
Abstract
We test several methods of ostracod-based palaeoenvironmental reconstruction using indicator species approach, mutual ecological/climatic range methods, transfer functions, modern analogue technique and morphological variation within Cyprideis torosa in reconstructing the site evolution of a late Quaternary small lake basin in Thuringia, Central Germany. Sediment sections containing a diverse ostracod fauna were studied and compared with those from modern water bodies of Thuringia. Palynological investigations were executed to reconstruct the environmental conditions in the catchment area and for a obtaining a biostratigraphical framework. The brackish water ostracod Cyprideis torosa as well as the foraminifer Haplophragmoides indicate phases of saline groundwater influence, fed by salt bearing sediments of the Triassic basement. The accompanying freshwater ostracod fauna, however, reflects only low variations of salinity and temperature. Environmental changes in salinity, temperature and ecological stability indicated by microfossils and pollen are caused by an interplay of climatic shifts and the local geological and hydrological setting.
Keywords
Introduction
Athalassic saline waters are typical for arid regions of the world. Humid regions, however, can yield such continental brackish water bodies as well (Pint et al., 2012). Their existence is normally related to evaporite sediments producing brines in contact with groundwater and feeding the lake water through springs. Athalassic brackish waters of humid regions are normally geologically short living and small compared with many of those in arid regions. According to the literature, a number of ancient water bodies in Central Germany were indicated to be probably saline based on the occurrence of foraminifers far from the sea (Bartenstein, 1939; Wennrich et al., 2007). Two sources of saline influence of these modern waters are considered: evaporite bearing sediments of Zechstein or Middle Triassic rocks and mine dumps. We think, however, that the presence of continental salt water sites in Central Germany is not solely driven by distribution of evaporites and tectonic or erosional processes giving groundwater access to those evaporites in the underground, but is also influenced by a climatic component either by shifting the groundwater levels or by a relatively low precipitation/evaporation ratio which can additionally enhance the salinization of the surface water but, on the other hand, may restrict the leaching in the underground by less groundwater (Pint et al., 2012). A detailed study of Holocene sediment profiles from a former shallow saline lake at Siebleben near Gotha was therefore started for testing these assumptions.
We document the fossil ostracod associations of two cores from Siebleber Senke in order to reconstruct palaeoenvironmental changes. The following methods are applied: indicator species approach, mutual ecological tolerance methods (incl. Mutual Ostracod Temperature Range (MOTR) sensu Horne, 2007), transfer function for salinity, modern analogue technique and morphological variation within Cyprideis torosa (noding and sieve-pore shape). Palynological analysis provides the independent reconstruction of climate and vegetation changes as well as chronology. The synthesis of palaeoecological results from ostracod and palynological analyses produces a model for site evolution. Relying on this model, we intend to test our hypothesis of climatically driven salinization in a temperate humid region.
Setting
The Siebleber Senke (Depression of Siebleben) close to Gotha belongs to a number of natural saline inland water (athalassic) sites in the Thuringian Basin which are in contact with evaporite bearing sediments of Triassic or Permian age. The ascendance of brines is facilitated by NW-SE trending faults running through the study area (Seidel, 2003). In the Siebleber Senke, a fishpond (diameter about 400 m, maximum depth: 1.5 m, height a.s.l.: 275 m) was created in 1717 (Greifeld, 1966) in place of a small remnant lake of natural origin in this depression. The present day salinity of the small pond is 0.5 psu, and the water is
Bartenstein (1939) reported the brackish water foraminifer Haplophragmoides for the first time from Siebleber Senke. However, he could not find living individuals. Empty tests of Haplophagmoides (Figure 1) sampled in 2005 were given to the authors by H Huckriede (TLUG Weimar). Their excellent preservation indicates a modern or sub-recent population in the lake. The index brackish water ostracod Cyprideis torosa could not be found in surface samples. Sediment cores, however, contain a typical freshwater macro- and microfauna including horizons with high numbers of Cyprideis torosa, accompanied by mainly halotolerant taxa like Limnocythere inopinata and Heterocypris salina. Their lateral spacing indicates a former brackish lake in the area as already assumed by Greifeld (1966) in her unpublished Diploma thesis.

Haplophragmoides sp. from surface sediments of the modern fishpond in the Siebleber Senke. The material was provided by H Huckriede (TLUG Weimar).
To understand the development of the Siebleber Senke, Greifeld (1966) described trenches, soundings and a core (Sieb, 1966) re-analysed for Ostracoda in the present study. According to her results, the sediment thickness in the depression reaches between 1 and 7 m, whereas the deepest part is situated in the western area (Figure 2). Core Sieb 1966 starts with a basal unit of clay, which contains small pieces of the Triassic bedrock (5.25–5.00 m). Up to 3.80 m, it is followed by grey-green clayey silt with solitaire gypsum pieces. The next bright layer up to 2.95 m has a high level of carbonate with scattered gypsum; therefore, it is interpreted as lake marl. Grey-green clayey silt with gypsum characterizes the next layer up to 1.70 m. Up to 0.8 m, a lake marl horizon occurs again. The last layer up to 0.2 m contains brownish silt with plant remains.

Topographic map showing contour lines of the Siebleben depression, core positions, positions of radiocarbon datings and correlation of the cores based on lithological and faunistic index horizons (map after Greifeld, 1966; redrawn); A – core Sieb 01/06, B – Sieb 1966. Contour lines outside the depression are omitted.
Material and methods
Sampling and analysis of collection material
Three sediment cores from nearly the same position in the area of the Siebleber Senke are investigated in this study (Figure 2). The first of them with a length of 525 cm (core Sieb 1966, 50°56′08″N, 10°45′40″E) was drilled in 1965 in the framework of a Diploma thesis by Ingrid Greifeld (1966). In the lower part of this core, pollen grains were analysed for chronological interpretation of the late Glacial (Lange, 1966). The complete ostracod fauna from this core (46 samples) is archived in the Natural History Museum of Berlin within the Diebel-Pietrzeniuk ostracod collection. This material was re-analysed for this study. In 2006 and 2010, two parallel cores, c. 500 m southern of core Sieb 1966, in one position were drilled for investigations of both pollen and ostracods with a manual gauge (core Sieb P/06 – 385 cm; core Sieb P/10 – 235 cm). Whereas core Sieb P/06 covers the Preboreal and Boreal, core Sieb P/10 includes the sequence from Boreal to modern. The correlation of the cores was possible using marker horizons, based on the mass occurrence of Cyprideis torosa and lake marl layers, and show only variations in small centimetre scale (Figure 2). From core Sieb P/06, 61 samples were taken for ostracod analysis covering the whole core length and 34 samples for palynology covering the Holocene. The ostracod samples have 25 g wet weight each, the pollen samples a volume of 1 ml each.
Sample preparation and documentation
The ostracod samples were completely sieved over a 200-µm sieve using tap water. The sieve residue was then oven-dried at 50°C. All ostracod valves were picked from the residues, identified, counted and stored in microslides. Taxonomic identification relies on Meisch (2000) and Diebel (1965, 1968) and was done under a low-magnification stereoscopic microscope. The excellent plates by Fuhrmann (2013) supported identification additionally when using SEM and light microscope photographs. Ilyocypris species were identified using valve size, morphology (marginal spines and nodes) and the presence and degree of marginal ripplets in the posterior-ventral part of the adult left valves sensu Van Harten (1979) and Janz (1994). The Ilyocypris sp. could not be assigned to a species given in Meisch (2000); the marginal zone of the left valve of this species is smooth without ripplets.
The studied material, except this from the Diebel-Pietrzeniuk collection, will be transferred to the museum Mauritianum in Altenburg (Thuringia), where the large Fuhrmann collection of Quaternary ostracods from Central Germany is also housed.
Preparation of pollen samples follows Faegri and Iversen (1989); additionally, the samples were sieved through an 8-µm Nylon mesh with ultra sound to remove clastic material. For analyses, the samples were embedded in silicon oil. Pollen identification was carried out with the help of Moore et al. (1991), Reille (1992) and Beug (2004) and the pollen collection of the Friedrich Schiller University Jena. Pollen counts are presented as a diagram with regional and local part and with ecological summary curves. In the same process, charcoal pieces were counted to identify periods with higher fire activity, which could point to dryer climate conditions or an increasing human impact (Schneider et al., 2016).
Statistical analyses
The modern training set of ostracods from Central Germany by Pint et al. (2015) complemented by another modern training set from the Baltic Sea coast (Frenzel et al., 2010) limited to oligohaline samples allowed the setup of an ostracod-based transfer function for salinity. The modern training set comprises 73 samples, 26 from Central Germany and 47 from the southern Baltic Sea coast, and covers salinities from 0.1 to 6.1 (psu). The performance calculated by bootstrapping is R2 = 0.74 and an RMSEP (root mean square error of prediction) of 0.9 for the WAPLS model (Weighted Averaging Partial Least Squares) of the transfer function.
