
Abstract
Humans are increasingly viewed as active agents of environmental and land-cover change in the moist Neotropics. While the scale and extent of pre-Columbian anthropic impacts are actively debated, the effects of post-Contact patterns of land use are rarely examined over the long term, defined here as centennial timescales. This article examines a putative area of historical low human impact located in the western Guiana Shield, the upper Cuao River, using an exploratory agent-based modelling approach. Based on an extensive ethnographic literature on the Piaroa, who have inhabited the region for at least four centuries, the model investigates the legacy effects of ethnographic patterns of land use in the interval between European Contact and the present. Model outcomes indicate that the potential range of anthropic changes to the environment of the study area is significantly greater in scale than previously assumed. Interpretative discrepancies between present vegetation conditions and the model are likely the product of sparse palaeoecological and archaeological research in the upper Cuao. More broadly, the results imply that small-scale agriculture and agroforestry can lead to extensive and persistent structural changes to ecosystems in relatively short timescales. The experiment bolsters existing cautions against assuming the ‘natural’ baseline of Neotropic forests based on present appearance. As a form of middle-range theory, the model demonstrates how computational approaches can promote closer integrations between ecological, archaeological, and ethnohistorical data, as well as frame the expectations of future research.
Introduction
The impact and legacy of indigenous land-use practices continues to be a contentious area of research in the Neotropics (McMichael et al., 2012; Piperno et al., 2015, 2017; Stahl, 2015; Ter Steege et al., 2013; Watling et al., 2015, 2017). Current debates in archaeology and historical ecology have moved on from dichotomous ‘cultural parkland’ versus ‘pristine forest’ scenarios (Denevan, 1992b; Heckenberger et al., 2003; Meggers, 2003), and focus now on defining the scale and extent of environmental modifications effected by the human inhabitants of Amazonia (Clement et al., 2015; Mayle and Iriarte, 2014). Critical readings of the ethnohistorical record frequently inform discussion on these topics, either to contrast with archaeological data or to serve as a point of departure for useful comparisons between observed and archaeologically inferred behaviours (Stahl, 2015; Whitehead, 1998). Crucially, the ‘wilderness’ described in early European records of the Neotropics is now recognized to be, in many cases, an artefact of a post-Contact transition to an anthropogenic, yet largely human-free, environment that bears little resemblance to the pre-Columbian period (Denevan, 2016; cf. McMichael et al., 2012). Although colonization had disastrous consequences, which left measurable signals in the palaeoenvironmental and archaeological records (Bush et al., 2008; Faust et al., 2006), total demographic collapse and transition to historical patterns was not the only possible outcome (Ramenofsky, 1987; Thornton, 1987). The following examines the impact of historical indigenous land use in the interval between Contact and the present, in order to refine hypotheses and theoretical expectations for further field research
This study identifies the long-term (centennial) consequences of historical indigenous land use as an area in need of scholarly attention, particularly where ethnohistorical, palaeoecological or archaeological data are sparse. Short-term research on swidden agriculture (defined here as sub-generational) has for decades formed a programmatic part of Tropical forest research in ecology, geography and archaeology (Cairns, 2015; Conklin, 1961; Harris, 1971). Within the context of the European colonization of the Americas, suggested as one potential ‘tipping point’ marking the onset of the Anthropocene (Erlandson and Braje, 2013), swidden agriculture emerged as the dominant agrarian subsistence strategy among post-Contact indigenous groups. This occurred under a particular set of internal and external societal pressures, as one strategy of many along a spectrum of possible responses by indigenous groups (Arroyo-Kalin, 2012; Balée, 2013: 185; Mayle and Iriarte, 2014; Neves, 2013). Swidden farming was also likely precocious among the first tropical forest cultivators, certainly making it a part of the complex legacy of human inhabitation in the Neotropics (Kawa, 2016; Kirch, 2005; Oliver, 2001; Piperno and Pearsall, 1999; Roosevelt, 2013). If the (circum-)Amazonian world is to be represented in the Anthropocene debate, it becomes necessary to be able to define the scale of historical indigenous environmental impacts in precise terms.
In parallel to historical ecology, yet largely isolated from it, is the growing adoption of simulation and model-based inferential frameworks in archaeological science (Kohler and Van der Leeuw, 2007; Lake, 2014; cf. Johnson et al., 2005), including studies of coupled human-environmental systems and land-cover/land-use change (LUCC) in the past (Barton, 2014; Barton et al., 2010; Wilkinson et al., 2007). Driven by questions about human-environmental impact, ecologists and geographers increasingly deploy computational approaches for exploring complex interactions between agents and ecosystem processes (Parker et al., 2003; Perry and Millington, 2008). Exploratory agent-based modelling (ABM), using known behaviours as a point of departure, is ideal for testing intuitions or hypothesis about a target system (Davies, 2016: 83). Computationally minded archaeologists have already noted the synergy between experimental simulation on one hand (Godfrey-Smith, 2006; Potochnik, 2012) and middle-range theory in archaeology on the other (Davies, 2016; Premo, 2007). Simulation is perhaps best conceptualized as a ‘virtual laboratory’ (Magliocca and Ellis, 2016) that serves to formalize assumptions about the operation of a system of interest, quantify variability in model outcomes and, ultimately, test these against knowledge claims. This is the approach adopted here.
