
Abstract
Current knowledge of climate change effects on forest ecology and species conservation should be linked to understanding of the past-time. Abies pinsapo forests constitute a model of an endangered ecosystem, highly vulnerable to ongoing warming, whose populations have been declining for centuries, while the drivers of this local depletion trend remain poorly understood. We hypothesized that long-term disturbances, both human- and natural-induced, have shaped A. pinsapo forests, contributing to these decline processes. Until today, studies using fossil pollen record to identify past climate impacts and land-use changes on A. pinsapo populations have not been done. Here, we investigate forests’ dynamics since the late Holocene (1180 cal. AD to present) in Southern Iberian Peninsula from a fossil pollen record by comparing the results obtained with climate fluctuations and land-uses changes. The pollen sequence shows a phase of stability during the Islamic Period (~1180–1400 cal. AD; ‘Medieval Climate Anomaly’), followed by increasing degradation at Christian Period concurrent with ‘Little Ice Age’ (LIA) (ca. 1487–1530 cal. AD). The Modern Period (1530–1800 cal. AD; LIA) is linked to intensive forest management, related to the naval industry. Afterwards, a progressive reduction is recorded during the Contemporary Age period (‘Industrial Period’) until ‘Recent Warming’. In short, historical severe forest management coupled with increasing aridity since LIA appear to influence A. pinsapo forest current species composition and poor structural diversity. These disturbances might be limiting the resilience of A. pinsapo forests under a climate change scenario. A selected forest management could promote a more complex forest structure.
Keywords
Introduction
Mediterranean mountain forests offer a suitable framework to investigate the complex interactions between climate trends and human activity from decadal to millennial timescales (Roberts et al., 2011). Some relict conifer forests are among the most remarkable elements of these Mediterranean landscapes, especially owing to the diversity of their species (Fady, 2005), standing in this region since the Eocene-Oligocene (Barrón et al., 2010). The fossil record supports that Mediterranean forests harbored this significant diversity of conifers, with several Pinaceae species, some of them extinct at present (Postigo-Mijarra et al., 2009, 2010)
Despite the loss of Pinaceae biodiversity and the extinctions described by several authors along the Neogene-Quaternary (Postigo-Mijarra et al., 2009, 2010), some Pinaceae species have been preserved in Mediterranean mountains up to present-day. Currently, they display an ecologically and geographically restricted range, spreading along disjunct mountain areas (Farjon, 1990; LePage, 1999). Abies genus is one of the most common Pinaceae taxa in the Mediterranean, being a group of species closely related genetically but occupying disconnected and sometimes limited areas around the Mediterranean Basin. This group consists of two sections: Abies and Piceaster (Arista and Talavera 1994). Abies section is composed by eight species, including an interspecific hybrid, whose distribution areas are described in Linares (2011). Piceaster section shows a Western Mediterranean distribution and comprises four species: Abies pinsapo Clemente ex Boiss. in Southern Spain; A. marocana Trabut and A. tazaotana Côzar ex Huguet del Villar, both present in the Western Rif (Morocco); and A. numidica Carrière located in Babor mountain (Algeria). The above mentioned taxa are considered as Tertiary relict species, which show fragmented populations of a common ancestor with a wider distribution throughout the mountains of the Mediterranean region (Alba-Sánchez et al. 2010, 2018; Jaramillo-Correa et al., 2010; Linares, 2011; Pérez-Latorre and Cabezudo, 2012).
More specifically, A. pinsapo (pinsapo fir; Cabezudo et al., 2010) populations show a limited and fragmented range along Western Baetic Range (Linares and Carreira, 2009). Furthermore, these populations constitute a model of an endangered ecosystem, highly vulnerable to ongoing climate change, in particular as regards increasing drought (Linares et al., 2011a, 2011b; IUCN Red List: www.iucnredlist.org), among other threats. But generally, the future of the circum-Mediterranean firs, especially Piceaster section, is seriously threatened due to global warming, decreased water availability and the pests associated with climate change (Aussenac, 2002). In fact, warming and drought are among the main stressing factors for several tree species worldwide, leading to local extinction and forest dieback (Allen et al., 2010). Drought may be particularly dramatic in areas already subjected to water deficit (Peñuelas and Boada, 2003), such as the southern Iberian Peninsula, where the Mediterranean climate is typified by severe summer dryness. The current drying summer trend has been related to forest dieback, characterized by tree growth decline and mortality (Camarero et al., 2017). Indeed, previous studies have pointed out that current A. pinsapo growth decline is related to long-term temperature rise, as well as recurrent severe drought events (Linares et al., 2010).
Nevertheless, climate-stress interacts with long-term disturbances and the human-use legacy. Such disturbances, both human and natural induced, have shaped forest dynamics, influencing species composition, stand structure, and functional processes (Dale et al., 2001). In areas subjected to intensive forest management, such as the Mediterranean region, historical thinning, harvesting, or reforestation can influence forest species composition and structural diversity even for centuries (Camarero et al., 2017).
Besides, long-term forest degradation has been related to declining genetic diversity. In this sense, Sánchez-Robles (2013) provides evidences of a homogeneous overall genetic structure across A. pinsapo range using nuclear (nSSR) and chloroplast microsatellites (cpSSR), and amplified fragment length polymorphism (AFLP). This lack of genetic differentiation among populations suggests that the current geographic areas are a result of a recent fragmentation rather than current gene flow, due to the restricted pollen dispersal of the Abies genus (Mazier et al., 2008) and the geographic isolation of the studied populations (Sánchez-Robles et al., 2012). Moreover, in each of the mountain ranges, there are genetically depauperate forest masses, mainly due to the isolation favored by genetic drift. Everything commented above could lead to depauperated genetic diversity and species adaptive capacity. Most likely, an unfavorable environment along Quaternary climate cycles, combined with the already noted historical land-use, favored strong demographic and genetic bottlenecks. Fady et al. (2008) studies support this idea, identifying a genetic diversity gradient among Mediterranean conifers, lower in the most Western than in Eastern ones. So, the case of A. pinsapo appears to be consistent with the hypothesis of genetic bottlenecks in Western conifers over the Quaternary.
On summary, conservation and restoration of forest ecosystems facing global warming need to integrate long-term stand dynamics’ knowledge (Edwards and Mason, 2006; Linares et al., 2011a). Then, significant efforts are still required to identify and evaluate the multiple drivers’ legacy which have led to historical A. pinsapo forest decline. A separate exploration of the causal factors, such as past climate and historical land-uses, will be key to understand their long-lasting interactions over centuries. Resolving this challenge could be feasible through pollen fossil record.