Zonation, correlation and chronology of cores
An ostracod-based zonation for the studied core was set up relying on ostracod distribution data, whereas the palynological zonation refers to changes in pollen composition (Moore et al., 1991). The sediment cores of 1966, 2006 and 2010 could be correlated based on lithological features like grain size, colour, carbonate content as well as pollen and ostracod distribution (Figure 2). Problematic is the differing description of core 1966 because of a deviant definition of marl and carbonate content. For the ostracod zonation, applied for the cores 1966 and 2006, the clear appearance and disappearance of taxa were used for the definition of the zonal boundaries. Furthermore, pollen zones of this study could be correlated based on a late glacial pollen diagram composed by Lange (1966) as stratigraphical framework for Greifeld (1966). An approximately chronological interpretation was possible using other dated pollen diagrams from this region as base for comparison (Meschner, 2008; Schneider, 2014). Palynological results of the oldest part are given in a simplified pollen diagram by Lange (1966). The original dataset of this diagram is not available. The basis of calculation in 1966 and today is different and thus the diagram of Lange (1966) is reconstructed here. Starting with the Holocene, a pollen diagram of the depression is presented in Figure 6b.
For dating the profiles, two samples were used for AMS-14C-Measurement (Table 1). Measured radiocarbon ages were calibrated using Calib 7.1 and the IntCal13 calibration curve (Reimer et al., 2013), and ages are given in calibrated years Before Present (cal BP) with a 2 sigma error age range.
Details of radiocarbon datings in core Sieb P/06.

Palaeoenvironmental analysis
We apply a range of methods for palaeoenvironmental reconstruction to the Siebleben profile. These are index species approach, ecological classification, MOTR method, Mutual Ostracod Salinity Range (MOSR) method, modern analogue technique and ostracod-based transfer functions for temperature and salinity (Frenzel et al., 2010). Additional information on salinity is provided by morphological analysis of Cyprideis torosa valves.
The index species method relies on the occurrence of a characteristic species like Cyprideis torosa as true brackish water indicators in our material to figure out saline influences in the pond. It gives no detailed results, but it is easy and fast to use. Ecological classification of ostracod groups and the mapping of their distribution is a classical and reliable method but shows mainly trends and no quantitative reconstructions for environmental parameters. We rely on ecological ostracod classifications by Meisch (2000) and Fuhrmann (2006, 2013). The MOTR method combines species distributions from a modern non-marine ostracod database and GIS-derived air temperature ranges (Horne, 2007; Horne and Mezquita, 2008). The MOTR reconstructions are presented as ranges (maximum and minimum values) for mean January, mean July and mean annual air temperature. The MOTR reconstructions are presented as ranges (maximum and minimum values) for mean January, mean July and mean annual air temperature. The used databases are NODE and OMEGA (Ostracod Metadatabase of Environmental and Geographical Attributes; Horne et al., 2012). For the Recent sites of the Thuringian Basin, MOTR is applied as a test for its performance (Pint et al., 2015) and thereafter for each ostracod zone of the core Sieb 1966 reconstructing palaeo-air-temperatures. The MOTR results are presented with 3 pt mean smoothed values (Table 2).
MOTR ranges in 3 pt mean in °C (Celsius) of the five zones.
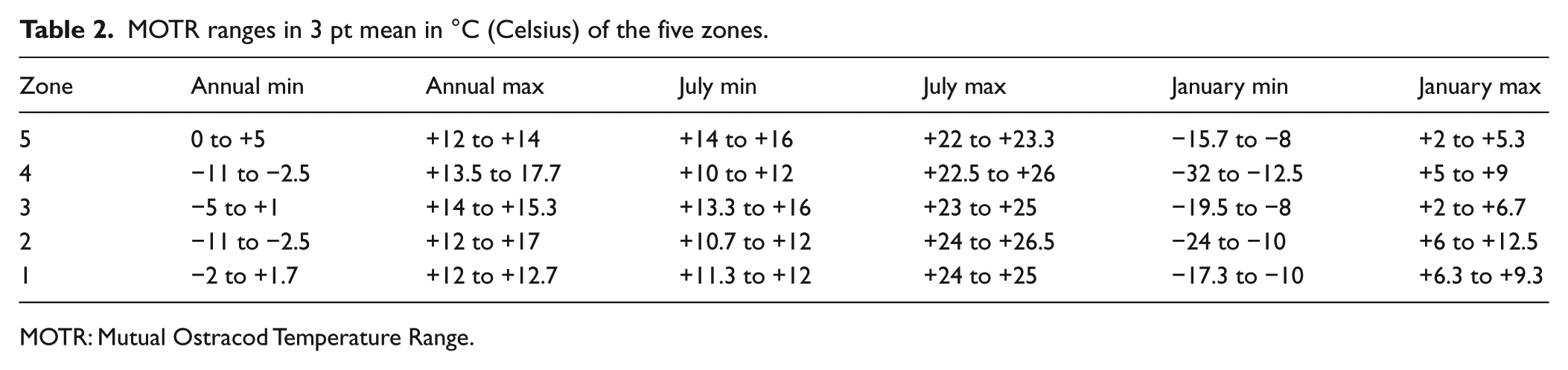
MOTR: Mutual Ostracod Temperature Range.
Salinity estimation is based on mutual salinity tolerances of the encountered ostracod taxa in a similar way as for MOTR; the general principle of this approach is not new, but we introduce it here for the first time as a systematic MOSR method based on specified datasets. Salinity ranges of individual taxa are taken from Frenzel et al. (2010) and Meisch (2000). The data used by Frenzel et al. (2010) are collected on the southern Baltic Sea coast and cover salinities from 0.05 to c. 20 relying on 745 samples.
Additional information on palaeosalinity is provided by sieve-pore analysis sensu Rosenfeld and Vesper (1977). These authors recognized that the shapes of sieve-pores of Cyprideis torosa and salinity are correlated. The increase in round-shaped sieve-pores indicates a decrease in salinity. With the aid of a transfer function (Pint et al., 2012), it is possible to calculate salinity directly from the percentage of round sieve-pores:

where S is the salinity and RS is the percentage of round sieve-pores.
It has to be stressed, however, that this formula is based on marginal marine populations and that it is not known so far if a water chemistry different from that of marginal marine settings influences the proportion of round sieve-pores. This problem is similar for noding of Cyprideis torosa. In marginal marine environments, Cyprideis torosa develops nodes below a salinity of about 7 psu. Below 2 psu, mostly more than 50% of valves of a population are noded (Frenzel, 1991). This phenomenon is caused by osmotic problems during moulting, resulting in bulges of the soft body and the carapax (Frenzel et al., 2012).
In general, the palaeoecological interpretation of fossil pollen and spores was based on uniformitarian principles (Jacomet and Kreuz, 1999) to divide pollen types into different local and regional groups. Lange (1966) reassigned the ecological separation between local and regional components for the late glacial and early-Holocene in core Sieb 1966 and aquatics are missed in her study. In order to correlate the stratigraphical classification of core Sieb 1966 and core Sieb P/06 during early-Holocene, the interpretation of Sieb P/06 follows the published interpretation model of Lange (1966).
During the later Holocene, the authors use the modern plant ecology of Ellenberg et al. (1992) for ecological classification of the pollen types that allows the reconstruction of vegetation pattern in the catchment as base for the stratigraphical interpretation.
The values of Chenopodiaceae are, in middle Europe, normally interpreted as ruderal indicators. In Thuringian areas characterized by gypsum and salt they document relatively high values and could coincide with the changes in salinity in the depression, which is observable in modern studies from Thuringia (Schuster et al., 2010). Consequently, they have been treated as local indicator. The high values in Poaceae are also typical for this kind of archive in Thuringia, such as are seen in different depressions in the Thuringian Basin (Meschner, 2008) and the Werra valley (Schuster et al., 2010). The often used local name for such depressions in the Thuringian Basin is ‘Ried’ or ‘Rieth’, which hints at the characteristic vegetation form with different species of Cyperaceae and Poaceae.
Both pollen types, Chenopodiaceae as well as Poaceae, are therefore interpreted as a local signal in the mid and late-Holocene. During the early-Holocene, they seem to be part of the regional vegetation.
Results
Lithology
The lithology of Greifeld’s (1966) core Sieb 1966 is given in chapter 2 (Settings). As expected, the lithologies of our cores Sieb P/06 and P/10 and Sieb 1966 look similar to each other, only the thickness of the described strata varies in low centimetre scale (Figure 2). The 3.85-m-long core Sieb P/06 mainly consists of greenish or greyish clayey silt with small intercalations of dark silt (2.12–1.91 m and 0.78–0.73 m). Horizons with a high content of calcareous material, interpreted as lake marl, are visible in a depth of 2.40–2.18, 1.60–1.47, 0.93–0.87 and 0.30–0.15 m.