The study aims to refine current understandings of the scale and temporality of impacts caused by interfluvial slash-and-burn (swidden) agriculture in the moist Neotropics. Clearer insights into the legacy and effects of post-Contact patterns of swidden agriculture are crucial to the agenda of historical ecological research, and Anthropocene studies more broadly. The following synthesizes a significant body of ethnographic work from the Orinoco, focusing on the Piaroa indigenous group (see Overing and Kaplan, 2011; Zent, 1992), to investigate the variation in ecosystem disturbances under swidden farming systems. This research was carried out in the context of a larger project investigating indigenous occupations along the Middle Orinoco (Lozada Mendieta et al., 2016). The model generalizes nearly half a century of anthropological and historical research on Piaroa subsistence strategies; however, it does so usefully and explicitly (Epstein, 2008). I argue that the lack of archaeological and environmental field research in the study area presents a gap in knowledge which ethnographically informed computational models may be particularly suited to tackle.
Background to model: Piaroa interfluvial subsistence
The Piaroa occupy the western edge of the Guiana Shield, an upland interfluvial zone in what is today southern Venezuela. This area has historically received scant attention from archaeologists and palaeoecologists, yet is rich in ethnographic data. The Piaroa are traditionally highly mobile farmers whose subsistence is heavily reliant on manioc cultivated in short-lived plots of cleared and burned forest, which is supplemented with fishing, hunting and agroforestry. It is highly likely that because of disease, conflict, slaving and new technologies following European contact, the ethnographically observed settlement pattern is contingent on recent historical processes (such as the gradual spread of metal tools) rather than being the end state of an uninterrupted cultural evolution (Denevan, 1992a; Huber and Zent, 1995: 48; Zent, 1998). Since the rural expansion of the Venezuelan market economy and healthcare programmes in the 1980s, the Piaroa have increasingly settled permanently along the main channel of the Orinoco and its navigable tributaries to access to these services (Freire, 2007; Freire and Zent, 2007; Mansutti Rodriguez, 1988). Modes of subsistence recorded in the highland interfluvial zones, now considered their ‘traditional’ homeland, differ considerably from modern practices. In comparison to the Orinoco floodplain, game is more dispersed, the rivers support fewer fish and cropping cycles are shortened while fallows are lengthened because of the shallow soils (Huber and Zent, 1995: 50). Still, Zent (1995: 87), citing Gilij (1987), considers the Piaroa to have maintained stable settlement in the uplands for 400 years at a minimum. As contact before the 18th century was sporadic and archaeological data are non-existent in the interfluvial zone, this is an estimate for the duration of occupation (Mansutti Rodriguez, 1988: 9; Overing and Kaplan, 2011: 506; Zent, 1992: 48). Sáliva-speaking ancestors of the Piaroa were present in the upper-Middle Orinoco before this, but the circumstances and duration of pre-Columbian settlement here are unknown at present.
Upland Piaroa settlements in the catchment of the Cuao River, a black water tributary of the Orinoco (Figure 1), are typically organized around a single large house (Piaroa: isode). This valley forms the focus of the present study. Isodes are co-inhabited by an extended family or household group of several extended families, consisting of 16–50 individuals (1–10 nuclear families), although actual settlement population fluctuates somewhat because of high individual mobility. Camping trips, visits and temporary relocation can take place in a given year, with the home isode and its extended family group acting as an anchor (Zent, 1998: 255). Overall population density is very low, with at most 10 isodes per 100 km2 ‘neighbourhood’ (Monod, 1970; Overing and Kaplan, 2011). Isodes can reportedly be occupied for as little as 1 year before relocation, or for 10 years or more (Heckler, 2004: 204; Zent, 1992).

Overview map of upper Cuao, showing historic Piaroa settlements recorded by Zent (1992) and modern Piaroa settlements visible in remote sensed imagery. Map inset: Time series of land-cover change from 2000 to 2014 based on data in Hansen et al. (2013). Bottom right: Histogram of swidden plot area (hectares) based on a survey of these data. The distribution of area is log-normal.