Until today, studies using fossil pollen record to identify the effects of past climate impacts and land-use changes on A. pinsapo populations have not been done. Here, we present the original first A. pinsapo fossil pollen record: (1) to provide the first precise record of A. pinsapo population dynamics in Southern Iberia during the late Holocene; (2) to elucidate whether the historical A. pinsapo forest changes are related to climate or human disturbances; and (3) to help the decision-making process for future management of the A. pinsapo forests.
Methods
Study area and target species
Sampling was performed in Sierra de las Nieves Natural Park (Málaga) at the westernmost Baetic ranges, located in the south of the Iberian Peninsula (Figure 1). Three geological units are defined within the Baetic ranges: Internal Zones, External Zones, and Campo de Gibraltar complex. The study site is found within the Lower Miocene Numidoide Formation of the Campo de Gibraltar Complex, mainly made of sheared brown clay and quartzarenites with typical features of a tectonosedimentary ‘mélange’ (Lozano et al., 2010). The site is set on impermeable clay and sandstone land. On the surface, clays are snuff brown due to alteration processes, while in the deepest parts clays show darker colors.
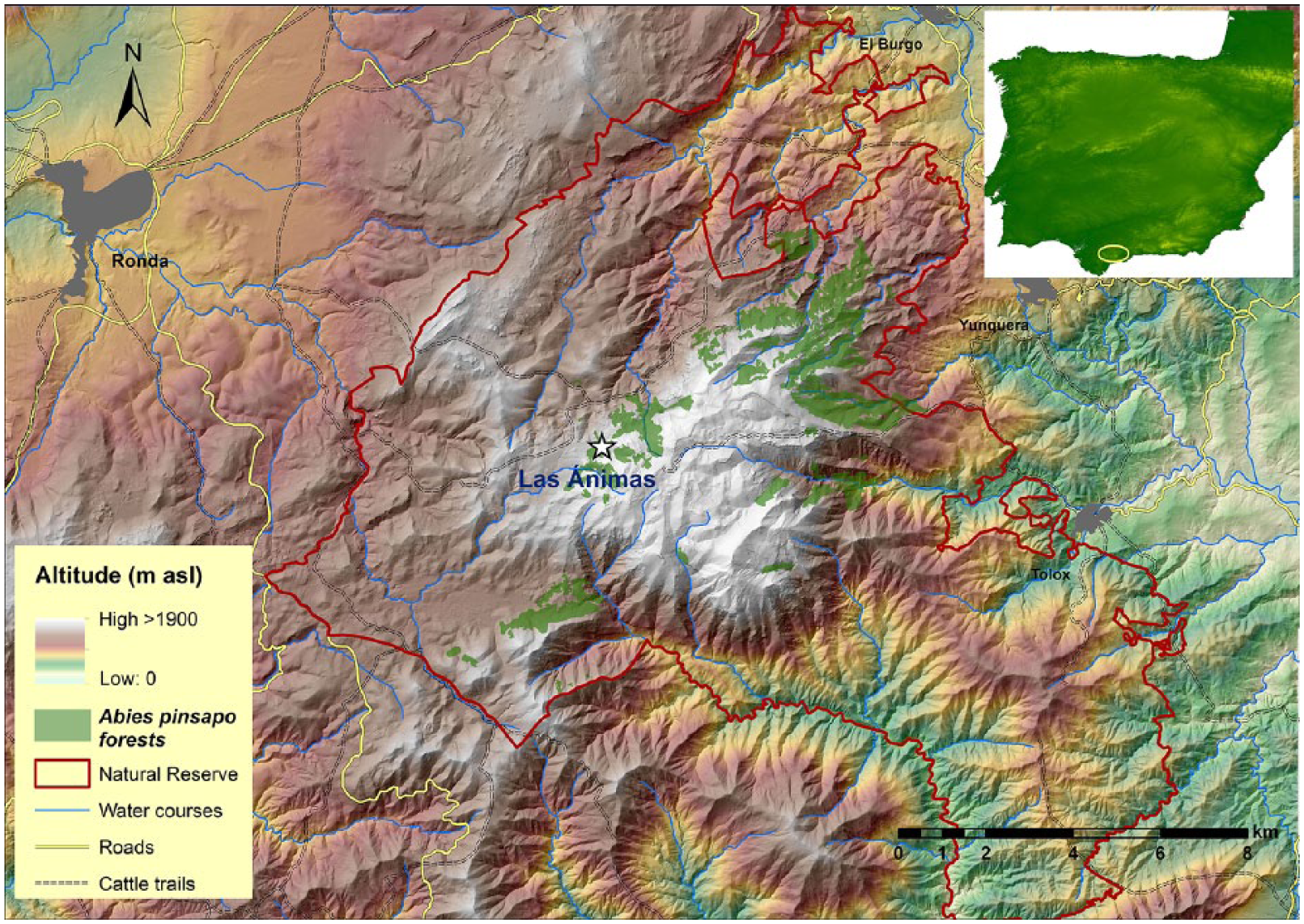
Location of Las Animas marsh (white star), showing the current Abies pinsapo distribution range in Southern Spain (green area).
The climate is Mediterranean pluviseasonal-oceanic type (www.globalbioclimatics.org) with a relatively intense summer drought, a very rainy season that runs from autumn to spring, and a high summer temperatures regime, although the proximity of the sea (it is ranging from 3 to 40 km for the different pinsapo fir populations) softens them. Sierra de las Nieves mountain highlights by the marked character of orographic precipitation. Rainfall values vary from 900 to 1600 mm at lower and high elevations, respectively. With respect to average temperatures, there are high seasonal fluctuations between summer, when the maximum temperature of the warmest month ranges from 24°C to 25°C, and winter, when the minimum temperature of the coldest month is between −1.5°C and 1°C. Finally, the annual average temperature ranges from 9°C to 11°C (Valladares, 2009).
The most representative vegetation is the A. pinsapo forest, dominated by the Iberian endemism A. pinsapo (Pinaceae). A. pinsapo is restricted to three populations on calcareous, dolomite, and serpentine substrates in the Baetic mountains, which occur from 900 to 1800 m a.s.l. in the Sierra de Grazalema (Cádiz province), Sierra de las Nieves, and Sierra Bermeja (both in the province of Málaga).
The pinsapo fir forms monospecific forests (pure forest) with a dense canopy, ranging from 1200 to 1700 m a.s.l, with species such as Helleborus foetidus, Hedera helix, Rubia peregrina, Daphne laureola subsp. latifolia, or Iris foetidissima growing in the understory, while Crataegus monogyna subsp. brevispina, Rubus ulmifolius, Prunus spinosa, Ulex baeticus, and Erinacea anthyllis appear in forest clearings as successional shrubland stages (Pérez-Latorre et al., 1998).