Ostracoda
The state of ostracod preservation is good in general (Figure 3). Only a few valves are broken or show traces of dissolution. A total of 21 ostracod species were identified in the sediments of Siebleber Senke (see Appendix 1). For one of them, Limnocytherina santipatricii, only one valve was recorded, and it is therefore not included in distribution charts and ecological analysis. The most common species are Limnocythere inopinata, Ilyocypris bradyi, Ilyocypris gibba and Candona candida (Figures 4 and 5). A complete list of taxa can be found attached as a taxonomic reference list.

Ostracods of the Siebleber Senke; black scale bar = 0.1 mm. 1 – Pseudocandona sucki (left valve), 2 – Pseudocandona marchica (left valve), 3 – Pseudocandona rostrata (left valve), 4 – Candona neglecta (left valve), 5 – Limnocythere blankenbergensis (left valve), 6 – Candona candida (left valve), 7 – Heterocypris salina (right valve), 8 – Limnocythere inopinata (right valve), 9 – Ilyocypris gibba (left valve), 10 – Cyprideis torosa (right valve), 11 – Darwinula stevensoni (left valve), 12 – Ilyocypris bradyi (right valve), 13 – Cyclocypris ovum (left valve), 14 – Cypris pubera, 15 – Cyclocypris laevis (left valve).
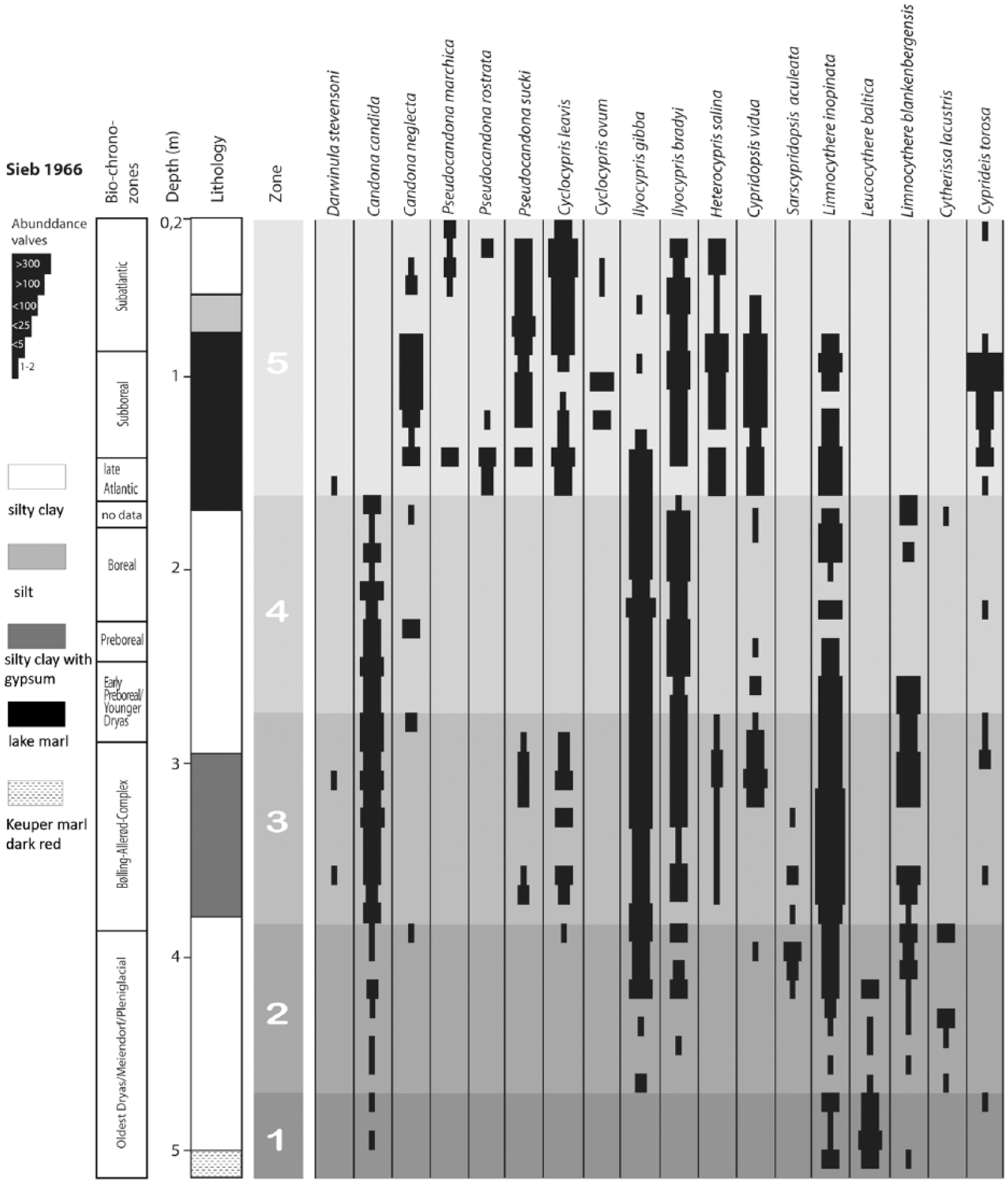
Lithology and ostracod distribution of the core Sieb 1966 and the hereon based unit differentiation. The chronozones are based on the pollen diagrams by Lange (1966) (Figure 6a) below 230 cm, and on this study above 230 cm (Figure 6b and c).
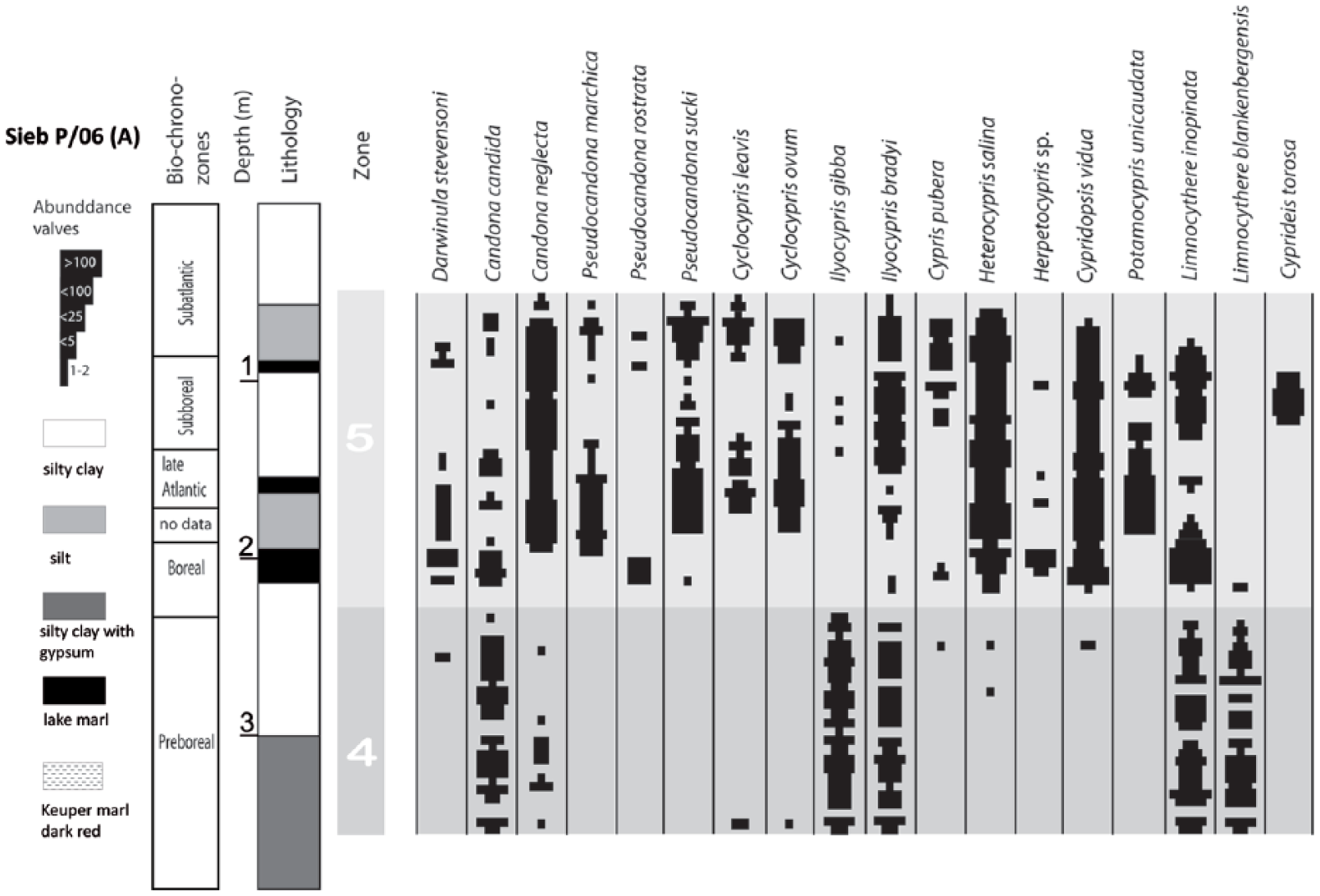
Based on ostracod distribution, a differentiation of core Sieb 1966 into five units (Units 1–5) and core Siebleben P/06 into two units (Units 4 and 5) is possible (Figures 4 and 5). The distribution of ostracods and especially a distinct horizon with a mass occurrence of Cyprideis torosa allow a correlation of the zones of both cores:
Unit 1 (525–470 cm). Only a few ostracods were found in this unit. The dominating species is Limnocythere inopinata; Limnocythere blankenbergensis, Limnocytherina santipatricii and Cytherissa lacustris occur very rarely.
