
Abstract
Marginal stands of balsam fir (Abies balsamea [L.] Mill.) and eastern white cedar (Thuja occidentalis L.) are found north of their limits of continuous distribution in eastern North America. Regional-scale paleoecological studies have suggested that fir and cedar populations could have had larger extents in the past. This study aimed at verifying this hypothesis at the local scale. Wood charcoal fragments were collected from the soils of two marginal fir and cedar stands as well as from 15 sites in the surrounding forest matrix where the species are absent currently. Anatomical identification and radiocarbon-dating showed that fir was more extensive in the study area until about 680 cal. BP, representing up to 31% of the charcoal assemblages at sites where it is currently absent. The evidence is less conclusive for cedar, however, although some of the charcoal fragments from the matrix sites could have been either fir or cedar (undistinguishable). Most of the dated fir/cedar charcoal in the matrix were from the ‘Medieval Warm Period’ (ca. 1000 cal. BP), suggesting contraction may have occurred at that time. Marginal fir – and possibly cedar – stands are thus relics of once more extensive populations. Fire is likely the main factor having contributed to the contraction of the species’ distributions. Fir and cedar are now relegated to areas where fires are less frequent and severe, such as the shores of lakes and rivers.
Introduction
In the face of climate change, tree species will either respond by adaptation, migration, or local extinction (Aitken et al., 2008; Corlett and Westcott, 2013; Walther et al., 2002). Climate models predict that the speed of climate change will require faster migration rates than those observed during the postglacial period (Corlett and Westcott, 2013; Malcolm et al., 2002). Some species, including anemochores (Nathan et al., 2011), will not be able to keep pace with these changes (Clark, 1998; Corlett and Westcott, 2013; Dyer, 1995; Malcolm et al., 2002). In such a context, marginal populations could play a key role in adaptation and migration by acting as dispersal outposts (Hampe and Petit, 2005; Lesica and Allendorf, 1995; Thomas et al., 2004).
Marginal populations are located beyond a species’ limit of continuous distribution. Some of their characteristics might limit their capacity to act as dispersal outposts. First, according to the central-marginal hypothesis, marginal populations often have lower genetic diversity than central populations (Eckert et al., 2008; Pandey and Rajora, 2012), although not always (Gamache et al., 2003; Xu et al., 2012). Second, competition by other species more adapted to local conditions could constrain the ability of marginal populations to expand in response to climate change (Case and Taper, 2000; Messaoud et al., 2014). Third, the small size of marginal populations makes them vulnerable to disturbance, and hence local extinction (Alleaume-Benharira et al., 2006; Kawecki, 2008). To predict how marginal populations will respond to future climate change, it is necessary to understand how they have responded in the past (Hampe and Petit, 2005; Pardi and Smith, 2012).
In eastern Canada, the distributions of balsam fir (Abies balsamea [L.] Mill.) and eastern white cedar (Thuja occidentalis L.) follow a latitudinal gradient. These species maintain marginal populations more than 100 km north of their continuous distribution limit, particularly in areas characterized by low fire incidence, such as the shores of lakes and rivers (Paul et al., 2014; Sirois, 1997). Fir and cedar are late-successional species, poorly adapted to fire (Frank, 1990; Johnston, 1990), and wildfires become larger and more severe from south to north (Bergeron et al., 2004).
Paleoecological studies conducted at large spatial scales have suggested that marginal fir and cedar stands are relics of formerly more extensive populations (Ali et al., 2008; Liu, 1990; Lucas, 2017; Richard, 1993). Larger and more severe fires during the Neoglacial period (after ca. 3000 cal. BP; Ali et al., 2012; Remy et al., 2017) are being pointed to as the possible mechanism to explain fir and cedar contraction. While these studies provide regional information, the Holocene dynamics of fir and cedar populations remain to be explored at the local scale.
Given that forest dynamics are not only influenced by the regional climate but also by local factors (Ali et al., 2009; Cyr et al., 2007; Senici et al., 2015), regional-scale proxies (e.g. pollen) can mask the dynamics occurring at the local scale (Davis, 2000). For example, wildfires produce heterogeneous patterns related to variations in local conditions (Kafka et al., 2001; Mansuy et al., 2014; Turner and Romme, 1994). Consequently, nearby sites are not necessarily subject to the same fire regime and may have different dynamics (Barrett et al., 2013; Clarke, 2002; El-Guellab et al., 2015; Ouarmim et al., 2014). It is therefore relevant to document past forest dynamics at small spatial scales, notably to provide a framework for ecosystem-based forest management (Bergeron et al., 1998; Lindbladh et al., 2013; Lindenmayer et al., 2006).
