
Abstract
The archaeological research focusing on El Mirador Cave (Sierra de Atapuerca, Spain) has revealed a succession of occupation levels in a dung layers context that span from the early Neolithic to the middle Bronze Age. The robustness and coherence of the chronological dates of the sequence have contributed to framing the beginning of farming practices on the northern Plateau of the Iberian Peninsula. This study focuses on the palynological analysis of the sedimentary sequence, spanning from ca. 7970–7770 to ca. 3390–3070 cal. yr BP. The results have allowed us to identify a landscape of mixed forest with evergreen and deciduous oaks and pinewood. Despite the discontinuities in such sediments, some fluctuations between different categories of anthropogenic taxa can be observed throughout the sequence, illustrating greater or lesser pressure from livestock or agriculture. From the Neolithic to the Bronze Age, we documented a reduction in the tree cover because of increased human pressure typical of the Neolithisation process, while the relationship between environment and society changes.
Introduction
The palynological studies of most of the deposits on the Iberian Peninsula indicate that human communities started to modify the landscape about 7500 years ago (Pérez-Obiol et al., 2011; Riera-Mora et al., 2004). However, the definitive establishment of ‘cultural landscapes’ came later, during the Chalcolithic and Bronze Age (Atienza-Ballano, 1995; Carrión, 2002; Carrión et al., 2001a, 2001b, 2007; Ruiz-Zapata et al., 2003, 2006).
Human occupation has played a decisive role in shaping the present landscape of the Peninsula since the middle Holocene. The effects of human activities on the environment are reflected in the changes in the composition of plant communities because of deforestation, use of fire, agricultural practices and use of grassland areas as pastures for livestock. Still, anthropogenic changes are detected erratically on the Iberian Peninsula, especially in the pollen records obtained in mountainous areas (Carrión et al., 2010; Riera-Mora et al., 2004; Sánchez-Goñi and Hannon, 1999). The palynological analysis of archaeological deposits clearly gives us the opportunity to more directly determine the magnitude of the anthropogenic footprint on the landscape (Bueno et al., 2002; Burjachs and Expósito, 2007; Carrión et al., 2010; Delibes de Castro et al., 2015; López-García and López-Sáez, 1994, 2000; López-Sáez, 2002; 2014; López-Sáez and López-Merino, 2007; López-Sáez et al., 2006, Ontañón et al., 2013; Ruiz-Zapata et al., 2003).
In addition to palynological analyses, the identification and quantification of micro-algae, fungi, chitinous remains and other organisms (non-pollen palynomorphs (NPPs)) have emerged in recent decades as a further source of information. The input of NPPs contributes additional information about the local environment that framed the formation of the deposits, as well as about the intensity of the anthropogenic activities that took place at a given site (López-Sáez and López-Merino, 2007; López-Sáez et al., 1998, 2000; Van Geel et al., 1989).
The aim of this study is to contribute to the knowledge of the Neolithisation process in the Iberian Peninsula from the palynological evidence of the farming practices identified in El Mirador sequence. Almost the entire deposit is made up of stabling layers originating from in situ burning of animal dung. This resulted in a typical fumier with a peculiar appearance that comes from the alternation of burned and non-burned livestock dung (Angelucci et al., 2009; Vergès et al., 2002, 2008, 2016).
Generally, these kinds of accumulations are rich in archaeobotanical material because they are formed from the repeated burning of waste generated after the use of these caves as a fold for cattle (Brochier et al., 1992; Canti, 1999; Vergès et al., 2008). Burning an accretion of faeces serves several purposes: deworming, reducing the volume of the accumulated manure and so on. This activity was very common during the Neolithic and Bronze Age in Europe (Badal, 1999; Charles, 1998) and has been identified at numerous sites, such as Arene Candide in Italy (Maggi, 1997), Caune de Belesta in France (Brochier et al., 1998) and Cova de les Cendres (Badal, 1999), Los Husos (Fernández-Eraso, 2010; Polo-Díaz and Fernández-Eraso, 2008), Cova Gran (Polo-Díaz et al., 2014), El Mirón (Peña-Chocarro et al., 2005), Peña Larga, Peña Parda and San Cristóbal (Ruiz-Alonso et al., 2016) inter alia on the Iberian Peninsula.
The study area and archaeological background
The Sierra de Atapuerca (Burgos, Spain) is located in the foothills of the Iberian Range and is composed of Upper Cretaceous limestone and dolomites (Cenomanian and Turonian). It is a complex groundwater-source karstic system that originated during the Neogene/early Pleistocene in the late Cretaceous (Ortega et al., 2013; Zazo et al., 1983) and continued developing in the Quaternary.
The mountain range is located at a biogeographic crossroads where the north sub-plateau, Iberian Range and Cantabrian Range converge. This geographical context in which the Eurosiberian and the Mediterranean regions converge contains a mixture of ecological trends (Pérez-González et al., 1995, 1999; Torres, 1976; Zazo et al., 1983, 1987).
Current vegetation is a mixed open continental forest of Quercus rotundifolia (=Quercus ilex subsp. Ballota) and Quercus faginea with an Atlantic influence evidenced by the presence of Erica vagans L. and Quercus pyrenaica, which grows on the Arlanzón terraces on siliceous soils. The intensity of the anthropic activities such as fuelwood exploitation and grazing has influenced its physiognomy, characterised by the bushy appearance of oaks (García-Antón et al., 1995).
El Mirador Cave is located about 1033 m a.s.l. at 42°20′58″ N and 03°30′33″ W, on the southern slope of the Sierra de Atapuerca (Figure 1). The cave has a shelter morphology due to the collapse of its dome and is about 15 m deep. The Edelweiss speleological group carried out the first archaeological work in the 1970s (Ortega and Martín, 2012; Osaba and Ruiz de Erenchun, 1978), and the activity of clandestine diggers subsequently affected the deposit. Finally, the cave is being systematically excavated since 1999.
