
Abstract
The aim of this work is to reconstruct the periods of growth and decline of human populations in Morocco and their potential impacts on the landscape over the past 10,000 years. In order to estimate the trends in the human population size between 10,000 and 3000 years ago, we used a summed probability distribution (SPD) of radiocarbon dates from a wide range of archaeological sites throughout Morocco. Landscape changes were identified and quantified from a dataset of fossil pollen records. Different anthropogenic pollen markers, as well as natural vegetation groups and taxonomic richness were used to analyse the relationship between long-term trends in human population expansion or regression and type of impact on the landscape. The sub-regions of Morocco have different topographies and climates, which have either favoured or prevented the establishment and/or spread of human populations. In order to identify the areas most significantly impacted by humans and the timing of such impacts, we have reconstructed and compared the same past anthropogenic and landscape proxies along with the population trends within the lowlands and mountainous areas. The lowlands were more strongly impacted earlier in the Holocene than the mountainous areas. Anthropogenic markers indicate that farming expanded in the lowlands during the first major expansion of human populations between ca. 7200 and 6700 cal. yr BP at the start of the Neolithic period. In the Atlas and Rif Mountains, anthropogenic impact is not clearly detectable in any of these areas before 4000 cal. BP.
Introduction
Humans have been present in the northwest corner of Africa throughout the Quaternary. For example, fossil remains of Homo sapiens more than 300,000 years old were recently discovered in Jebel Irhoud, near the town of Safi in Morocco (Hublin et al., 2017). This time span, which encompasses the latter part of the Palaeolithic period, witnessed three climatic cycles with marked interglacials that were similar to the Holocene, and long glacial periods (e.g. Hays et al., 1976). During the most recent glacial period, around 20,000 years BP, glaciers developed even at low latitudes, such as within the Mediterranean (Hughes et al., 2006), including North Africa, at elevations as low as 2000 m a.s.l. (Hughes et al., 2011, 2018). As a consequence, plant and animal species were naturally constrained, which reduced their range substantially and allowed them to persist only in refugial areas (e.g. Hewitt, 2000). Likewise, humans would have adapted their population size and habitat locations in response to past climate fluctuations (Roberts et al., 2018; Van de Loosdrecht et al., 2018).
The recolonization of new suitable habitats by humans after the last glacial period from scattered populations was probably neither synchronous throughout the Mediterranean nor continuous and homogeneous during the Holocene warm period (Hajar et al., 2010; Mercuri and Sadori, 2012). This may explain the asynchronous dating of the beginning and end of the Neolithic in the Mediterranean (Linstädter et al., 2018; Morales et al., 2013). In terms of impact on the landscape, as soon as human populations began to settle and/or to spread in the Mediterranean basin they left clear imprints of their activities (cultivation, fire, domestication, clearing, use of tools, etc.) directly in the areas they occupied and indirectly in the fossil records preserved in wetlands sediment archives and lakes. Most Holocene fossil records tend to show that there were natural changes during the first 1000 years of the Holocene when climate mainly forced ecosystem changes, which was followed by complex interactions with increasing human interference. Some authors have proposed that climate was the main driver of synchronous ecosystem changes in the Mediterranean during the entire Holocene and that landscape changes cannot be attributed to human activity alone (Jalut et al., 2009). Other scholars have argued that there is an interplay between climate, humans and Mediterranean ecosystems, which becomes complex to unravel when aridity increased during and after the mid-Holocene (Carrión et al., 2010; Mercuri, 2008; Sadori et al., 2011).
In Morocco, the earliest Holocene human use of plant resources was detected in the semi-arid lowlands of the northeast Moroccan hinterland. Charcoal samples from rock shelters (Görsdorf and Eiwanger, 1999) and Epipalaeolithic open-air sites (Ibouhouten et al., 2010; Linstädter et al., 2012; Mikdad et al., 2000) provide 14C ages between 11,700 and 7800 cal. yr BP. In the Middle Atlas Mountains, there are indications of early Holocene occupation during the Epipalaeolithic with chronological evidence dating to around 8400 cal. BP from Ouberid cave (Mikdad et al., 2012). However, the ecological impact of Epipalaeolithic hunter-gatherers on the early Holocene landscape was minor, therefore, this early occupation is not clearly reflected in secondary environmental archives. The first evidence for human impact on vegetation cover is provided by recent and precise archaeobotanical studies that have dated the onset of early Neolithic occupation in northern Morocco, close to the Mediterranean Sea, between 7700 and 7200 cal. yr BP (Linstädter et al., 2016; Zapata et al., 2013).