Unit 2 (470–390 cm). Ostracod diversity increases distinctively. The dominant species are Limnocythere inopinata, Limnocythere blankenbergensis and Ilyocypris gibba. Less abundant are Sarscypridopsis aculeata, Cytherissa lacustris, Ilyocypris bradyi, Leucocythere baltica and Candona candida.
Unit 3 (390–285 cm). Ostracod diversity and abundance further increase. Dominating species are Candona candida, Limnocythere inopinata, Limnocythere blankenbergensis, Ilyocypris gibba, Ilyocypris bradyi and Cypridopsis vidua.
Unit 4 (285–165 cm). Whereas ostracod abundance is similar to that of the unit below, diversity decreases. Candona candida, Limnocythere inopinata, Limnocythere blankenbergensis, Ilyocypris gibba and Ilyocypris bradyi are still dominant, in contrast to the rare occurrence of Candona neglecta and Cypridopsis vidua.
Unit 5 (165–20 cm). This uppermost part of the section contains the highest diversity and abundance of Ostracoda. Cyprideis torosa appears in the lower part of the unit and reaches maximum occurrence in 115–95 cm. A total of 20–30% of the valves are noded. Sieve-pore analysis of the valves shows a mean percentage of 74% rounded sieve-pores. Abundant are Candona neglecta, Limnocythere inopinata, Ilyocypris bradyi, Cypridopsis vidua, Cyclocypris laevis, Heterocypris salina and Pseudocandona sucki. Less abundant are Ilyocypris gibba, Cyclocypris ovum, Pseudocandona rostrata, P. marchica and Darwinula stevensoni.
Palynology
Pollen concentration and preservation are very variable over the profiles. In core Sieb 1966, pollen are completely absent between 300 and 280 cm, 265 and 235 cm as well as in core Sieb P/06 and P/10 between 185 and 169 cm.
In core Sieb 1966 (Lange, 1966), the first pollen section (410–280 cm) documents (with the exception of two samples) a distinct Betula dominance followed by Pinus. Heliophilous herbs and open land species are regularly present. The local pollen component shows very low values. After a hiatus in pollen preservation (300–280 cm), a pollen association with high values in Pinus, lower values in Betula and a high amount of heliophilous herbs as well as Cyperaceae and Poaceae is documented between 280 and 265 cm. A second gap (265–235 cm) disrupts the pollen record, which is followed by an increase of Corylus values up to 20% and also first thermophilous trees between 235 and 215 cm. Trees now dominate the vegetation with nearly 90%.
Core Sieb P/06 shows a different pollen composition in the lower part (Figure 6b). The section between 385 and 290 cm reflects a Pinus dominance (40–60%), Betula values around 20% with relatively high values in heliophile herbs (Plantago major type, Artemisia, Cichorioideae). Pediastrum occurs as local component and increases between 290 and 207 cm core depth accompanied by an additional rise in Cyperaceae and Poaceae. A rise in Corylus combined with lower values in open land communities marks the last zone.

Palynological diagrams: (a) pollen diagram of core Sieb 1966 based on remarks of E Lange (1966); (b) pollen diagram of core Sieb P/06; (c) pollen diagram of core Sieb P/10.
In core Sieb P/10 the last pollen zone is also very clearly to correlate to core Sieb P/06 and show equivalent vegetation patterns until 185 cm. After this period, a phase without pollen preservation is documented until 169 cm. Then the pollen sequences show distinctly higher values in Quercus, Ulmus, Corylus, Tilia, Betula and Alnus. The local pollen component documents relatively high values of cane brake (Dryopteris, Sparganium, Equisetum, Typha latifolia) and wet meadow species (Filipendula ulmaria type, Cyperaceae, Galium type) as well as aquatics (Lemna, Pediastrum). Between 144 and 127 cm, Pinus is decreasing while Quercus and grassland communities rise in abundance. Within the local vegetation, aquatics are decreasing. The following section (127–87 cm) is once more characterized by higher values in Quercus, Corylus, Picea and grassland communities. Pinus and Ulmus show distinctly lower values. Aquatics like Pediastrum as well as wet meadow indicators are regularly found and Chenopodiaceae show increasing values. The vegetation development in the core section between 87 and 72 cm documents a rise in Alnus, Betula and Pinus as well as indicators for stronger human impact (Cerealia type, ruderal species), while Quercus, Tilia, Ulmus and Corylus decrease. In the local vegetation, Lemna, Potamogeton, cane brake communities with Equisetum, Dryopteris and Sparganium as well as Cyperaceae are documented. During the next phase (72–55 cm), Pinus and Quercus as well as indicators for meadows and open land communities are increasing in proportion. In contrast, aquatics (Lemna, Potamogeton) and plants of cane brake habitats are decreasing. In the upper phase between 55 and 44 cm, most tree species decrease and cereals as well as grassland and ruderal species increase in number. The local component documents cane brake species especially with Equisetum and Thelypteris as well as wet meadow types like Poaceae, Cyperaceae and Cichorioideae.
Interpretation and discussion
Biostratigraphical interpretation
According to the regional pollen composition documented in core Sieb 1966 by Lange (1966), the phase between 410 and 285 cm is to correlate with the late Glacial (Figure 6a). Warmer interstadial phases and cooler stadial phases are documented by different values of open land communities and forest habitats within the analysable sequences. The permanently high values of Betula could document the correlation of this phase into the Bølling Interstadial (Litt and Stebich, 1999; Schneider, 2006), but the value changes in heliophilous herbs within this phase point to different stadials and interstadials. Probably Betula was also a part of the local vegetation and grew in the marginal areas of the depression. For solving this problem, Lange (1966) discussed two different chronological models – one according to tree pollen composition and another one according to general pollen combination with open land communities. Between 315 and 300 cm in core Sieb 1966, the Artemisia values reach more than 30%. Such high values are characteristic for steppe communities during the cold stadials (Litt and Stebich, 1999). In this case, trees as well as other open land communities document very low values, which indicate a very local signal of Artemisia as a result of populations at the margins of the depression. A hiatus in pollen between 300 and 280 cm in Sieb P/10 is possibly stratigraphically correlated to the Younger Dryas period. The following section (280–250 cm in core Sieb 1966, 385–207 cm in core Sieb P/06) reflects pollen spectra with higher values in Pinus, lower values in Betula and a high amount of open land communities pointing to an early Preboreal vegetation phase (comp. Schneider, 2006). This is also confirmed by the temperature curves (Figure 7). The rise of Cyperaceae and Poaceae between 290 and 207 cm in core P/06 is comparable to the pollen association in core Sieb 1966 (Lange, 1966) between 280 and 265 cm. From 235 cm, the rise of Corylus and first proofs of thermophilous trees indicate the early Boreal, which is also visible in the pollen diagram of core Sieb P/06 and P/10 (Figure 6b and c).
