 2011). Finally, several morphogenetic phases related to Holocene climatic variability have been identified in the lower Rhône basin (south of France) (Arnaud-Fasseta et al., 2010).
We should also stress that palynological, sedimentological and geochemical analyses, mainly carried out on continental lakes of the Western Mediterranean, reveal changes in vegetation dynamics, prominent peaks of xerophytes and, particularly, water level fluctuations which have been related with global climatic oscillations in the Holocene (Carrión, 2002; González-Sampériz et al., 2006; Jalut et al., 2000; Jiménez-Moreno et al., 2015; López-Blanco et al., 2012; Magny et al., 2007, 2009, 2011; Moreno et al., 2008). However, for the past 2500 years the answer seems to have been conditioned by both Holocene climatic variability and by human action (Burjachs et al., 1997; Butzer, 2005; Pérez-Obiol et al., 2011; Riera et al., 2004).
Coastal lagoons and flood plains are environments highly vulnerable to global warming and the rapid rise in sea level, as they are very exposed to processes such as flooding, subsidence, sediment scarcity and coastline erosion (Anthony et al., 2014; McCarthy et al., 2001). Our study focuses on the coastal lagoon of Valencia, located in an alluvial plain of the Spanish Mediterranean coast. The morphology of this plain is characterised by alternating small coastal barrier–lagoons, flood plains and deltas of river system tributaries (Figure 1). For this reason, the Holocene evolution is closely linked with specific lagoonal and marine processes as well as fluvial dynamics. These coastal lagoon environments preserve an almost continuous record of Pleistocene and Holocene intervals (Torres et al., 2014; Viñals, 1991). On a Holocene timescale, marine, continental and transitional environments have been characterised in several cores. Many studies have addressed eustatic changes in sea level (Blázquez, 2005; Blázquez and Usera, 2010; Torres et al., 2014; Viñals, 1991; Viñals and Fumanal, 1995) and others have provided data on geomorphological and environmental changes on the Holocene (Blázquez and Ferrer, 2012; Carmona and Ruiz, 2011; Ferrer and Blázquez, 2012; Goy et al., 2003; López-Buendía et al., 1999; Marco-Barba et al., 2013a, 2013b; Mediato and Santisteban, 2006; Ruiz and Carmona, 2005; Zazo et al., 2008).
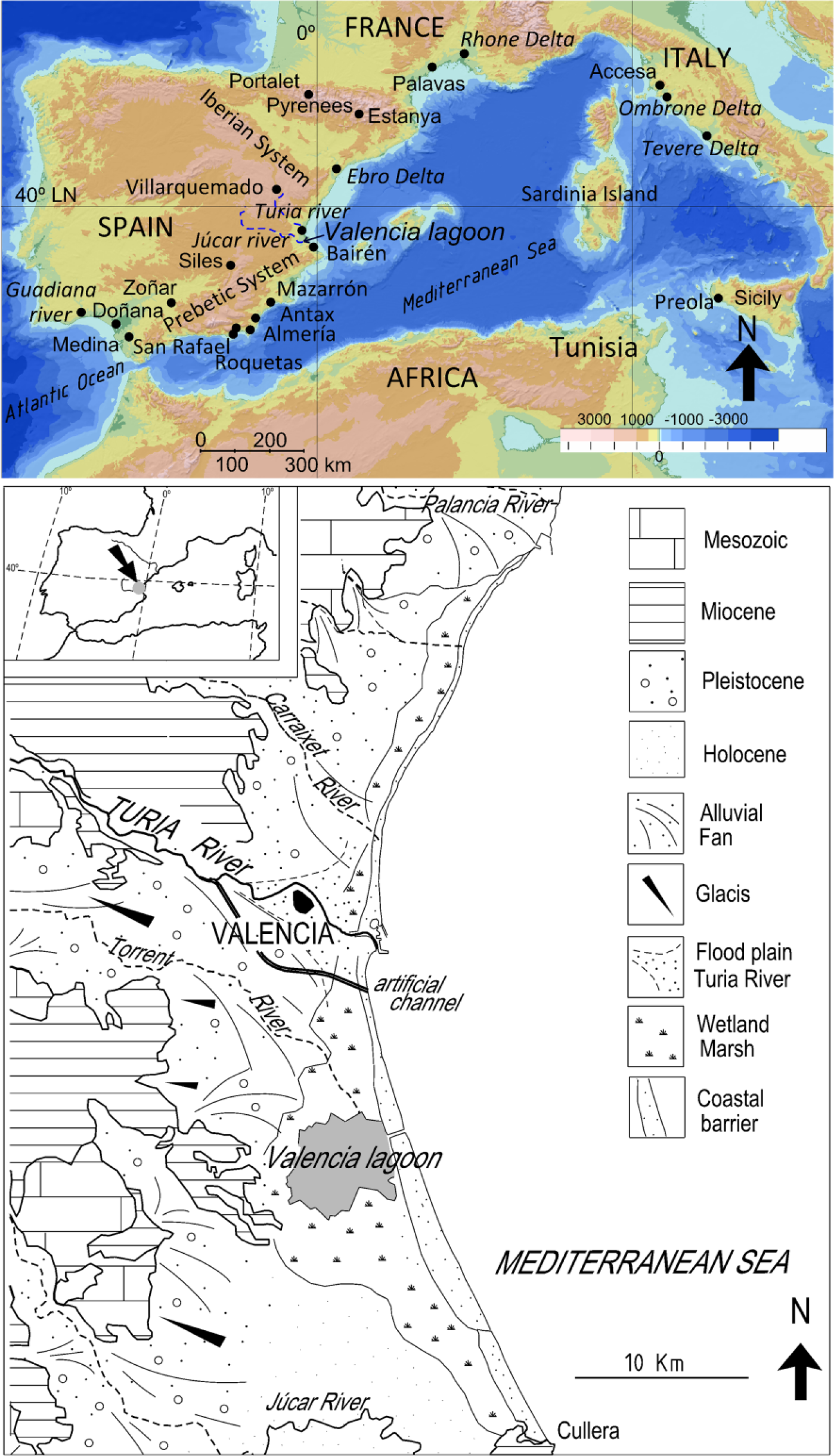
Upper: location of the lagoon of Valencia in the Iberian Peninsula and sites mentioned in the text. Guadiana river (Fletcher et al., 2007); Doñana (Jiménez-Moreno et al., 2015); Antas, San Rafael, Roquetas de Mar (Pantaleón-Cano et al., 2003); Villarquemado (Aranbarri et al., 2014); Zoñar (Martín-Puertas et al., 2008); Siles (Carrión, 2002); Mazarrón (Navarro-Hervás et al., 2014); Stanya (Morellón et al., 2009); Medina (Reed et al., 2001). Lower: Morphological patterns of the Valencian coastal plains.
Assessing future impacts on these highly dynamic coastal environments calls for appropriate knowledge of their processes, as well as the changes they have undergone in different timescales. Valencia’s barrier–lagoon system is one of the most important on the Spanish Mediterranean coast. Previous research showed that inland deltas and flood plains of the tributary rivers (Turia and Júcar) formed the infill of the late-Holocene palaeolagoon (Carmona and Ruiz, 2011, 2014; Rosselló, 1972; Ruiz and Carmona, 2005). In this context, the main aims of this article are as follows: (a) to analyse the geomorphological processes and evolution of a sector of the lagoon from the middle Holocene to the historical epoch and (b) characterise the possible responses of the geomorphological system to middle–late Holocene climate events. To this end, we rely on a detailed geomorphological study of the coastal barrier–lagoon and flood plain systems and analysis of the sedimentology, malacology and foraminifera of a well dated record (14C) obtained in the northern sector of the lagoon.