Although fishing, hunting and wild resource gathering feature in the diversified Piaroa subsistence base, significant time and effort is invested in the swidden agricultural system, characterized by a short cropping phase (6 years or less) followed by long fallows of 20 years or more (Zent, 1992: 184). Manioc (Manihot esculenta) and maize (Zea mays) together comprise nearly 80% of a harvest and supply the bulk of Piaroa caloric needs, with other wild and cultivated foods providing mainly supplementary nutrition (Overing and Kaplan, 2011: 514; Zent, 1992, 1998). Swidden plots, always cleared by slashing and burning climax forest, range in size from 0.25 to 6 ha, a figure that varies based on the number of individuals a plot will support. A nuclear family will generally self-sustain with a single swidden plot. A survey of 178 swidden plots in the upper Cuao corroborates this (Figure 1, bottom right), and shows a notable skew towards such plots (<2 ha in area, Perera, 1986; Zent, 1992: 190). When the productivity of a coeval group of ageing fields drops off because of vegetation succession and weed growth, members of an isode will relocate to the site of a former plot and establish a new settlement. Altering the forest cover exposes the thin tropical soils to depletion or erosion, in particular if secondary regrowth is cleared again after an insufficiently long fallow (<20 years) (Fölster, 1995: 70). The viability of subsequent cropping cycles and fallows will be severely curtailed if this occurs. Similarly, maintenance of swidden plots for longer than a few years risks limiting the regeneration potential of subsequent regrowth to bush fallow (Eden, 1974: 48). These pathways are typically avoided if at all possible, since fallowed plots and secondary forest both supply ecosystem services through improved hunting and gathering grounds (Harris, 1971; Zent, 1995). To summarize, Piaroa swidden farming in the upper Cuao is an interplay between meeting subsistence requirements, monitoring existing plots and managing wild resources effectively, while avoiding intensive localized pressure on forest cover and overtaxing a limited pool of labour (Mansutti Rodriguez, 1988; Zent, 1992).
Modelling Piaroa subsistence practices and forest succession in the upper Cuao allows some important aspects of post-Contact land cover and land use to be discussed. The implementation of a small number of conservative and straightforward decision-making routines and parameters is based on reasonable assumptions and derivations from the ethnographic literature. For instance, the model uses a static or declining proxy for population, and swidden plots provide consistent returns over their period of cultivation. The world initializes as a homogeneous primary forest, in which site selection is mostly unconstrained and guided only by economic logic insofar as it impinges on farming. It slightly underestimates starting population density as roughly 1 isode per 118 km2, below that estimated by Monod (1970), and does not consider isode migration or fission–fusion dynamics. These serve to usefully constrain the number of factors considered in the parameter space, and focus the discussion of the output. Rather than precisely mirroring every aspect of Piaroa settlement patterns and subsistence behaviour, the model provides a reasonable approximation, which allows for inferences to be made on long-term human-environmental interaction.
Model overview and design
The above account of Piaroa socioecology is implemented computationally through a coupled agent-based and cellular automata (CA) model based on spatial grid cells in NetLogo 6.0.1 (see Luus et al., 2013; Mena et al., 2011; Moreno et al., 2007; Wilensky, 1999) to create an abstraction of observed behaviour capable of investigating its long-term operation. The model parameters are summarized in Table 1. For the study, individual isodes are treated as agents, with population per isode generalized to the nuclear family level. At the start of a given run of the simulation, the number of families is randomly assigned a value of 1–10, which determines the number of swidden plots an isode will produce after each relocation to meet requirements. Isodes are otherwise identical and do not interact, except for avoiding ‘claiming’ active swidden plots owned by another isode. As isode relocation is directly linked to the agricultural strategy (new house plots are always established in old fields), a single parameter handles both swidden logistical range and mobility magnitude (parameter 2). During a run, isodes monitor returns from surrounding grid cells in range each time step and relocate to establish new swidden plots (conucos) when requirements are not being met by existing fields. Grid cells targeted for swidden clearing always consist of fallow secondary or primary forest. The model assumes that historically observed clearance practices, employing efficient metal tools, are present at the start of a run. Stone axes, for example, in a pre-Contact scenario, would require different model assumption as to the time and energy cost of creating clearings (Denevan, 1992a). The choice of new house gardens when relocating (parameter 3) can be varied to be a random cell out of the pre-existing swidden plots owned by an isode, or optimized to always be the plot with the least regrowth. If, in any case, there is an insufficient quantity of grid cells to meet requirements in logistical range, an isode may enter a stressed state when relocating and lose a family (to death or outmigration). The likelihood of stress events occurring is controlled by the magnitude of the deficit of available land.
Summary of parameters in the Piaroa swidden model. The range of values swept in the simulations reported here are shown in square brackets.

Vegetation succession and forest transition is handled as an integer by the CA model, mapped onto a simplified emic schema of forest regrowth stages (see Zent, 1992: 331, 1995: 91; Table 2), which functions on a per-grid cell basis. Grid cells regenerate from initial clearance to initial secondary regrowth, with an equal likelihood of adding no growth (zero), little growth (1) or significant growth (2) per time step. Each time step in the model approximates a season, with two seasons per year. This introduces a degree of stochasticity as to when isode relocation will take place, as plots may regenerate unevenly. As grid cells of secondary regrowth provide some returns (1/10th of a swidden plot) in the form of mature tree stands and improved hunting, a sufficient quantity of old gardens in logistical range could theoretically offset the gradual losses of some fields becoming unproductive.
Vegetation succession in the Piaroa swidden model. Below the thick line, succession is modelled probabilistically as described in the text. Final stage regrowth is indistinguishable from primary forest after a century of regeneration (Zent, 1995). The model also tracks grid cells that have reverted to this state.