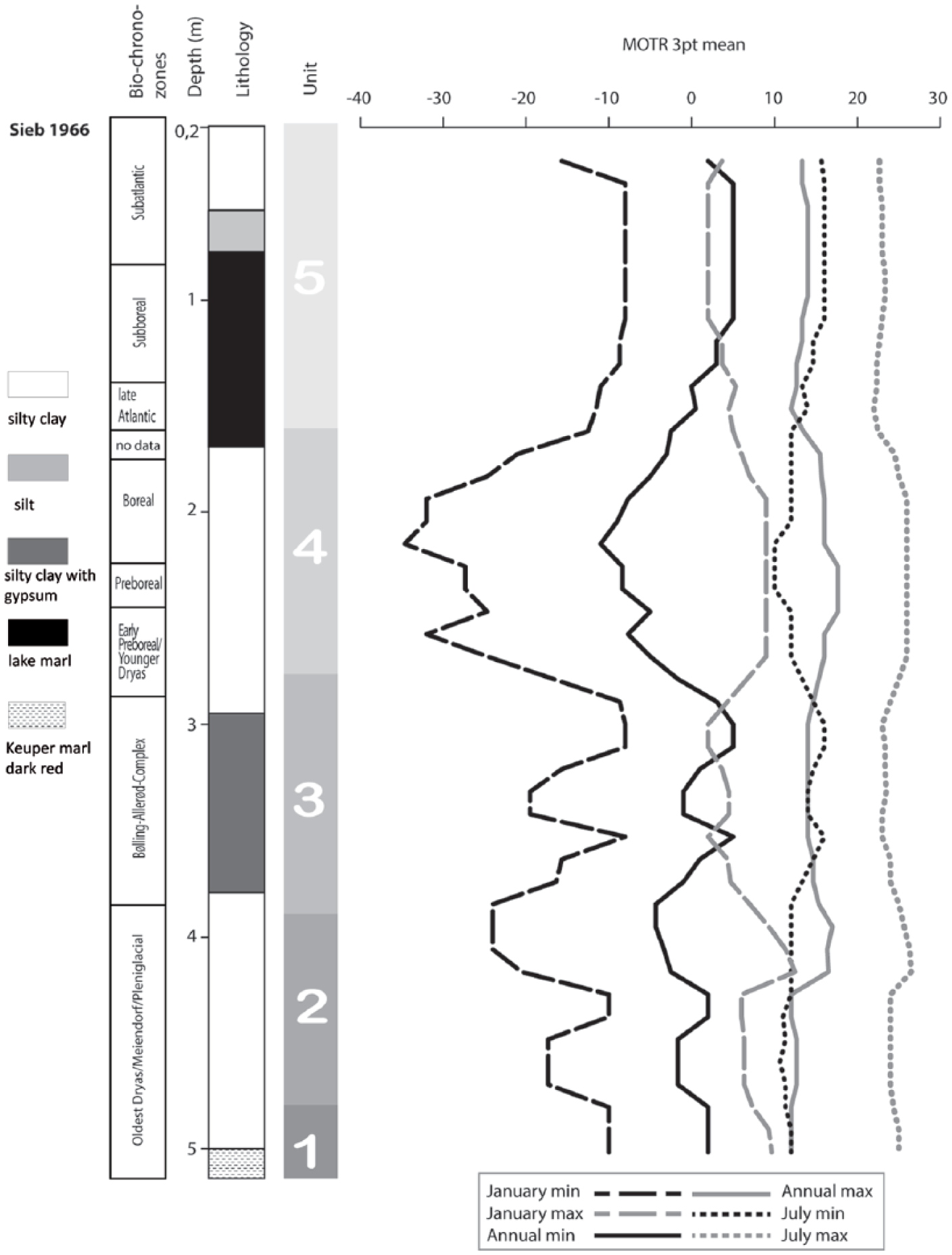
Synopsis of Mutual Ostracod Temperature Range (MOTR) and Transfer function (TF) results of the ostracod species of Siebleben based on the ostracod distribution of core Sieb 1966.
An AMS radiocarbon dating, taken from gyrogonites of 350–355 cm in Sieb P/06, presents an age of 11,605–11,187 cal. BP. This points to a reservoir effect affecting skeletons of aquatic organisms in the lake. Due to the lack of any terrestrial datable material, it was not possible to calculate the reservoir effect; the chronological difference between the end of the Preboral phase and the beginning of the Boreal suggests a minimum error of around 500 years (Bos et al., 2007). After the boreal Corylus peak, a sediment accumulation without pollen conservation is documented in all cores and is typical for this landscape (Schneider, 2012). In this position, Lange (1966) stopped pollen analysis in core Sieb 1966 and the following Holocene development bases on the investigation of core Sieb P/10 (Figure 6b).
In core Sieb P/10 the last pollen zone is very clearly to correlate to core Sieb P/06 and shows similar vegetation patterns until 185 cm. The core section in Sieb P/10 starting at 167 cm shows a typical mixed forest with Tilia, Ulmus and Quercus which points to the late Atlantic (Firbas, 1949; Schneider, 2006). The following zone (144–87 cm) reflects the biozone of the Subboreal with an increase in Quercus, Picea and Fagus. According to the changes in local scale, the zone is divided into two subzones, the early Subboreal (144–127 cm) and the late Subboreal (127–87 cm). Shells of the gastropod Gyraulus sp. of the horizon (Sieb P/06 120–115) with the highest abundance of Cyprideis torosa were AMS radiocarbon dated to 4977–5286 cal. BP (Table 1). As for the other radiocarbon date, a reservoir effect can be assumed.
The pollen composition of the next section (87–55 cm) documents the early Subatlantic phase with an increase in Fagus, Picea and Abies together with a rise in pioneer trees. It shows in its second part a further increase in Quercus and Pinus as well (Schneider, 2006). In the younger phase between 55 and 44 cm, most trees are decreasing and indicators for anthropogenic use (Behre, 1981) are rising distinctly in proportion. This pollen composition indicates the late Subatlantic (Firbas, 1949).
A biostratigraphical identification of sediment units from the core Sieb 1966 based on ostracods is only roughly possible. The ostracod-based stratigraphical classification for Central Germany provided by Fuhrmann (2013) allows a differentiation between late Glacial and Holocene sediments for our section, but no further differentiation within the Holocene. Considering the general shallow water character of the palaeohabitats at Siebleben, the ostracod faunas of Unit 1 and Unit 2 reflect late Glacial conditions (Leucocythere baltica, Cytherissa lacustris). All the following units are typical for warmer conditions. The occurrence of such an ostracod fauna more typical for the Holocene below 300 cm within a late Glacial pollen association indicates interstadial conditions which could be typical for late glacial interstadials (Litt and Stebich, 1999; Schneider, 2006).
Palaeoenvironmental reconstruction
The following palaeoenvironmental reconstruction for the core Sieb 1966 is based on ecological interpretation of ostracod associations following data given in Meisch (2000), Fuhrmann (2006) and Pint et al. (2012) applying MOTR, MOSR, ostracod-based transfer function, palynological analysis as well as lithological observations. In general, the ostracod distribution patterns of the units are different from those of recent sites (Pint et al., 2015). However, all sites except Stausee Kelbra are small and mostly temporary waters. Even the ostracod assemblage of the small modern pond of the Siebleber Senke is quite different from the associations from the cores. It is dominated by Cyprois marginata and Cypris pubera, typical for small temporary ponds.
Palaeotemperature reconstructions made with the MOTR method often produce wide ranges of air temperatures which are difficult to interpret because the actual temperature could have been anywhere within the given range.
Salinity analyses show that, in general, curves of transfer function and modern analogue lay within the MOSR. One notable exception is located in zone 5 based on the short-term dominance of Cyprideis torosa which produced a higher peak of the salinity transfer function curve.
Unit 1 (Oldest Dryas/Meiendorf/Pleniglacial; 525–470 cm)
The ostracod record starts with a typical pioneer assemblage (Limnocythere inopinata, Candona candida and Leucocythere baltica) characteristic for the late Glacial (Diebel, 1965; Frenzel and Viehberg, 2004; Griffiths and Evans, 1995). More than 80% of the valves indicate permanent and cold freshwater – these species belong to the ‘oligostenothermal/boreo-alpine’ group of Fuhrmann (2006). The MOTR method provides a mean annual air temperature range of +2 to +12°C. This result is mainly controlled by Leucocythere baltica (for the MOTR, the synonymous Leucocythere mirabilis is used), the most common species in Unit 1. The estimated air temperature range is very broad because of the low species number and does not facilitate precise palaeoclimatic reconstruction of the initial lake phase. We assume a permanent water body, probably a shallow lake, under cold climatic conditions. The missing pollen record could be caused by a fluctuating water level in the shallow marginal zone of the lake. A saline influence is not recognizable if looking for brackish water species, despite the very rare occurrence of the brackish water ostracod Cyprideis torosa. MOSR provides a large range of salinity from 0 to 5. The ostracod-based transfer function for salinity estimates 1.5–2.3 psu, which indicate oligohaline conditions making a saline water discharge probable.
Unit 2 (Oldest Dryas/Meiendorf/Pleniglacial: 470–390 cm)
The much higher ostracod diversity and abundance of this unit point to ecologically improving and probably more stable environmental conditions likely coupled to higher productivity. The high proportion (40%) of species living in permanent waters indicates a permanent water body. Regarding the presence of mesorheophilic and the lack of polyrheophilic taxa, a shallow lake seems most likely (Pint et al., 2015) which is also documented in the pollen results by Lange (in Greifeld, 1966) with wide wet meadow areas (Galium, Thalictrum, Cyperaceae) within the depression. Using the classification by Fuhrmann (2006), rheophobic species are dominant. The typical glacial species Leucocythere baltica indicates cold climatic conditions (Diebel, 1965) in agreement with Cytherissa lacustris which is regarded as a cold water species (Meisch, 2000). On the other hand, both species perish within this unit and polythermophilic species start to increase in abundance. This attests to at least warmer summers than in the previous unit. The MOTR analysis suggests an annual temperature range of −9 to +16°C within the zone, controlled by Leucocythere baltica and Candona candida. Maybe a distinct continentally of the regional climate produced cold winters and warm summers. The ostracod-based salinity transfer function estimates oligohaline conditions (c. 2) as before. Palynological analyses of Lange (in Greifeld, 1966) show a mosaic of light birch forests and open land habitats with grasses and heliophilous herbs like Cistaceae, Plantago spec. and Artemisia during this period.