Wood charcoal resulting from forest fires resist degradation and can be preserved in forest soils for millennia (De Lafontaine and Asselin, 2011). Their anatomical identification to the genus or species level provides information on the composition of burned stands (Scott and Damblon, 2010; Thinon, 1992). Hence, soil charcoal is a robust proxy that can be used to reconstruct the long-term history of vegetation and fire at a fine spatial scale (Asselin and Payette, 2005; Figueiral and Mosbrugger, 2000; Nelle et al., 2013; Payette et al., 2017; Talon et al., 2005; Thinon, 1992).
This study used charcoal analysis to verify, at the local level, whether marginal fir and cedar stands in the northern boreal forest of eastern Canada are relics of formerly more extensive populations. More specifically, it aimed to determine whether – and when – fir and cedar were present at sites where they are currently absent. The potential causes of fluctuations in fir and cedar abundance in the northern part of their ranges are also discussed.
Material and methods
Study area
The study area is located in the black spruce – feather moss bioclimatic domain of western Quebec (between 49°15′N and 49°55′N and between 78°30′W and 79°15′W). It is a low-lying area consisting mainly of clay deposits from proglacial lake Ojibway (Veillette, 1995) and organic deposits in the lower slopes. The climate is of the humid continental subpolar type. The mean annual temperature and mean annual precipitation at the nearest weather station (Joutel, 55 km from the study area) were 0°C and 909 mm, respectively, for the 1981–2010 period (Environment Canada, 2018).
The area is dominated by coniferous stands, mainly composed of black spruce (Picea mariana [Mill.] B.S.P.). Fir and cedar, while within their range of distribution, are much more scarce than further south and fail to occupy all suitable sites (Messaoud et al., 2007b; Paul et al., 2014), and jack pine (Pinus banksiana Lamb.) is mostly found on drier sites. White birch (Betula papyrifera Marsh) and trembling aspen (Populus tremuloides Michx.) are the main deciduous species, mainly found in early successional stands. The undergrowth is dominated by shrubs of the ericaceous family (Rhododendron groenlandicum (Oeder) Kron and Judd., Kalmia angustifolia L. var angustifolia and Vaccinium spp.).
Sampling
Two marginal fir and cedar populations accessible by road were identified (TOC and CDC, hereafter ‘marginal sites’). Both marginal sites were located on the shores of small lakes ca. 50 km apart. In addition, 15 sites were randomly selected from the inland forest of the study area (hereafter ‘sites in the forest matrix’), representative of the diversity of tree species composition and edaphic conditions (Table 1).