Forest succession (from bush to secondary forest, and from secondary to climax vegetation) is modelled probabilistically (parameter 4). Every time step, grid cells with incipient or mature secondary status (Table 2), counts the number of surrounding primary forest cells. Every grid cell of primary forest in the Moore neighbourhood (up to eight neighbouring cells) increases the likelihood that stage succession will occur in increments of 0.025 per cell. The contribution of each cell is varied as a model parameter (see Table 1), giving a range of values from 0.025 (a single primary forest neighbour at the lowest parameter setting) to 1 (eight neighbours with primary forest at the highest parameter setting). This implementation of forest succession approximates the gradual colonization of fallowed plots by pioneer species and the maturation of economic tree species planted by Piaroa at the start of its life cycle. It also reflects the long secondary regrowth period (on average a century) typical of the nutrient-poor and fragile soils of the study area.
The landscape employed in the simulations is a 609 × 426 cell digital terrain model of the upper Cuao watershed (3568 km2), resampled to half of its original resolution of 90 m to improve processing time. Use of the terrain model (parameter 5) does not affect the behaviour of either isodes or cells, but allows certain results of the parameter sweeps to be visualized more easily. The initial state of the landscape assumes 100% primary forest cover. As noted above, there were undoubtedly legacy effects of settlement in the landscape prior to the Piaroa uptake of swidden agriculture. The present state of knowledge in the study area only allows these to be guessed at, and for present purposes, legacy effects have been left out. Each run of the model lasts for 800 time steps, equivalent to 400 years, or stops when all isodes have disbanded or expired. This timeframe represents a hypothetical maximum for the presence of ethnographic Piaroa subsistence behaviour in the study area: the 17th century to the present. Two sets of runs were carried out separately because of prohibitively long processing times for large numbers of continuously measured runs. The first set measured the end state of the landscape and isodes over 100 repetitions of each parameter combination (4500 runs), while the second set measured model outputs diachronically over just 30 repetitions of each parameter combination (1350 runs).
Results
This study modelled Piaroa land use through the interplay between mobility, population and forest regeneration potential to investigate legacy effects on the landscape of the upper Cuao. Figure 2a summarizes the effects of isode mobility on the composition of primary forest in the study area. This compares the number of grid cells that are never cultivated (old growth) against the total amount of primary forest (old growth + reverted primary forest grid cells) at the end of 800 model time steps, equivalent to four centuries. Grid cells of reverted primary forest are former swidden plots that have succeeded back to climax vegetation after a century or more of fallow, making them functionally indistinguishable from old growth primary forest to both Piaroa and non-Piaroa. The pattern implies that as logistical radius (parameter 2) increases, the total amount of primary forest remains stationary and very close to 98%, while unmodified primary forest sees a steady and proportional decrease. Isodes in very low mobility regimes (logistical radius ≤3) experience frequent stress events because of the local depletion of arable grid cells, with coeval losses of population. Loss reduces subsistence requirements, with the consequence of fewer cultivated grid cells overall, leading to a lower ratio of reverted to old growth grid cells. Probability of regeneration (parameter 3) has a negligible effect on the above outputs.
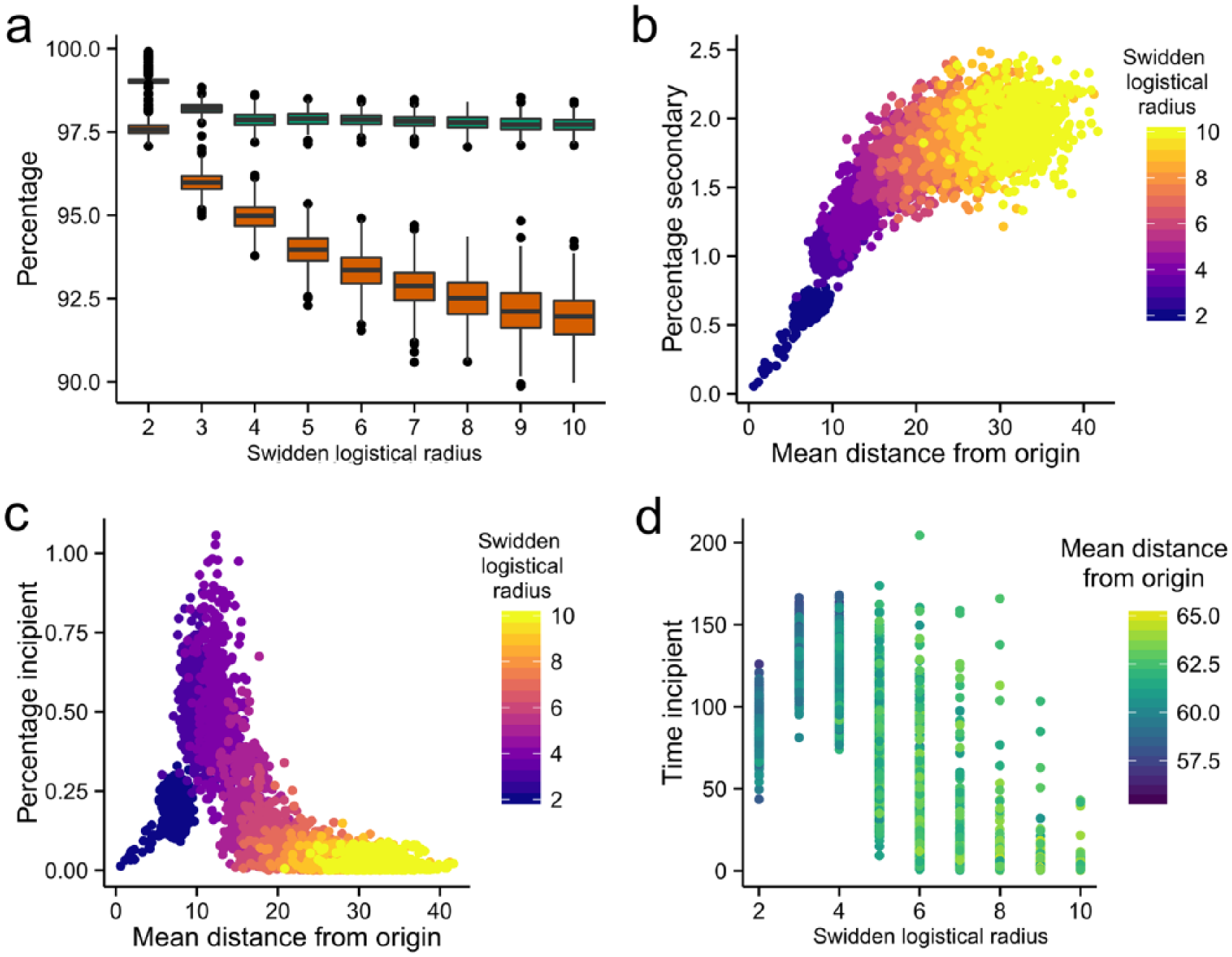