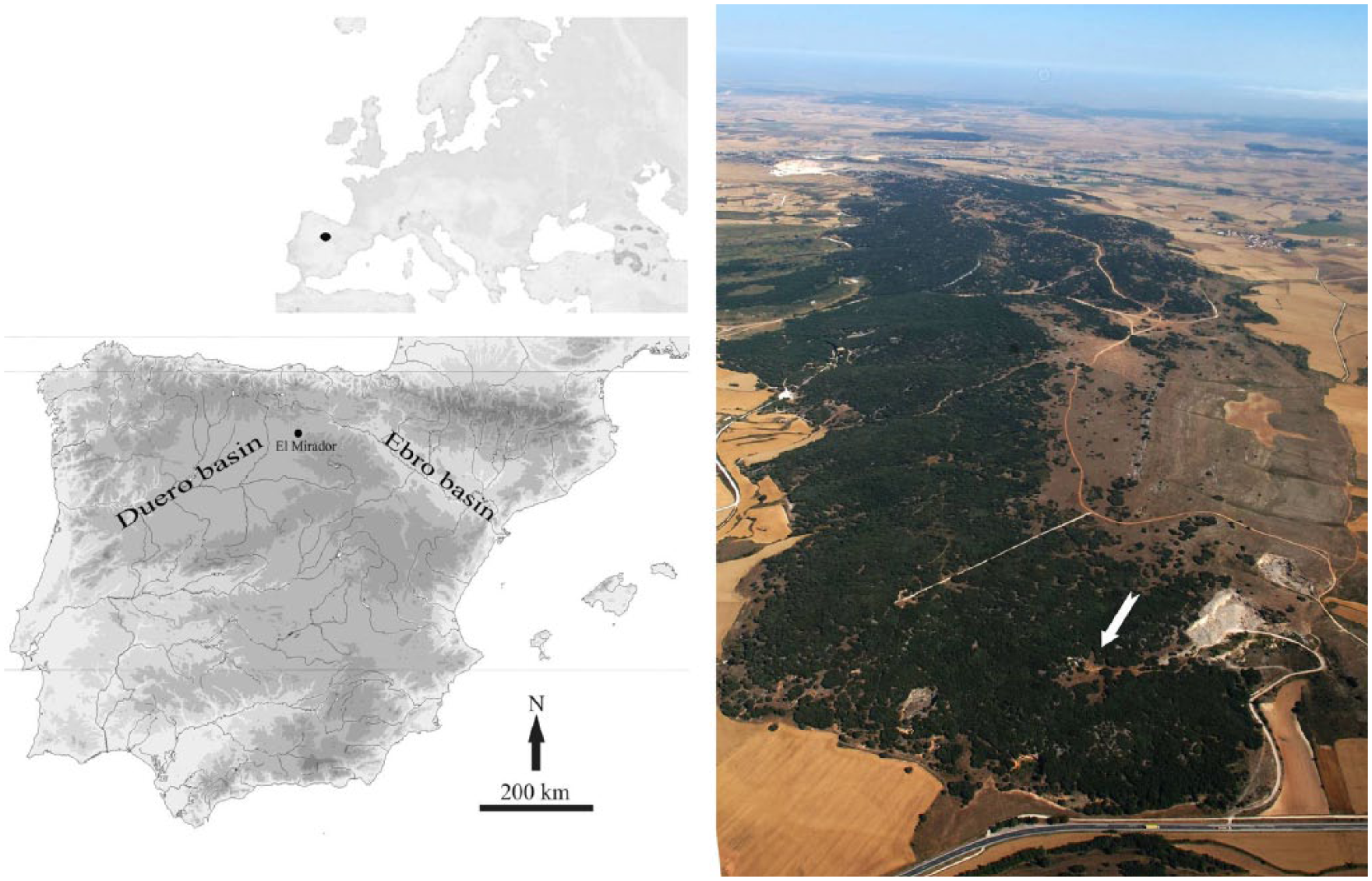
The location of El Mirador Cave and the main associated river basins. The arrow in the picture points to the location of El Mirador Cave with respect to the Sierra de Atapuerca.
The Holocene sequence discovered after the initial survey is 5.5 m thick and includes 24 levels, ranging from the early Neolithic to the Bronze Age (Figure 2). Most of these units exhibit scarce postdepositional modifications and an exceptional degree of preservation (Angelucci et al., 2009). The upper levels, MIR3A and MIR4, are assigned to the middle Bronze Age, level MIR5 is a disruption in the overall sedimentary dynamic when the cave was used as a burial site during the Chalcolithic (Vergès et al., 2016) and from MIR6 to the base of the sequence the deposit is clearly related to a Neolithic horizon (Vergès et al., 2008).

Stratigraphic sequence of the test trench at El Mirador Cave (Sierra de Atapuerca, Burgos, Spain).