Indirect evidence from fossil pollen records suggests that human impact has strongly increased over the past 4000 years in Morocco and became increasingly apparent at both low and high elevations (Campbell et al., 2017; Cheddadi et al., 2015; Lamb et al., 1991). In addition to fossil pollen records, model simulations show that during the historic period, forest cover on usable land may have dropped dramatically from an estimated 98% in 3000 BP to 31.7% in AD 1850 (Kaplan et al., 2009), in relation to the expansion of the human population. In Morocco, cedar forest cover in the Rif mountains decreased by about 75% between 1960 and 2010 (Cheddadi et al., 2017).
Human activities can be identified in the fossil pollen records through the occurrence and abundance of those taxa that are considered anthropogenic indicators (Behre, 1981; Mercuri et al., 2011). Changes through time in the human population size can also be estimated from the summed probability distributions (SPD) of archaeological (e.g. anthropogenic) radiocarbon dates (Crema et al., 2016; Gamble et al., 2004; Palmisano et al., 2017, this volume; Shennan et al., 2013; Timpson et al., 2014; Weninger et al., 2009; Williams, 2012; Zielhofer et al., 2008).
In this study, we estimated the size of the human population in Morocco for the time period between 10,000 and 3000 cal. BP and compared the population variation during the Holocene with several anthropogenic pollen indicators as well as with reconstructed natural vegetation groups and past taxonomic richness derived from an extensive dataset of fossil pollen records.
Morocco has a wide range of topographies with the highest elevations ranging from 2500 in the Rif mountain chain up to more than 4000 m a.s.l. in the High Atlas. There are also large coastal lands and plains, which are intensively cultivated today. These complex topographic features have almost certainly constrained the spread and settlement of humans throughout the Moroccan landscape. The main objectives of this study are threefold: (1) to reconstruct the overall human demographic changes in Morocco during the later prehistory, (2) to identify different past human activities in different areas using a set of anthropogenic markers from a dataset of fossil pollen records and (3) to evaluate the spread of human activities and their impacts both on the lowlands and on the mountain landscapes.
Materials and methods
Anthropogenic markers, natural vegetation groups and taxonomic richness
The pollen dataset used in this study includes 22 records from different areas in Morocco, obtained from both original authors and digitized published work (Figure 1a, Table 1). There are two sites, Lake Hachlaf and Lake Sidi Ali, from which two records were collected from each lake. We integrated these duplicated records because they encompass different time periods. The Moroccan pollen records were produced by various analysts over a ca. 40-year period, between 1976 and 2017 and have been dated relatively accurately by conventional and AMS radiocarbon dating. Some of the oldest pollen records (Reille, 1976, 1977, 1979) have been dated using very few radiocarbon dates. In three pollen records (Marzine in the Rif, Tessaout and Tizi Inouzane in the High Atlas) the published chronologies were based on just one 14C date (Reille, 1976, 1977), while one record (Iguerda-Ait-Amama in the Middle Atlas) has not been dated by any radiometric methods (Reille, 1976). The age/depth models that we have built for these four pollen records are based on the published original author’s assumptions based on stratigraphic markers and expertise. The time spans proposed by the original author (Atlantic, Sub-Boreal and Sub-Atlantic periods) have been used to build a quantitative chronology for the four pollen records.