Geographical setting of the study area
The coastal lagoon of Valencia is located in the transition area of the Iberian system and External Prebetic zone of the Betic Cordillera, affected by extensional tectonics during the Neogene period (Simón, 1984; Vegas, 1992) (Figure 1). The distensive processes compartmentalised the coast into staggered blocks towards the coast, orienting a NE-SW strip of Pleistocene alluvial fans. The distal part of the fans overlaps the Holocene coastal alluvial plain formed by flood plains and lagoon–barrier systems (Figure 1). From the dynamic perspective, coastal processes are controlled by waves, wind and longshore drift (N-S). The tidal influence, with average daily oscillation around 15 cm, is negligible. Sediment delivery from the river mouths to the coast has shaped successive systems of wave dominated deltas slightly prominent seawards in the main rivers (Ruiz and Carmona, 2005). The barrier beaches of these deltas link to the coastal lagoon–barrier systems whose Holocene dynamic has been widely studied on the adjacent continental shelf (Albarracín et al., 2012; Alcántara et al., 2012). The rivers Turia and Júcar flow across the coastal plain, with extraordinary floods having exceeded 3700 and 15,000 m3/s, respectively (Carmona and Ruiz, 2000). During the Holocene, a common inland delta of both rivers would be the basis of the formation and filling of the lagoon (Rosselló, 1972; Ruiz and Carmona, 2005). Over recent millennia, the riverine inputs of these watercourses have formed two contiguous flood plains in the lagoon’s continental perimeter. The lagoon now has a water surface of 25–28 km2 and a maximum depth around 160 cm (Rosselló, 1995).
Geomorphological features of the coastal plain
Interpreting georeferenced historical maps (18th and 19th centuries), LiDAR (Light Detection and Ranging) altimetry (MDT) and a photomosaic (orthophoto) of year 1956 of the lagoon environment allowed us to characterise a geomorphological model consisting of several sub-environments such as lagoons, freshwater marshes, flood basins, flood plains, fluvial levees and coastal sand barriers (Figures 1 and 2).

Left: Digital Elevation Model LIDAR (Light Detection and Ranging) 1 m. Right: Geomorphological map of the coastal barrier–lagoon of Valencia through time. The lagoon of 19th century corresponds to Sea Chart of 1877 (Ruiz and Carmona, 2005). The lagoon of 18th century corresponds to ‘Mapa de reposición de mojones de los límites de la Albufera’ (Carmona and Ruiz, 2014). Location of other previous cores around the Valencia lagoon discussed in the text.
The lagoon
Historical cartography (the 300 last years) enabled us to analyse the evolution of the lagoon perimeter and transformation of the environment into a flood plain. The geomorphological processes are related to lagoonal inland delta progradation and accretion of alluvial levees on the shores of the lagoon. These processes determine a progressive evolution towards continentalisation. Change starts with the compartmentalisation of the lagoon into topographically depressed areas (flood basins) with different degrees of siltation. This process causes the emergence of a diversity of transitional wetlands. Many of these environments are recently formed. According to historical cartography, in the 18th century the lagoon shore extended roughly 4–5 km further north than it does today and, in that era, the freshwater marshes of the lagoon’s perimeter extended several km northwards, where they connected with wetlands of the medieval epoch (Carmona and Ruiz, 2014) (Figure 2). Wetland environments around the lagoon have undergone an alluviation process and have been drained, cultivated and built upon in historical times (Carmona, 1990; Glick, 1970; Ruiz, 2002; Sanchis, 2001).
The current assemblage of foraminifera in Valencia’s lagoon has not yet been studied. In any case this lagoon is highly anthropised today and current foraminifera distribution would not serve as a reliable indicator to compare with that present in a natural environment. This is particularly true for assemblages found in levels prior to the 18th/19th century. Ostracod palaeointerpretations clearly indicated a previous brackish stage (prior to 1850) that changed to a freshwater environment (Marco-Barba et al., 2013a). Now, it is an oligohaline coastal lagoon. According to Soria et al. (2005), the lagoon currently is freshwater and its conductivity level does not exceed 2000 µS/cm. The lagoon waters correspond to the following relations: SO4 = ><C1–>AIK and Na+><Ca++><Mg++>K+ (Vicente and Miracle, 1992).
The flood plains
The long stretch of low or zero gradient (between 0 and 1 m a.s.l.) around the lagoon today seems to be compartmentalised by gentle topographic prominences of elongated layout, corresponding to the fluvial levees of ancient deltaic mouths of the Turia river (Brosquil) and the ephemeral stream of Catarroja. Among the levees, there are flood basins of plane topography depressed by several decimetres. Alluvial surfaces of convex morphology extend inland, constituting the continental flood plain of the affluent rivers. All these wetland sub-environments are sheltered from marine influence by the medieval alluvial ridge of the Turia (la Punta) and its extension into the narrow coastal barrier (Carmona and Ruiz, 2014; Ruiz and Carmona, 1999).
Today, the entire perimeter of the lagoon is affected by overflowing of the main tributary river systems of Turia and Júcar and the ephemeral stream of Catarroja. Thus, the wetland environments of the lagoon’s perimeter appear buried under a flood plain layer roughly 2 m thick, deposited mainly in historical times, especially in Medieval and Modern intervals (Carmona, 1990; Carmona and Ruiz, 2011; Carmona et al., 1994). It is important to highlight that river flooding events have been a recurrent process since the late-Holocene. In the palaeohydrological and geoarchaeological record of the Turia river flood plain, we identified two phases of high flooding frequency and magnitude; the oldest corresponds to a period from Bronze Final period (2750 cal. yr BP) and the second, more recent, to Islamic times (11th century; 1000 cal. yr BP) (chronology derived from archaeological remains) (Carmona and Ruiz, 2011) (Figure 3). A third phase, corresponding to the ‘Little Ice Age’ (LIA), was analysed by means of an approximately 700-year period database (historical chronicles) in the Júcar and Turia rivers. All these phases have been linked to global climate changes in the Holocene (Carmona and Ruiz, 2011; Ruiz et al., 2014).
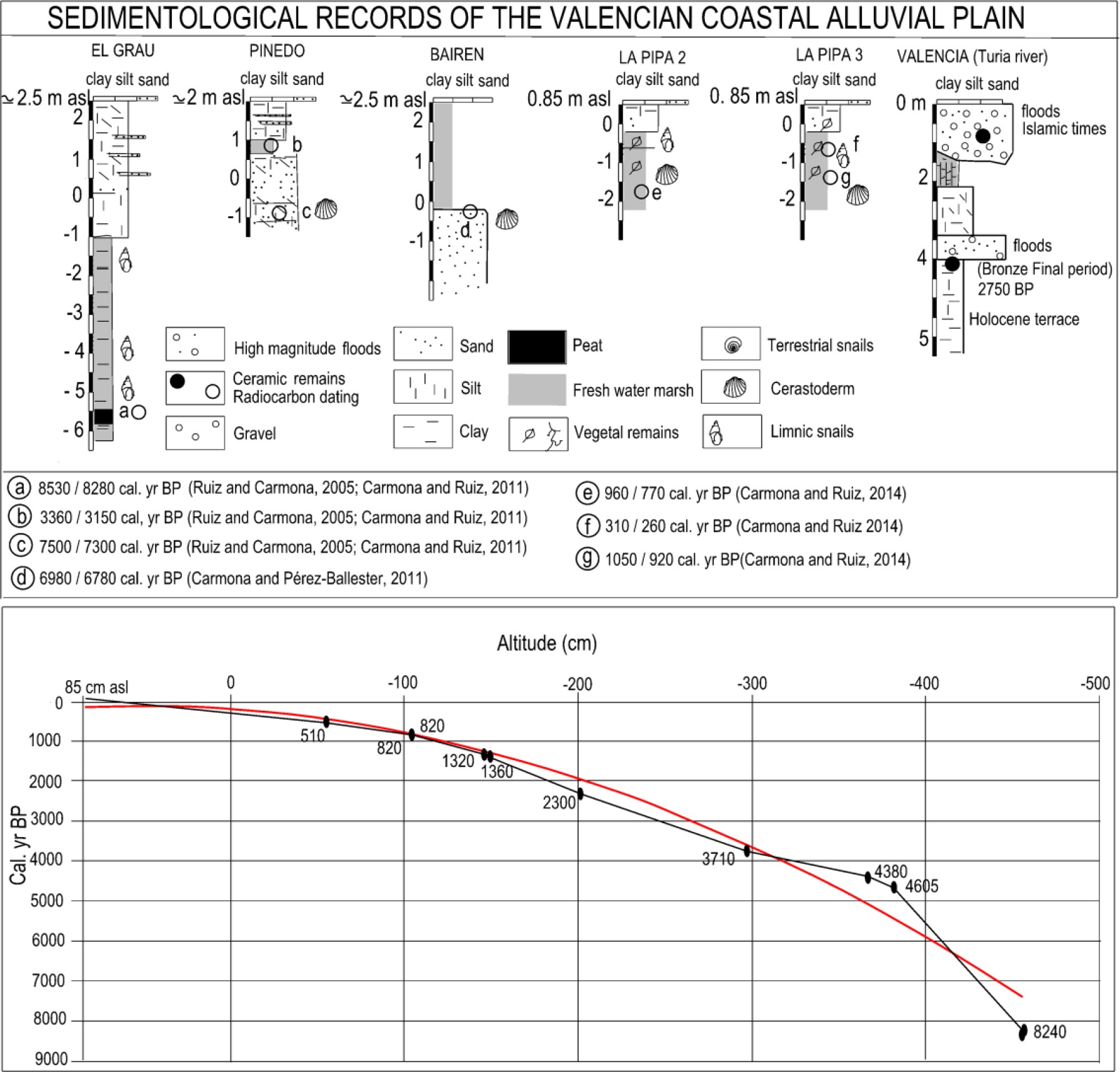
Upper: Other previous sedimentological records from the Valencian coastal alluvial plain. Lower: Composite time–altitude (cm) model based on 14C dating (Table 1) and calibration curve following Reimer et al. (2013) of SAL/S1 and SAL/S2 cores.