Characteristics of marginal sites and sites in the forest matrix.
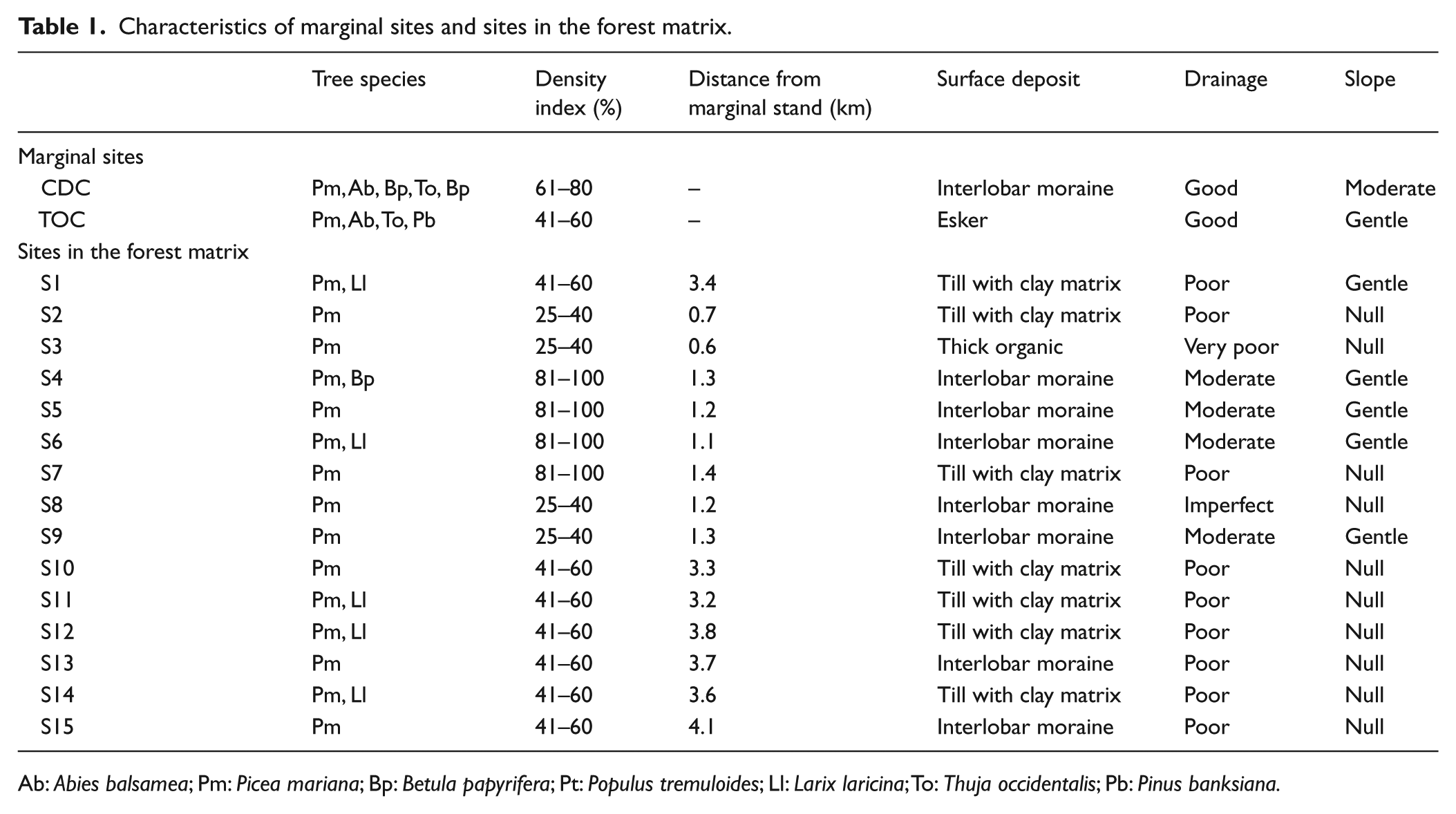
Ab: Abies balsamea; Pm: Picea mariana; Bp: Betula papyrifera; Pt: Populus tremuloides; Ll: Larix laricina; To: Thuja occidentalis; Pb: Pinus banksiana.
Each site was subdivided into microsites where soil samples were taken (averaging 950 cm3). At the marginal sites, microsites were sampled at the corners of three 4 m × 4 m quadrats (12 microsites in total): one quadrat at the center of the marginal site and two quadrats 150 m on each side of the central quadrat. Each site in the forest matrix consisted of one quadrat with four microsites sampled at each corner. At each microsite, charcoal was collected in the mineral soil horizon and at the contact between the organic and mineral horizons (Payette et al., 2012).
Charcoal extraction and identification
Soil samples were immersed in a 2.5% sodium hexametaphosphate solution – (NaPO3)6 – for 12 h to disperse aggregates. The samples were then sieved (2 mm mesh) under a gentle water jet. Charcoal extraction was carried out by flotation and manual sorting under a binocular microscope (Thinon, 1992). Charcoal fragments >2 mm were selected for analysis as they represent local information (Asselin and Payette, 2005).
After extraction, charcoal fragments were identified to the species or genus level according to a reference collection and to wood anatomy references (Panshin and De Zeeuw, 1980; Schweingruber, 1990) and separated into six groups: (1) Abies balsamea; (2) Thuja occidentalis; (3) Abies/Thuja (the criteria for distinguishing cedar from fir – for example the parenchyma wall – are not always visible; Jacquiot, 1955; Panshin and De Zeeuw, 1980); (4) Pinus banksiana; (5) other gymnosperms; and (6) angiosperms. Each charcoal was weighed and the results are presented as the relative mass for each group (Ali et al., 2005).
Radiocarbon dating
A total of 20 fir and Abies/Thuja charcoal fragments (3 within each marginal site and 14 in the forest matrix sites) were dated using the Accelerator Mass Spectrometry (AMS) technique. Samples were prepared for radiocarbon dating at the Centre d’études nordiques (Laval University, Quebec, Canada) and dates were obtained from the Keck Carbon Cycle AMS Facility (University of California, Irvine, CA, USA). The radiocarbon dates were calibrated using the Intcal 13 database (Reimer et al., 2013) of the online Calib program (version 7.1) (Stuiver et al., 2017).