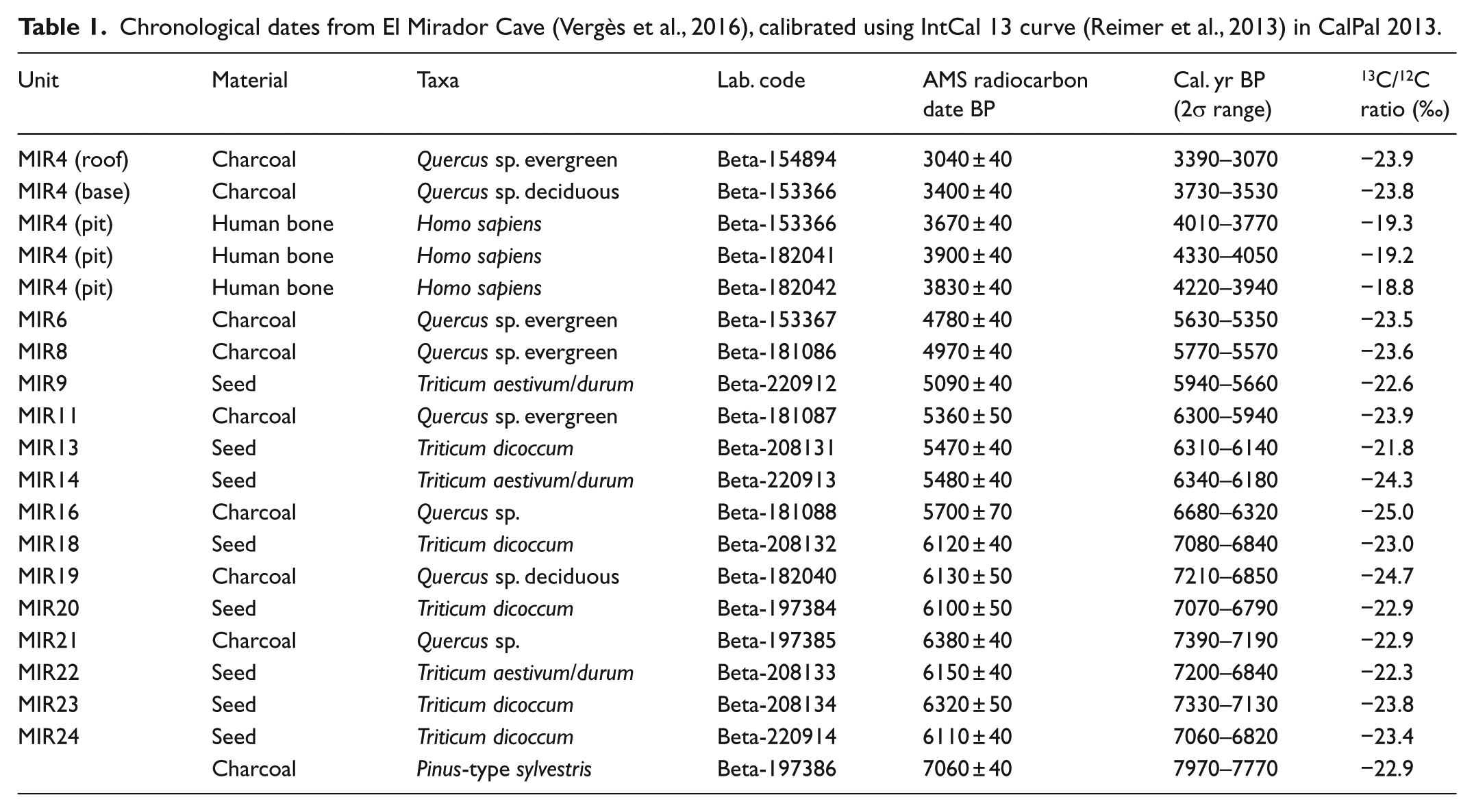
Chronology is based on a set of radiometric dates (Table 1) obtained from charred plant remains.
Chronological dates from El Mirador Cave (Vergès et al., 2016), calibrated using IntCal 13 curve (Reimer et al., 2013) in CalPal 2013.
The archaeological record, typical of a settlement with agricultural and livestock practices, is quite complete and includes one Bronze Age human burial (3580 ± 40 BP) with evidence of cannibalism, pottery, macro- and micromammal remains and lithic artefacts (Bañuls et al., 2013; Cáceres et al., 2007; Gómez-Sánchez et al., 2014; López-García et al., 2008; Lozano et al., 2015; Martín et al., 2009, 2014, 2016a, 2016b; Moral del Hoyo et al., 2003–2004; Oms et al., 2014; Saladié et al., 2015; Vergès et al., 2008). However, the archaeobotanical record is the most complete, although it has only been partially studied (Allué and Euba, 2008; Cabanes et al., 2009; Euba et al., 2016; Expósito and Burjachs, 2016; Rodríguez-Cruz and Buxó, 2008; Rodríguez-Cruz et al., 2016).
Materials and methods
The palynological samples were collected simultaneously to the archaeological work. The sampling was adapted to the complex stratigraphy of the deposit and to the overlap of the sedimentary facies. The samples were collected on a succession of sediment layers proposed by the archaeologist. We focused our palynological analysis on the south section of the archaeological survey, which contains virtually all the documented levels.
The samples were treated with HCl, NaOH and HF, and flotation was performed with Thoulet’s solution (Burjachs et al., 2003; Goeury and De Beaulieu, 1979). Fossil pollen, spores and NPPs were identified using published keys (Jarzen and Elsik, 1986; Miola, 2012; Moore et al., 1991; Reille, 1992, 1995; Van Geel, 1978, 1986) and a modern pollen reference collection.
A set of 50 samples were analysed from 21 levels (MIR4–MIR24). We counted a minimum of 100 grains of pollen as indicated by the methodology established for the palynological analysis of archaeological deposits (Bryant and Holloway, 1983, 1996; Janssen, 1981; McAndrews and King, 1976; Sánchez-Goñi, 1993) with an Olympus Cx41 microscope at 600 magnifications.
