
Abstract
At the biogeographical limit between the boreal and the subarctic domain in northeastern Canada, peatlands are mainly oligotrophic fens characterized by a dominance of aquatic microforms such as pools and wet hollows. These peatlands present similar features as the appa mires in Scandinavia. They show evidence for recent water-table rise indicated by tree mortality, physical degradation of strings, and pool expansion. This study aims to evaluate the timing of pool inception and their impact on the long-term dynamics of these microforms within two patterned oligotrophic fens in the Laforge region, northern Québec. Plant macrofossil and testate amoeba analyses from sediment underneath pools were used along with radiocarbon dating to reconstruct peatland pool dynamics over the Holocene. Our data indicate that wet hollows or shallow pools developed at minimal ages between ~4200 and ~2500 cal. a BP. Pool initiation in the peatlands of the Laforge region corresponds to the climate shift toward cooler and wetter conditions from the onset of the late-Holocene cooling. We suggest that the pool developed as secondary features influenced by short growing seasons, low accumulation rates, and wet conditions, which in turn affected the wettest microforms to shift into permanent pools. The differential response of microforms to shift in surface wetness shows the complexity of processes involved in pool initiation.
Keywords
Introduction
In the Northern Hemisphere, peatlands are widespread ecosystems of the boreal and subarctic landscape and play a key role in the global carbon cycle and feedbacks to climate change (Yu, 2011). In Canada, peatlands cover about 12% of the land area and are particularly abundant in the Hudson Bay and James Bay regions (Tarnocai et al., 2005). In eastern Canada, surface peatland ecology, hydrology, and morphology change along important latitudinal and continental–oceanic climatic gradients (Foster et al., 1988; Garneau et al., 2014; Glaser and Janssens, 1986; Payette and Rochefort, 2001). The ombrotrophic peatlands which dominate the boreal regions are gradually replaced by oligotrophic peatlands in subarctic latitudes with abundance of pools gradually increasing toward the north. In the Laforge region in northern Quebec (54°N; 72–73°W), the patterned fens are characterized by flooded surfaces with high dominance of coalesced pools with narrow strings covering up to 42 % of their surfaces (Cliche Trudeau et al., 2014; White and Payette, 2016). These pools comprise residual hummocks, strings and floating peat carpets, and the inundation and mechanical destruction of their margins suggest that the aquatic compartments are expanding at the expense of terrestrial surfaces (Cliche Trudeau et al., 2013, 2014; Dissanska et al., 2009; White and Payette, 2016).
The present-day surface patterning observed in the fens of the Laforge region was previously attributed to a general rise in water tables that started ~3000 years ago and being particularly active during the ‘Little Ice Age’ (Van Bellen et al., 2013). More recently, Arlen-Pouliot and Payette (2015) suggested that the increase in precipitation since the mid-18th century in boreal and subarctic Quebec was responsible for pool formation in these peatlands. The evolution of these systems toward aquatic ecosystems may have major consequences on their carbon sequestration functions and ecological functioning. The transformation of terrestrial vegetated microforms into wet hollows and pools enhances peat decomposition with significant potential increase in emissions of greenhouse gases such as CO2 and CH4 (Cliche Trudeau et al., 2014; McEnroe et al., 2009; Pelletier et al., 2007; Waddington et al., 1998). It is important to understand the mechanisms involved in the long-term development of these pools to better predict their evolution in the context of ongoing climate change.
Research on Holocene peatland development has expanded over the past decade in boreal and subarctic bogs of eastern Canada (Beaulieu-Audy et al., 2009; Garneau et al., 2014; Lamarre et al., 2012; Loisel and Garneau, 2010; Magnan et al., 2014; Van Bellen et al., 2011a, 2011b, 2013). However, little attention has been given yet to the Holocene dynamics of pools in the patterned fens of the subarctic regions. The origin and development of pools and strings in peatlands have been studied for more than a century in Europe (Auer, 1920; Nlisson, 1899; Osvald, 1923; Von Post and Sernander, 1910), and many theories have been proposed since to explain their initiation (e.g. topographic, biotic, and climatic) (Belyea, 2007; Belyea and Lancaster, 2002). Overall, the processes involved in pool formation and linkages with climate changes remain poorly documented in northern peatlands. Most studies on Holocene development of pools and hollows were conducted in bogs (e.g. Foster and Wright, 1990; Glaser and Janssens, 1986; Hulme, 1986; Karofeld, 1998, 2004), and much less attention was given to the processes controlling their formation in fens (Foster and Fritz, 1987). White and Payette (2016) have analyzed 24 fens with different sizes and pool structures in the James Bay region and suggested that pools formed and expanded mainly due to physical factors such as peatland slope and area, peat thickness, and watershed size. On the contrary, the hypothesis of groundwater control suggested by Comas et al. (2011) to explain pool formation might be considered as an explanatory factor not necessary underneath pools but when integrating fluxes from adjacent mineral aquifer surrounding the ecosystems (Bourgault et al., 2016).
We hypothesized that pools in subarctic peatlands developed as secondary features under wet and cool conditions of the late Holocene (Van Bellen et al., 2013). These conditions were likely associated with short growing seasons, low peat accumulation, and high water tables that affected the wettest microforms to shift into permanent pools. The objectives of our study were to reconstruct the long-term dynamics of pools within oligotrophic fens in the subarctic Quebec region using reconstruction of peat stratigraphy underneath pools. More specifically, we aimed to evaluate (1) the timing of pool inception in these peatlands over the Holocene and (2) the influence of internal processes and regional climate over long-term pool dynamics.
Study area and sites
In the northeastern portion of the La Grande Rivière watershed (54°N; 72–73° W; James Bay region), peatlands cover ~15% of the land area (Tarnocai et al., 2005). They are mainly represented by patterned oligotrophic fens with permanent pools and seasonally flooded hollows, interspersed between strings (White and Payette, 2016). In this region, peatlands developed shortly after the retreat of the Laurentide Ice Sheet from 7500 cal. a BP (Dyke, 2004). The Tyrrell sea waters penetrated 200–250 km inland (Vincent, 1989) where ombrotrophic peatlands (Sphagnum bogs) developed. East of the marine limit, peatlands are predominantly minerotrophic (poor fen; Arlen-Pouliot and Payette, 2015).