Results
Charcoal fragments were found at all sites and a total of 3435 charcoal fragments were identified. At the marginal sites, fir and cedar (combined percentages of Abies balsamea, Thuja occidentalis, and Abies/Thuja) accounted for 17–53% of the charcoal assemblages (Figure 1). Jack pine, still present today around the TOC site, accounted for 6–13% of the charcoal assemblages at this marginal site. Other charcoal taxa at the marginal sites were mostly other gymnosperms (27–58%), with some angiosperms (2–9%), and 6–29% of non-identifiable taxa.
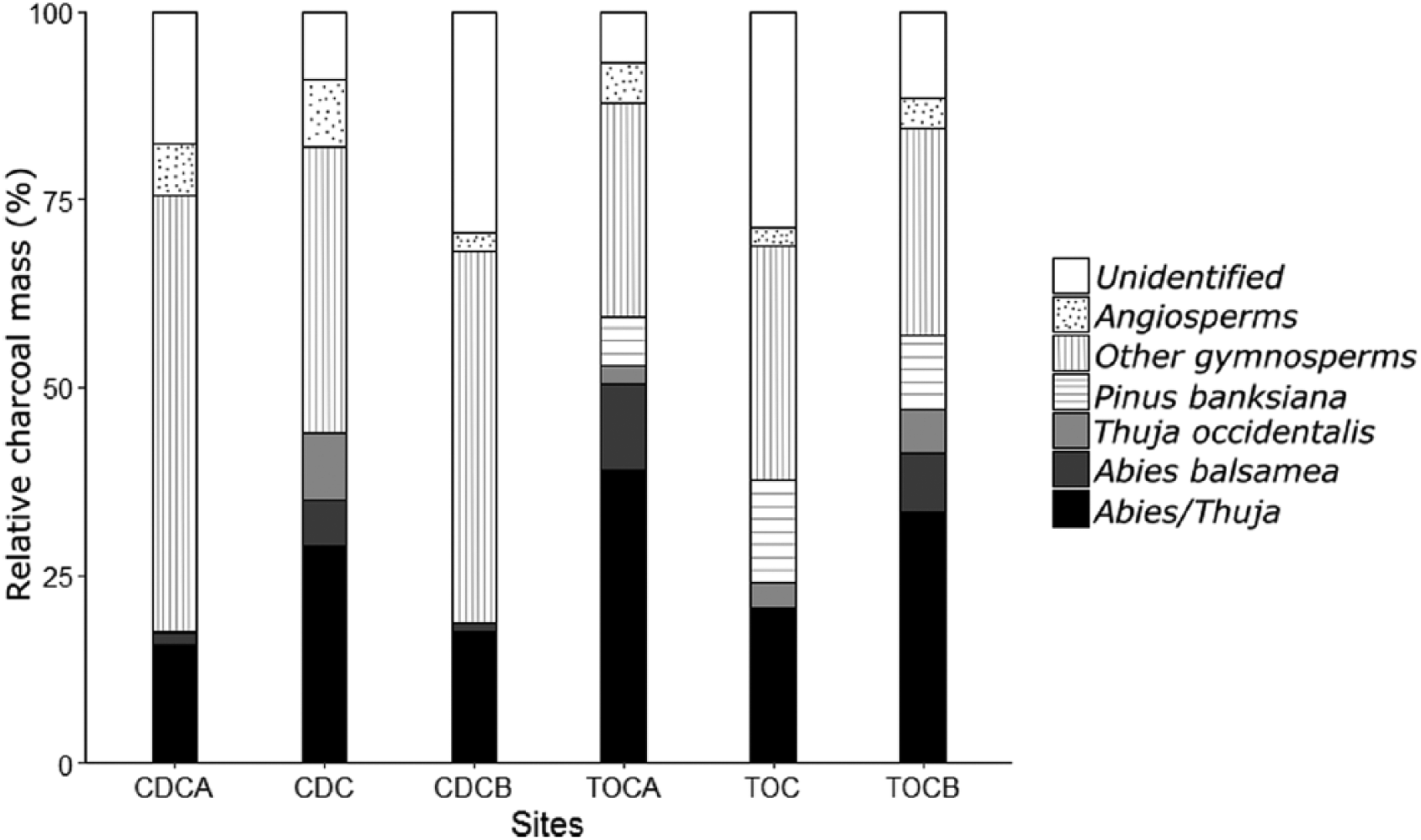
Relative mass (%) for each charcoal category within marginal sites (CDC and TOC) and 150 m on each side of the center of the marginal sites (A, B).