Data processing and graphic representation were performed with the help of the software Tilia (Grimm, 1991–2011). The results are expressed in relative frequencies separately for pollen and NPPs. Pollen percentages of trees, shrubs and herbs were calculated based on a pollen sum, excluding Asteraceae and Cerealia-type because they have a specific pollination system and/or are favoured by anthropic activity. For the palynological concentration (PC), we used the volumetric method proposed by Loublier (1978). Local pollen assemblage zones were determined with a cluster analysis using CONISS (Grimm, 1987).
In order to simplify the results, we created a synthetic diagram in which the pollen and NPP taxa appear grouped according to their ecological affinities (Table 2).
Allocation of pollen and non-pollen palynomorphs to groups with ecological affinity (Behre, 1986; Carrión et al., 2010; Galop and López-Sáez, 2002; Hillbrand et al., 2012; López-Sáez et al., 1998, 2000; Ruiz-Zapata et al., 2006).

Results
The characteristics of the deposit, as well as their taphonomic history, have conditioned the unequal preservation of pollen and spores. Still, despite the irregularities, we could guess the plant environment and we have identified the most essential features in terms of its evolution.
A cluster analysis made it possible to divide the sequence into two main palynological zones (A and B) and two subzones (A1 and A2/B1 and B2; Figures 3–5 and Table 3).
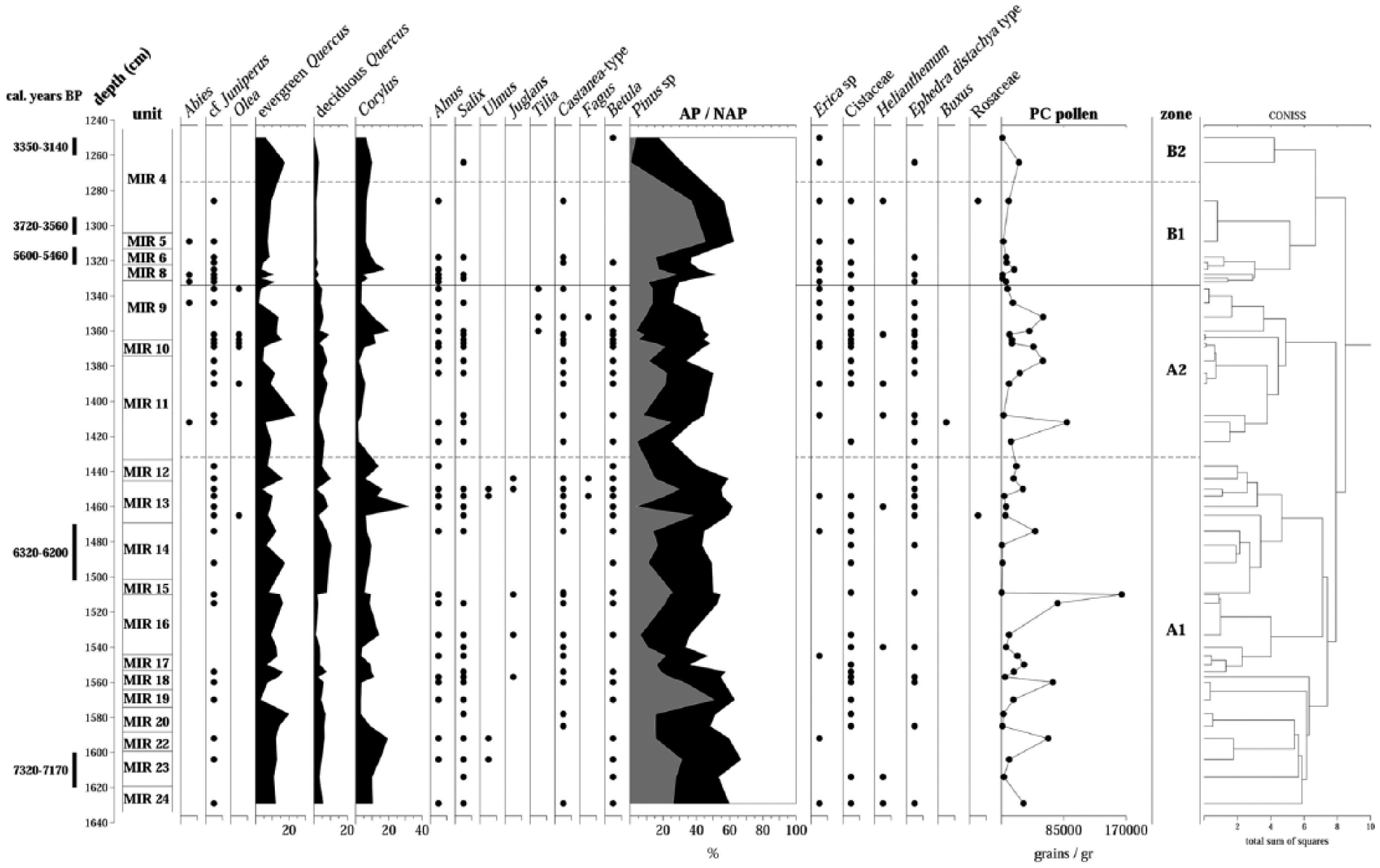
Diagram of the relative frequencies of the tree and shrub taxa of El Mirador Cave. Points indicate frequencies below 2%.

Diagram of the relative frequencies of the herbaceous taxa of El Mirador Cave. Points indicate frequencies below 2%.

Diagram of the relative frequencies of non-pollen palynomorphs (NPP) of El Mirador Cave. Points indicate frequencies below 2%.
Description of palynological zones established by CONISS (Grimm, 1987).