The coastal barrier
The coastal barrier that closes off the present-day lagoon in Valencia starts from the delta mouth of the Turia, whose sedimentary inputs are distributed by the drift current (N-S direction) along the regularised coastline and on a gently sloping continental shelf (Rosselló, 1972, 1995; Sanjaume, 1974). At the northern end (Turia river delta) the barrier is totally anthropised by the Valencia city port installations. To the south, the barrier presents different morphologies closely related with Pleistocene calcarenite outcrops and neotectonics (Carmona and Ruiz, 1999). The stretch that cuts off the study area from marine influence, Pinedo-El Saler, is formed by a narrow, single ridge 330 m in width and 2 m a.s.l. in height (Figure 2). In this sector (towards the more inland continental area), we examined the stratigraphy of the Holocene coastal barrier in an excavation (Pinedo core in Figure 3). This sequence is composed of a barrier whose base is formed by sandy sediments of a beach dated around the 7500–7300 cal. yr BP. This beach level is buried by grey silt from a backbarrier coastal lagoon (dated around 3360–3150 cal. yr BP) and dune sand sediments (undated). The top level consists of layers of sandy silts from the flood plain of the River Turia (Carmona and Ruiz, 2011).
The Saler barrier section extends southwards with a morphology of two prograding ridges, of greater width (980 m) and height and dune alignments of up to 5 m (Sanjaume, 1974). In a core extracted in the El Palmar area, Santisteban et al. (2009) and Marco-Barba et al. (2013b) identified several Holocene barrier units which the authors correlated with progradation series of the Ebro Delta (Somoza et al., 1998) and with the Holocene prograding beach ridges of the south-eastern Spanish coast (Goy et al., 2003; Zazo et al., 2008).
Methods
The methodology behind this study is based on the integration of geomorphological, stratigraphic, malacological, sedimentological and micropalaeontological techniques (foraminifera, charophytes, ostracods, etc.) and 14C dating.
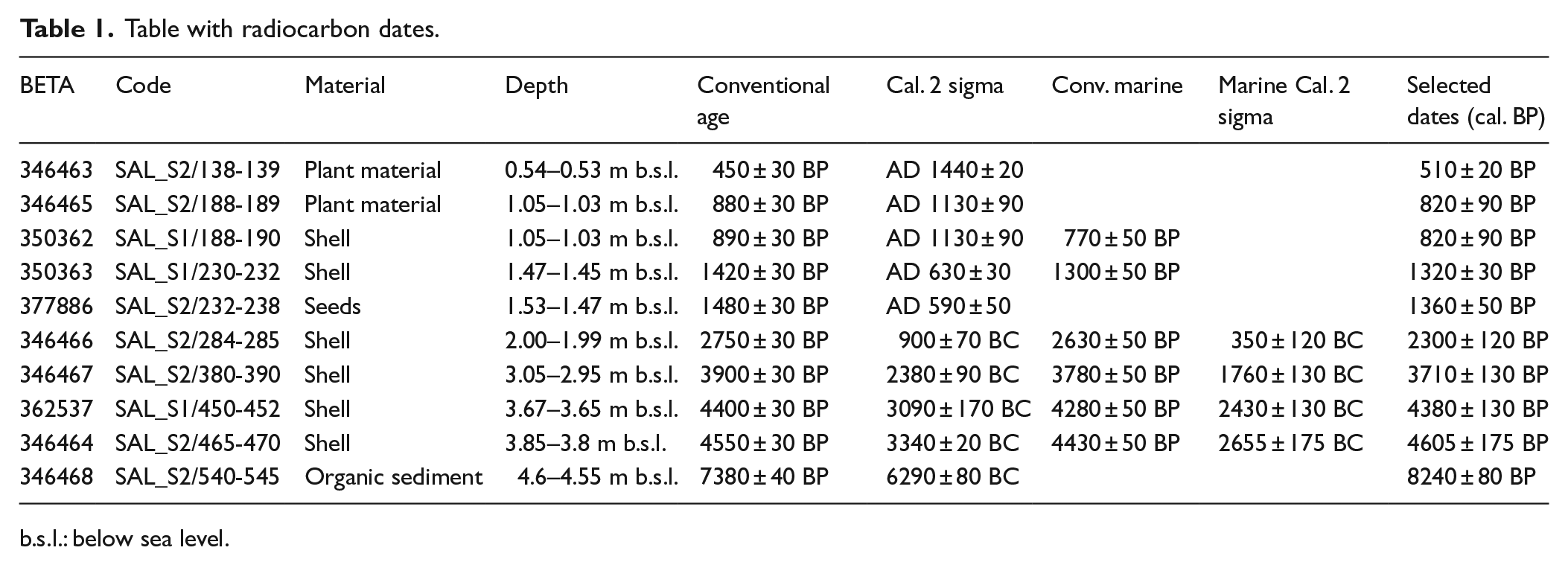
The geomorphological study was carried out by identification of current forms and processes in the field, photo-interpretation of 1956 aerial photomosaic 1:33,000 scale and microrelief analysis using a 1- and 5-m LiDAR Digital Elevation Model (DEM) (by TERRASIT and CNIG). The use of 1956 aerial photographs (known as the ‘American flight’) made possible to recognise landforms just before rapid urban development during the second half of the 20th century. Historical cartography was used to identify morphological features of the coastal lagoon, freshwater marshes and river flood plains in the Modern Era. Two continuous sedimentary sequences (cores SAL/S1 and SAL/S2) 0.85 m a.s.l., 2 m apart from each other were extracted (in 2013) using a manually driven 50 cm × 5 cm ‘Russian’ corer (peat sampler). Core SAL/S1 (39°22′53.45″N, 0°21′43.24″W) was 500 cm long, while SAL/S2 core (39°22′53.55″N, 0°21′43.34″W) was 570 cm. The rationale behind the double coring was to multiply the chances to obtain material suitable for radiocarbon dating and to increase the sediment available for multi-proxy palaeoenvironmental studies. The sequences were correlated following sedimentary parameters although, due to the proximity of the coring points, corresponding sedimentary phases were found at equal depths. A total of 10 radiocarbon dates were obtained from different material from both cores, whose upper extreme was considered to correspond to a contemporary date (Table 1). Analytical data (sedimentology and micropalaeontology) presented in this work correspond to core SAL/S2.
Table with radiocarbon dates.
b.s.l.: below sea level.
Visual descriptions of the texture, colour and identification of the malacofauna and molluscs in the core served to initially distinguish between brackish and freshwater assemblages. The micropalaeontological study was based on the benthic foraminifera and other microfossils (charophytes, ostracods, etc.). Systematic sampling involved collection at intervals of 10 cm along the core. The material was washed through sieves with mash diameters of 0.063 mm. A total of 57 samples of approximate volume 100 cm3 were examined for micropalaeontological analysis. The analysis was conducted at the micropalaeontological laboratory of the Catholic University of Valencia. Foraminifera tests were picked using a binocular stereomicroscope after flotation (when necessary) in dense liquids such as trichloroethylene and until a representative number of 300 individuals per sample were obtained. The resulting data were quantitatively analysed, and Diversity Index (Shannon and Weaver, 1949), Dominance, Equitability and Fisher’s Alpha (Fisher et al., 1943) were calculated in order to establish the composition, proportional abundance and dominance of the species identified in the samples. The percentage of allochthonous species is calculated according to the autoecology of the species and its conservation status and especially according to the association of living foraminifera found in the inner shelf on adjacent sectors (López-Belzunce et al., 2014; Usera and Blázquez, 1997). The authochthonous taxa comes from the association studied in the current marshes and in Holocene sedimentary records of similar lagoons and nearby, such as Torreblanca (Guillem, 2008), Peñíscola (Usera et al., 2006), Oliva-Pego (Torres et al., 2014), Javea (Fumanal et al., 1993), Albufereta (Ferrer and Blázquez, 2012) and Elx (Blázquez and Usera, 2010). Different palaeoenvironments have been determined, based on the foraminiferal content classified according to Loeblich and Tappan (1988). The AphiaID number from the World Register of Marine Species (WORMS: http://www.marinespecies.org/) has been incorporated for every described species (Table 2). Determination of the foraminifera assemblage was based on the relative abundance of the dominant species.