Two small patterned fens have been studied in the Laforge region, approximately 440 km east of the James Bay at the boreal/subarctic ecotone in northern Québec. Aéroport (54°6′2″N, 72°30′59″W; 402 m a.s.l.) and Ours peatlands (54°2′56″N, 72°27′25″W; 460 m a.s.l.) are located, respectively, at 16 and 26 km south of the Laforge-1 hydroelectric power station (Figure 1).
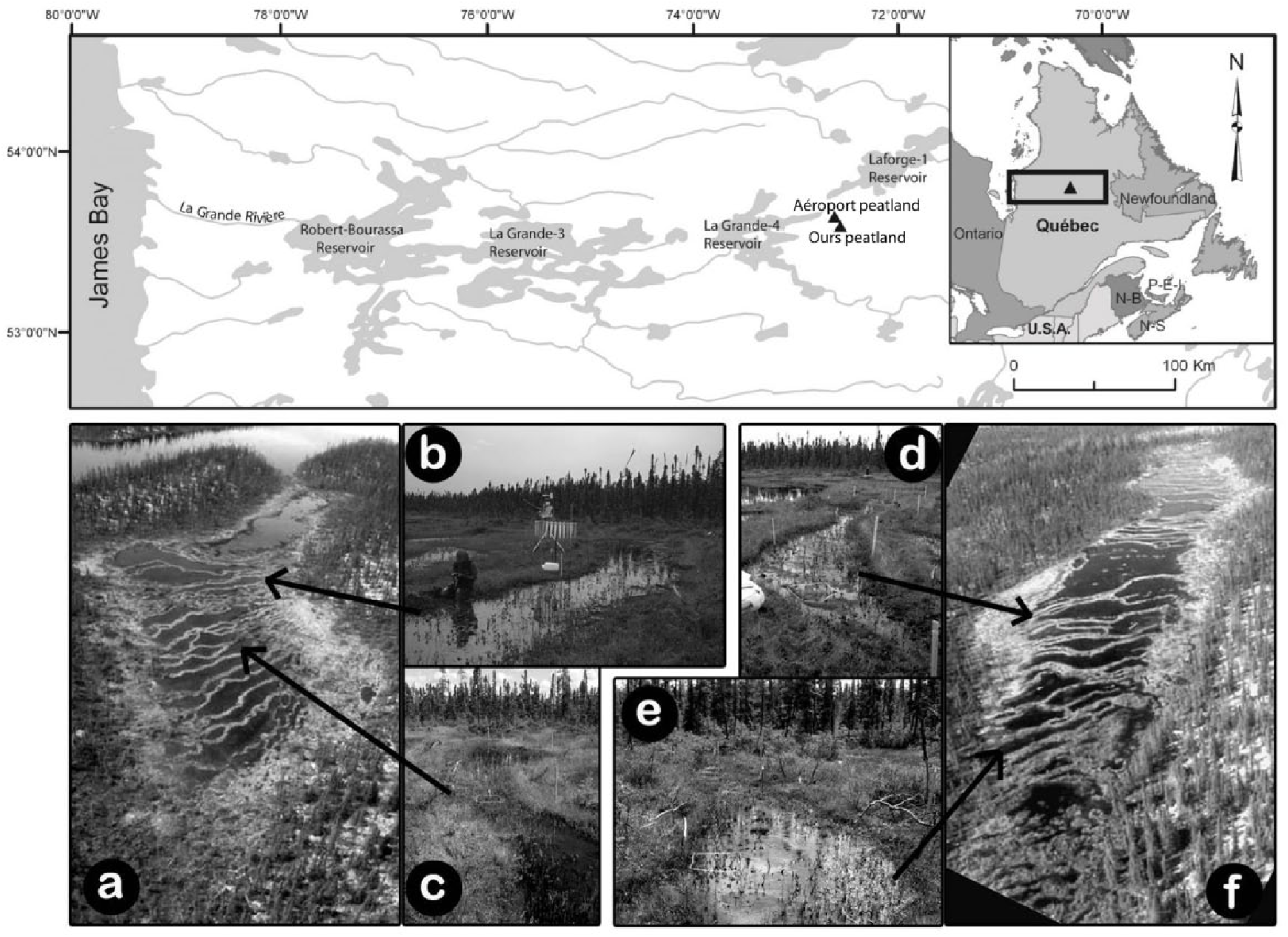
Location of the two studied peatlands in the La Grande Rivière watershed in Québec, northeastern Canada (upper panel). Photographs of the two studied ecosystems: (a–b–c) Aéroport and (e–d–f) Ours.
The studied peatlands are largely treeless and dominated by sedges (Carex exilis, Carex oligosperma, Carex limosa) and mosses (mainly Sphagnum fuscum and Sphagnum cuspidatum with Gymnocolea inflata) accompanied by ericaceous shrubs (e.g. Kalmia polifolia, Rhododendron groenlandicum, Chamaedaphne calyculata, Andromeda glaucophylla, and Vaccinium oxycoccos). They are located in depressions topographically delimited by Quaternary till deposits over the Precambrian Shield. The two studied peatlands have well-developed surface pattern with lengthened, narrow (1–5 m) shallow pools (mean depths of 5–30 cm) and small hollows oriented perpendicularly along the longitudinal axis and the slope. Pools and hollows are separated by narrow strings topped with low hummock–lawn microforms (Figure 1).
Aéroport peatland covers an area of ~0.02 km2 and has a slight slope of ~1% oriented ENE-WSE. A small outlet in the western portion of the peatland basin flows into an adjacent shallow lake. The deepest portions of the basin contain the largest pools (100-m long and 10- to 25-m wide) (Figure 2) and are located on the west section underlain by fine sands and silts likely deposited during a phase of high lake level following ice retreat. The eastern part of the peatland lies over deepened bedrock and sand deposits, probably corresponding to the former shoreline of the adjacent lake. The presence of submerged strings and hummocks indicates the coalescence of small pools.