Discussion
The vegetation and environmental evolution
The pollen spectra identified suggest that the landscape may have been made up of a mixed forest of Pinus, deciduous and evergreen Quercus and other deciduous trees like Corylus, Fagus and Juglans. We identified a riverside community comprising Salix, Tilia, Ulmus, Alnus and hydro-hygrophilous taxa like Cyperaceae, Typha–Sparganium and the aquatic Myriophyllum. The presence of these hydro-hygrophilous taxa implies the existence of a water source near the cave on the past. At least Castanea and deciduous Quercus (e.g. Q. pyrenaica) may have lived on the acidophilous Arlanzón terraces, and some species of mountain pines and Betula must be regarded as having been transported by wind over a long distance from the higher elevations of the neighbouring regional mountain ranges, like the northwest foothills of the Iberian System (Carrión et al., 2010; Franco-Múgica et al., 2001; López-Sáez et al., 2010). It must be emphasised that the continuous presence of Castanea wich matches with its expansion from the northeast to the northwest of the Iberian Peninsula, during the middle Holocene (García-Antón et al., 1990). With respect to Abies, it should have travelled from the central and western Pyrenees, where its expansion has been documented after 6000 BP (Alba-Sánchez et al., 2009). Meanwhile, Fagus could be linked with the documented communities of the Iberian System (López-Merino et al., 2008) or the Cantabrian massifs and Pyrenees (Martínez-Atienza and Morla-Juaristi, 1992) that have acted as refuge areas until the beech expansion in the middle Holocene (Magri, 2008).
Heliophilous taxa such as Cistus or Helianthemum would have lived in the open areas within the forest. In the meadows, we would have found Poaceae, Asteraceae, Chenopodiaceae, Plantago spp., Apiaceae, Artemisia, Urticaceae, Ranunculaceae, Asphodelus and other herbs. Some of these deforested areas may have been devoted to the cultivation of cereal crops, but not continuously.
Therefore, the relatively diverse plant taxonomical composition, and its development throughout the profile, suggests the existence of a temperate climate context with some fluctuations in temperature and humidity, marked by oscillation in the representation of thermophilous and mesophilous trees and the riparian community.
The most temperate and wet phase was detected at the bottom of the sequence, where we found the highest values of arboreal taxa. This stage is consistent with the climatic stability and the thermo-mesophilous taxa expansion described in the climatic optimum (ca. 10,000 and 6500 cal. yr BP) for the Mediterranean area (Giralt et al., 1999; Jiménez-Moreno and Anderson, 2012), the north (Peñalba-Garmendia et al., 1997; Sánchez-Goñi and Hannon, 1999) and the southwest Iberian Peninsula (Fletcher et al., 2007; Reed et al., 2001).
The most important trend in the evolution of the vegetation is the progressive decrease in the arboreal pollen percentage throughout the Neolithic occupation. We observed a gradual opening up of the environment, which became much more intense as we approached the Bronze Age levels. This involved the appearance of heliophilous trees and shrubs, the development of grassland areas and the increase in vegetation associated with crops (Figure 4).
In fact, like at El Mirador, the arboreal pollen values from palynological studies conducted at archaeological sites in the north and centre of the Iberian Peninsula are generally low (Atienza-Ballano, 1995; Bueno et al., 2002; López-García and López-Sáez, 2000; López-Sáez and Burjachs, 2002; López-Sáez and López-Merino, 2007; Ruiz-Zapata et al., 2003, 2006). This is often directly related to human disturbance since AP rates are much lower than those evidenced in natural sequences for this geographical area and chronological period (López-García, 1978).
With regard to the relatively high and constant representation of pine through the sequence, it should be noted that pine is also the most represented arboreal taxa in natural and archaeological deposits available for this geographical area and chronological period (Carrión et al., 2010; Franco-Múgica et al., 2001; García-Antón et al., 2011; López-Sáez et al., 2010). In fact, the presence of pine is considered to be linked to regional vegetation and to be directly related to the surrounding mountain ranges of the Northern Plateau because of its great capacity for pollen dispersal. Pine is present from the late Pleistocene in many of the sequences of the central and coastal Iberian Peninsula (Carrión, 2002; Franco-Múgica et al., 2001, 2005; López-Sáez et al., 2014; Peñalba-Garmendia, 1994) and would have prevailed during the first half of the Holocene, with some oscillations.
This constant presence of pine during the Holocene on the Northern Plateau of the Iberian Peninsula is evident in sequences such as Cueva del Moro (López-García and López-Sáez, 1994), Gallocanta (Burjachs et al., 1996), Espinosa de Cerrato (Franco-Múgica et al., 1996) and Lillo (Franco-Múgica et al., 1997), which show the great prevalence of pine forests in the Mediterranean region. In fact, Pinus dominated, together with Quercus, Betula and Juniperus, from the Last Glacial Maximum until the influence of the establishment of deciduous trees in Atlantic areas during the Holocene (Carrión et al., 2010; Peñalba-Garmendia, 1994).
In any case, although the elevation and location of the deposits imply variations in the representation of pine and other taxa, the general trend is a fluctuation in the role of Pinus and Quercus based on small climatic variations or human pressure (Carrión et al., 2010). In fact, it seems that Pinus pinaster could be dominant in some of the regional mountain ranges and also could have been part of the mesophytic forest formations with Quercus (Carrión et al., 2000, 2007).
In the case of El Mirador, palynological data reinforce the idea of the coexistence of both types of forests during the Holocene and both regional and local presence of Pinus. The anthracological data are consistent with that coexistence throughout the sequence, but with a dominance of Quercus compared with the occasional presence of Pinus sylvestris type. The palynological data, however, illustrate a close similarity in the dynamic of the Pinus and Quercus curves. The lines only occasionally diverge, which may correspond to an aridification episode, as evidenced in Zone B1 (Figures 3 and 4). In fact, this phase related to MIR5 and the base of MIR4 level lacks evidence of anthropogenic impact on vegetation. This fact also occurs throughout the Peninsula, the central area of the Iberian Peninsula, and could match with the 4.2 ka cal. BP event (Magny, 2004; Magny et al., 2009).