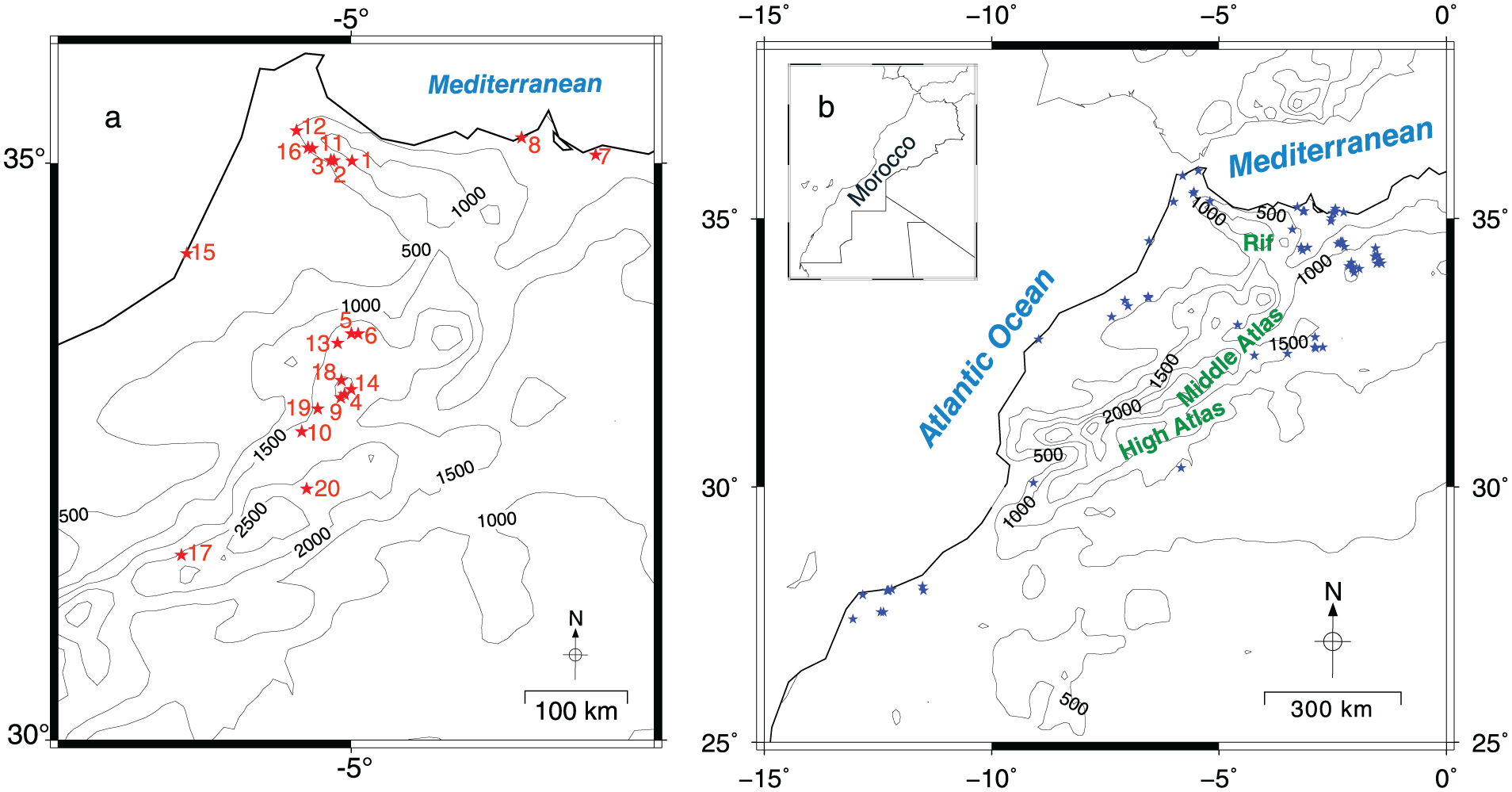
(a) Location of the fossil pollen records (red stars) used for reconstructing anthropogenic markers, vegetation groups and pollen taxonomic richness. Pollen record numbers are those referred in Table 1. (b) Location of the archaeological sites (blue stars) from which we have obtained 14C measurements for evaluating past changes in the human population in Morocco (SPD).
Pollen records used in this study (displayed in Figure 1a). All pollen records from Reille (1976, 1977, 1979) have been digitized from the original published pollen diagrams and the raw data were computed using the pollen sums.