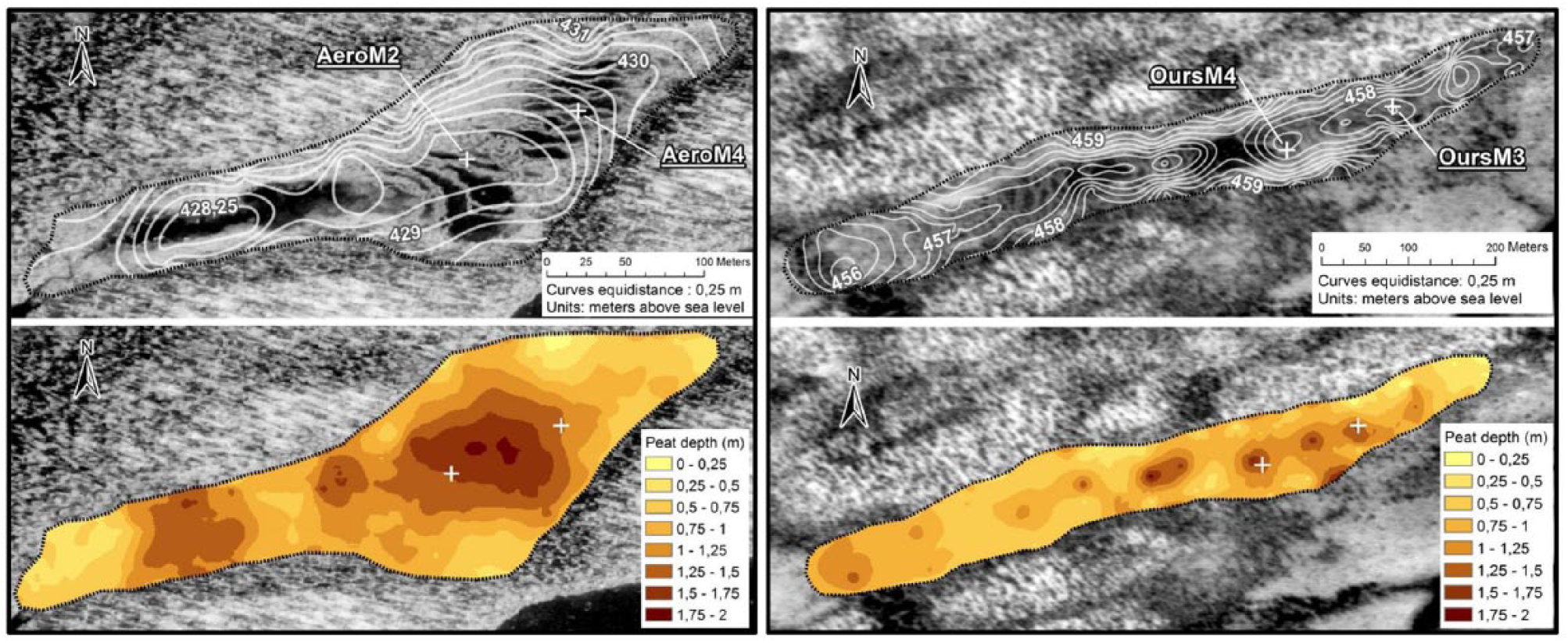
Peatland basin topography shown in elevation in meter above sea level (upper panels) and measurements of peat depths (lower panels). The location of the two coring sites in each peatland (Aéroport on the left and Ours on the right) is indicated by a white cross.
Ours peatland has an area of ~0.03 km2 and developed into a topographic depression oriented ENE-WSW delimited by two parallel fluted hills (Figure 2). The underlying mineral substrate mainly consists of fluvial sands and patches of gravels. The basin morphology is characterized by two sub-basins that induce a bidirectional water flow from the center toward the edges. The largest and deepest pools are found in the central and thickest portion of the peatland. They enclose hummock-type islands and string-like submerged crests likely formed by the coalescence of pools.
The regional climate is continental subarctic with a mean annual temperature of −4.3°C ranging from −24.1°C in January to 12.8°C in July and mean annual precipitation of 738 mm (NLWIS data; Huchinson et al., 2009). The region belongs to the spruce woodland bioclimatic region, characterized by acidic soils and dominated by black spruce (Picea mariana (Mill) BSP) and lichens of the genus Cladinia and Cladonia.
Materials and methods
Characterization of peatland surface and mineral basin topography
The surface microtopography of Aéroport and Ours peatlands has been evaluated using a differential global positioning system (DGPS) Trimble RTK 5800 with a high horizontal and vertical spatial resolution of 1–2 cm and 3–5 cm, respectively. The peat thickness and peatland basin topography (relative altitude m a.s.l.) were determined over the entire peatlands with a Hiller probe (Fries and Hafsten, 1965) following gridded 40-m intervals at Aéroport peatland (total 62 depth measurements along 11 transects) and 50-m intervals at Ours peatland (total 76 depth measurements along 14 transects). Each peatland basin topography measurement was plotted using the ‘Topo to Raster’ interpolation method by ArcGIS 9.2 software (ESRI, Redlands, CA, US) to create digital elevation models (DEM; Figure 2).
Field sampling
Following the microtopographic surveys in each site, pools were selected for coring during the next winter season. Peat cores were sampled in the center of two representative pools from Aéroport (AeroM4 and AeroM2) and Ours peatlands (OursM3 and OursM4). AeroM2 was collected in the center of the peatland where peat is nearly the thickest and AeroM4 in the upstream sector, the two sites being located 40 m apart (Figure 2). In Ours peatland, the two sampled pools (OursM4 and OursM3) are located 85 m apart within the same and deepest sub-basin in the eastern sector (Figure 2).
The four studied pools are shallow with maximum water depths ranging from 14 to 35 cm. AeroM2 and AeroM4 pools are 30 and 35 m long and 1–5 m and 2–4 m wide, respectively (Figure 2). OursM3 and OursM4 pools are 15 and 20 m long and 1–3 m and 0.4–1 m wide, respectively (Figure 2). A CRREL mechanical ice-corer (75 mm diameter; Brockett and Lawson, 1985) was used to extract the upper frozen peat bottom sediment, and the underneath unfrozen and unconsolidated sediment was sampled with a Kajak-Brinkhurst (KB) gravity corer (Brinkhurst et al., 1969; Glew, 1989; 50 cm × 5 cm diameter). A Livingstone piston-corer (Livingstone, 1955; 100 cm × 5 cm diameter) was used for deeper consolidated organic sediments. Frozen sediments were wrapped in plastic foil and stored in a freezer at −5°C. The top unconsolidated sediments were cut into 1-cm-thick slices in the field using the KB extractor (Glew, 1988). Livingstone peat cores were wrapped in PVC tubes and aluminum foil and stored at 4°C until analyses in laboratory.