So, overall, we can confirm the existence of a phase of increased aridity in the upper part of the sequence, which would coincide with the aridity episodes described for the interval 5300–3540 cal. yr BP, closely linked to the expansion of pine, sometimes in conjunction with evergreen Quercus (Carrión et al., 2010). In El Mirador, the beginning of this phase coincides with level MIR5 (chronologically between 3560 and 5600 cal. yr BP), and the rise of Pinus starts becoming irrefutable in MIR4. It is precisely in this level, which belongs to the Bronze Age phase, where we observed the maximum representation of pine and an increase in the Mediterranean trends and aridity characteristic of the beginning of the middle Holocene in this geographic area and that would have influenced the decline or disappearance of Ulmus, Juglans, Tilia and Fagus populations (Carrión et al., 2010; Franco-Múgica et al., 2001, 2005; García-Antón et al., 2011; Gil-García et al., 2002; López-Sáez et al., 2003; Muñoz-Sobrino et al., 2004). The incidence of aridity and the intensification of human pressure are also reflected in the anthracological record, which shows a change in dominance from deciduous Quercus to evergreen. This change is noticeable from MIR14 (6320–6200 cal. yr BP) and is definitely consolidated at the top of the sequence (Allué and Euba, 2008).
It is difficult to document the influence of these xerophytic phases in the settlements in the vicinity of the Sierra de Atapuerca, although some evidence of the occurrence of some of these aridity episodes has been found. This is the case of the 8.2 ka BP event, verified from the pollen record of Cañada de la Cruz, which caused the migration of most of the hunter–gatherer communities from the central Ebro basin (González-Sampériz et al., 2009). In our case, it was not possible to assess the impact of natural forest fires so closely related to processes of aridity. The genesis of the deposit does not allow us to differentiate between charcoals from fires and those generated by anthropic systematic burning. At any rate, the incidence of decreased rainfall is parallel to the progressive increase in indicators of anthropic pressure. Thus, the oscillation of the representation of arboreal pollen at El Mirador is clearly accompanied by the appearance of vegetation typical of a deforestation phase, the development of anthropozoogenous grasslands and the proliferation of nitrophilous or ruderal plants (Barbier et al., 2001; Behre, 1981; López-Sáez et al., 1998).
Human activities and anthropogenic modifications
Although one of the objectives of this work has been the landscape reconstruction in the vicinity of the Sierra de Atapuerca during the Neolithic and Bronze Age, the most important trend identified in the sequence analysed is the role of vegetation related to a clear anthropised environment.
It should be noted that in the data from bogs and palaeolakes of the north and central areas of the Peninsula, such as Comeya (Jiménez-Sánchez et al., 2003), Monte Areo and Alto de la Espina (López-Merino, 2009), Puerto de los Tornos (Peñalba-Garmendia, 1994), Turbera de la Piedra (Muñoz-Sobrino et al., 1996), Quintanar de la Sierra (Peñalba-Garmendia, 1989), Hoyos de Iregua (Gil-García et al., 2002), Lago del Carrizal (Franco-Múgica et al., 2005), Espinosa de Cerrato (Franco-Múgica et al., 2001) and Cañada de la Cruz (González-Sampériz et al., 2009), the first pollinical evidence of the start of anthropisation appears only in the early Neolithic.
Nevertheless, the first clear signs of human impact are not found in the Iberian System until the late Bronze Age (Ruiz-Zapata et al., 2002), when the higher elevation areas of the Northern Plateau began being colonised. This late sign of human communities’ influence on the landscape is surely related to the elevation at which these deposits are found. It seems obvious that the first more or less permanent settlements that exerted pressure on the environment were located in the lowlands and valleys, so a pollen analysis that only provides a regional view of the landscape would not detect these signs (Carrión et al., 2010). This increase in the phenomenon of anthropic activity has been widely observed in most palynological sequences available for the north and the central areas of the Iberian Peninsula at sites such as Can Roqueta (Burjachs and Expósito, 2007), Lago Enol (López-Merino, 2009), Rascafría (Ruiz-Zapata et al., 2006), El Portalón (Ruiz-Zapata et al., 2003) and Covatilla (Atienza-Ballano, 1995).
The palynological analyses of archaeological deposits such as Azutan (Bueno et al., 2002), Barruecos (López-Sáez et al., 2007), Cueva de Chaves (López-Sáez et al., 2006), La Cueva de los Gitanos (Ontañón et al., 2013) and Dehesa Río Fortes (López-Sáez, 2002) reflect an evident human modification of the vegetation on the north and centre of the Iberian Peninsula at the end of the Atlantic period (Carrión et al., 2010; Delibes de Castro et al., 2015; López-Sáez et al., 2014) as in the rest of European continent (Zvelebil, 1989).
The first archaeobotanical evidence of agricultural practices on the Iberian Peninsula was documented on the east coast, at Mas d’Is, Valencia, dating to ca. 6600 ± 50 BP (Bernabeu-Aubán et al., 2003). The available data from inner Iberia have documented the agriculture practices in some sites in the Meseta as La Lámpara and La Revilla (Rojo-Guerra et al., 2006) and Cueva de la Vaquera (Estremera, 2003), among others, around 6400 cal. yr BP. In the northeast of the Iberian Peninsula, Can Sadurní (Blasco et al., 2005) has reported evidences of farming activities at 6450 ± 50 BP, and in the rockshelter of Chaves, located in the Ebro valley, cereal pollen have been identified between 6230 and 6120 BP (López-Sáez and López-García, 2000).