Summary graphs of 4500 model runs. (a) Summary boxplots of old growth primary forest + reverted primary forest (green) in the upper Cuao shown, compared to the percentage of old growth primary forest alone (red). As mobility increases, the proportion of old growth decreases while total quantity of primary forest remains stable. (b) Percentages of forest grid cells with anthropogenic modifications against mean isode distance from starting position. Horizontal axis covaries with parameter governing isode mobility. Above x = 10, the range of the y-value broadens considerably. (c) Grid cells with scrubland regrowth peak noticeably in a short interval, while very low and very high mobility regimes are comparable in impact. (d) The effect of isode mobility magnitude on the average number of time steps spent as incipient forest.
The total area roamed by a co-resident group of families is what Zent (1992) would term their ‘neighbourhood’. Forest succession in the upper Cuao is observably affected by the average distance travelled by isodes from their origin, which is a function of logistical radius (Figure 2b and c). Using this as a proxy for isode neighbourhood, changes in the proportion of grid cells with scrubland (‘incipient’) regrowth and secondary forest both correlate with neighbourhood size. Plotting the proportion of secondary regrowth against the proxy for neighbourhood, the variable describes an S-shaped curve which fans out considerably as mobility increases, reaching a plateau at approximately 20 grid cells from the origin (see Figure 2b). A plausible mechanism behind this pattern is that high-mobility isodes are both (1) unlikely to suffer stress events and thus maintain starting populations (because of a comparatively large logistical radius for establishing new plots) and (2) more likely to move further away from their starting position and disproportionately target old growth primary forest over fallowed secondary forest for new swidden plots.
The highest proportion of incipient regrowth in the study area (up to 1% of all cells) is caused by isodes that maintain a neighbourhood of 8–15 grid cells (Figure 2c). This can be suggested to be the tipping point at which isodes can move a sufficient distance from their previous house garden to avoid stress events (thus maintaining population levels) while creating new swidden plots close enough to old plots to adversely affect their odds of regeneration to secondary forest. This results in isodes creating large tracts of scrubland that have difficulty succeeding beyond this stage of regrowth because of a lack of nearby old growth forest from which to draw pioneer species. The prevalence of incipient regrowth does not appear to impact on the relative proportion of secondary forest, however.
The average time grid cells spend as scrubland (a cumulative metric over the duration each run) is also correlated with mobility magnitude. The peak in the latter between values of 2 and 4 (Figure 2d) is likely because of the same mechanism identified above: short average distances between successive house gardens. Because of the proximity of a given isode to its point of origin, and hence fallowed plots, cleared grid cells are more likely to border grid cells from previous cropping cycle. In one exceptional run, the mean time grid cells spent as scrubland was 200 time steps, or a quarter of the total length of the run. Runs with a logistical range of eight or higher display a sharp contraction in the average time spent as scrubland. This is because an increase in the logistical radius of an isode causes an exponential growth in the quantity of grid cells available for cultivation, reducing the likelihood that fallowed swidden plots will affect the regeneration potential of new plots. This is not true when mobility is lowered, which the results imply leads to outcomes with numerous small yet persistent grid cells of scrubland.