Unit 3 (Bølling/Allerød-Complex – Younger Dryas: 390–285 cm)
Ostracod diversity and abundance further increase. This observation and the lithological change from silty clays to calcareous muds point to increased temperatures and productivity. Cold stenothermal species disappear and taxa preferring warm water such as Ilyocypris gibba, Cypridopsis vidua and the eurythermal species Limnocythere inopinata (Fuhrmann, 2006; Meisch, 2000) show highest proportions within the entire succession. Hence, warm waters of the shallow lake in summer and cold winters as indicated by the high proportion of Candona candida, a winter form (Meisch, 2000), are assumed. The MOTR analysis suggests a mean annual temperature range of −7 to +16°C, and similar variations as in Units 1 and 2. Except the upper limit of July, which is determined by Cyclocypris laevis, the MOTR is mainly controlled by Pseudocandona sucki, which occurs frequently. The temperature range is not significantly different from that of the preceding unit, but the limits of the winter, summer and annual ranges are closer to each other. The pollen record between 390 and 300 cm documents, with the exception of two samples, still a distinct Betula dominance followed by Pinus. A short peak of non-arboreal pollen (upland herbs), Artemisia, Chenopodiaceae and Thalictrum in between (Lange, 1966; Sieb 1966), points to a short phase with dryer conditions possibly during the Allerød. In general, the local pollen component shows very low values and Pediastrum increases, which hints at open water conditions until a depth of 300 cm. These conditions are also visible by high percentages of ostracod species living in permanent water bodies, and the rheoeuryplastic to mesorheophil association indicates a shallow lake. Furthermore, because Cyprideis torosa has no desiccation resistant eggs (Heip, 1976), the brackish water character of the water body has to be concluded as permanent at least in parts of it. The small percentage of species from perennial waters could reflect a littoral zone with water level fluctuations, or the transition to temporary water. The mesohalophilic species increase remarkably in proportion and a small but growing proportion of the brackish water ostracod Cyprideis torosa appears. This pattern reflects an increasing saline influence by salty brines from the underground. The ostracod-based salinity transfer function estimates slightly increased (c. 2.5) oligohaline conditions. The sieve-pore analysis on valves of Cyprideis torosa indicates a salinity of about 1 psu, also indicated by 20–30% of noded valves of Cyprideis torosa. According to Frenzel et al. (2012), this corresponds to a salinity of approximately 1.7 psu.
Unit 4 (Younger Dryas, Preboreal, early Boreal and late Atlantic; 285–165 cm)
Unit 4 is characterized by decreasing diversity of the ostracod associations pointing to less stable or degrading ecological conditions. Lowered proportions of polythermophilic species and a more than doubled proportion of oligothermal species sensu Meisch (2000) point to lower water temperatures and coincide with a lithological change from calcareous mud to siliciclastically dominated sediment. Regarding the high percentage of species living in permanent waters and the lack of such from temporary waters, the Siebleber Senke was at least partly covered by a permanent lake. The local vegetation is characterized by fluctuating values of local pollen types and points to a shallow water system where two dryer phases led to temporary drying disabling pollen conservation. Hence, permanent water bodies were present but did not exist over the whole time period under consideration.
Between 280 and 265 cm, a wetter phase allows the reconstruction of wet meadow communities with Cyperaceae in the marginal areas of the water filled depression (Lange, 1966). The regional pollen combination at the transition between late Glacial and Holocene indicates a light pine forest with a high amount of heliophilous herbs, while birches grew in the surrounding area of the depression and in the floodplains. The section between 265 and 235 cm is characterized by a pollen hiatus in core Sieb 1966 (Lange, 1966), but not in core Sieb P/10. The reason for this phenomenon is unclear. One possible explanation is the very local change of subsurface depth because of the pattern of salt solution in large subrosion hollows (Igl, 2000).
Between 235 and 215 cm in core Sieb 1966 as well as between 207 and 185 cm in core Sieb P/06, pine–birch forests with an increasing amount of hazel as understorey and also first elms, limes and oaks as indicators for warmer conditions grew in flood plain areas. At the end of the zone, a rise in charcoal pieces points to an increase in natural fire activity (Vannière et al., 2008).
Clearly reduced numbers of poly- and mesohaline ostracod species indicate a dwindling saline influence. This is reflected by salinity estimations of less than 1 by the transfer function. The MOTR analysis provides a mean annual temperature range of −11 to +17°C, mean January of −32 to +9°C and mean July of +10 to +26°C. Actual temperatures could have been anywhere within these ranges, but the minimum value for mean January temperature is significantly lower than in the other units, with a corresponding cooling trend reflected in the mean annual and mean July curves, suggesting the possibility of significantly colder winters and possibly cooler summers. The lower limit of the July range is defined by the frequently occurring Limnocythere inopinata and the upper limit is set by Candona candida and the very rare Cytherissa lacustris. This represents a dryer and more continental climate than in the other zones.
Starting with 192 cm, sediments are changing from lake marl to a very thin layer with small stones and concretions which is overlain by black brown silty clays with bad pollen preservation. In core Sieb P/10 between 185 and 169 cm, the sediments show a hiatus in pollen preservation. According to the pollen combination below and above this section, the hiatus covers the period between beginning of the late Boreal and end of the early Atlantic. Different investigations in several geo-archives in Thuringia document a drastic change in sedimentation during this period, indicating lower lake or groundwater levels (Schneider, 2012), the decrease in sedimentation rates in bogs and fens or in some cases the loss of material because of erosion processes (hiatus). The ostracod assemblages do not show any changes until the end of the pollen hiatus, but due to sampling strategy the differences in this short part of the section are probably not detectable.
Unit 5 (late Atlantic to late Subatlantic; 165–20 cm)
Unit 5 yields the maximum ostracod species number of the studied core. A reason for this high diversity could be the higher winter temperature as indicated by returning calcareous sedimentation and a slightly higher proportion of meso- to polythermophilic species. The air temperature range of the MOTR analysis is only controlled by Pseudocandona sucki and comprises a mean annual temperature range of 0 to +14°C. The MOTR of zone 5 reflects the highest winter and the lowest summer temperatures of the section with less variation. Up to 144 cm, a mixed forest with oak, elm, hazel and lime grew in the surrounding area, while birch and alder dominated in the wet areas. The local pollen component documents a system with large cane brake and wet meadow areas as well as very shallow open water habitats with Lemna and Pediastrum. The very high number of charcoal pieces implicates a higher intensity in natural fires as a result of dryer climate conditions. On the other hand, this phenomenon could also suggest an increasing human impact, which is proven by pollen grains of cereals and forest disturbing indicators like Pteridium. Later, pine woods decrease and mixed forests with a high amount of oak extend in the pollen zone of 144–127 cm. Increasing grassland communities point to stronger anthropogenic influences. Within the depression, aquatics are decreasing in number and document a fen with small open water areas. The following section (127–87 cm) is once more characterized by a spreading of oak, hazel, and grassland communities, which points to an anthropogenic use by coppice with standards. Wet meadow indicators like Cyperaceae and the algae Pediastrum extended in the archive and represent more humid conditions. Here, the abundance maximum of the brackish water ostracod Cyprideis torosa is reached indicating higher salinities and permanent waters. This increase coincides with a drastic rise in Chenopodiaceae and offers evidence for a local interpretation and also an indicator function of this pollen type during periods with higher salinity. Local spreading of Chenopodiaceae in salt solution depressions is actually visible in the Werra valley (Schuster et al., 2010). The vegetation development in the upper section (87–72 cm) of Unit 5 documents very disturbed forest habitats with pioneer trees (birch, pine, alder) according to human impact. Tree species of the submontane forest like beech, spruce and fir extend and show forest regeneration in the neighbouring regions. The depression is characterized by a permanent water surface and wide margins with cane brake and wet meadows. During the next phase (72–55 cm), pine and oak are spreading again, while all other tree species decline, and indicators for meadows and open land communities are rising in proportion. This combination points to a clear human impact. In the depression, open water and cane brake habitats decrease when Cyprideis torosa disappears, probably because of the loss of a permanent brackish water body. Between 55 and 44 cm, most forest habitats are cleared and indicators for arable land (cereals), meadows (Plantago lanceolate, Plantago major, Polygonum bistorta) and ruderals (Artemisia, Polygonum aviculare) document an intensively used landscape (Meschner, 2008; Schneider, 2006), which enclosed a wet depression, where reed communities and wet meadows extended. A relatively high salinity of 1–3 between 160- and 100-cm sediment depth and the very high proportion of Cyprideis torosa imply permanent brackish water conditions, a strong ecological stressor for most of the freshwater species. Therefore, we assume a more and more fragmented lake broken into separated smaller basins with different habitats and varying salinity. A fluctuating water level and rain-wash could cause a mixing of different associations over longer time. The highest proportion (8%) of species living in temporary waters within the core fits this model. A slight influence of a surface inflow is also recognizable. The lower species number in the upper half of this unit and the change to siliciclastic sedimentation reflects the terminal silting up phase of the basin development. The highest numbers of phytophilic ostracod species indicate the existence of a broad reed fringe during silting up of the lake, similar to present day conditions.