In this way, Neolithisation process implies significant changes in the relation between environment and human communities. These transformations spread throughout the Iberian Peninsula at various stages of inland colonisation through the channels of major rivers of the Mediterranean coastal area (Bernabeu-Aubán et al., 2014). This process starts in the first half of the eighth millennium completing the cultural and social change that had begun in the southeast of the European continent in the ninth millennium (Marinova et al., 2012; Revelles, 2016).
The palynological sequence of El Mirador reflects reliably the introduction and settlement of this new production system characterised, in the Iberian Peninsula, for the great diversity of the crops including a large variability of cereals and vegetables, from ca. 7970 to 7770 cal. yr BP (Zapata et al., 2004).
In El Mirador, the agricultural practices have already been demonstrated through the data provided by the archaeobotanical analysis (Rodríguez-Cruz and Buxó, 2008). The presence of seeds of Triticum aestivum/durum and Triticum dicoccum was identified from MIR6 to MIR24, as well as, exceptionally, Hordeum vulgare var. nudum. Agricultural by-products have also been identified in 10 levels, although with a greater presence at the base of the sequence (from MIR19 to MIR24).
The palynological analysis revealed that the Cerealia-type curve is continuous throughout the entire sequence with some fluctuations, perhaps related to the nature of the activities carried out by the communities who occupied the cave. Indeed, there seems to be an alternation between livestock and farming practices that also involves changes in the values of the nitrophilous vegetation component, in accordance with the intensity of the occupations. Still, it is necessary to thoroughly assess the significance of the oscillation of the different indicators. For example, at the top of the sequence, the decline in nitrophilous plants, coinciding with a rise in cultivation, does not appear to be exclusively related to a lesser degree of anthropisation, but rather to a change in land use strategies. According to López-Sáez et al. (2000), areas previously occupied by nitrophilous and ruderal plants were afterwards used for crops, and this could have been the case at El Mirador.
Nevertheless, in some cases, it has identified a very intensive development of livestock. Thus, the progressive decrease in Poaceae observed in some moments of the sequence was probably related to the high incidence of farming practices in an arid or less humid environment, which may have been decisive in the decrease in grassland areas near the cave at the beginning of the Bronze Age. So the increase in percentages of pastoral pressure indicators that match with the minimal representation of Poaceae could be related to the depletion of these types of herbaceous plants by overgrazing (Burjachs et al., 1997) and could involve the existence of a more sedentary mobility pattern in these communities.
In any case, the Cerealia percentage exceeds 3% in 27 of the samples analysed, which may be related to the existence of crop fields near the cave (Diot, 1992; Richard, 1995). Values close to 10%, identified in some of the levels of the sequence (Figure 4), have been attributed, however, to the existence of some degree of anthropogenic manipulation (Diot, 1992) or to an indirect contribution of anthropogenic origin (Bower, 1998; Diot, 1992; Robinson and Hubbard, 1977).
The impact on the environment intensified in the Bronze Age levels is evidenced by an increase in anthropogenic indicators. The exceptional location of the settlement probably made it possible to establish cultivation areas in the vicinity and to use large areas of grassland. In addition, the strategic location of El Mirador cave, connecting two large valleys, suggests that this site that would have been frequented by pastoralist groups of the Northern Plateau moving to the area of Cantabrian influence or the Iberian System to reach high-mountain pastures.
The only point in which there is no evidence of these activities at El Mirador is in MIR5 which, as mentioned above, represents a unique and uncommon phenomenon. Thus, palynologically, this level yielded evidence of plants and NPPs opposite to those associated with anthropogenic contexts, as well as the existence of a particularly dry climate context.
Regarding the contribution of NPPs and considering the functionality of the infilling and the fact that it was subjected to periodic burning for the purpose of cleaning and deworming, one would expect greater taxonomic variability and a greater percentage of NPPs, and especially coprophilous and carbonicolous ones. The low presence of these kinds of microorganisms leads us to consider that the systematic fires may have inhibited the activity of these saprophytic organisms.
The oscillation in the representation of taxa associated with nitrogenated contexts (Figure 6) indicates that the changes in the trophic conditions can be explained by the increase or decrease in phosphate- and nitrate-rich waste related to pastoral pressure (Van Geel et al., 1981). Naturally, the systematic burning of infill drastically reduces the eutrophic content of the soil, which is regenerated through the proliferation of species like Gloeotrichia, a pioneer alga that colonises nitrogen-poor substrates (Van Geel, 1986; Van Geel et al., 1984, 1989), or through a new occupation of the cave.