At the lower limit of the pinsapo fir pure forest (1000–1200 m a.s.l.), the mixed-forest develops and it is composed of Quercus ilex subsp. ballota, Q. faginea, Q. suber, Pinus halepensis, or P. pinaster combined to pinsapo fir. Whereas at the upper limit (1700–1800 m a.s.l.), the pinsapo fir forest contacts with high mountain shrub communities (Juniperus communis, J. sabina, Berberis vulgaris subsp. australis, Erinacea anthyllis, Vella spinosa, Bupleurum spinosum, Hormatophylla spinosa, Astragalus granatensis, etc.), together with relict populations of Quercus faginea subsp. alpestris, as well as Acer granatensis, Sorbus aria, and Taxus baccata (Pérez-Latorre et al., 1998).
Coring and chronology
The coring was carried out in a higrophilous grassland (Las Ánimas: 36°41.9′N; 5°1.8′W; 1403 m a.s.l.) dominated by Schoenus nigricans and Molinia caerulea subsp arundinacea. A 235-cm sediment core was extracted in March 2008 using a Rolatec RL-48-L drilling machine equipped with a hydraulic piston corer. The sediment cores were wrapped in film, put in core boxes, and stored in a dark cool room at 4°C prior to laboratory sub-sampling and analysis. Then, the core was sectioned into continuous 5-cm-thick slices.
Ten samples were 14C dated using AMS technique. The AMS dating was conducted at Scottish Universities Environmental Research Center (UK). The dates were calibrated using IntCal13 age calibration curves (Reimer et al., 2013). An age–depth model was produced using Clam 2.2 software (Blaauw, 2010). The best fit was obtained applying a smoothing spline (error-weighted with smoothness set at 0.5) to the available radiocarbon dates. Confidence intervals of the calibrations and the age–depth model were calculated at 95% (2σ).
Pollen analysis
Pollen analysis was carried out on 39 sub-samples of 5 cm3 along the core length. All samples were treated chemically (HCl, KOH, HF) following the standard procedures described by Moore et al. (1991), using Thoulet solution for densimetric extraction of pollen and non-pollen microfossils (Goeury and de Beaulieu, 1979). Macrofossils were not discerned throughout the core. Palynomorphs were identified at 400× and 1000× magnification with the help of the reference collection of the Institute of History of the CSIC-Madrid, as well as identification keys and photo atlases (Beug, 2004; Cugny et al., 2010; Moore et al., 1991; Reille, 1999; van Geel, 2001). Pinus pinaster and Quercus suber pollen differentiation was possible following Carrión et al. (2000a, 2000b). Pollen concentration (grains cm−3) was estimated by adding a Lycopodium tablet to each sample (Stockmarr, 1971). Values were divided by deposition time (year cm−1) to calculate pollen accumulation rate (PAR; grains cm−2 year−1). Pollen counts of up to 400 grains of total land pollen per sample were identified and counted. Pollen of aquatic or wetland plants as well as spores and non-pollen palynomorphs (NPP) were excluded from the pollen sum. The pollen diagram has been plotted against age using TGview (Grimm, 2004). Local pollen assemblage zones (LPAZ) were constructed on the basis of agglomerative constrained cluster analysis of incremental sum of squares (Coniss) with square root transformed percentage data (Grimm, 1987).
Charcoal analysis
The number of charcoal debris was calculated under microscope along with the identification of pollen and classified into two size classes (Vannière et al., 2008), namely >125 and <125 µm, indicating local and regional fires, respectively (Whitlock and Larsen, 2001). Charcoal accumulation rate (CHAR) was calculated by dividing the concentration of microcharcoals by the sedimentation rate (cm year−1) and is expressed in particles cm−2 year−1 (Long and Whitlock, 2002). It should be noted that the pre-treatment process can damage or split original large charcoal debris into smaller pieces. Yet, since all samples were processed with the same procedure, we assume that this systematic bias was uniform across samples and therefore did not influence the temporal variation of charcoal abundance. In fact, Carcaillet et al. (2001) and Yin et al. (2016) confirmed a similar pattern when sieving and pollen-slide charcoal series were compared.
Results and discussion
Chronology
Table 1 shows the AMS-radiocarbon data with 2σ range of calibration from Las Ánimas marsh, and Figure 2 displays the age–depth model calculated for the study core using Clam 2.2 software (Blaauw, 2010).
AMS-radiocarbon data with 2σ range of calibration from Las Ánimas marsh.

C: charcoal; OS: organic sediment.

Age–depth model calculated for Las Animas core. The solid black line is the best-fit spline generated by the CLAM 2.2, while the gray envelope shows the final 95% confidence intervals based on 1000 iterations.
Pollen, NPPs, and CHARs
A total of 34 pollen, spore, and NPPs types were identified. The results of identification and counting are presented as percentage diagrams. Herbs and NPPs were also grouped according to their ecological affinities (Figure 3). The percentage pollen diagrams can be divided into four zones (LPAZ) which were divided into six subzones (LPAZ-1 (1a-1b) to LPAZ-4 (4a-4b)) using a constrained cluster analysis (Grimm, 1987). The six subzones description indicating percentages are shown in Table 2. A synthetic diagram is also included (Figure 4), in which main environmental changes, and a summary of selected pollen types, NPPs, and CHAR over the last ~800 years in the Animas marsh are shown.