Discussion of proxies and methods
The MOTR method was applied to the five assemblages identified in the sequence (Figure 7), using calibrations (Table 3) based on those published by Horne and Mezquita (2008) but revised by reference to the OMEGA (Horne et al., 2011) in a few cases of species which occur in North America as well as Europe. Several species are not calibrated and could not be used in the analyses, because they are extinct or because of taxonomic uncertainty: Ilyocypris gibba, Ilyocypris bradyi and Limnocythere blankenbergensis. Cyprideis torosa has not been calibrated because, as a brackish water species, its distribution is not well-documented in the non-marine databases used. The MOTR results are illustrated in Figure 7; they show disappointingly little variation, probably because of the relatively low number of calibrated species in each assemblage, although the reconstructed temperature ranges are at least consistent with other palaeoclimate proxy data for this region and interval. For example, for Unit 3 the MOTR July range is +12 to +23°C and the January range is −10 to +7°C. Pollen-based palaeotemperature reconstructions by Davis et al. (2003) indicate summers around 2° cooler and winters 7° cooler than today (i.e. around +15 and −8°C, respectively), both values falling within the MOTR estimates. Similarly, pollen-based palaeotemperatures for July and January for the past 6000 cal. yr (mid- to late-Holocene) have varied by less than 1° either way from modern values (Davis et al., 2003). This falls within the MOTR estimates for Unit 5 (Figure 7), while, as already noted above, the MOTR-based reconstructions possibly indicate a significant cooling in Unit 4, which corresponds at least in part with the Younger Dryas cold interval that immediately preceded the Holocene.
MOTR calibrations based on Horne and Mezquita (2008) with updates from OMEGA, December 2011.
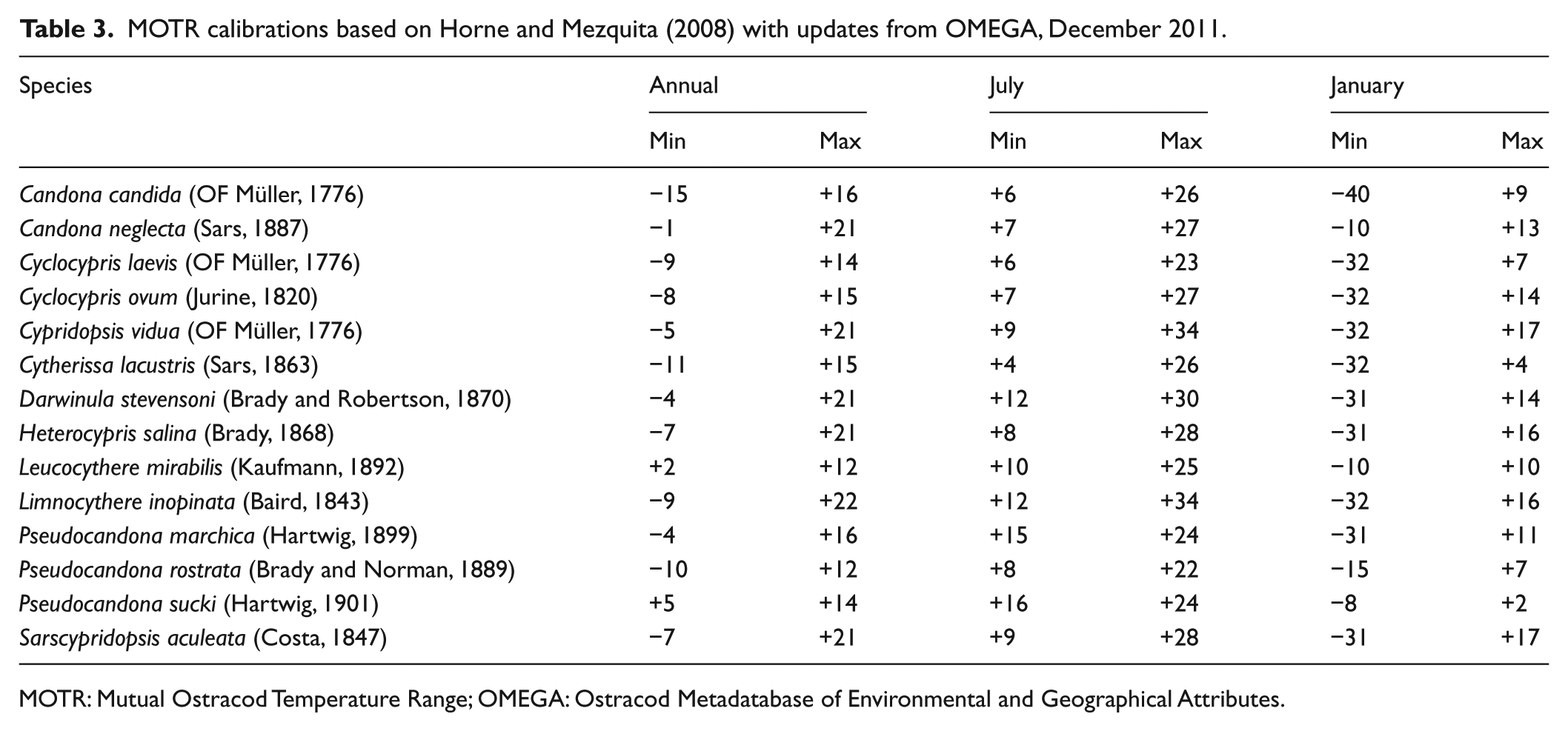
MOTR: Mutual Ostracod Temperature Range; OMEGA: Ostracod Metadatabase of Environmental and Geographical Attributes.
For the Recent ostracod association in Thuringia (Pint et al., 2015), the MOTR method was applied for all species having a reference in the MOTR calibration list (Horne and Mezquita, 2008). The reconstructed air temperatures for the region of these sites lie (July: +17.5°C, January: −0.5°C) within ±2°C of modern measured mean air temperature values. This means that the test of MOTR on Recent sites of the Thuringian Basin works very well and can be applied to the section of Siebleben. The MOTRs of the fossil units are mainly controlled by species with low relative abundance and low frequency. As expected, units containing low species numbers produce a wide range for temperature estimation. The estimations by MOTR for the Siebleben core present no distinct variation between the units and are usually less sensitive than index species methods or ostracod-based transfer functions. One reason is the exclusive application of absolute ecological tolerances and for a small number of species only. On the other hand, Ilyocypris gibba and Ilyocypris bradyi, two of the most frequent species of the Siebleber Senke, have not yet been calibrated.
Similar to the MOTR method, the salinity range of the MOSR method is controlled by a few species. Using salinity tolerance ranges from Frenzel et al. (2010), the salinity estimations are 0–3 for Unit 1, 0.5–1.8 for Unit 2, 0.5–4 for Unit 3, 0.5–1.8 for Unit 4 and 0.5–3 for Unit 5 (Figure 8). The ostracod-based transfer function for salinity, its modern analogue counterpart, sieve-pore and node analyses in Cyprideis torosa fall within the limits of the MOSR method which outlines extreme values taken from the literature (Frenzel et al., 2010). The other methods show variations not recorded by MOSR indicating oligohaline values between 1 and 3 psu in general with the exception of the Cyprideis torosa horizon as mirrored by the modern analogue technique and Unit 4 at the limit to freshwater conditions. We conclude a higher sensitivity of the methods relying on transfer functions and intraspecific morphological variability compared with the mutual range methods based on species presence alone instead of relative abundances.