Micropalaeontological data. Relative abundances of the principal assemblages. Allochthonous species in bold.

The sedimentary analysis core was carried out by the Geomorphology Laboratory at the Valencia University Geography Department. Systematic sampling involved collection of the sediments at intervals of 10 cm interspersed between the micropalaeontological samples. The colour was checked with dry samples and the Munsell charts. Texture was determined by dry sieving (sand fraction > 4 phi) and pipette method (silt and clay fractions). Grain size parameters were calculated with Gradistat software package (Blott and Pye, 2001). The sedimentological information was completed with binocular stereomicroscope observations of sand grains.
Cores SAL/S2 and SAL/S1 were taken in the freshwater marshy and flood plain environments bordering the northern continental sector of the lagoon at an absolute height of 0.85 m a.s.l. (Figure 4). A total of 10 radiocarbon dates (Table 1, Figures 3 and 4) were obtained from different material from both cores. Calibration of shell material dates was performed by BETA Analytic Inc. and incorporated local reservoir correction. However, marine correction from SAL/S1 188-190 and SAL/S1 230-232 samples was ruled out because of their correspondence with dates obtained from plant material (SAL/S2 188-189) and seeds (SAL/S2 232-238) at similar depths (Table 1, Figures 3 and 4). The increase in water salinity (corresponding to a higher marine influence), the sediment accumulation rate and the integrity of the time–depth vector made us incorporate the local reservoir correction for the last four radiocarbon dates obtained from shell material. The information is presented in chronological dates cal. yr BP.
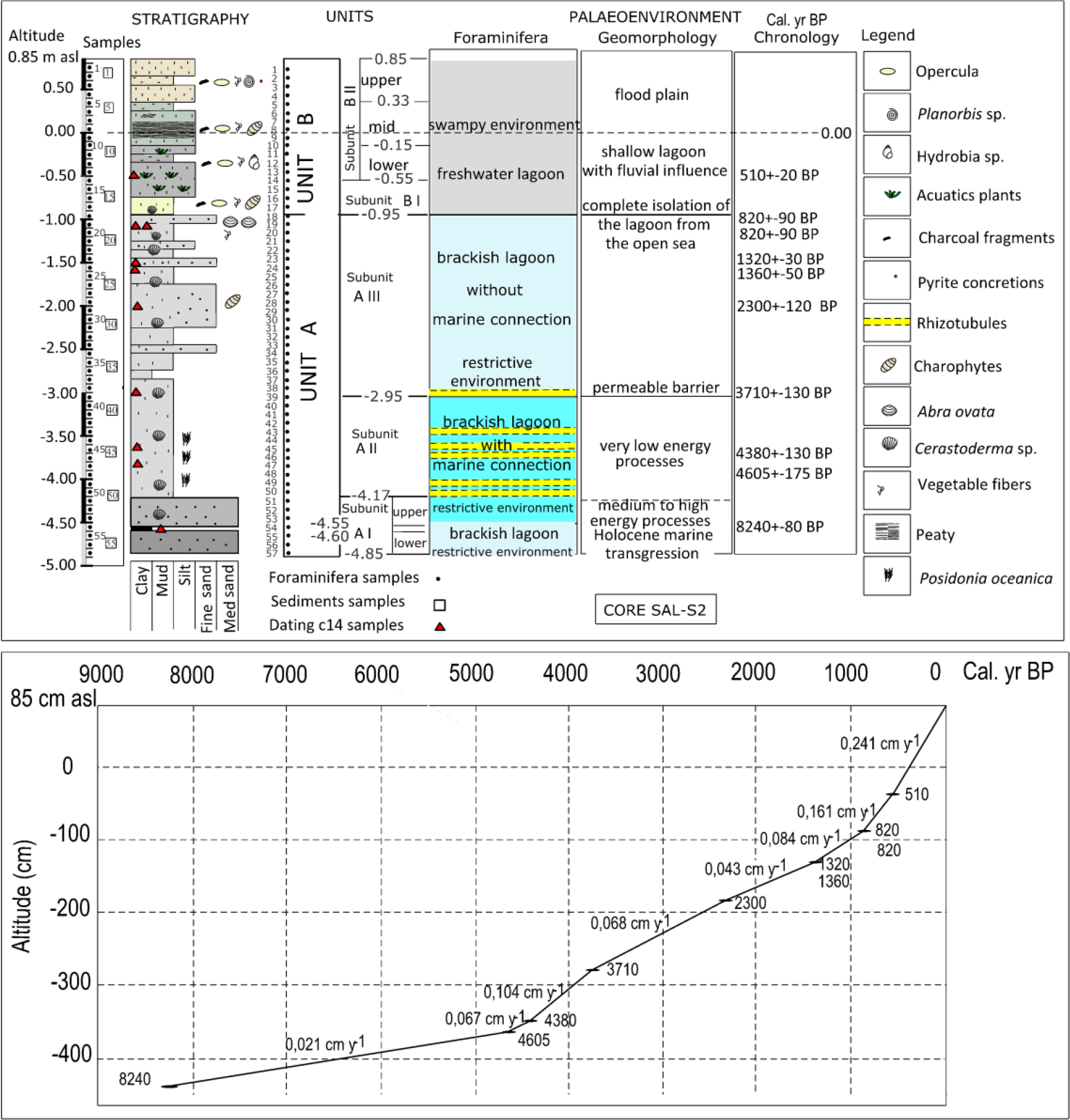
Upper: Lithostratigraphical units, foraminiferal interpretation, geomorphological processes, chronology and succession of palaeoenvironmental phases throughout the lagoon record SAL/S2. Lower: 14C dating (Table 1) and accretion rates of SAL/S1 and SAL/S2 cores.
All spatial information was georeferenced in CAD (Microstation V8i) on a vector-based digital map on a scale of 1:5000. This base was organised as an ArcGIS (Geographic Information System) and used to plot the geomorphological mapping, stratigraphic study and analysis of geomorphological changes.
Results: Sedimentary record
Visual descriptions and biofacies and sedimentological analyses of the cores initially enabled us to distinguish two main sedimentary environments: units A and B (Figure 4). In the SAL/S2 core, unit A appears at the base, between 4.85 and 0.95 m below sea level (b.s.l.), as a grey coloured silty clay textured layer with an abundance of Cerastoderma sp. shells. Unit B corresponds to the top layer of the sequence (between 0.95 m b.s.l. and 0.85 m a.s.l.) and is a greyish brown clayey silt stratum with variable organic matter content, remains of aquatic vegetation and freshwater gastropod fauna. This top layer unit was previously studied and dated in two nearby cores (Figure 3) (Carmona and Ruiz, 2014). Furthermore, the stratigraphic column incorporates the sedimentological, micropalaeontological and chronological data (Figure 4). The sedimentological information, foraminifera and chronological data of both units are described in detail below.
Environmental facies of Unit A
The sedimentological features and micropalaeontological study of unit A allow us to distinguish three sub-environments: A I, A II and A III (Figure 4).
Subunit A I
Corresponds to basal sector (4.85–4.17 m b.s.l.) in which two levels are distinguished (lower and upper), separated by a peat layer.