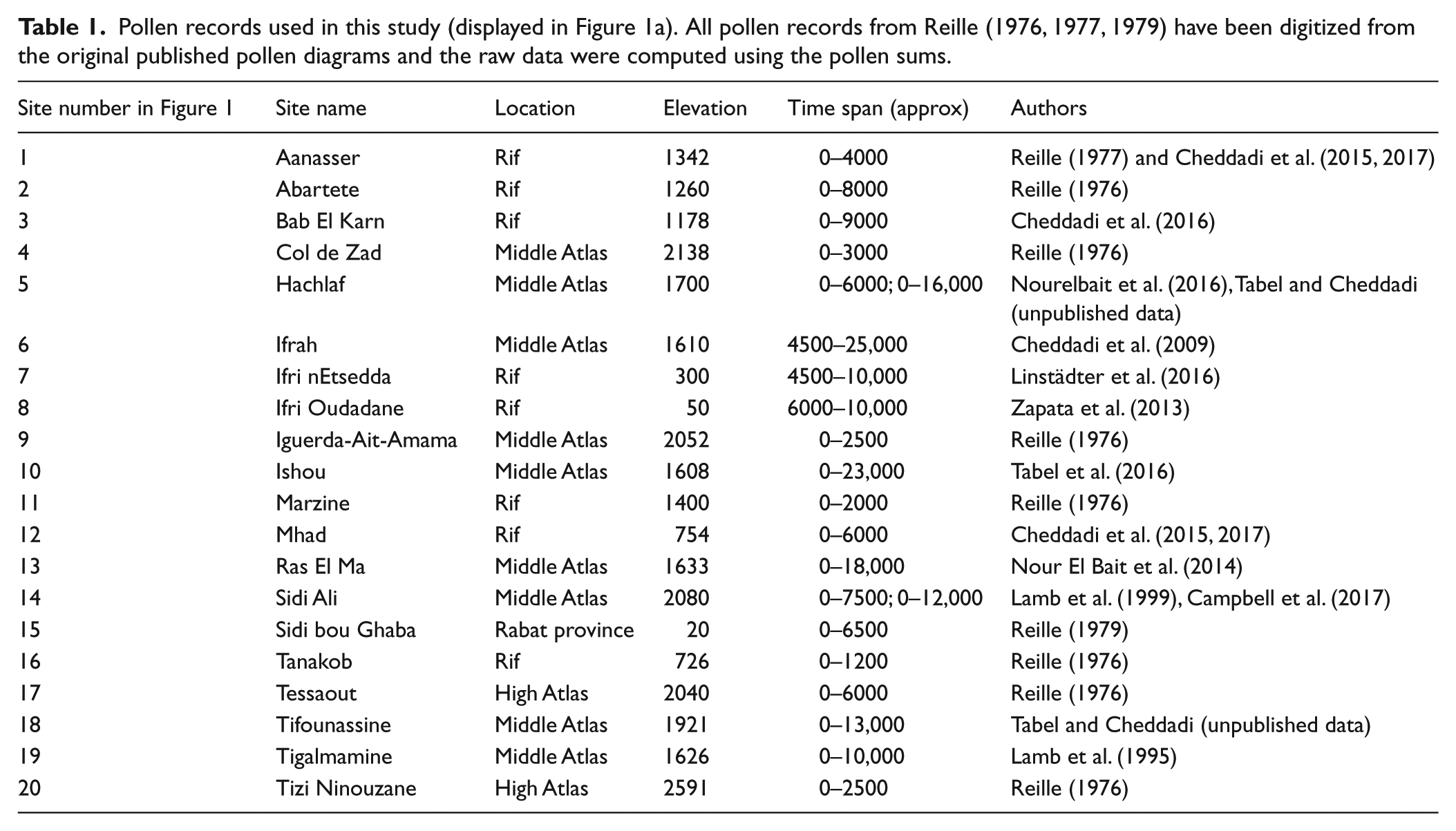
Within the dataset, there are five pollen records located at elevations lower than 800 m a.s.l. Two of them encompass the early Holocene and three others only cover the mid–late Holocene (after 6500 BP). Thus, in the dataset used in this study, there are more pollen records that encompass the entire Holocene period in the mountains than in the lowlands. Such bias in the duration and spatial distribution of the pollen records in Morocco must be taken into account when interpreting the occurrence of anthropogenic markers and overall vegetation changes. All of the pollen records compiled have been archived in a MySQL database, which has a structure that is compatible with the European Pollen Database (www.europeanpollendatabase.net). The published Moroccan pollen data will be contributed to the European Pollen Database (Leydet, 2007–2018).
We defined five anthropogenic pollen markers (APMs) from the taxa identified in the fossil pollen records (Table 2). These APMs are based on earlier published research and are considered indicative of human activities (Behre, 1981; Mercuri, 2008; Mercuri et al., 2011, 2013a, 2013b; Sadori et al., 2011). The reconstructed APMs include the following:
Anthropogenic pollen index (API)
Regional pastoral indicators (RPI)
Anthropogenic nitrophilous herbs (ANH)
Cultures and crops (CC)
Olea-Juglans-Castanea-Vitis (OJCV)
Taxa used to identify different human impacts within the Moroccan pollen records.

API = Anthropogenic Pollen Index; RPI = regional pastoral indicators; ANH = Anthropogenic nitrophilous herbs; CC = cultures and crops; OJCV: Olea-Juglans-Castanea-Vitis.