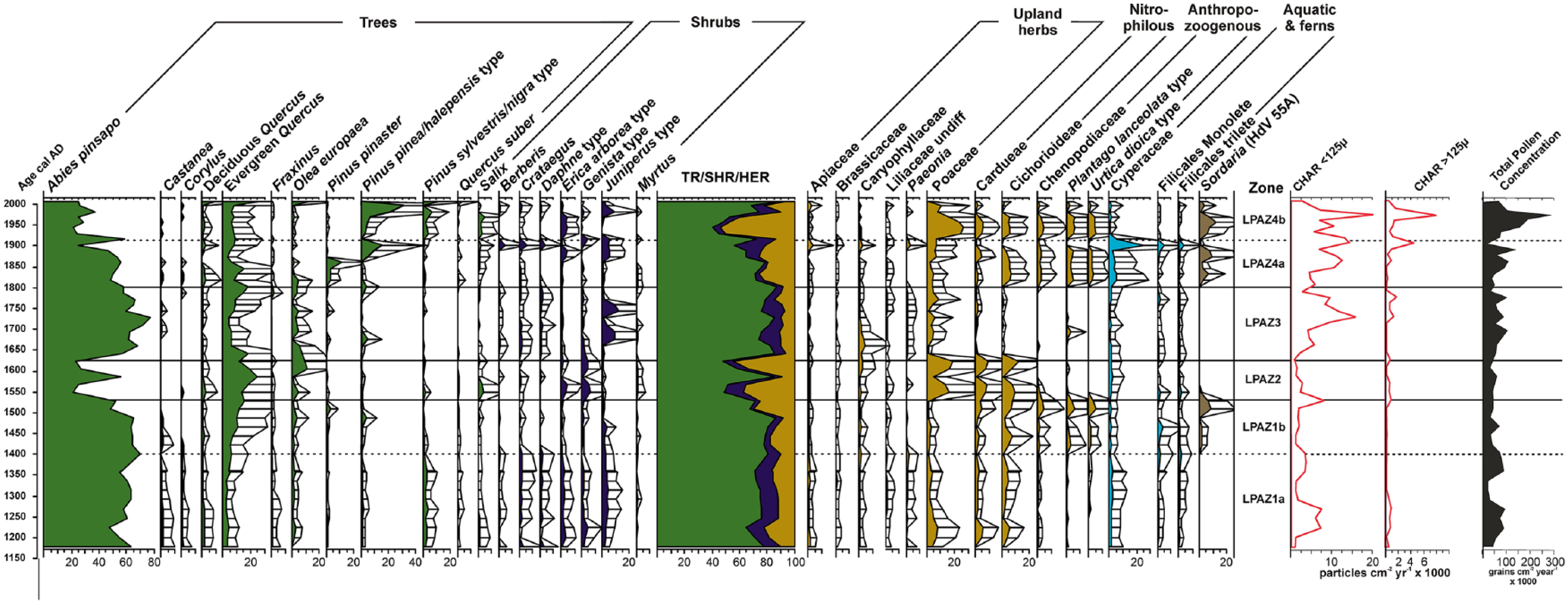
Summary diagram including selected pollen types, NPPs, CHAR, and total pollen concentration.
Description of pollen zones indicating percentages.
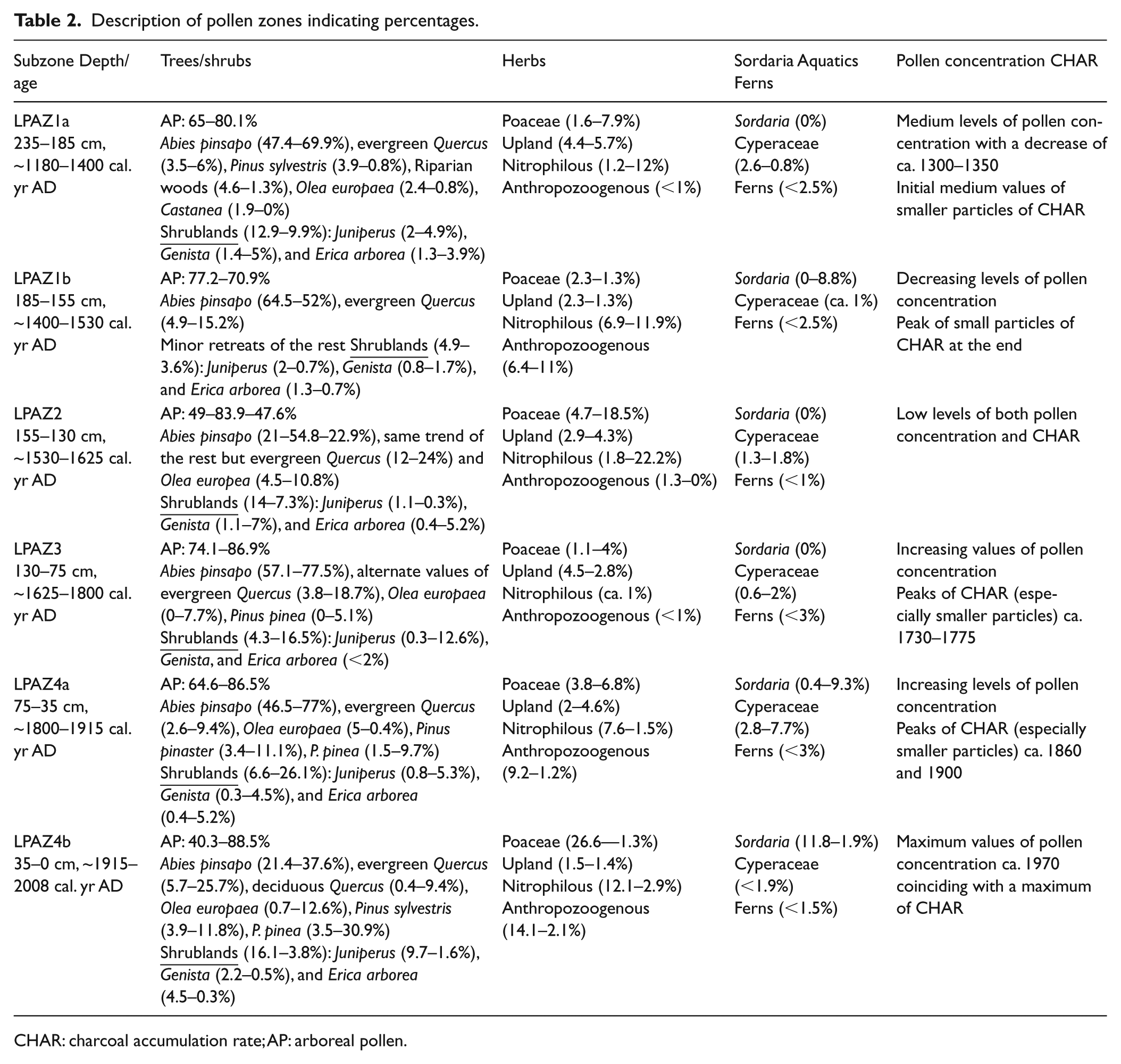
CHAR: charcoal accumulation rate; AP: arboreal pollen.

Main environmental changes over the last ~800 years in the Animas marsh. Abies pinsapo and Quercus ilex pollen; for the explanation of the pollen groups: Poaceae and Upland herbs; Coprophilous (Sordaria), Nitrophilous, and Anthropozoogenous (Anthropogenic Communities). Charcoal accumulation rate (CHAR) and classification into two size classes (CHAR ⩽ 125 CHAR ⩾ 125). NAO Index from a climate proxy reconstruction from Greenland (Olsen et al., 2012); reconstruction of the total solar irradiance (Bard et al., 2000); annual reconstruction of Northern Hemisphere surface temperatures (Wahl et al., 2010); oscillations in precipitation recorded by Mg/Al ratio and Rb/Al ratio in the Alboran sea and MCA (‘Medieval Climate Anomaly’), LIA (‘Little Ice Age’), IP (‘Industrial Period’), and RW (‘Recent Warming’) (Martin- et al., 2010); dry (yellow) versus humid (light blue) phases from the first eigenvector of the west Balearic-Algerian Basin 306G core (Nieto-Moreno et al., 2013).