Fir was present in the charcoal assemblages of nearly half of the sites in the forest matrix (Figure 2), while cedar has not been identified. However, as cedar is often present in old fir stands, and given the high proportion of the Abies/Thuja category at some sites, cedar nevertheless could have been present. By combining the relative mass of Abies balsamea and Abies/Thuja, we can say that fir – and possibly cedar – used to be present at 12 of the 15 sites in the forest matrix and represented up to 31% of the charcoal assemblages (site S9). The remaining charcoal fragments were mainly other gymnosperms (13–94%), with very few angiosperms (0–45%) and 6–44% unidentified fragments. The 12 sites in the forest matrix with fir and/or cedar in their charcoal assemblages had variable characteristics. They included both open and closed forests, drainages ranging from very poor to moderate and various surface deposits.

Relative mass (%) for each charcoal category in sites in the forest matrix.
The radiocarbon dates obtained from Abies/Thuja charcoal fragments were between 3755 cal. BP and the modern era (AD 1950) (Table 2). The youngest dates (180 cal. BP to modern) were all from marginal sites. In the forest matrix sites, 11 of the 14 dates were from 679 cal. BP to 1283 cal. BP.
Radiocarbon dates for 20 Abies/Thuja charcoal fragments from marginal sites and from sites in the forest matrix.

Discussion
This study confirmed that fir populations used to have larger extents than today in northern Quebec, and that a climate-related change in the wildfire regime is responsible for the contraction of the species’ distribution during the Neoglacial period. The evidence is less conclusive for cedar, however, although some of the charcoal fragments from the matrix sites could have been either fir or cedar (undistinguishable). According to previous paleoecological studies, fir and cedar arrived in the study area shortly after the retreat of proglacial lake Ojibway, in a landscape dominated by black spruce (Ali et al., 2008; Carcaillet et al., 2001; Richard, 1980). Fir and cedar proliferated during the Hypsithermal period (ca. 6000–3000 cal. BP), before declining gradually during the Neoglacial period (Ali et al., 2008; Liu, 1990; Richard, 1993; Viau et al., 2006). Although this study does not cover the beginning of the Holocene, the oldest date obtained (3755 cal. BP) corresponds to the end of the Hypsithermal period.
The Neoglacial period was characterized by cool, wet summers in eastern North America (Ali et al., 2012; Carcaillet and Richard, 2000). Studies in several lakes 0.2–60 km from the study area suggested that the cooler and wetter climate after 3000 cal. BP would have led to less frequent, but larger fires (Ali et al., 2012; Oris et al., 2014; Remy et al., 2017), probably more severe. As fire size and severity play a key role in long-term vegetation trajectories (Bergeron et al., 2004; Remy et al., 2017), the shift from frequent small fires to infrequent large fires likely explains the contraction of fir – and maybe also cedar – distribution during the Neoglacial period. A similar shift in fire regime was also suggested to have caused balsam fir to disappear from taiga sites located north of the study area (Arseneault and Sirois, 2004; Gennaretti et al., 2014). Fir and cedar do not have an aerial seed bank (Greene et al., 1999; Rowe and Scotter, 1973) and have low dispersal capacity (Cornett et al., 1997; Frank, 1990; Johnston, 1990). Furthermore, seed production by cedar is low in the northern part of its range, where this study took place (Visnadi, 2014). To recolonize burned sites, fir and cedar must count on the seed rain from a nearby remnant stand or from surviving mature individuals within the burn (Asselin et al., 2001; Zasada et al., 1992). Large, severe Neoglacial fires would have thus limited fir and cedar postfire recruitment.
The contraction of fir – and possibly cedar – distribution seems to have accelerated around 1000 cal. BP, during the ‘Medieval Warm Period’ (Hunt, 2006; Viau and Gajewski, 2009). Indeed, most of the dated Abies/Thuja charcoal fragments in the forest matrix were concentrated in a ca. 500-year period centered on 1000 cal. yr BP (Figure 3), a period well known in the area for its increased fire activity (Ali et al., 2009; El-Guellab et al., 2015; Oris et al., 2014; Remy et al., 2017).