Sediment characterization
The frozen cores were cut into 1-cm-thick slices with a band saw in the frozen chamber of the former National Glaciology Laboratory of the Geological Survey of Canada (GSC, Ottawa, Canada). The Livingstone peat cores were cut into 1-cm-thick slices. Sub-samples (1 cm3) were collected at 1 cm intervals in each core for analyses of bulk density with overnight drying at 105°C and combustion at 600°C for 3 h to determine ash-free bulk densities (AFBD) and mineral contents (Chambers et al., 2010). Sediment stratigraphy from the different cores was reconstructed based on the bulk density data.
Plant macrofossil analysis
Peat sub-samples of 5 cm3 were collected at 4 cm intervals for macrofossil analysis and prepared following the protocol of Mauquoy et al. (2010). Samples were heated for 10 min in a KOH-5% solution and gently washed with distilled water through a 125-µm sieve. Plant remains were counted (e.g. seeds, leaves, and needles) or estimated as relative abundance (%) using a gridded Petri dish (e.g. Sphagnum, brown mosses, and Cyperaceae). Plant remains were identified using the reference collection of the Laboratory of Continental Paleoecology (GEOTOP, UQAM). Nomenclature follows Marie-Victorin (1997) for vascular plants, Crum and Anderson (1981) for brown mosses, and Bastien and Garneau (1997) for Sphagnum. Macrofossil remains were grouped into five peat types (Sphagnum, brown mosses, Cyperaceae, ligneous, and unidentified organic matter (UOM)) and represented as cumulated percentages curves. In some levels, an important fraction of the highly humified sediment was lost through sieving process (up to 80%). The volume of the fine peat fraction (FPF) <125 µm lost by sieving was estimated in percentage.
A detrended correspondence analysis (DCA) was performed on the whole plant macrofossil dataset (four cores combined) using PCORD (McCune and Mefford, 2006) in order to explore the associations between taxa in the fossil assemblages. Only the taxa expressed in relative abundance (%) have been included in the analyses, and taxa with low occurrence have been down-weighted. The DCA axis 1 scores were used along with other proxies (e.g. AFBD and FPF) to establish visually the diagram zonation.
Testate amoeba analyses
Peat samples (2 cm3) were taken at 4 cm intervals along AeroM2 and OursM4 cores for testate amoeba analyses on the same levels used for macrofossil analyses. However, many levels could not be analyzed due to poor test preservation and very low concentrations. For AeroM2 core, only the upper 40 cm contained sufficient test concentrations. Sub-samples were processed following the protocol of Hendon and Charman (1997). They were spiked with a Lycopodium tablet, boiled in distilled water for 10 min and washed through 355 and 15 µm sieves. The material retained between the two sieves was stained with neutral red and mounted on glass slides. We aimed to count a minimum of 100 tests under the microscope following the taxonomy of Charman et al. (2000). A weighted average tolerance (WA-tol) down-weighted model with classical deshrinking was used to reconstruct past water table depths (WTDs) with the C2 software version 1.7.2 (Juggins, 2007) using the transfer function of Lamarre et al. (2013). Only samples with >75 tests were used in the reconstructions as lower counts tend to provide unreliable values (Payne and Mitchell, 2009). The rotifer Habrotrocha angusticollis was excluded from the WTD reconstructions. The two testate amoebae diagrams are provided as Supplementary Material (Figures S1 and S2, available online).
Radiocarbon dating
In all, 28 samples of terrestrial plant remains (mainly Sphagnum stems) were collected along the main stratigraphic transitions and submitted to the Keck Carbon Cycle laboratory (University of California, Irvine, US) for AMS radiocarbon dating (Table 1). Radiocarbon dates were calibrated to calendar years before present (BP, that is, AD1950) using the CALIB 6.0.1 software using the INTCal09 calibration curve (Reimer et al., 2009; Stuiver et al., 2010). Ages were rounded to the nearest decade.
AMS radiocarbon dates and calibrated ages from Aéroport and Ours pool cores.
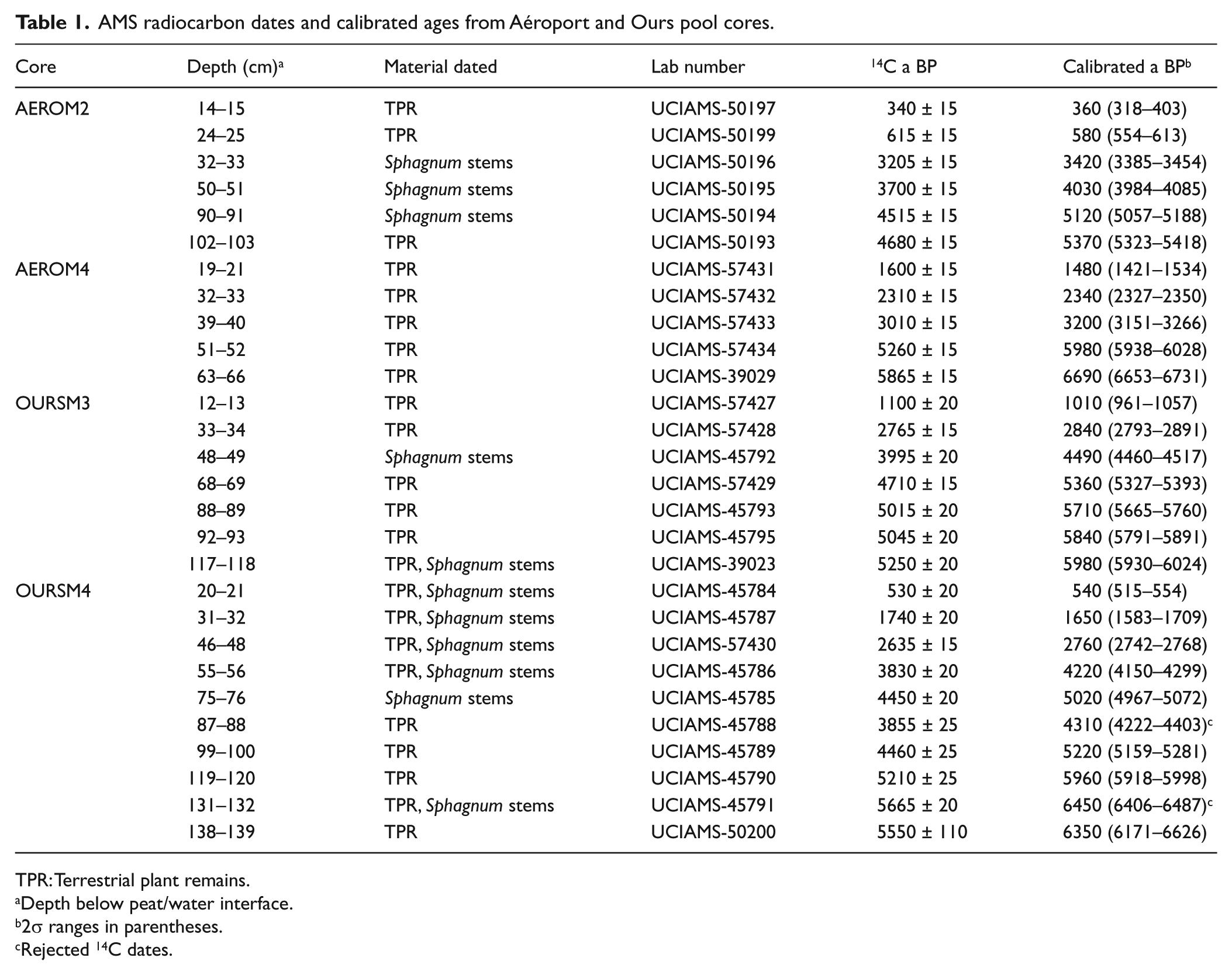
TPR: Terrestrial plant remains.