Islamic Period: A. pinsapo forest, a low human-disturbed landscape (ca. 1180–1400 cal. AD)
This phase coincides with the Late Muslin Iberian domain period in Al-Andalus territory. Climatically, this subzone (LPAZ1a) corresponds to Medieval Climatic Anomaly (MCA; 800–1300 cal. AD) (Nieto-Moreno et al., 2013) with a warm climate and relatively arid conditions in the Mediterranean Iberia (Figure 4). According to Moreno et al. (2012), MCA was a period characterized by decreased lake levels, more xerophytic and heliophytic vegetation, a low frequency of floods, and less fluvial input to marine basins. The scarceness of Cyperaceae and ferns at local scale could confirm this general picture (Figure 3, Table 2).
Under the above scenario of dry and warm conditions, pinsapo fir pollen have reached very high levels along this record, displaying a wide extension of pure and mixed-forest (ca. 1180–1300 cal. AD; Figure 3), which persisted beyond the MCA period up to ca. 1487 cal. AD in the LIA period (Figure 3). The collected data provide insights into the well-development state of pinsapo fir forests throughout the whole second half of the Muslin Iberian period covered by the Animas’ palynological record, although signs of recurrent slight retractions are also observed. Arboreal pollen (AP) shows high levels (65–80.1%), mainly dominated by A. pinsapo (47.4–69.9%; Table 2) which displays slight signs of decay/recovery along this stages. In addition, the well-development state allows the expansion of shrub communities and herbaceous species characteristic of this type of forest. Shrub communities display high, although decreasing, levels as time progresses (Juniperus, Genista, Erica arborea, Crataegus, Daphne and Berberis) (Figure 3). Herbs keep relatively low levels, alternating their peaks with those of tree species, while upland herbs (4.4–5.7%) like Apiaceae, Brassicaceae, Caryophyllaceae, Liliaceae, or Paeonia rather follow the arboreal pattern, also showing the high levels of diversity on A. pinsapo forest.
At the lower limit of pinsapo fir pure forest, the mixed-species forest develops, with species such as evergreen Quercus, deciduous Quercus, P. pinaster, and P. halepensis, which exhibit insignificant fluctuations during this phase (Figure 3). It is worth noting the influence of vegetation on a regional scale. The abundance of P. sylvestris pollen (<3.9%) probably comes from surrounding mountains, showing slight signs of decay. Riparian woods (<4.6%) appear to be likely growing on well-development state and extensively across the landscape, although they show signs of decay, probably due to the effects of MCA. Finally, the presence of Myrtus pollen may come from more thermophilic environments.
Regarding land-uses, during the period of Al-Andalus, a shift in the approach to territory and sustainable resource management emerged on the base of intensified agriculture through irrigation (Civantos and María, 2014; Pérez Ordóñez, 2005). In fact, the fossil data provide insight on the sustainable management of the pinsapo fir forests, which left no strong signs of human-impact on the Malaga mountains. Among other benefits of irrigation such as extending the harvest time and boosting crop yields (Trillo San José, 2005; van der Veen, 2010), it allowed the cultivation of plants from India, Southeastern Europe, Southeastern/Southwestern Asia and Africa, some of which are reflected in the pollen diagram (Figure 3). Nut and fruit trees, such as Corylus avellana (hazelnut), Castanea sativa (chestnut), Juglans regia (walnut), Prunus dulcis (almond), and Ficus carica (fig tree) were abundant and provided important winter food. Morus alba (mulberry) was introduced by the Moors for silkworm rearing (García and Sánchez, 1981, 1988, 1992; Vaquerizo Gil, 2017).
According to Trillo San José (2005), the yield of traditional rainfed crops (cereals and olive trees) also were improved by means of irrigation. More productive harvests per year would have been ensured, with wheat and barley being dietary staples. The irrigation of cereals could be evidenced by the abundance of nitrophilous herbs (1.2–12%) during this phase (Figure 3, Table 2), consisting of Cardueae and Cichorioideae. However, contrary to expectations, the olive yield at this time appeared to be low, as reflected by the fossil sequence (2.4–0.8%; Figure 3).
Also, Watson (1983) suggests the irrigation water even provided nutrients to such an extent that animal manure was hardly needed, whereas poultry fertilizers and plant remains were commonly used. The lack of anthropozoogenous herbs like Chenopodiaceae, Plantago lanceolata, and Urtica dioica (1% altogether), as well as Sordaria spores (0%) in this subzone confirms the absence of large-scale livestock breeding (Figures 3 and 4) and implies the decoupling between agriculture and livestock farming in opposite of the feudal system widely spread in Europe (Watson, 1983). In addition, the low percentage of local microcharcoal (CHAR > 125 µm) tends to suggest lack of exploitation or clearing inside the fir forest to encourage the livestock. All the evidences given suggest that the raising of livestock and herding of cattle appear to be secondary in the local economy (Trillo San José, 1999), in contrast to neighboring regions where merino sheep were the basis for trade and, therefore, was widespread by means of the transhumance, that is, a traditional livestock practice (Klein, 1920; López Saez et al., 2009, 2013; Robles-López et al., 2017; Svensson and Costello, 2018).
Christian Period: Increasingly disturbed landscape after Islamic Period decline (ca. 1400–1530 cal. AD)
This phase starts before the conquest of Ronda (Malaga) by the Christian Kingdom at 1485, which involves the incorporation of the feudal system common throughout Europe (Pérez Ordóñez, 2005). An increasing transformation of the surrounding mountains and land-use management took place along the second subzone LPAZ 1b (Figure 3). This stage is included in the LIA, characterized by a generalized increase in water availability (Benito et al., 2003; Moreno et al., 2008), due to a rise in winter rainfall (negative NAO phase; Nieto-Moreno et al., 2013; Figure 4), and cooling oscillations which probably led to glaciers advance in surroundings mountains such as Sierra Nevada (Gómez Ortiz et al., 1996).
The generalized cool and rainy framework might have favored that forest ecosystem remained almost unaltered in an early stage, but progressively decaying over time, as pollen record indicates (Figure 3). A gradual decline in the cover of A. pinsapo (64.5–52%) accompanied by a possible advance of evergreen Quercus (<15.2%) at the lower forest limit were observed (Table 2). Such landscape change may be related to specific clearing events as a result of intensified human use during the Spanish occupation of medium-elevation forests. The lower amounts of CHAR particles could indicate few regional and local fires, suggesting the lack of fires for forest management, likely stimulated by humid and cooling conditions. A peak of local fires at the late stage was detected possibly caused by human action and a recovery of dry conditions (Figures 3 and 4). The degeneration of the forest caused understory species also to become sparser and, as a result, the percentages of shrublands (Juniperus, Genista and Erica; Table 2) fell. The lower pollen count of riparian species (i.e. Fraxinus, Salix; Figure 3) suggests that this type of vegetation was also reduced by human action, as climatic conditions prevailing in the area at this period tends to promote the spread and maintenance of these forests.