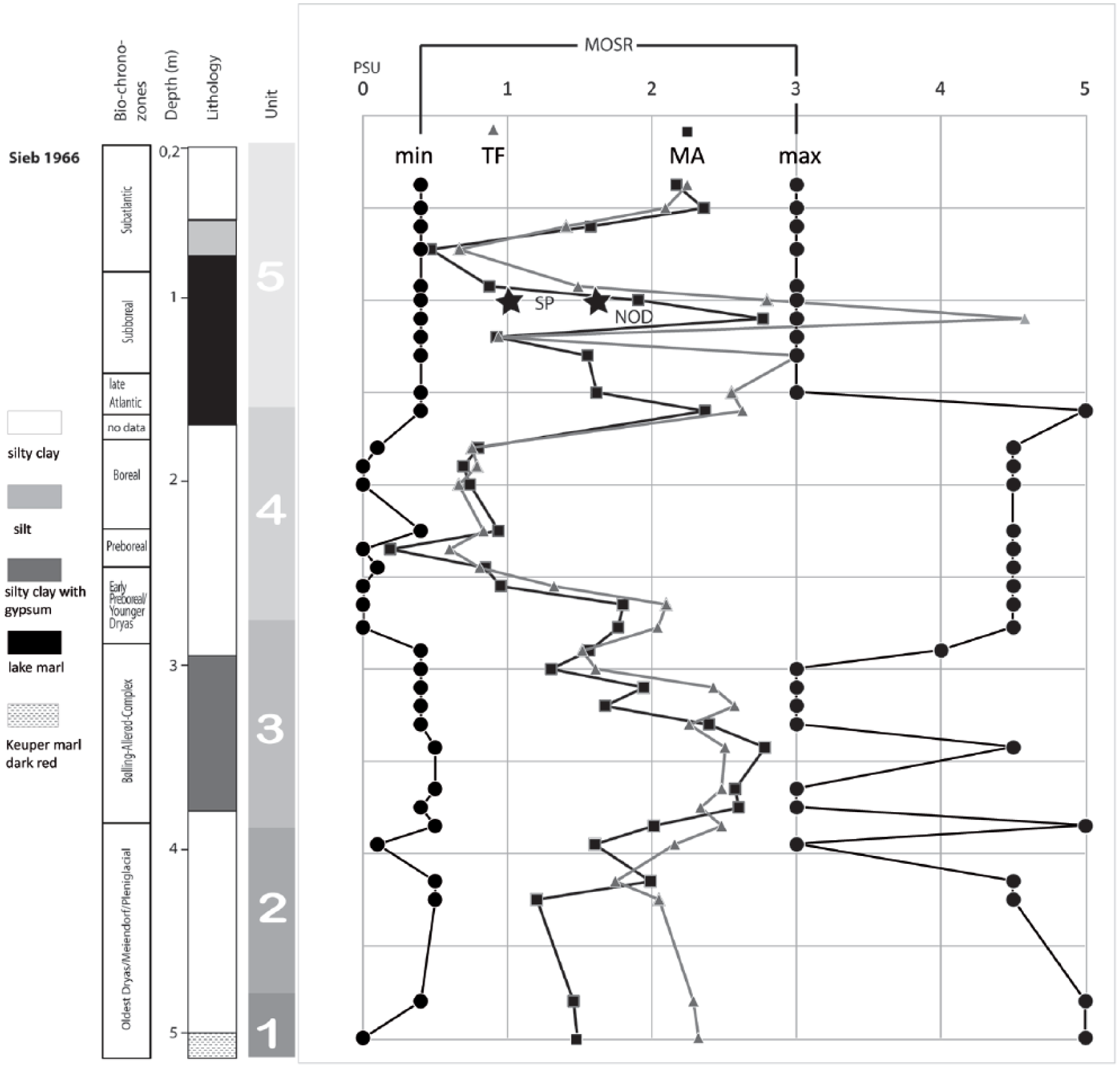
Synopsis of Mutual Ostracod Salinity Range (MOSR), Transfer function (TF) and Modern Analogue (MA) results based on the ostracod distribution of core Sieb 1966. The stars indicate estimated salinity values based on sieve-pore analysis (SP) and node analysis (NOD).
Elevated salinity in a continental water body could be explained either by salt brine inflow or negative precipitation/inflow–evaporation balance. A climatically induced salinization would be caused by stronger continentally (colder winters and hotter summers). Such a climate pattern would be recognizable in reconstructing air temperatures by MOTR. The only zone of the studied core revealing more continentally is Unit 4. The ostracod association of this unit, however, does not indicate elevated salinity but the lowest values of the entire succession. We can therefore conclude for Siebleben that higher salinity is driven by brines caused by subrosion.
The ostracod associations of core Sieb 1966 reflect permanent water bodies in the Siebleben basin from the base of the core to the surface. The uppermost samples point to silting up of the lake. In contrast, the pollen analyses show three periods without pollen preservation, pointing to phases with temporary drying out. Two of those phases are documented during the late Glacial, the third one is visible from the start of the late Boreal to the end of the early Atlantic. The reason for this phenomenon is the different chronological resolution of the samples and preservation problems. While pollen grains are destroyed very fast under aerobic conditions (Moore et al., 1991), remains of ostracods and foraminifers are not sensitive to this. Sedimentation hiati could have destroyed the pollen during phase of drying out.
A brackish water influence is visible during the warmest periods, in the Allerød and Atlantic to Subboreal, times which are reported to be wettest with widely traceable subrosion processes in the study area (e.g. Schneider, 2006). Hence, climatic conditions are the driving factor for subrosion or, inversely, salt water signals reflect humid phases in climate evolution of this area with evaporites not deep below the ground. Ostracods, in this case the brackish water indicator Cyprideis torosa, prove to be a valuable salinity proxy.
Shells of freshwater molluscs found on reed beds in Thuringia like those of the Esperstedter Ried and Grossengottern Ried point to larger postglacial lakes that have vanished. Despite the occurrence of halophytic plants, the modern ostracod fauna in the recent residual ponds situated in those reed areas do not contain brackish water ostracod taxa (Pint et al., 2015). The extension of postglacial lakes in Central Germany in space, time and its climatic implication needs further investigations to figure out saline water influences as local events or probably climatic controlled effects.
Summary of late glacial to Holocene site evolution
The documented late Glacial period (525–c. 260 cm in core Sieb 1966) is characterized by the existence of a water body in the Siebleber Senke. Warmer and more productive phases of the Bølling–Allerød-Complex are indicated by changes in the ostracod and pollen association. The water of the Siebleben shallow lake(s) was slightly brackish (oligohaline) and the lake level fluctuated. We assume subrosion of Keuper salt by a rising groundwater table as the origin of the salinization. The slightly rising salinity points to increasing saline water discharge caused by more precipitation during the Bølling–Allerød-Complex if compared with dry and cold climate conditions of the Dryas. Additionally, dissolution of salts from sediments of the periglacial landscape through increased precipitation could add to salt inflow. The landscape was dominated by birch woods and open land vegetation.
During the early Preboreal (>285 to at least 250 cm in core Sieb 1966 and 385–207 cm in core Sieb P/06), small groups of pine and birch surrounded the temporary waters in a slightly cooler climate. Saltwater discharge decreased because of a sinking groundwater table. Hazel and thermophilous trees appear in the pine and birch woods of the early Boreal (>235 to at least 215 cm in core Sieb 1966, 207–185 cm in cores Sieb P/06 and P/10) in a warmer climate. Apparently, salt water discharge does not play a role in this time. Hence, we assume small temporary rain fed ponds in the depression of Siebleben.
Between 183 and 167 cm (core Sieb P/10), a gap in pollen preservation documents a change in hydrology from late Boreal to early Atlantic.
In the late Atlantic (167–145 cm in core Sieb P/10), the temperature increased as indicated by a higher proportion of meso- to polythermophilic species, higher ostracod diversity, tree pollen (e.g. oak and hazel) and returning calcareous sedimentation. The pollen record reflects shallow open water habitats, cane brakes and wet meadows in the depression of Siebleben. The area was surrounded by a mixed forest often affected by natural fires caused by dryer climate conditions. Subsequently, the increasing anthropogenic influence during Subboreal (145–85 cm in core Sieb P/10) becomes clear because of the extension of grassland communities and forest use indicating tree pollen combination (Behre, 1981). The areas of open water decreased within the depression. The high abundance of Cyprideis torosa and halophytic plants demonstrates brackish conditions of the still permanent water. The increasing salinity is likely caused by the inflow of saline waters deriving from leaching of salt bearing sediments of the Triassic basement. During the progressive siltation, the water was likely separated into small ponds with different ecological conditions.
Conclusion
Although the ostracod distribution of the section of Siebleben suggests strong ecological variations, not all the results of the applied methods reflect these changes. The reasons for the partly low performance of mutual ecological tolerance methods as well as the transfer function are the low species number and often the dominance of very tolerant species. The lack of studied comparable recent water bodies complicates an actualistic analysis. However, the combination of the pollen and ostracod datasets provides a consistent basis for the palaeoenvironmental reconstruction of the Siebleber Senke. A larger lake existed there during the Holocene and was temporarily influenced by salt brines. These probably short-term periods are likely caused by subrosion of salt bearing sediments of the Triassic underground. Wetter climate periods enhanced subrosion by a higher groundwater level producing salty brines locally.
The presented study demonstrates the need for more ecological data of several ostracod species as well as for investigation of palaeoecology of extinct species. This would enhance the performance of MOTR and the transfer function considerably. We can already state, however, that the combination of ostracod and fossil pollen analyses allows a comprehensive reconstruction of hydrological and climatic conditions as well as site evolution, also for the surrounding area. The different methods complement each other and enable conclusions, which would be impossible by using only one method.
Footnotes
Appendix 1
Acknowledgements
David Lazarus (Museum für Naturkunde Berlin) enabled access to the Diebel-Pietrzeniuk collections.
Funding
The author(s) received no financial support for the research, authorship and/or publication of this article.