We selected these five APMs to allow comparisons with a selection of similar studies based on six other regions spanning the Mediterranean and a Mediterranean-wide synthesis of the Holocene population and landscape change (see Bevan et al., this volume; Roberts et al., this volume). In Morocco, these APMs were reconstructed for three separate regions: the Rif Mountains, the Atlas Mountains and lowland sites (located at an elevation lower than 800 m a.s.l.). The three sub-regions were then amalgamated and reconstructions were produced for the entire country as one overall entity that takes all of the pollen records into account (Figure 2). One palynological difference with these similar studies carried out in other Mediterranean regions is the exclusion of Artemisia from the APMs, because it is a natural dominant taxon in Moroccan steppe landscapes which occurs in the mountainous areas (e.g. Saadi and Bernard, 1991). Thus, Artemisia was not considered an anthropogenic marker in this study. In addition to the APMs, we reconstructed the pollen taxonomic richness (Figure 3) using rarefaction analysis (Birks and Line, 1992) for the same areas and pollen records as the APMs. Fossil pollen samples represent a partial representation of the anemophilous plants, because of the differing pollen productivity of different plants and their dissimilar dispersal capacity. The number of identified and counted pollen grains is also often, if not always, different from one analysed sample to another within the same record. Rarefaction analysis provides an unbiased estimate of the number of taxa in a fossil sample, which allows a comparison of the pollen analyses from different samples in the same record (Birks and Line, 1992). However, one should keep in mind that the pollen taxonomic richness does not represent a measure of the species diversity, sensu Shannon or Simpson indices, as one pollen taxon may correspond to one or several species and the pollen percentage in a fossil sample does not correspond to the number of occurrences of the species in the studied site. Modern human activities often result in a negative impact on species diversity. Pollen taxonomic richness is not a direct indicator of past human disturbances, but it may help comprehend whether the past human demographic changes had a negative or a positive impact on species diversity.

Percentages of anthropogenic pollen markers (API, RPI, ANH, CC and OJCV) and arboreal pollen taxa (AP) during the Holocene in the Atlas mountains (red), the Rif mountains (green), the lowlands (sites below 800 m a.s.l., blue) and Morocco (black, including all pollen records).

Estimated pollen taxonomic richness using rarefaction analysis from the lowland sites (below 800 m a.s.l.), the Rif and Atlas Mountains, and all datasets from Morocco.
In addition to the APMs and pollen taxonomic richness, we also used clusters of pollen taxa to identify four natural vegetation groups (Figure 4) that represent the main ecosystem types in Morocco (Quézel and Médail, 2003). These natural vegetation groups are ordered by increasing elevation: (1) evergreen trees and shrubs, (2) deciduous trees, (3) mountain conifers and (4) steppe (Table 3). This is based on grouping pollen samples into ‘vegetation clusters’ according to their taxa assemblages and builds on the cluster analysis approach used by Woodbridge et al. (2018) and Fyfe et al. (2018).
Pollen percentages of four natural vegetation groups.
Pollen taxa grouped in the four natural vegetation groups.

Demographic change over the Holocene
In the past two decades, one of the most popular proxies for inferring demographic trends in the prehistoric period has involved SPDs of archaeological radiocarbon dates as a result of increasingly sophisticated methods (Balsera et al., 2015; Bevan et al., 2017; Bocquet-Appel et al., 2009; Capuzzo et al., 2018; Crema et al., 2016; Palmisano et al., 2017; Rick, 1987; Shennan and Edinborough, 2007; Shennan et al., 2013; Timpson et al., 2014). The SPD results from ‘counting up’ (summed in the manner of a histogram) the calibrated raw radiocarbon years of each organic sample, which are expressed as probability statements with error ranges. This is based on the assumption that the more people living in a given region, the more the archaeological remains, the more the organic materials, and the more the samples that can be collected for radiocarbon dating (Rick, 1987). Such indicators do not offer good evidence for absolute numbers of a human population but rather give an idea of the relative intensities of population and proportional change through time. Although SPDs of radiocarbon dates have been extensively used by archaeologists for modelling population fluctuations in prehistory, they face several challenges such as biases in research strategies, budgets and interests that can undermine a random sample of human activity in every archaeological phase.