As opposed to an urban and monetary economy during the Islamic period, the new settlers tried to adapt the Andalusian lands to their autarkic economic and social structure (Pérez Ordóñez, 2005) causing severe changes in the land uses in various ways. First, the development of the extensive rainfed crops in the mountain areas (e.g. cereals, olive trees, and grapevine), as could be reflected by the proliferation of nitrophilous herbs (6.9–11.9%) as well as Olea pollen (<4%), while decreasing percentages of upland herbs (2.3–1.3%) and Poaceae (2.3–1.3%) are also recorded (Figure 3; Table 2).
Second, large-scale animal raising, causing both activities, crops and livestock, to be combined again. High levels of grazing, anthropozoogenous herbs (<11%) and Sordaria spores (<8%), are now found (Table 2). In short, the resulting data suggest that a new economic structure led landscape to become progressively more disturbed mainly due to grazing and rainfed areas.
Early Modern Age Period (Habsburg dynasty): Serious deforestation events in favor of shipping building (ca. 1530–1625 cal. AD)
Heavy deforestation events of pinsapo fir forests occurred during Philip II’s reign (1556–1598). This period was marked by global exploration and territorial expansion across the Atlantic and Pacific oceans, leading the Spanish monarchy to become the preeminent power in Europe (Parker, 2000). This zone (LPAZ-2) is part of the LIA but contrary to the previous stage a dry period occurs at AD 1500–1590 (Nieto-Moreno et al., 2013; Figure 4). However, these drier and cooler climatic conditions in the area do not appear to be the main drivers of these sharp decreases (Figures 3 and 4).
The severe deforestation episodes of A. pinsapo forest (Figure 3; Table 2) came as a result of providing timber for shipping building. Lumber was essential for maintaining the fleets that controlled Spain’s transatlantic empire in the 16th century, and the masts and planking of the Spanish Armada are thought to have been made from A. pinsapo, although this has not yet been confirmed (García Esteban and de Palacios, 2007). In parallel, as a symbol of the Christian conquest, churches were built in the conquered territories. For instance, there is evidence that Santo Domingo convent (Sanlúcar de Barrameda, Cádiz) was built using pinsapo fir (AD 1528–AD 1605; Cruz-Isidoro, 2011).
The contraction of the forest matches with the expansion of shrubs such as Genista (<7%) or Erica (5.2%) as well as with a major spread of upland herbs (<4.3%) and Poaceae (<18.5%) (Table 2). At this point, despite its overall increase during this period, the oak forest (<24%) underwent a similar fluctuation as that of A. pinsapo. Therefore, human pressure at the lower limit of the A. pinsapo pure forest, bordering the mixed-forest (ca. 1000–1200 m), may have been one cause for the invasion and rise in elevation of Q. ilex.
Nevertheless, Pinus pinaster and P. halepensis in the mixed-forest did not follow the same pattern as the oaks did, but rather declined sharply for naval shipbuilding (Monchet, 2016). Supporting this idea, no CHAR data could attribute this deforestation to fire or charcoal making (Figure 3). The bulk of evidence shows that the clearing of the forests for shipbuilding during Felipe II’s reign was ecologically destructive, as described by Parker (2000).
Regarding traditional farming, the crown permitted the population’s access to forest resources. It could not prevent people from cutting wood for individual use because so many people depended on the forest (Wing, 2012). Also noteworthy, other land uses are reflected by the increasing percentages of Olea europaea (<10.8%), specifically during the second deforestation event (Figure 3; Table 2). Therefore, this rainfed crop was presumably introduced at the lower elevation of the pinsapo fir forest where trees were cleared. Simultaneously, nitrophilous herbs significantly increased (<11.9%), apparently confirming human pressure on the lower forests limits for agriculture (Table 2). Surprisingly, during this period the signs of livestock were scarce, anthropozoogenous herbs (<1.3%) and Sordaria (0%). These low levels appear to indicate that cattle grazing occurred far below the fir forest.
Modern Age Period (Habsburg-Bourbon dynasty): Starting point for pinsapo forest conservation and management (ca. 1625–1800 cal. AD)
Tree felling as a strategic resource for shipbuilding during the Habsburg dynasty marked the turning point in timber depletion and the beginning of Spanish forest management. The 17th- to 18th-century transition coincides with an LIA stage of alternating humid and dry phases. Two rainfall anomalies were registered in southern Spain (AD 1590–1650 and AD 1650–1750; Rodrigo et al., 1999). Whereas a dry period occurred from AD 1500 to 1590 coupled with a lowering of temperature (Maunder minimum; Nieto-Moreno et al., 2013).
To prevent further Iberian deforestation, strict regulations and forest conservation (Martínez and González, 2014), including afforestation for navy use, were implemented as standard practices from the 16th century on (Wing, 2012). During the 18th century (Bourbon dynasty), the “ Forest Ordinance for the Navy (1748)” was the first law for forest protection in Spain (Bauer, 1980; Rozas, 2003) and established the political basis for oceanic exploration by the Spanish monarchy (De Aranda Antón, 2003).
Forest conservation, which started in ending 16th century, appears to have been successful at the beginning of the LPAZ-3 zone, given an initial Pinsapo fir recovery (Figure 3), and due to the onset of wetter conditions (Figure 4; Nieto-Moreno et al., 2013), thus leading the A. pinsapo forests to reach their maximum expansion (~77.5%) ca. 1725 cal. AD. The high pinsapo fir values also correlated with Q. ilex minimums, possibly as a result of continuous conservation and afforestation since the 16th century (Wing, 2012). Furthermore, the aforesaid cooling event (Maunder minimum; Figure 4) could have favored the spread of the fir-forest at the lower limit (Alba-Sánchez et al., 2018; González-Hernández et al., 2015), perhaps as a result of competitive displacement of Q. ilex.
The high values of the fir forest also coincide with faint signals of human pressure, especially related to grazing, reflected by low levels of coprophilous fungi and anthropozoogenous herbs (<1%). Low values were found for nitrophilous herbs (ca. 1%) and Olea europaea (<7%) over the period (Figure 3). The traditional uses of forests by local communities (firewood, lumber, open grazing, land clearance by fire) were severely restricted following the “Forest Ordinance for the Navy (1748)” (De Aranda Antón, 2003). This mild human-impact also agrees with sudden shifts in the rainfall regime and the cooling maximum during the LIA (Figure 4), which could have hampered the development of traditional field activities in the surrounding areas. Several floods caused by the Guadalmedina river (Malaga), and epidemics, accompanied by several successive poor harvests during the 17th century have been documented (Ringrose, 1998). The traditional use of forests by local communities (tree lopping, gathering wood for lumber, uncontrolled grazing, land clearance by fire) was severely restricted, and agents of the Navy periodically surveyed the forests, logging trees adequate for shipbuilding.