Spatial visualization of some examples lends additional support to these trends, and provides some useful contrasts between the influences of mobility magnitude on forest composition (Figure 3a). Larger distances between new settlements results in more spatially diffuse tracts of anthropogenic forest, although the absolute quantity of secondary forest is greater than in low-mobility regimes (see Figure 2b). Simultaneously, the amount of incipient regrowth is minimized by the distances between swidden plots. Large swidden logistical radii lead to the appearance of tracts of modified forest following long-term abandonment (>200 time steps), and the formation of diverse mosaics of active swidden plots at different stages, secondary regrowth and reverted primary forest (see Figure 3a, inset). Smaller radii (low mobility) epitomizes the opposite, displaying spatially circumscribed zones of secondary regrowth that frequently centre on groups of grid cells with scrub vegetation (Figure 3b). These were observed to form through the mechanism hypothesized from Figure 2c and d, where isodes are unable to move far from old grounds, prolonging the regeneration of incipient vegetation further. The temporality of anthropogenic land-cover change is therefore an important factor to consider.

(a) Patterns of secondary regrowth in a high-mobility, high-regeneration probability run. For clarity, grid cells with secondary regrowth are recoloured to purple, while reverted primary forest is dark green. Inset: Mosaic of farmed swidden plots, reverted old growth, and actively farmed plots. Starting isodes = 30, swidden radius = 10, regeneration probability = 0.125, time steps = 800, random seed = 1950. (b) End state of a low-mobility, high-regeneration probability run in the upper Cuao. Groups of grid cells with incipient scrubland vegetation have been recoloured red for clarity. Secondary and primary regrowth are both displayed as dark green. Large groups of red grid cells rarely form under other circumstances. Starting isodes = 30, swidden radius = 4, regeneration probability = 0.125, time steps = 800, random seed = 1950.
Examining model output diachronically permits three broad trends for incipient vegetation formation to be defined (Figure 4). The first (A) indicates that low mobility causes an early and rapid spike in the quantity of incipient forest plots, irrespective of regeneration potential. It was hypothesized that this would cause frequent stress events among isodes, leading to a reduction in total population and hence anthropogenic impact. This is not, however, borne out by the experimental results; the legacy effects of early large-scale clearances persist until the end of runs. Regeneration probability appears to exacerbate the magnitude of this outcome. The second trend (B) displays a slower rise in incipient forest cover followed by plateaus or continued upward slopes towards the end of runs. Finally, and in the majority, are the set of parameter combinations which result in the creation of few, if any, persistent grid cells of incipient regrowth (C). The mean trend is essentially flat. It is worth noting that, except for a single parameter combination (mobility = 6, probability = 0.025), mobility is the key determinant of incipient vegetation trends, while probability of regeneration only affects the magnitude of the trend.

Diachronic data on the quantity of grid cells with incipient secondary vegetation under all combinations of regeneration (right) and mobility (top) parameters. Red lines are the mean of 30 runs, with the data range in grey envelopes (total: 1350 runs). Three principal trends can be highlighted. A: Rapid initial rise, followed by slow reduction. B: Moderate to low rise followed by sustained growth. C: Low initial rise with little to no growth. Twenty-four out of 45 runs (53.3%) are in this category.
An additional experiment was conducted over a limited number of parameter combinations to investigate changes in the returns of active grid cells over time (Figure 5). The model was allowed to run for three times the number of time steps as in the previous simulations (2400) to detect any long-term diachronic trends in its output that shorter runs might not capture. The most significant qualitative differences in the shapes of these curves are again in low-mobility regimes. Except for where population starts to collapse within a few hundred time steps, the mean returns from active fields (1) become stationary almost immediately. Conversely, the returns from secondary forest (2) fluctuate significantly in the first 1000 time steps and only flatten out after approximately 1500 time steps, far beyond the duration investigated above. Kolmogorov–Smirnov tests indicate significant differences between all four curves in both time series at p < 0.001, in addition to the qualitative variability between parameter combinations. Extrapolating from Figure 5b, the ratio of secondary forest to primary forest appears to reach a dynamic equilibrium, where on average only approximately 2000 grid cells of secondary forest exist any point in time, or <2% of the modelling domain. Secondary forest which has succeeded to climax vegetation (reverted forest) can be predicted to continue to expand over time (see Figure 2a). Finally, while active plots and secondary fields may appear comparable in terms of the sums of their returns, it is worth noting that swidden plots in this model are by definition located within the logistical radius of isodes. Secondary forest, on the other hand, can be widely distributed in the landscape of the upper Cuao (see Figure 3a) and are not necessarily exploited synchronically.

Time series in returns of active grid cells over time: (a) cultivated plots and (b) secondary forest. Data ranges are shown in grey (darker shades indicate overlap), with smoothed means for swidden radius = 4 (purple), 6 (pink), 8 (orange), and 10 (yellow). Regeneration probability (0.075) and number of isodes (30) were held constant over 10 repetitions of each swidden radius value. Random seed = 1950.