Dates when fir/cedar were present in marginal sites (stars) and in sites in the forest matrix (dots).
The role of local factors in the fire regime is well documented (Bergeron et al., 2004; Mansuy et al., 2014, 2010; Parisien and Moritz, 2009; Parks et al., 2012). In this study, evidence of past fir/cedar populations was found at sites with varied topography, surface deposit, drainage and species composition. Therefore, disappearance of the two species from these sites suggests that regional climatic factors might have supplanted the effects of local factors on the fire regime (Senici et al., 2015). For example, sites S9 and S15 had the highest proportion of fir/cedar in their charcoal assemblages (31% and 28%, respectively). Site S9 is an old spruce stand with a thick Sphagnum carpet, while site S15 is in a topographic depression. These features are known to limit fire spread and severity (Cyr et al., 2007; Mansuy et al., 2014; Ouarmim et al., 2015). Being firebreaks, these sites succeeded in maintaining a high portion of fir – and possibly cedar – during the first half of the Neoglacial. But firebreaks were ineffective against the large and severe fires of the ‘Medieval Warm Period’.
In addition to fire, other potential factors have been suggested to explain the decline in fir/cedar in northern Quebec: reproductive failure, competition from black spruce and lack of safe sites for germination (Gauthier et al., 2000; Messaoud et al., 2007a, 2007b, 2014; Paul et al., 2014; Visnadi, 2014). Nevertheless, fire seems to be the overarching constraint to fir and cedar spread north of their continuous range (Ali et al., 2008; Paul et al., 2014). The historical presence of fir/cedar in the forest matrix sites up to about 680 cal. BP suggests that these species can grow in the study area. This corroborates the results of previous studies having shown that many sites theoretically favorable to fir/cedar are not occupied currently (Messaoud et al., 2007b; Paul et al., 2014), as fire kills mature trees, thus limiting regeneration, paving the way for black spruce to take advantage (Messaoud et al., 2014, 2007b). Today, fir and cedar are found in areas with low fire incidence (Denneler et al., 2008; Ouarmim et al., 2015; Sirois, 1997).
Interestingly, marginal sites burned during the last two centuries, but were recolonized by fir and cedar afterwards. This would tend to confirm the role of outposts that marginal sites could play in response to climate change in northern Quebec. In general, the high humidity or poorly drained areas such as wetlands and shores of lakes and rivers limit fire propagation or reduce severity (Araya et al., 2016; Cyr et al., 2005; Nielsen et al., 2016; Senici et al., 2015). Reduced fire severity in marginal sites means that some mature individuals survive and serve as seed sources for post-fire recruitment (Asselin et al., 2001; Zasada et al., 1992). However, for fir and cedar to expand beyond the area of influence of lakes and rivers, climatic conditions will have to be wetter, which does not seem to be part of the predictions for the coming decades (Flannigan et al., 2009; Hély et al., 2010). It is therefore possible that fir and cedar will remain confined to marginal sites.
Conclusion
The presence of fir – and possibly cedar – in Holocene charcoal assemblages from sites where they are currently absent confirms formerly more extensive distributions in the northern part of their current range in Quebec. This study thus corroborates, at the local scale, the results of previous regional-scale paleoecological studies. Wildfire appears to be the main factor responsible for range contraction of both species by eliminating seed sources, hence relegating fir and cedar to sites protected from severe and recurring fires, such as the shores of lakes and rivers. However, for these sites to act as outposts for the northern expansion of fir and cedar in response to climate change, climatic conditions will have to favor the establishment of a regime of light and infrequent fires, which is unlikely given predictions.
Footnotes
Acknowledgements
The authors thank Aurore Lucas for initiating the study, Pierre Grondin for help in the field and Cécile Fouquemberg, Mathieu Frégeau, Marion Lestienne and Vanessa Pilon for help with charcoal identification. Martin Lavoie and Hubert Morin provided insightful comments on an earlier draft.
Funding
This research was funded by the Natural Sciences and Engineering Research Council of Canada (NSERC), the Fonds de recherche du Québec – Nature et technologies (FRQNT), the Centre National de la Recherche Scientifique (France), the MITACS program in association with the Ministère des Forêts, de la Faune et des Parcs du Québec (MFFP) and the Institut Écologie et Environnement through the GDRI ‘Forêts froides’.