Depth below peat/water interface.
2σ ranges in parentheses.
Rejected 14C dates.
Age–depth models were developed using linear interpolation between each calibrated 14C date (Figure 3). We used linear interpolation because dating was realized at the main stratigraphic transitions where changes in sedimentation rates can often be reconstructed. Two 14C dates were rejected in the modeling of OursM4 because they were inconsistent with the adjacent 14C dates (Table 1) as either too young (OursM4 87–88 cm) or too old (OursM4 131–132 cm) likely due to a contamination of pool sediment by younger plant material or redeposition of older plant remains. However, we found no other evidence of major sediment reworking in the peat stratigraphy.
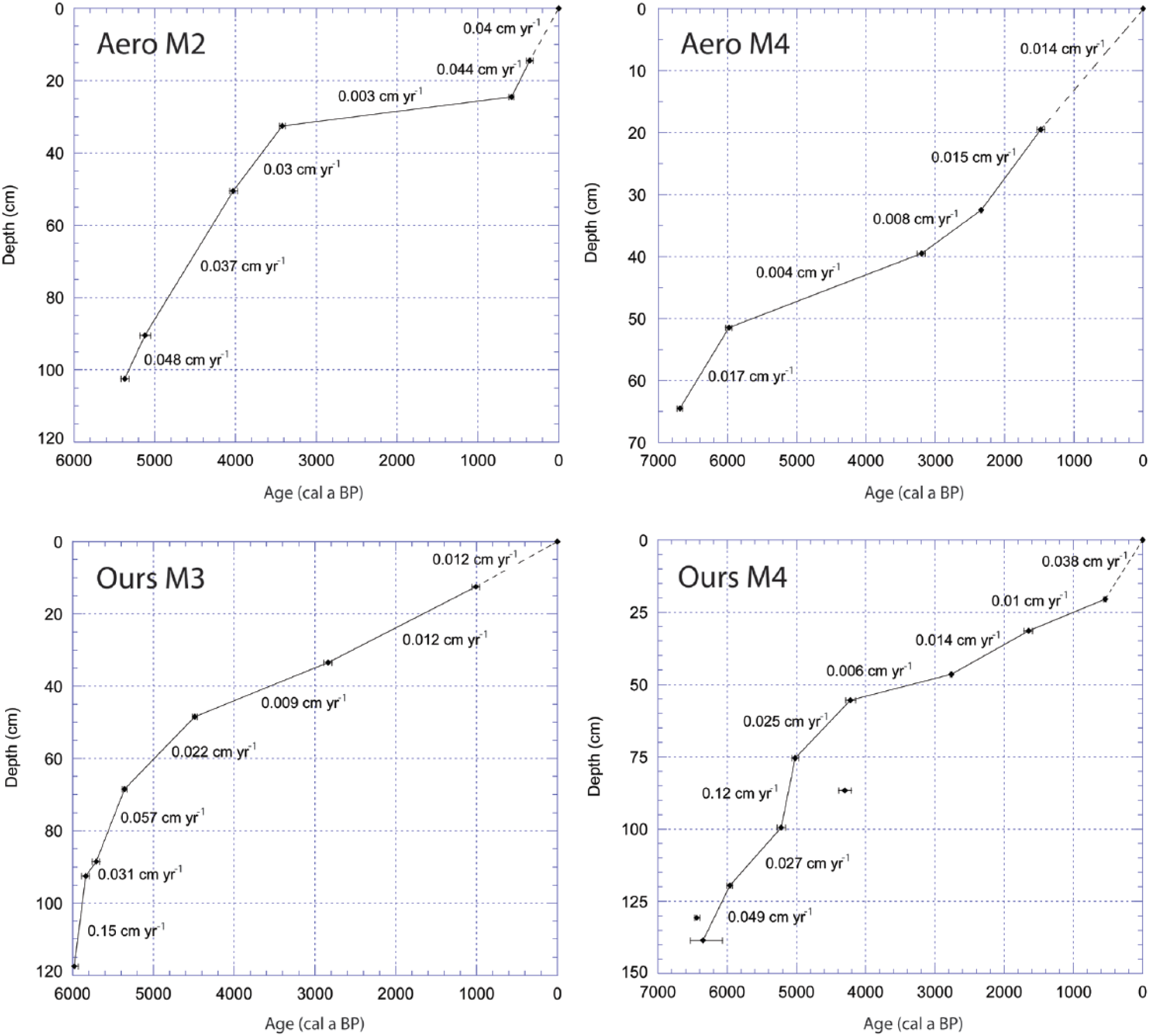
Age–depth models of the four studied pool cores.