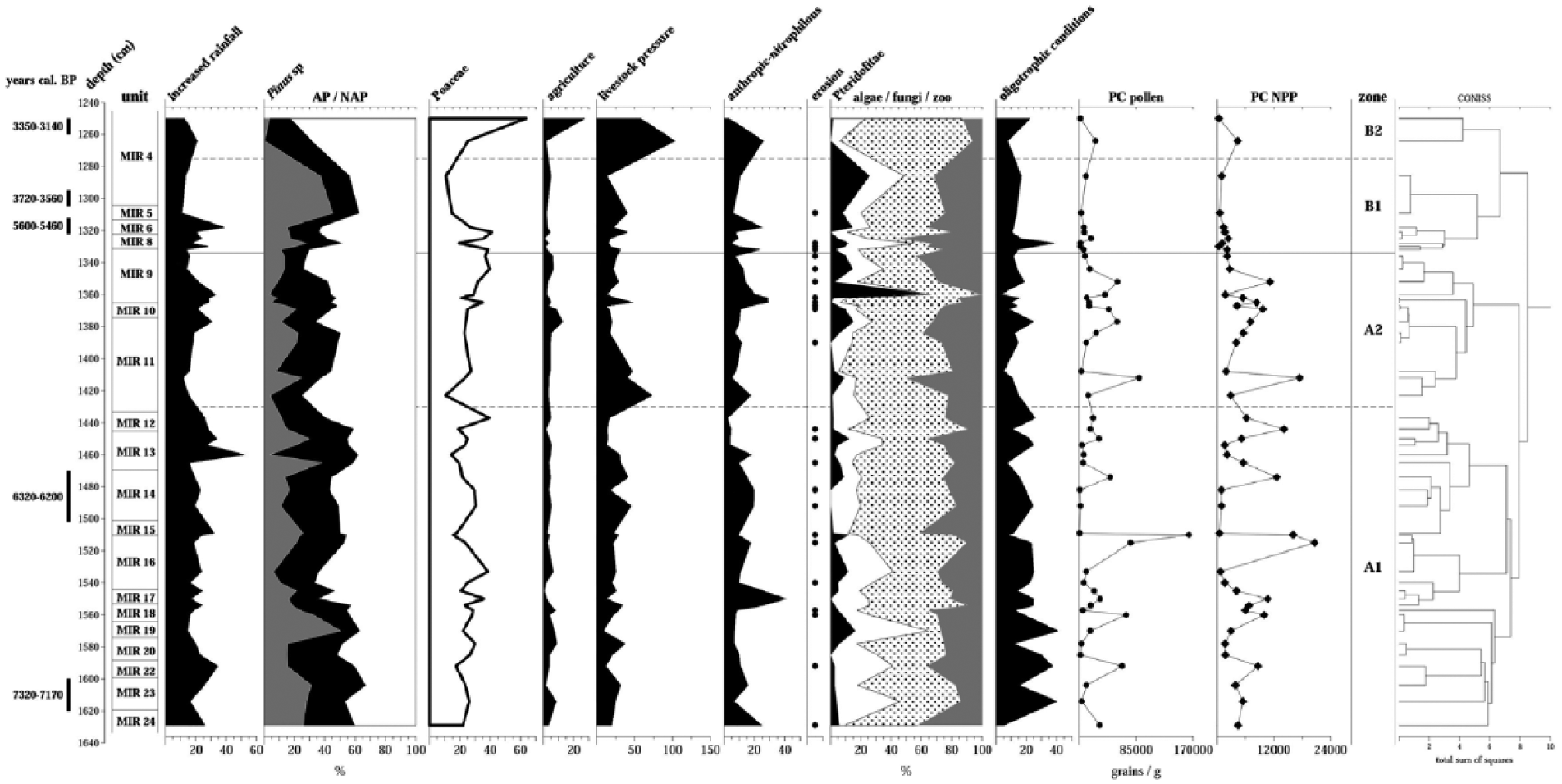
Synthetic palynological diagram of El Mirador Cave. The groups are formed according to the taxa allocation shown in Table 2.
Conclusion
This first palynological analysis of the complete sequence of El Mirador cave has shown the existence of an open landscape formed by a mixed forest of pines and oaks from the early Neolithic to the Bronze Age. Mixed in with these forest formations were other deciduous trees such as Corylus or Castanea-type, which grew on the acidophilous terraces. The riparian forest was made up of Salix, Tilia and Alnus. Betula, Abies and Pinus were probably distributed among the mountain ranges of the region.
The different plant communities documented and the values of the arboreal pollen (always around 50%) suggest that the landscape had a clear open sub-Mediterranean character with a clear importance of sclerophyllous trees and pine woodlands like is typical of the continental central areas of the Iberian Peninsula in this chronological frame (Burjachs et al., 1997; Carrión et al., 2010; Pérez-Obiol, 2007; Pérez-Obiol et al., 2011).
The decline in forest formations, the occupation of the open areas with heliophilous trees and bushes and the increase in grasslands and scrublands suggest the typical phases of landscape degradation. Thus, this evolution towards a highly anthropised environment implies the progressive increase in nitrophilous and anthropozoogenous taxa associated with areas dedicated to grazing and cultivation. Therefore, as would be expected given the use of the cave as a fold, the community most widely represented is the group of grazing plants.
The data indicate that the human groups visiting El Mirador from the early Neolithic to the Bronze Age carried out livestock rearing and cereal cultivation practices and made use of the available forest resources in the chronological frame that the beginning of this mode of production was documented in the north of the central area of northern Plateau of the Iberian Peninsula.
The exceptional resolution of the palynological data, as well as the excellent chronological adscription of documented levels in El Mirador, makes this palynological record essential for the characterisation of the implementation of the Neolithic economic system in the Sierra de Atapuerca biogeographic crossroad, little explored so far.
Footnotes
Acknowledgements
The authors are deeply grateful to the Atapuerca research team and the fieldwork participants. They also thank Ethel Allué, Carmen Expósito and Isabel Cáceres for their help and comments that have certainly helped improve the manuscript. The authors also acknowledge the comments and suggestions made by the anonymous reviewers.
Funding
The Ministerio de Ciencia e Innovación (N_CGL2009-12703-C03-02/BTE, HUM2007-62915/HIST, HAR2008-01984/HIST) and the Ministerio de Economía y Competitividad (CGL2015-65387-C3-1-P (MINECO/FEDER)) and the Generalitat de Catalunya (2014SGR900, 2014SGR899) supported this research. Funding for the fieldwork came from the Consejería de Cultura y Turismo, the Junta de Castilla y León and the Fundación Atapuerca.