Later, from ca. 1725 to 1800 cal. AD, the A. pinsapo forest progressively declined, coinciding with a peak in CHAR. The A. pinsapo decline agrees with the spread of Juniperus spp. and Quercus ilex. Juniperus appears to have spread around the treeline in response to previous cooling conditions (Figure 3). Meanwhile, the Q. ilex increase could be related both to the changing climate and the steady decline in the pinsapo fir forests, facilitating the spreading of the oaks at the lower limit.
The renovation of the Spanish navy in the mid-18th century owing largely much to the reforms of Fernando VI led to improved management of domestic timber for shipbuilding. A shipyard using forest resources (La Carraca, Bay of Cadiz) has been documented (Wing, 2014), but it is unknown whether this wood was fir. In 1749, the crown ordered the felling of trees in the mountains of Malaga province in order to build six ships in the Cartagena shipyards (De Aranda Antón, 2003). Curiously, this shipbuilding coincides with the shrinking pinsapo fir forest (Figure 3). However, there is no conclusive study demonstrating that A. pinsapo lumber was suitable for shipbuilding. Extensive evidence has shown the use of A. pinsapo timber for civil-engineering structures (Ronda bullring seating in 1785, territorial demarcation, Ronda bridge scaffolding in 1751–1793; Becerra Parra, 2006; García Esteban and de Palacios, 2007; Guzmán Alvarez et al., 2013). A. pinsapo also provided wood for building houses (Guzmán Alvarez et al., 2013) given that this was not prohibited by the crown.
At a local scale, the construction of a tin factory has been documented in Júzcar (Malaga) in 1730 (under the reign of Felipe V), which fostered an economic recovery (Gómez Zotano, 2004). Nevertheless, the socioeconomic resurgence led to a serious environmental deterioration of the surrounding forests due to the newly permitted mining, logging, and charcoal production. The trees most affected were chestnut, oak, and A. pinsapo. The increase of CHAR (Figure 3) appears to indicate local and regional burning.
Contemporary Age period: Increasing forest decline under liberal principle of private property and industrialization expansion (ca. 1800–1915 cal. AD)
Despite the 19th-century political, economic, and social transformation in Spain with the advent of modern industry, agriculture and animal husbandry remained the predominant sector of the economy. The prevalence of moist climatic conditions continued during most of the IP (‘Industrial Period’; Nieto-Moreno et al., 2013; Figure 4). A negative mode of the NAO persisted during this time, with moist conditions governing the southern Iberian region, as evidenced by instrumental precipitation series since AD 1750–1950 (Rodrigo et al., 1999). Our data may also support this evidence, reflecting moist conditions in the sharp increase in Cyperaceae (2.8–7.7%; Table 2). Anyway, in terms of forest dynamics, climate played a secondary role.
The administration of Spanish forests dramatically shifted to the liberal principle of private property over the prior communal land ownership (Bauer, 1980). In 1833, the General Ordinance of Forests allowed the unrestricted elimination of forests for common grazing, serving agricultural interests. Forest decline progressed over the early decades of 19th century (Figure 3), suggesting intense grazing pressure in the study area, Sierra de las Nieves, during this stage. For instance, in the first third of the century, the density of livestock in Malaga province increased brusquely (Figure 3). The anthropozoogenous herbs (until 9.2%) as well as coprophilous spores (until 9.3%) began a sharp increase on the Malaga mountains (Table 2).
Later, the Mendizabal Desentailment of the liberal government (1837 and 1855) was the most critical event for the fate of the Spanish forests because of a further land privatization (Valbuena-Carabaña et al., 2010). Millions of hectares of forests were logged and converted to agriculture (Bauer, 1980). However, A. pinsapo forests survived because toward the mid-19th century they were transferred to state ownership during this process (Bauer, 1980; Gil and Torre, 2007), resulting in a slight forest recovery (A. pinsapo, P. pinaster and Q. ilex) (Figure 3). This paralleled a reduction in human pressure at this time (see decrease of Sordaria, nitrophilous and anthropozoogenous herbs; Figure 3). Furthermore, the 1863 National Forest law prevented forest destruction and, supported by science, established the basis for modern forestry policy (Bauer, 1980; Marraco Solana, 1991). This law preserved the A. pinsapo forest to remain as communal, prohibiting its transformation into pasture (Bauer, 1980; Figure 3).
In the second half of the 19th century, an intensive steady regression of forest cover (A pinsapo and Q. ilex) was detected. Several factors could have been responsible. On one hand, a period of prosperity in Malaga province began due to new industry based on Marbella blast furnaces and Malaga ironworks (Guzmán Álvarez et al., 2013; Montoro, 2007). The production of charcoal from A. pinsapo and Q. ilex forests on the surrounding mountains (mainly in Sierra Blanca and Sierra Real) made this company the major iron supplier in Spain. Moreover, the Algeciras-Ronda railroad was built using A. pinsapo lumber (García Esteban et al., 2010; Soto-García, 2006), although, apart from the Grazalema mountains, the origin of the timber is unknown. In addition, A. pinsapo forests were seriously threatened by woodcutting for domestic firewood, for covering ice cellar during the hot season, and also for paper pulp (Barbey, 1931; Ceballos and Ruiz de La Torre, 1979; Ceballos and Vicioso, 1933). A sharp A. pinsapo minimum (ca. 1875 cal. yr AD) might have been prompted by recurring local fires (see CHAR Figure 3).
Contemporary Age Period: Multifactorial inheritance lead to a heavily deforested landscape (ca. 1915 cal. AD–present)
At the beginning of the 20th century, the Iberian landscapes were drastically deforested. As will be discussed below, the overall decline of A. pinsapo forests appears to have been human-induced (Soto-García, 2006), although a dry period (Figure 3; Nieto-Moreno et al., 2013; Rodrigo et al., 1999) since AD 1950 and jointly rising temperatures (Crowley, 2000) could have exacerbated the decline and hampered tree regeneration (Cabezudo et al., 1995). The sudden onset of a positive-NAO index led to dry winters in the Mediterranean region (Trigo et al., 2002; Trouet et al., 2009; Wanner et al., 2001) until the present day.