Results
DCA ordination
The DCA ordination shows coherent associations between the plant macrofossil types along the main axis of variability (axis 1) which corresponds with a surface wetness gradient in fens (Figure 4). DCA Axis 1 and Axis 2 explain, respectively, 21.5 % and 14.6 % of the variability in the macrofossil assemblages. The bryophytes typical of submerged conditions in wet hollows or shallow pools (e.g. Scorpidium scorpioides, Drepanocladus spp., Sphagnum cuspidatum) are found on the right-hand side (high values). The plants typical of drier terrestrial microforms such as lawns and hummocks (e.g. Sphagnum sect. Acutifolia, Sphagnum papillosum, and Picea mariana) are found on the left-hand side (lower values). The position of UOM toward the center is attributed to opposite conditions: very dry decomposed peat or very wet hollow peat.
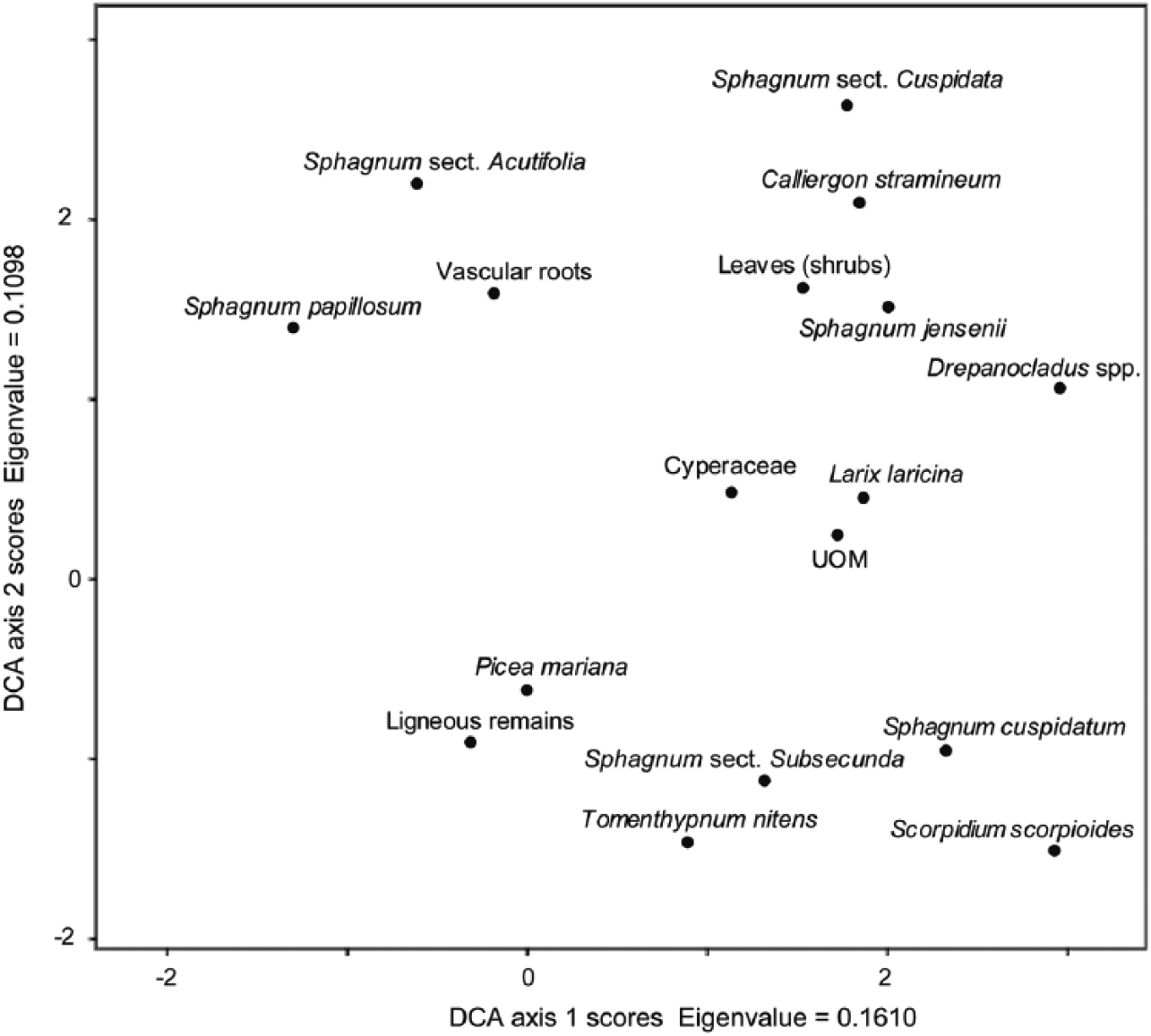
DCA ordination of the plant macrofossil data (four cores from two sites combined).
Macrofossil data
At AeroM2, peat started to accumulate in a semi-aquatic environment (Characeae, Daphnia, Trichoptera, Menyanthes trifoliata) colonized by the brown mosses Drepanocladus spp. and Scorpidium scorpioides which indicate rich minerotrophic conditions (Zone 1; 5370–4900 cal. a BP; Table 2 and Figure 5). Sphagnum sect. Cuspidata and Chamaedaphne calyculata established between 4900 and 4100 cal. a BP, suggesting a shift toward poor minerotrophic conditions (Zone 2). Picea mariana and Sphagnum sect. Acutifolia were present between 4100 and 3400 cal. a BP (Zone 3) in a section of highly decomposed peat (high UOM and FPF) as the conditions were likely drier and ombrotrophic. A return to semi-aquatic conditions after 3400 cal. a BP (Zone 4) is suggested by the presence of Sphagnum sect. Cuspidata along with few remains of Daphnia and Nymphaeaceae. The very low PAR (0.003 cm a−1) between the levels dated at 3400 and 600 cal. a BP may be related to high peat decomposition within wetter hollows. The high abundance of conifer needles after 4000 cal. a BP suggests tree mortality due to possible flooding. The testate amoeba assemblages mainly dominated by Amphitrema wrightianum and Difflugia globulosa (Figure S1, available online) indicate relatively stable water tables near or above the surface (−5 to 0 cm) from 3800 cal. a BP.
Details of macrofossil diagram zones.

PAR: peat accumulation rate; WTD: water table depth; FPF: fine peat fraction; AFBD: ash-free bulk densities.

Macrofossil diagrams of AeroM2 and AeroM4 cores. The basal sediment with high mineral content >50% is shaded in light gray. lvs: leaves; sds: seeds; PAR: peat accumulation rate; WTD: water table depth; FPF: fine peat fraction.