Lower level A I
From 4.85 to 4.55 m b.s.l. Foraminifera and sediment samples: 57–54. A stratum 30 cm thick, sandy mud texture, with a sand percentage between 27% and 37% and dark to very dark grey in colour (2.5Y 4/1). The sand fraction consists of heterometric calcite and quartz grains, medium to fine texture, of different colours and shell fragments of Cerastoderma sp. and gastropods (Hydrobia sp.) which increase towards the top layer (samples 54 and 55). The microfossils study indicated the presence of species of foraminifera of brackish and restricted waters: Ammonia tepida (Cushman), with 82.7% average relative abundance per subunit, Haynesina germanica (Ehrenberg), with 8%, and Cribroelphidium excavatum (Terquem), with 2.9%, especially at the base (sample 57), as well as the ostracod Cyprideis torosa (Jones), corroborating the environment. At the top, some eroded tests of marine species were recorded, such as Triloculina trigonula (Lamarck), with 1.2% of relative abundance per subunit and Quinqueloculina seminula (Linné) with 0.9%. Results of the diversity indices in this level indicated an environment with lower diversity, as corresponds to restricted waters. The medium value of 10 different species per sample was obtained. Fisher’s alpha diversity presented higher values at this level (Table 2). As regards Dominance, all samples have high values, close to 1, indicating the presence of dominant species such as the species mentioned previously. The equitability values are consequently lower with the values of Dominance. The subunit has a top layer rich in organic matter level (4.60–4.55 m b.s.l.) dated around 8240 ± 80 cal. yr BP. The lower level A I may be interpreted as a brackish lagoon of moderate energy in restricted environment.
Upper level A I
From 4.55 to 4.17 m b.s.l. Foraminifera and sediment samples: 53, 52, 51. Towards the top layer and in sharp contact with the peat, there is a level 38 cm in thickness, sandy mud or muddy sand texture, grey coloured (5Y 5/1), waterlogged and with abundant Cerastoderma sp. shell fragments (several centimetre diameter). The sand fraction (31%–57% of the total sample) presents a considerable increase in bioclasts, with abundant fragments of Cerastoderma sp., slightly rounded and some rhizotubules. The sands are predominantly quartz and medium to fine texture. The dominant foraminiferal assemblage is formed by a mixture of brackish organisms: A. tepida (82.7%), H. germanica (8.9%), Pseudolachlanella eburnea (D’Orbigny) (1.9%), C. excavatum (1.8%), with another of normal-marine salinity foraminifera, lowest representation, formed by Adelosina bicornis (Walker & Jacob) (1.18%), Adelosina laevigata (D’Orbigny) (0.5%), and so on (Table 2). This assemblage indicates a restricted brackish water environment with marine communication and shows the continuity of the association registered to the top of the previous level. From the taphonomic standpoint, the tests of marine origin are less well preserved compared with the brackish, which would indicate possible transport of these tests from the open sea. The high sand content in the sample and abundance of bioclasts denote medium to high energy processes.
Regarding the diversity indices, they are very similar to lower level A I. Although there are no differences in the values of diversity in Subunit A I analysed, differences were observed in the conservation of tests: the upper level presents more eroded tests. Furthermore, at this level the percentage of allochthonous taxa increases from 4.8% (lower level) to 7.2%.
Subunit A II
From 4.17 to 2.95 m b.s.l. Foraminifera samples: 50–40; sediment samples: 50 to 38–39. The textural shift towards muddy sediments characterises Subunit A II: a waterlogged sedimentary stratum, light grey in colour (2.5Y 7/1), with a muddy texture and high clay content (around 74%). At this subunit, abundant shells of Cerastoderma sp. several centimetres in diameter appear, in some cases with both valves complete. Between 3.45 and 4.05 m b.s.l., the sediment contains many fibres (millimetre size) of Posidonia oceanica (L.) Delile. In the whole of Subunit A II, the sandy fraction is drastically reduced (4%) and composed of abundant shell fragments and scarce quartz and calcite sand (very fine). In samples 43, 45, 46, 49 and 50, massive calcium carbonate precipitation processes are observed, which in the majority of cases are rhizotubules. These sediments, together with micropalaeontological evidence, are interpreted as episodes of saturated environments showing a recurring pattern. In the two episodes at the base (samples 45, 46, 49 and 50), this massive carbonate precipitation takes place in a restricted environment, given the predominance of brackish water tests as A. tepida (78% of average relative abundance, in first and second episodes), H. germanica (13.9%), C. excavatum (3%) and P. eburnea (2.6% in samples 45 and 46). In some cases (sample 49), of the freshwater ostracod Ilyocypris gibba (Ramdohr) is recorded. The saturated episode of sample 43 records an abundance of shell fragments (Cerastoderma glaucum (Poiret) and other bivalves, as well as marine and brackish gastropods such as Hydrobia sp.) although the assemblage of foraminifera is similar to the previous saturated episodes, especially to samples 49 and 50. The rest of Subunit A II presents a mixed assemblage of marine species (Ammonia beccarii (Linné), with 5% of average relative abundance, Pseudotriloculina rotunda (D’Orbigny), with 2%, Miliolinella subrotunda (Montagú), with 1.3%) and brackish species mentioned, indicating a restricted environment connected with the open sea. Sample 44 presents oogonia of Charophytes (Lamprothamnium papulosum (Wallr)) and many valves of the ostracod C. torosa (ostracodites) indicating the stability of the brackish water bodies. From the taphonomic viewpoint, a worsening of conservation is also observed in the tests of marine origin.
The values of the Shannon diversity index are higher than those obtained for Subunit A I. The dominance of the species did not provide clear results and the equitability is higher (with respect to Subunit A I). The percentage of allochthonous taxa is nearly 8.3% although this should be taken with caution (as in lower levels) because, in an open environment, it is not possible to reliably differentiate which species are adapted to it and which ones are drawn from the open sea. There are no palaeoenvironmental differences between the top of A I Subunit and the entire A II Subunit. In both cases, the lagoon is connected to the sea, where restricted conditions prevail. Nevertheless, the drastic change of Subunit A II to muddy textures (high clay content) indicates that it is a very low energy lagoon with predominance of decantation processes (flocculation) in very calm waters. Datings carried out in Subunit A II give dates of 4605 ± 175 cal. yr BP (3.8–3.85 m b.s.l.), 4380 ± 130 cal. yr BP (3.65–3.67 m b.s.l.) and 3710 ± 130 cal. yr BP (top layer of the unit at 2.95–3.05 m b.s.l.).
Subunit A III
From 2.95 to 0.95 m b.s.l. Foraminifera and sediment samples: 39–18. In this subunit, the sediment is the same light grey colour as in the previous level (2.5Y 7/1) and likewise contains abundant shells of Cerastoderma sp. (1–3 cm diameter). The texture is slightly different, varying between mud and sandy silt, because of the slight and variable increase in sand throughout the record, especially between 2.25 and 1.75 m b.s.l. The sand fraction is composed almost entirely of Cerastoderma sp. fragments, with a slight proportion of very fine quartz sand. In this subunit, we find abundant gyronites of Characeae (L. papulosum) and no remains of P. oceanica plant fibres were identified. Biogenic residues are especially abundant among micropalaeontological samples 23–21, including foraminifera tests. From samples 21 to 39, we detected abundant carbonate precipitate, reaching maximum saturation (rhizotubules) in sample 39. At the top of the subunit (0.95 m b.s.l.), there is a layer with a high concentration of bivalve Abra ovata (Philippi) (several millimetre diameter) and Cerastoderma sp. shell fragments. After sample 39, the micropalaeontological content of Subunit A III indicates disconnection from the marine environment. The assemblage of foraminifera found is from restricted and brackish waters: A. tepida (74.9%), H. germanica (14.8%), C. excavatum (4.48%) accompanied by abundant shell fragments formed by an amalgam of brackish bivalves and gastropods, as well as ostracods (C. torosa). The percentage of allochthonous taxa is nearly 3%. From the taphonomic standpoint, the foraminifera tests are very well preserved. Regarding the diversity index, the average value of species was nearly 5, lower than the other (Subunit A II); consequently, the values of diversity decrease again (Table 2). This can be a response to the isolation of the brackish lagoon. The above data allow us to interpret this environment as a brackish lagoon of low energy, albeit slightly higher than that of the underlying stratum. Five 14C datings were carried out, with the following results: 2300 ± 120 cal. yr BP (1.99–2.00 m b.s.l.); 1360 ± 50 cal. yr BP (1.47–1.54 m b.s.l.); 1320 ± 30 cal. yr BP (1.45–1.47 m b.s.l.); 820 ± 90 cal. yr BP (two datings in shell and plant samples) (1.03–1.05 m b.s.l.).