Discussion
The aim of this experiment was to constrain the range of plausible scenarios for the scale of impacts of Piaroa land use. Returning to this, assertions that the upper Cuao is a pristine environment largely unaffected by significant LUCCs over its history of human inhabitation (Freire, 2007; Harris, 1971; Huber and Zent, 1995; Perera, 1986; Zent, 1992, 1995) must be questioned in light of the modelling results. Instead, the modelling results provide quantitative backing to the idea that groups of small-scale agriculturalists can cause significant qualitative changes to an environment. In the context of Piaroa land-use patterns, the notion that shifting cultivation is necessarily a ‘high mobility, low impact’ adaptation ought to be modified. The results imply that the quantity of old growth forest affected by human action in the interval since Contact may be up to four times greater than previously assumed, given the parameter space explored in this case. Counterintuitively, high mobility exacerbates this pattern rather than diminishes it. The deleterious effects of shifting cultivation (deforestation, transition to savannah and attendant losses of resilience and biodiversity) emerge only under the most extreme combinations of low mobility and environmental vulnerability. Under conditions where forest succession is impacted over the long term, this manifests in a spatially circumscribed manner, centred on individual Piaroa isodes and their ranges (see Figure 3b).
Shifting swidden cultivation has been compared unfavourably with ‘the cumulative impact of passive, small- to medium-scale and localized disturbance’, meaning agroforestry (Stahl, 2015: 1600). While slash and burn farming is arguably an active form of landscape intervention, its impact is demonstrably cumulative while almost by definition local in scale. In this respect, it is very similar to the various types of terra firme agroforestry documented elsewhere in Amazonia that the above quote refers to. Its persistent reputation as the bête noire of Neotropical subsistence, viewed as limiting to cultural development, wasteful and ultimately unsustainable, is only just being shed (Arroyo-Kalin, 2012; papers in Cairns, 2015). The depth of indigenous knowledge on managing swidden plot nutrient cycles, including biodiverse secondary forest, provides useful templates for exploring this topic computationally (Denevan, 1992a; Harris, 1971). It is clear that the intensive and sedentary pre-Columbian agricultural systems on the margins of rivers, such as those responsible for the creation of anthropogenic dark earths in the central Amazon basin between 2500 and 500 BP, were far from being the only response to subsistence requirements. Indigenous people developed and deployed a variety of different systems depending on the setting and historical circumstances they faced (Arroyo-Kalin, 2010; McMichael et al., 2014; Neves, 2013), each of which left imprints on the landscape. Against this backdrop, the Piaroa modelling experiment must be tempered with palaeoenvironmental data implying low-impact and temporary human occupation in the Amazonian interfluves since the initial colonization of the basin (see Levis et al., 2012; McMichael et al., 2012; Stahl, 2015; Watling et al., 2015).
Although subordinate to manioc production, the Piaroa obtain a great variety of non-domesticated resources from tracts of secondary forest (Overing and Kaplan, 2011; Zent, 1992). In the context of this experiment, it is the emergence of diffuse yet extensive secondary forest tracts which promote high logistical mobility as a comparatively sustainable pattern of land use. Conversely, low-mobility regime runs evidence how the odds of isode survival and the rate of forest succession are adversely affected by the expansion of large scrubland tracts. The fact that the upper Cuao today remains dominated by closed canopy forest suggests that pre-20th-century land-use patterns tended towards the equivalent of ‘high mobility’ runs. Because of the diffuse nature of Piaroa swidden farming, extant tracts of secondary forest (and succeeding climax communities) may become nearly undetectable to the untrained eye. Heckenberger et al. (2007: 197) note that satellite imagery shows Amazonia to be largely flat and green, yet a look under the canopy reveals high biocultural diversity. Johnson et al. (2005) observe,
[A]rchaeologists are not very good at estimating in their heads the effects over long periods of time of slow processes […], particularly when those have to be weighed against a countervailing force such as forest regrowth. It is important to address such processes in a quantitative framework to allow them to be understood. (p. 104)
Taken together, the above observations by Heckenberger et al. (2007) and Johnson et al. (2005) highlight a tension between ‘deep time’ on one hand, and modern observational data on tropical forests and their inhabitants on the other (e.g. Politis, 2007; Rival, 2016). While prior accounts of tropical swidden farming societies (e.g. Conklin, 1961; Harris, 1971; Zent, 1992) are not incorrect or guilty of errors of omission, the shallow time depth of ethnographic fieldwork relative to long-term ecological processes may be a contributing factor to this apparent discrepancy. Historical ecology aims to understand landscapes as historically contingent and emergent artefacts of human activity, the products of centuries (if not millennia) of small-scale actions enacted across the entire biosphere. Consequently, understanding how short-term mechanisms create enduring anthropic patterns of landscape heterogeneity is high on the agenda (Balée, 2006). Because of the nature of the archaeological and palaeoenvironmental records, however, the focus tends to be on the longue durée of human-environmental history through the aggregation of numerous unresolvable individual actions (Isendahl, 2016).