A. pinsapo forest contracted notably in the beginning of this subzone (Figure 3). The critical event for the forest decline was the emergence of intensive cultivation of former forest lands and intended mainly for animal feed (Silva Pérez, 1999). La Mesta (transhumance system) dissolution led to more extensive grazing and an uncontrolled use of forests and grasslands which likely contributed also to increase wildfire incidence (Camarero et al., 2018). A similar picture is observed in other areas such as the Iberian Central Mountains (Abel-Schaad et al., 2014; Franco-Múgica et al., 2001; López-Sáez et al., 2016).
The intensification of agrarian practices led to more sedentary livestock farming (Ruiz and Ruiz, 1986). Concomitantly, Poaceae (26.6%), nitrophilous (12.1%), and anthropozoogenous communities (14.1%) (Table 2) rapidly expanded. CHAR, especially smaller particles, registered significant peaks throughout this period. Thus, all evidences appear to indicate that fires were set to clear areas for cattle grazing at a regional scale.
Later, the opening of a market economy (around 1950s) caused the collapse of some traditional agricultural systems, resulting in a rural exodus (Silva Pérez, 1999). The steady shift in land uses (Figure 3), that is, the spread of thickets, reduction of pasturelands (Poaceae) and grazing areas (Nitrophilous and Anthropozoogenous herbs), as well as the reforestation with tree species (Pinus spp.) could have accelerated the marginalization of these rural areas. In addition, depopulation could have led to forest fires as a result of poor forest management (Marraco Solana, 1991), reflected by the CHAR results since cal. yr 1950. A pronounced increase in CHAR > 125 µm occurred roughly the 1990s, indicating local fires near the study area.
An ambitious reforestation was implemented until the 1970s, covering 3 million hectares. This process, for short-term economic return, committed fundamental mistakes in terms of the species of trees selected as a result of contradictory policies in the forest administration (Marraco Solana, 1991). This resulted in a significant rise of Pinus pinea/halepensis (30.9%) and even P. sylvestris (11.8%; Table 2).
Later on, following the implementation of conservation measures since mid-20th century, fragmented populations of A. pinsapo experienced an expansion and densification (Figure 3) from scattered remaining stands (Linares and Carreira, 2009).
Particularly noteworthy was the remarkable expansion of Q. ilex at the top of our sample, probably helped by the dry and warm climatic conditions mentioned above and the ending of charcoal making. Currently, the interspecific competition between A. pinsapo and Q. ilex at lower elevations is discussed by López-Tirado and Hidalgo (2014) in Sierra las Nieves. It appears that both long-term anthropization of Abies mixed-forest jointly to global warming have favored the expansion of Fagaceae versus Abies spp. (e.g. López-Merino et al., 2008). Low elevation A. pinsapo stands will be probably the most sensitive ones to future climate change (Linares et al., 2010; Navarro-Cerrillo et al., 2014), where according to this study other tree species better adapted to drought, such as Q. ilex, might lead to a shifting community, from pure fir forest to mixed stands.
Toward the end of the 20th century, most of the A. pinsapo forests were included in the so-called ‘Network of Protected Areas in Andalusia’: Sierra de Grazalema Natural Park (1984), Sierra de las Nieves Natural Park (1989), and Los Reales de Sierra Bermeja Nature Area (1989). Later, the two first forests were declared Biosphere Reserve by UNESCO in 1977 and 1995, respectively. And finally, both populations were included in the Habitats Directive (92/43/CEE). An action program for A. pinsapo conservation has been established during the last years and now conservation efforts focus on the development of fire prevention plans, pest control during severe drought, and experimental reforestations. The possibility of undertaking silvicultural treatment is also being considered for some areas, to reduce competition among individual trees (García Esteban et al., 2010; IUCN Red List: www.iucnredlist.org).
The coming future: Toward adaptive forest management under a global change scenario
Our results enhance the interest of the paleorecord within global change ecology, by providing well-documented instances of species and community responses to environmental and land-use changes at reliable spatial and temporal scopes. Hence, we conclude that the conservation status and forest structure of A. pinsapo forest rely to some extent on its previous history, where several driving factors, both human and climate induced, have left noticeable footprints. Our results suggest that relict A. pinsapo forests will probably undergo changes beyond the late Holocene recorded levels of community turnover and range shifts, because the magnitude of climate anomalies expected in the future are greater than those observed during the period studied here. Some A. pinsapo populations, like the remnants located at higher elevation, may experience a positive response to climate change in the future, while low elevation (González-Hernández et al., 2015) or high-density stands will likely be more sensitive (Linares et al., 2010). Here we find, in addition, that long-term legacies could be present as a contributing factor interacting with future climate change.
In view of the climate transition over the last centuries, the oncoming A. pinsapo forests dynamics might be different from those reported here during the late Holocene, as shown by the latest investigations in the current forest stands. As regards, the course involved in this recent large-scale forest decline, drought-induced tree mortality drives the functional and structural dynamics of forest ecosystems, both as a sink/source of carbon and nutrients, and as a mechanism of change in forest structure and species diversity. However, managing stand structure, including sizes diversity and tree spatial patterns, may boost the adaptive capacity of drought-sensitive tree species (Lechuga et al., 2017).
Conclusion
Our data show that the increasing human pressure has triggered the steady decline of A. pinsapo in the studied area. The intensification of agrarian practices since mid-15th century coupled with climate seasonality, as well as increasing aridity during LIA (a time of successive short and abrupt dry phases), and RW (a rising aridity period) were the main drivers.
After Islamic period, characterized by a sustainable management of A. pinsapo forest, with low human-impact and a more stable forest structure than today, an unsustainable land-use has been the common practice of successive societies. Historical land-use (clearing for shipping building, mining, charcoal, livestock, extensive agriculture, erroneous afforestation process, etc.) could have influenced pinsapo fir forest biodiversity, leading to a decreased structural complexity and homogenization, and finally to a loss of resilience within A. pinsapo forest to face global warming.
Although the future of A. pinsapo forests will be constrained by climate change, the primary driving factor will likely be the forest and land management types chosen to implement in situ. Re-establishing low impact traditional uses (low intensity grazing or thinning, etc.) could promote, in some cases, the development of a more complex forest structure, appropriate arboreal density, and higher biodiversity of forest species.
Footnotes
Acknowledgements
We are grateful to Consejería de Medio Ambiente (Andalusian Government) and José López Quintanilla for providing sampling permissions and access to the study area. Thanks to Andrés Perez-Latorre (UMA) for reviewing the manuscript. The authors thank the two anonymous reviewers for improving the final version with their constructive comments.
Funding
This study was supported by Research Program from the Andalusian Government: P11-RNM-7033 project ‘Paleobiogeografía y conservación de Abies pinsapo Boiss. y Cedrus atlantica (Endl.) Carrière: dos especies de la flora relicta del Mediterráneo occidental (RELIC-FLORA)’.