Environmental facies of Unit B
Unit B is 1.8 m thick (0.95 m b.s.l.–0.85 m a.s.l.). Foraminifera samples: 17 and 16; sediment samples: 17–1. This level corresponds to the top layer of the survey, and is in sharp contact with the previous level (Subunit A III). The colour of Unit B changes to olive brown tones. From the textural point of view, it is characterised by a clear increase in the percentage of silt compared with all the underlying levels (33%) and a very low sand content (around 3.4%). According to the textural classification, most of the samples are classified as silt and some as mud. As for biofacies, the brackish bivalve fauna disappears (Cerastoderma sp.) and centimetre-sized freshwater gastropods appear.
From the micropalaeontological point of view, there are no differences between samples 3 and 12 and samples 1 and 2 are interpreted as flood plain environment. There are hardly any foraminifera tests, and the biogenic residues are the same throughout the unit (fragments of ostracods and charophytes, opercula and gastropods such as Hydrobia sp.). Massive carbonate precipitation was observed in samples 9 and 10, precipitation of iron sulphide between samples 13 and 9 and the largest amount of charcoal was described in samples 4 and 5. The entire unit is interpreted as a freshwater environment, anoxic and in swampy facies.
From the textural and sedimentological viewpoint, two sub-environments, B I y B II, are undistinguished. In sub-environment B II, a gradual change can be observed between the B II lower, B II mid and B II upper levels.
Subunit B I
From 0.95 to 0.55 m b.s.l. The colour is light olive brown, with bands of yellow colourations. The sand fraction keeps the textural characteristics of the immediately underlying layer and is almost entirely composed of very fine shell fragments of Cerastoderma sp. with a slight proportion of very fine quartz sand. It contains gyronites of Characeae and white coloured vegetable fibres. From the micropalaeontological point of view, in this unit we recorded tests of the species A. tepida as well as brackish (C. torosa) and freshwater (I. gibba) ostracods. The abundance of carbonate determines a saturated environment. It is interpreted as a swampy environment. The fauna in general decreases dramatically in abundance compared with the underlying level (Table 2). From the taphonomic viewpoint, shells show a worse state of conservation compared with previous levels. This assemblage is interpreted as reworked (from previous deposits).
Subunit B II
Unit B II lower level
From 0.55 to 0.15 m b.s.l. This level is grey in colour. The sand is very fine, subangular to subrounded and basically calcareous. Between 0.55 and 0.53 and between 0.49 and 0.46 m b.s.l. we find macroremains of brown coloured aquatic plants and some peat. Towards 0.45 m b.s.l., pyrite concretions are appreciated. Finally, towards the top of the subunit (between 0.35 and 0.15 m b.s.l.), freshwater gastropods 0.5 cm in diameter appear (Bytynia tentaculata (Linne)), with abundant brown coloured vegetable remains and Chara sp. gyronites.
From the micropalaeontological point of view, no foraminifer tests were reported, with the exception of sample 13, where some tests of the suborder Trochamminina were detected. Freshwater ostracods and charophytes were also observed. In sample 12, the amount of carbon and peaty conditions increased. There are frequent carbonate precipitations, especially in the top layer. At 0.54–0.53 m b.s.l., we carried out a dating with the result of 510 ± 20 cal. yr BP.
Subunit B II mid level
From 0.15 m b.s.l. to 0.33 m a.s.l. This layer contains abundant charcoal, very fine calcareous sands, opercula, gyronites of Characeae and abundant shell fragments of millimetre-sized freshwater lagoon gastropods. Between 0.05 m b.s.l. to 0.15 m a.s.l., there are levels of organic matter. From the micropalaeontological point of view, no foraminifer tests were recorded.
Subunit B II upper level
From 0.33 to 0.85 m a.s.l. At this level, the texture is fine silt to mud and pale brown in colour (10 YR 6/3). It contains gastropods Planorbis sp. (millimetre size), opercula, abundant plant fibre debris and charcoal. Towards the base of this level, redox processes and accumulations of organic matter are observed. At 0.31 m a.s.l., there is a peat layer.
It may be concluded that the sharp decline in foraminiferal abundance and the presence of freshwater gastropods determined the change to a swampy or freshwater lagoon environment. This change takes place gradually in Subunit B I (between 0.95 and 0.55 m b.s.l.) where we can still appreciate the presence of quartz sand and millimetric-sized shell fragments of Cerastoderma sp. In the upper layers, the increase in silt proportion, the presence of macroremains of aquatic vegetation (undecomposed), the appearance of freshwater gastropods, the greater organic matter content and the appearance of charcoal and pyrite concretions indicate a swampy continental environment whose sedimentary infill would take place in a flood basin environment.
Discussion
El core analysed (SAL/S2) covers the evolutionary sequence of the palaeolagoon that was formed during the Holocene in this sector of the Valencian coastal alluvial plain until its conversion into the current freshwater marsh and flood plain of the Turia river. Our objectives were on one hand to analyse the geomorphological processes and evolution and, on the other, to characterise the possible responses of the geomorphological system to middle–late Holocene climate events. To understand the geomorphological dynamics and possible responses to global middle–late Holocene climate variability, it is necessary to relate lagoon, flood plain and coastal barrier processes. We distinguish four phases corresponding to the units and subunits described in the sedimentary record (Figures 4 and 5).
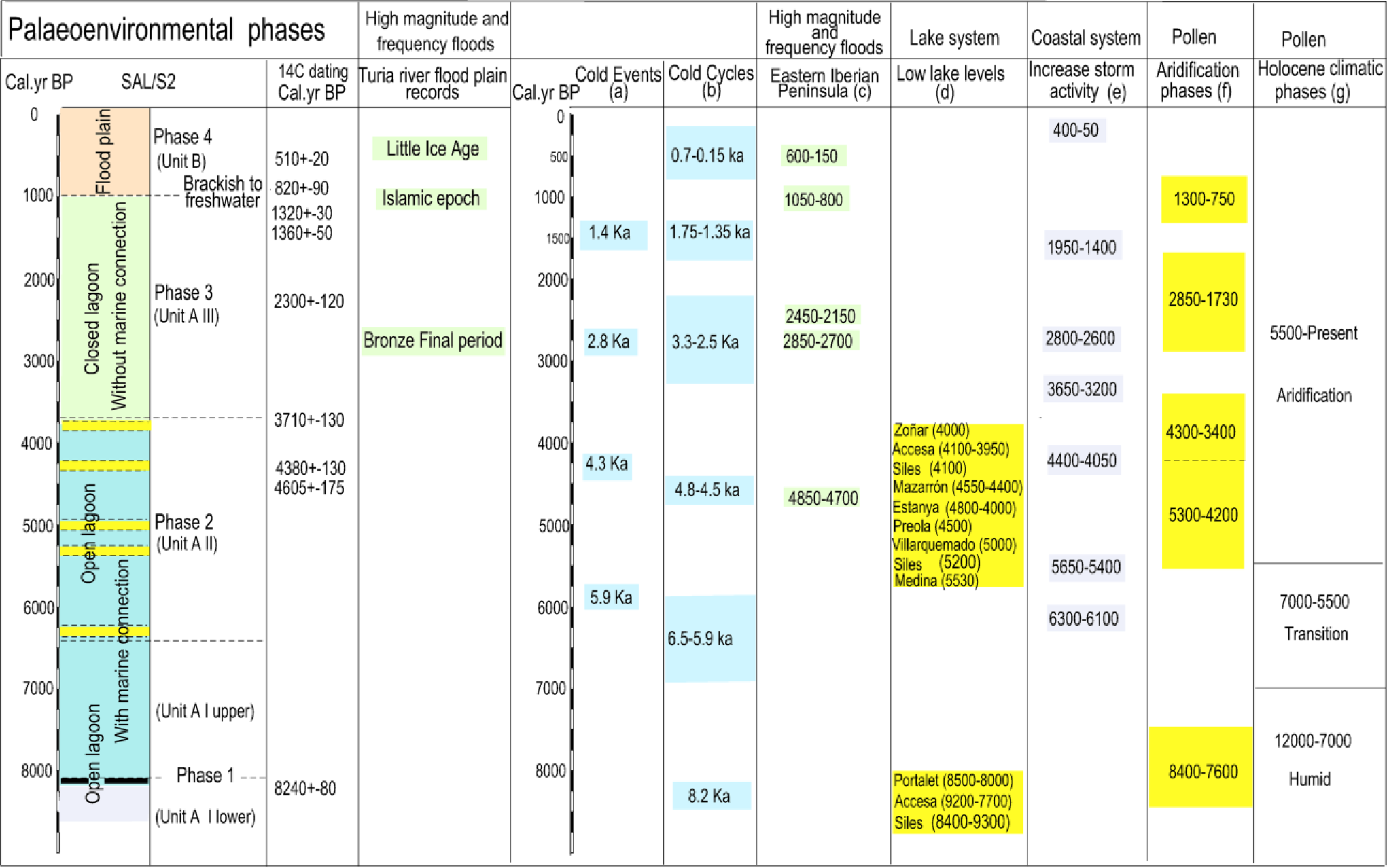
Correlation between palaeoenvironmental phases of the lagoon and other selected records: (a) Bond et al., 1997, 2001; (b) Wanner et al., 2011; (c) Benito et al., 2015; (d) Zoñar: Martín-Puertas et al., 2008; Accesa: Magny et al., 2007, 2009; Siles: Carrión, 2002; Mazarrón: Navarro-Hervás et al., 2014; Stanya: Morellón et al., 2009; Preola: Magny et al., 2011; Villarquemado: Aranbarri et al., 2014; Medina: Reed et al., 2001; Portalet: González-Sampériz et al., 2006; (e) Sabatier et al., 2012; (f) Jalut et al., 2000; (g) Jalut et al., 2000; Pérez-Obiol et al., 2011; Carrión et al., 2010.