Computational modelling grounded in ethnographic data to represent such actions has demonstrated one way in which this gap may be bridged, using simple generative rules to explore a series of ‘what if’ scenarios (Cegielski and Rogers, 2016; Davies, 2016; Lake, 2014; Magliocca and Ellis, 2016). Building on the premise that the Piaroa operated a system of swidden farming in the upper Cuao for approximately four centuries lends further weight to the idea legacy effects in tropical forests are more profound and pervasive than previously hypothesized. Going further, can Piaroa secondary forest tracts be identified as post-Contact equivalents of pre-Columbian ‘anthropic islands in a sea of forest’ (Neves, 2013: 379)? If that is the case, the results suggest that they are not only extensive but follow people around the landscape too. When viewed together with the ephemerality and frequent movement of isodes in this environment, an archaeology of the upper Cuao (like other interfluvial zones of the Neotropics) becomes a challenging prospect. Extant ecological indicators of disturbance and yet-to-be-collected palaeoecological records, with high-resolution radiometric age determinations, may be the only recourse to reconstructing the pathways taken by the post-Contact Piaroa and their predecessors.
The present study area centres on only one of the secondary rivers occupied by Piaroa groups (and other indigenous people) in the upper Orinoco valley (see Freire and Zent, 2007; Overing and Kaplan, 2011), which was selected as the modelling domain specifically for its relative isolation (see Zent, 1992). It is important to underline that it is not reasonable to expect these results to be mirrored exactly in other parts of the Neotropics where swidden agriculture is practised; the model design was deliberately particularistic (cf. Barton, 2014). Significant factors documented elsewhere, such as the re-use of pre-Columbian landscape alterations (Heckenberger et al., 2007), do not play a major role here. On a general level, however, the results do caution further against assuming that the baseline model for Neotropical forests should necessarily be a natural one, and here it is relevant to note that a consensus on the definition of a ‘natural’ Amazonian environment is itself elusive (Clement et al., 2015; McKey et al., 2010: 7827; Watling et al., 2017). Placing this in a pan-Amazonian context, understanding indigenous environmental impacts and their legacy may be a question of identifying and translating between patterns observable at different spatio-temporal scales (Arroyo-Kalin, 2010; Balée, 2013; Boivin et al., 2016; Levis et al., 2017). A key element of this will be correctly identifying the longevity, magnitude and cumulative effects of pre- versus post-Contact indigenous impacts in different locations (McMichael et al., 2017).
This study of the post-Contact Piaroa shows that simulation may prove a valuable tool in achieving this goal. Returning to the introduction, a fruitful path forward would be to develop comparative frameworks for the degree and types of coupled human-environmental systems rather than re-tread old debates contrasting spatially extensive, low-impact patterns with those which are spatially intensive and locally high impact. Without overstating the extendibility of this specific experiment beyond the upper Orinoco, simulation and computational methods can help bridge ethnohistory, archaeology, palaeoecology and the subsistence regimes inferred or hypothesized to exist from these sources of information on the past.
Conclusion
The kinds of data that historical ecologists, anthropologists and archaeologists are accustomed to handling demonstrably benefit from ever-closer integration. Advances in this area will be especially crucial to properly situating Amazonia in relation to human-induced changes in the biosphere observed globally (Erlandson and Braje, 2013; Kawa, 2016; Kirch, 2005). Computational modelling was adopted to characterize and develop a study of Piaroa ecodynamics, and proposed how discrepancies between the model and the reported environmental conditions of the upper Cuao may be resolved. By taking ethnographies at face value, the agent-based simulations suggest that the environments inhabited by these groups cannot be considered pristine by any metric, even in the relatively short time between European Contact to present. Furthermore, persistent concerns that conceptual models of subsistence in Amazonian risk overemphasizing some parts of a system to the detriment our understanding of the whole (Whitehead, 1998: 38) are not a concern of formal, explicit models. A deliberate epistemic focus on ‘imagined concrete’ properties (Godfrey-Smith, 2006: 738) of a real-world system, rather than precise isomorphism, enabled a robust consideration of the long-term effects of Piaroa swidden farming to be built from the ground up using aspects of known behaviours as a point of departure. The results suggest that the anthropic impacts of swidden farming in the modelling domain can manifest on very short timescales. This has implications for future investigations of the effect of millennia of agroforestry and intensification across the Neotropical world. As noted in the introduction, the ties between model-based science and middle-range theory (Davies, 2016; Premo, 2007) are worth exploiting for historical ecologists. Conceptual models, as semantic entities open to interpretation, will always have barriers between the intent of their original authors and their audiences. Although computational models are not a perfect replacement, they are also not vulnerable to this critique. With effort and informed theory-building, anyone can extend, modify or contest the model discussed here. Indeed, this is welcomed, and may be instrumental to framing expectations for future archaeological research into tropical forest cultures as agents of biodiversity and change.
Footnotes
Acknowledgements
This research was carried out as part of a Leverhulme Trust Research Grant (RPG-2014-234) to José Oliver (University College London (UCL) Institute of Archaeology). He and Natalia Lozada Mendieta provided valuable advice on earlier drafts of this paper. Stefani Crabtree, Benjamin Davies and Colin Wren are thanked for their critical comments on several earlier versions of the Piaroa swidden model discussed here, as well as the two anonymous reviewers. The Instituto Venezolano de Investigaciones Científicas is gratefully acknowledged for provisioning ethnographic literature that is hard to find outside of Venezuela. I also extend my utmost gratitude to our indigenous partners in fieldwork in Venezuela. Any errors are my own.
Funding
The author(s) received no financial support for the research, authorship and/or publication of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.