At AeroM4, Picea mariana (needles) was present on the site prior to the establishment of minerotrophic conditions confirmed by Scorpidium scorpioides and Drepanocladus spp. (Zone 1; 6700–5980 cal. a BP; Figure 5 and Table 2). PAR declined drastically around 5980 cal. a BP from 0.017 to 0.004 cm a−1 as brown mosses were replaced by Cyperaceae and ligneous remains (Zone 2). After 2340 cal. a BP, Sphagnum jensenii (sect. Cuspidata) and Drepanocladus spp. were present along with few aquatic proxies (Menyanthes trifoliata, Daphnia), suggesting that saturated conditions initiated at a maximal age of ~2340 cal. a BP and have persisted until today (Zone 3). Conifers and ericaceous shrubs (Andromeda glaucophylla, Chamaedaphne calyculata) were present nearby on drier microforms but gradually declined likely due to flooding.
At OursM3, initial rich minerotrophic conditions from 6690 cal. a BP (Nymphaeaceae, Cyperaceae, Myrica gale and Scorpidium scorpioides) shifted around 5980 cal. a BP to poorer fen conditions with Sphagnum and Drepanocladus mosses in the macrofossil assemblages (Zone 1; Figure 6 and Table 2). PARs were high and decomposition was minimal before 5600 cal. a BP. Bryophytes were replaced by Cyperaceae and Chamaedaphne calyculata between 5600 and 5100 cal. a BP (Zone 2). Conifers and ericaceous shrubs became more abundant after 5100 cal. a BP in the macrofossil assemblages. The presence of Scheuchzeria palustris and Sphagnum (sect Subsecunda and Cuspidata) suggests that a wet hollow developed between 5100 and 4200 cal. a BP (Zone 3). Sphagnum declined and peat decomposition increased after 4200 cal. a BP (high UOM and FPF) as the vegetation was composed of ligneous species and Cyperaceae (Zone 4). The very low PAR (0.009 cm a−1), high level of peat decomposition, and the presence of Trichoptera remains are indicative of shallow pool conditions after 4200 cal. a BP.

Macrofossil diagrams of OursM3 and OursM4 cores. lvs: leaves; sds: seeds; PAR: peat accumulation rate; WTD: water table depth; FPF: fine peat fraction. The basal sediment with high mineral content >50% is shaded in light gray.
At OursM4, the basal peat (Zone 1; 6350–6000 cal. a BP) contains many black spruce needles and some remains of Sphagnum sect. Acutifolia (Figure 6 and Table 2). The testate amoebae (Figure S2, available online) indicate dry surface conditions (dominance of Difflugia pulex, Trigonopyxis arcula, Nebela militaris, and Heleopera sylvativa) during this early stage likely corresponding to paludification over a forested site. From 6000 to 5100 cal. a BP (Zone 2), Cyperaceae became dominant accompanied by Drepanocladus spp., Menyanthes trifoliata, and Daphnia ephippia, suggesting the onset of wet conditions on the site. From 6000 cal. a BP onward, the inferred WTDs are above or near the surface (dominance of Amphitrema wrightianum) until present-day. Conifer needles are abundant along with Chamaedaphne calyculata between 5100 and 4200 cal. a BP (Zone 3), but the presence of Menyanthes trifoliata suggests the persistence of a wet hollow. Although water table was high between 6000 and 4200 cal. a BP (Zones 2 and 3), PAR remained relatively high varying from 0.025 to 0.12 cm a−1, suggesting a stable height between peat surface and WTD. After 4200 cal. a BP, the very low PAR (0.006 cm a−1) and high AFBD (Zone 4) combined with the presence of Daphnia ephippia suggest pool initiation with some inputs from nearby hummocks or strings (e.g. Cenococcum sclerotia and Picea needles). Surface conditions likely became too wet for the persistence of bryophytes and Larix laricina that declined.
Discussion
Initial context of peatland inception
The two studied peatlands developed relatively shortly after the retreat of the ice sheet from the region around 7500 cal. a BP (Dyke, 2004). Aéroport peatland basin was gradually isolated from the adjacent lake by isostatic uplift and lake level drawdown during the warmer mid-Holocene period (Payette and Filion, 1993; Viau and Gajewski, 2009). Peat accumulation started ~1300 years earlier at AeroM4 than at AeroM2 (6690 vs 5370 cal. a BP), which could be explained by its higher elevation in the mineral basin that was the first to emerge (Figure 2). At Ours peatland, land emergence was associated with reduced meltwater inputs into the fluvial plain while the glacier was receding. Peat accumulation started earlier at OursM4 (6400 cal. a BP) than OursM3 (6000 cal. a BP), which was drained slightly later due to its lower position in the basin (Figure 2).
Patterns and timing of wet hollow/pool development
In the two studied peatlands, pools developed as secondary features from vegetated wet hollows as previously documented in boreal peatlands (Belyea and Lancaster, 2002; Foster and Fritz, 1987; Foster et al., 1983, 1988). Our data suggest that wet hollow/pool developed significantly earlier in AeroM2 (3420 cal. a BP) than AeroM4 (2340 cal. a BP). However, there are larger uncertainties in AeroM4 core due to poor peat preservation between the levels dated at 5980 and 3200 cal. a BP so that pool may have developed earlier at this site (Figure 7). In both Ours sites, wet microforms developed rapidly after peat inception from 6000 cal. a BP and persisted until they both transformed into shallow pools at a maximum age of ~4200 cal. a BP.

Synthesis of wet hollow/shallow pool formation from Aéroport and Ours peatlands and the pollen-based reconstruction of annual precipitation anomaly for northern Québec (Viau and Gajewski, 2009).
Autogenic and climatic influence over pool/wet hollow formation
The development of wet hollow/shallow pools in the studied peatlands seems to have been primarily driven by site-specific microform feedbacks to water table rise (Swindles et al., 2012). The difference in the timing of pool formation at each site can be explained by divergent vegetation succession pathways and rates of vertical peat accumulation in microforms. Some initial microforms were gradually transformed due to wetter conditions that enhanced physical decay of vegetation at the peat surface. High water levels caused vegetation drowning within wetter hollows, whereas hummock plants were less affected (Arlen-Pouliot and Payette, 2015). At AeroM2, the transition from rich to poor fen conditions followed by drier bog conditions represents a typical hydrosere which may have been primarily driven by vertical peat accumulation (Hughes and Dumayne-Peaty, 2002). The three other studied sites have never reached ombrotrophic conditions before they transformed into aquatic ecosystems.