Phase 1: From the mid-Holocene marine transgression (8240 ± 80 cal. yr BP) (Subunit A I lower level) to the marine maximum: A coastal lagoon connected with the sea (Subunit A I upper level)
As shown in the record for the lower level of Subunit A I, during the marine transgression of the middle Holocene, this area was flooded by brackish waters which deposited a thin sandy mud and muddy sand sequence. The peat layer (4.60–4.55 m b.s.l.) dated around 8240 ± 80 cal. yr BP assigns an earlier date to this level of the core base. The dating correlates well with the chronology of the early peats analysed in the palaeovalley (Grau) of the River Turia (Figures 2 and 3), at very similar absolute height (−5 to −6 m b.s.l.) and dated at around 8530–8280 cal. yr BP (Carmona and Ruiz, 2011). From the palaeoclimate perspective, the date obtained in this peat layer matches the 8.2 ka cold event (Alley and Agustsdottir, 2005) and with one of the cold cycles of Wanner et al. (2011). It also coincides with a period of aridity identified in pollen records and with shallower conditions in lake water levels (Carrión, 2002 (Siles lake); González-Sampériz et al., 2006 (Portalet lake); Jalut et al., 2000; Magny et al., 2007 (Accesa lake)) (Figures 1 and 5).
The upper part of Subunit A I (4.55–4.17 m b.s.l.) corresponds to a lagoon environment of moderate–high energy (the highest in the entire core) with marine communication. Although we have no specific dating for this level, it could correspond to the maximum marine transgression in the area. Thus, this lagoon could be contemporaneous with the beach barrier identified at the base of the core of Pinedo on the nearby coast (Figures 2 and 3), dated 7500–7300 cal. yr BP (around −1 to 0 m b.s.l.) (Carmona and Ruiz, 2011). We think that the bioclastic sands of this upper section of Subunit A I would have come from the coastal zone and may have been introduced by coastal currents. On the other hand, it is possible that the beach dated in the Pinedo core may have been a spit/sandbar of a barrier island system formed during the transgressive phase. The chronology is consistent with the high Holocene sea level in the Bairén record (50 km to the south, Figure 1) corresponding to a sandy level in an infralittoral environment at similar absolute height and dated around 6980–6780 cal. yr BP (Carmona and Pérez-Ballester, 2011; Ruiz and Carmona, 2005) (Figure 3). The chronology of both sea levels is similar at the overall radiocarbon dates of the maximum Holocene marine transgression in the Mediterranean (Pirazzoli, 2005; Sabatier et al., 2010; Stewart and Morhange, 2009). Moreover, the data are consistent with stratigraphic models of Holocene barriers and lagoons (Dalrymple et al., 1994; Roy et al., 1994; Woodroffe and Saito, 2011) and also with the typology and chronology of the early barriers studied in the Atlantic Mediterranean linkage area that began to develop around 6500–7000 cal. yr BP (Dabrio et al., 2000; Goy et al., 1986; Zazo et al., 1994, 2008).
Phase 2: From 6450 cal. yr BP (approximately) to 3710 ± 130 cal. yr BP: A very low energy brackish lagoon with sea connection (Subunit A II)
Subunit A II (4.17–2.95 m b.s.l.) corresponds to a restricted lagoon environment clearly communicated with the sea. This lagoon is very low energy and presents active clay flocculation processes in very calm waters. The episodes of massive carbonate precipitation and rhizotubules indicate saturation processes that could be interpreted as a fall in water level. The age of the base of this level (around 4.17 m b.s.l.) interpolated between radiocarbon-dated horizons would indicate a starting date for the formation of this environment around 6450 cal. yr BP. The absolute 14C dating samples obtained at the top of this subunit cover a period of approximately 1000 years (4605 ± 175; 4380 ± 130; 3710 ± 130 cal. yr BP). Thus, Subunit A II would be contemporary with the period of marked increase in the aridity trend evidenced in the pollen sequences in this sector of the Mediterranean (Carrión et al., 2010; Fletcher et al., 2007; Jalut et al., 2000, 2009; Jiménez-Moreno et al., 2015; Pantaleón-Cano et al., 2003; Pérez-Obiol et al., 2011) and also with aridity on a global scale (Booth et al., 2005; Weiss et al., 1993). Furthermore, regional comparisons suggest a correlation of North Atlantic Holocene cooling events (peak 4.3 of Bond et al. (1997, 2001) and peaks 3 and 4.8–4.5 cal. yr BP cycles of Wanner et al. (2011)) and with dry conditions across southern Iberia and Northwest Africa (Fletcher et al., 2007) (Figure 5).
Moreover, it is also important to note that during this period abrupt drops in water levels are detected in several continental lakes of the Iberian peninsula (Aranbarri et al., 2014; Carrión, 2002; Martín-Puertas et al., 2008; Morellón et al., 2009; Reed et al., 2001) and in Mediterranean coastal lagoons such as Mazarrón harbour (Navarro-Hervás et al., 2014) (Figure 5). In Italy, several lakes evidence this same process. In the record of lake Preola, in southern Sicily, the period from 4500 cal. yr BP to present coincides with a lowering of the lake levels (Magny et al., 2011). Comparisons of the Accesa lake levels record (north-central Italy) with marine records from Adriatic Sea and terrestrial palaeoclimate records from Tunisia and Spain suggest that the complex climatic oscillations of this period (two phases characterised by wetter conditions dated to 4300–4100 and 3950–3850 cal. yr BP and a phase marked by drier conditions at 4100–3950 cal. yr BP) may have affected the whole central and western Mediterranean area and can pinpoint the moment of a crucial transition from mid to late-Holocene for Mediterranean and tropical areas (Magny et al., 2009).
Phase 3: From 3710 ± 130 cal. yr BP to 820 ± 90 cal. yr BP: A brackish lagoon without marine connection (Subunit A III)
The micropalaeontological information indicates the lagoon’s closure from the sea towards 3710 ± 130 cal. yr BP (dating at 3.05–2.95 m b.s.l., at the base of Subunit A III) and its complete isolation (change from brackish lagoon to freshwater lagoon) around 820 ± 90 cal. yr BP (dating at top layer of Subunit A III). Thus, throughout the chronological period covered by Subunit III, a shift gradually took place from a protected lagoon to a totally isolated lagoon environment. It should be noted that the timing of the start of lagoon marine disconnect is prior to the closure of the Atlantic estuaries of Spain (Cádiz-Huelva) dated at around 3100 cal. yr BP (Zazo, 2006).
The whole process is related to the dynamics of closure of the coastal barrier. So, although the only morphological expression on the nearby coast is the current (or sub-recent) thin barrier, the Pinedo record (Figure 3) located in a more continental sector, (away from the current coastline) (Figure 2), shows a barrier sequence whose morphology and dynamics would provide keys to help interpret the lagoon record. As mentioned in previous paragraphs, this sequence is composed of a beach barrier dated around 7500–7300 cal. yr BP. The beach level is buried by grey silt from a backbarrier coastal lagoon (dated around 3360–3150 cal. yr BP) (Carmona and Ruiz, 2011).
Regarding the chronology of shift from brackish to freshwater lagoon, the top layer dates of 820 ± 90 cal. yr BP (top of Subunit A III) coincide with those obtained at the same stratigraphic level (at similar depths) in nearby records (La Pipa 2 and 3 in Figures 2 and 3). The datings taken at these cores give dates of 1050–920 cal. yr BP (1.25 m b.s.l.) and 960–770 cal. yr BP (1.75 m b.s.l.) for the change (Carmona and Ruiz, 2014). Moreover, surveys carried out in the current lagoon show that the change from brackish to freshwater took place around 1110 ± 115 cal. yr BP (Sanjaume et al., 1992). On the other hand, we note that this closure process is synchronous with that detected in the Palavasian wetland (French Mediterranean coast) (Sabatier et al., 2010, 2012), where the faunal content (a clear shift in mollusc population characterised by an increase of the most typical lagoonal specie Hydrobia acuta whereas the number of marine species Bittium recticulatum decreases) revealed a major palaeoenvironmental change around 1000 cal. yr BP, which could have resulted from the total closure of communications between the lagoon and the sea around 730 ± 120 cal. yr BP (Sabatier et al., 2010).
On our coast, the definitive closure process of the lagoon may be placed in relation with the extraordinary sedimentation rate during the high-magnitude flood events of the 11th century, identified in the palaeohydrological record of the Turia river (Figure 3). The coastal progradation processes related with this extraordinary input of sediments would rethicken the ridge of the medieval mouth of the River Turia (La Punta) (Carmona and Ruiz, 2011). This phase also coincides with one of the concentration periods of high-magnitude flooding revealed in records of slack water sediments around 1000 cal. yr BP in rivers of the Iberian Peninsula (Benito et al., 2008, 2015; Machado et al., 2012; Thorndycraft and Benito, 2006a, 2006b) and Southern France (Benito et al., 2015), related to climatic–anthropogenic causes (Figure 5).
Finally, we should note that although several closure phases have been identified in this lagoon record, it is possible that coastal local neotectonics, already mentioned in the geomorphological description of the barrier (Carmona and Ruiz, 1999), may hinder the possible identification and correlation with the barrier progradation phases described in the southern sector (el Palmar record) (Marco-Barba et al., 2013b; Santisteban et al., 2009) and other Mediterranean areas (Goy et al., 2003; Zazo et al., 2008).
Phase 4: During the LIA the isolated freshwater lagoon environment became a swampy environment and finally a flood plain (Unit B)
From the palaeoenvironmental viewpoint, in Unit B we detect a fundamental change, where the isolated freshwater lagoon environment becomes a swampy and finally a flood plain environment. Around 510 ± 20 cal. yr BP (lower Subunit B II), the lagoon was colonised by abundant aquatic vegetation and freshwater gastropods. Towards the top of the sequence, the rate of sedimentation increases (0.241 cm/yr, the highest in the record) (Figure 4), a fact that may be related with the global change of the LIA period. Indeed, from AD 1545 to 1900 (405–50 cal. yr BP) there was a much greater frequency of flood events than at present in the coastal flood plains of the rivers Turia and Júcar (Ruiz et al., 2014). Frequent flooding caused an environmental change in the lagoon. On one hand, these changes referred to a notable increase in water tables, which must have affected the freshening process of the lagoon and, on the other, the extraordinary sediment supply by floods. The outcome is the formation of alluvial levees in the wetland environment (clearly shown in the LiDAR DEM). In poorly drained areas of low topography, freshwater marshes and shallow lagoons appear. All these environments are recorded in the sedimentary sequence of Subunit B II. The transition towards continentalisation gradually transformed the wetland into flood plain. On the other hand, the extraordinary sediment supply to the coast from the mouth of the River Turia could have conformed the current barrier set.
The phase of high frequency and magnitude of floods corresponding to the LIA period is of global scope (Arnaud-Fasseta et al., 2010; Bruneton et al., 2001; Glaser et al., 2010; Miramont et al., 1998; Pichard, 1995; Schmocker-Fackel and Naef, 2010) and coincidental with glacial advances in the Alps (Grove, 2001). It is important to highlight the effects of the extraordinary sediment inputs of rivers to the deltaic coasts during the LIA period. Amorosi and Milli (2001) observed that the progradation phase of the Tevere delta of roughly 4.5 km (10 m/yr) took place during LIA period. The same phase, with similar features but different progradation rates, has been recorded in other eastern Tyrrhenian deltas (Alessandro et al., 1990; Caputo et al., 1987). On the other hand, in addition to severe flooding, several authors report the incidence of intense coastal storms in centuries of the LIA period in Mediterranean barrier–lagoon systems of the French coast (Blanchemanche, 2009; Dezileau et al., 2011; Sabatier et al., 2008) (Figure 5).
Conclusion
We conclude that the coastal lagoon and flood plain constitute a system in which the processes are interlinked. Thus, the geomorphological evolution and the response to Holocene climatic and hydrologic variability reveal the multiple connections. Analysis of the sedimentary record and foraminifera of this coastal lagoon showed changes in the salinity and variations in texture and sedimentation rates, as well as groundwater fluctuations that reflect responses to global Holocene climate changes. The progressive closures of the lagoon could be related to periods of extraordinary river inputs of sediments in the coastal flood plains.
Dynamic and palaeoenvironmental data enable us to distinguish four different phases in the geomorphological Holocene evolution of the lagoon and coastal alluvial plain:
Phase 1. Around 8240 ± 80 cal. yr BP, a brackish lagoon of moderate energy and in restricted environment was formed, with an energy peak that could correspond to the maximum Holocene marine transgression. The peat layer dating (8240 ± 80 cal. yr BP) corresponds to the cold 8.2 ka event and low water levels in inland lakes of the Iberian Peninsula and Italy.
Phase 2. From 6450 cal. yr BP (approximately) to 3710 ± 130 cal. yr BP, a lagoon remained, in restricted environment connected with the sea, but with a notable energy decrease. The lagoon presents recurrent environmental saturation processes similar to those described in continental lakes of the Iberian Peninsula and other Mediterranean coastal lagoons, which could be interpreted as a fall in water level. This phase is contemporaneous with a period of increase in the aridity trend in the western Mediterranean and on a global scale. It is also roughly contemporary with global cold cycles.
Phase 3. From 3710 ± 130 cal. yr BP, a brackish lagoon closes its connection with the open sea and towards 820 ± 90 cal. yr BP a shift to a totally isolated lagoon environment took place (changing from brackish to freshwater lagoon). The process is synchronous to another of similar characteristics detected in a coastal lagoon of the French Mediterranean coast. On the other hand, the process of change from brackish to freshwater lagoon is coeval with a palaeohydrological phase of high flood frequency and magnitude in the flood plain of the River Turia and other rivers of the Iberian Peninsula and Southern France. These sedimentary inputs to the coastal barrier could have caused the lagoon’s complete isolation from seawater.
Phase 4. Freshwater lagoon environment becomes flood plain. The phase of high frequency and magnitude of floods corresponding to the LIA period, of global scope and coincidental with glacial advances in the Alps, would rethicken the most recent prograding set of the current barrier and form the flood plain on the top level of the sedimentary sequence.
Phases and processes in the coastal flood plain and barrier–lagoon system recorded in sedimentation could be placed in relation with global mid–late Holocene events. Many of these hydrogeomorphological processes are contemporary with specific cold events, such as phases 1, 2 and 4. We observed that the response in our hydrological system is variable, with either aridity–shallower conditions (phases 1 and 2) or high flooding frequency and magnitude (phase 4 LIA period).
Footnotes
Acknowledgements
The authors thank two anonymous reviewers for insightful critiques that improved the manuscript.
Funding
This work was supported by grant CSO 2012-32367, Department of Scientific and Technical Research. Ministry of Economy and Competitiveness (Spain). ‘Hydrogeomorphological processes and changes in Mediterranean coastal flood plains. Climate variability and human actions. A multiscale approach’. This work was partially carried out thanks to the support of Labex OT-Med (ANR-11-LABX-0061) and the A*MIDEX project (no. ANR-11-IDEX-0001-02), funded by the French Government’s ‘Investissements d’Avenir’ programme, managed by the French National Research Agency (ANR).