The formation of shallow pools in the studied peatlands from ~4200 cal. a BP coincides with a shift from drier to wetter climate conditions in Northern Québec (Figure 7; Viau and Gajewski, 2009) and is consistant with regional lake level rises recorded ~3500 cal. a BP (Miousse et al., 2003; Payette and Filion, 1993). The fens of the Laforge region may have been less resilient to past changes in atmospheric moisture balance due to their biogeographic location at the boreal–subarctic ecotone. In this region, the imbalance between high precipitation and low evapotranspiration, combined with low peat productivity associated with short growing season and low temperatures, may explain the general water table rise during the late-Holocene period (Garneau et al., 2014; Van Bellen et al., 2013). Moreover, the topographic context in which they developed may have influenced groundwater fluxes from the adjacent mineral aquifers (Bourgault et al., 2016) when considering the size of the watersheds (White and Payette, 2016)
Our data show a decrease in peat accumulation rates from ~4000 cal. a BP in lawn microforms located nearby on the same peatlands (Van Bellen et al., 2013). This slowdown in peat accumulation at the onset of the late-Holocene cooling may also have influenced an apparent rise in water tables and the flooding of low-lying terrestrial areas such as hollows and lawns. Once these wet hollow/pools have formed, they have persisted until today despite subsequent climate changes documented in Northern Québec such as the ‘Medieval Climate Anomaly’ (MCA) around 1100–800 cal. a BP and the ‘Little Ice Age’ (LIA) after 500 cal. a BP (Viau and Gajewski, 2009). This supports the idea that pools can be resilient and persistent microforms over millennia (Baird et al., 2016; Belyea and Clymo, 2001).
The combination of our results with those of Van Bellen et al. (2013), Bourgault et al. (2016) and White and Payette (2016) suggests that the flooding was not synchronous over the entire peatland during the middle to late Holocene and were influenced by a combination of climatic, hydrogeologic, and topographic factors. Our study shows that pool and wet hollow formation started from ~4200 cal. a BP in the central and deepest portion of the peatlands when wetter climate conditions were recorded in Northern Québec (Viau and Gajewski, 2009). Arlen-Pouliot and Payette (2015) suggested that pool formation and expansion in this region have been particularly active at the end of the ‘LIA’ and that this process remains active nowadays at the edge of these peatlands due to increased precipitation since the mid-18th century.
Reconstructing long-term pool dynamics using sediment underneath pools
The dating of pool inception with our methodological approach can only be interpreted as a maximal age because the upper peat column may have been partially altered through decomposition by an overlying pool (Foster and Fritz, 1987), and hence, the interpretation of long-term pool dynamics using sediment underneath pools must be done with caution. Hollows are usually characterized by very low peat accumulation rates due to a combination of active decomposition (Karofeld et al., 2008; Rydin and Jeglum, 2006) and low plant productivity (Belyea and Clymo, 2001; Seppälä and Koutaniemi, 1985). Nevertheless, the macrofossil records from pool sediment need to be interpreted with caution due to potential exogenous organic inputs transported by surface waters or derived from pool margins (Belyea and Lancaster, 2002; Foster et al., 1983; Foster and King, 1984). In our study, the periods of pool formation are also associated with abundance of shrubs and conifers remains which were likely derived from nearby terrestrial microforms as pools were expanding. Moreover, the abundance of conifer needles during the periods of pool inception (Figures 5 and 6) may reflect massive tree mortality due to the flooding. This interpretation is consistent with Arlen-Pouliot and Payette (2015) that suggested a massive deforestation of the patterned fens between 3000 and 1000 cal. a BP due to water table rise. Despite potential taphonomic issues, the coherent associations of plant taxa in the macrofossil assemblages and the consistency of the 14C chronology (Table 1) suggest that the pool cores were not considerably affected by mixing or redeposition processes.
In our study, it was difficult to clearly distinguish former wet hollows from shallow pools based on the plant macrofossil analyses. This is mainly because most aquatic proxies such as Menyanthes trifoliata have a relatively wide tolerance to submersion and can grow in both microforms. The Nymphaeaceae (e.g. Nuphar variegata) were the only plants clearly indicative of aquatic (pool) conditions but were rarely found in fossil samples. The testate amoeba analyses allowed a better understanding of past ecological and hydrological dynamics but did not allow to differentiate clearly wet hollows from pools, which might be due to the higher imprecision of WTD reconstruction in fens (Payne, 2011).
Conclusion
Our study showed that the development of wet hollows and pools in the patterned fens of the Laforge region in Northern Québec is a process that started at a maximal age of ~4200 cal. a BP. Pool formation seems to have been primarily driven by internal factors, but the transition toward the cold and wet late-Holocene period in northern Québec seems to have been an important triggering factor. At the onset of the late-Holocene cooling, a slowdown in vertical peat accumulation may have initiated a gradual rise in water table and the transformation of wet hollows into shallow pools in the central sections of the peatlands. This study highlighted the challenge of paleoecological reconstructions from pool sediments due to the complexity of peat accumulation and taphonomic processes in these aquatic microforms. However, our study suggests that sediment underneath pools can provide valuable records for understanding long-term pool dynamics. Such data are essential to evaluate the future ecological and hydrological dynamics of these ecosystems in the context of the ongoing climate change at northern latitudes.
Footnotes
Acknowledgements
Special thanks to Hans Asnong for his great implication in the project. We are also grateful to Valéry Hamel, Alexandre Lamarre, Caroline L’Heureux, Noémie Cliche Trudeau, Claire Lacroix, Simon Tardif, and Guillaume Saint-Jacques for laboratory and field assistance. We thank Dr Jocelyne Bourgeois for the use of the GSC frozen chamber-laboratory (National Glaciology Laboratory, Geological Survey of Canada, Ottawa) and Alayn Larouche for macrofossil identification advices. Thanks to Dr GT Swindles, Dr RJ Payne, and an anonymous reviewer for the constructive comments on the manuscript.
Funding
This work was financially supported by project NSERC-CRDPJ364636-08 (NSERC-Hydro-Quebec; PI: Professor Serge Payette), FRQNT (#117462-2006), and Ouranos Consortium.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.