Over the past 10 years, both the number and the accuracy of radiocarbon dates have increased in most newly investigated archaeological sites. This is the case in Morocco, where more than two-thirds of the radiocarbon dates used in this study were published in the past decade. In this study, we have estimated the human population size in Morocco from 270 uncalibrated radiocarbon dates, which have been collected from 83 archaeological sites (Figure 1b) between 10,000 and 3000 cal. BP. To our knowledge, the dataset used in this work represents the largest existing collation of archaeological radiocarbon data for Morocco. This number of dates collected (n = 270) exceeds the suggested minimum threshold of 200–500 dates to produce a reliable SPD with reduced statistical fluctuation for a time interval of 10,000 years; specific issues about sample size are discussed below (Michczyńska et al., 2007; Michczyńska and Pazdur, 2004; Williams, 2012: 580–581). We are aware of the limitations of our dataset and the results are a preliminary attempt to reconstruct demographic change given the spatially restricted archaeological data available in the area.
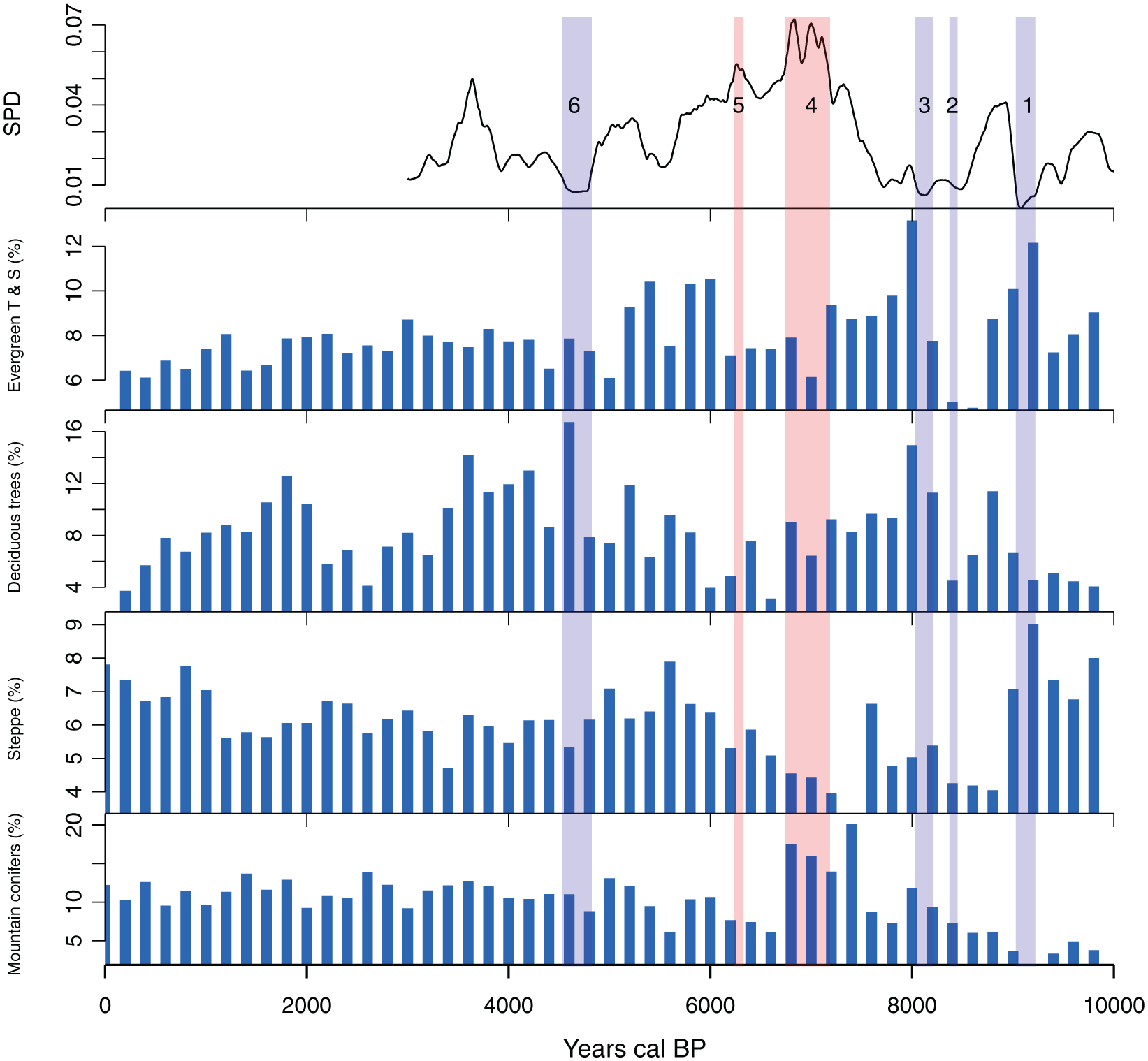
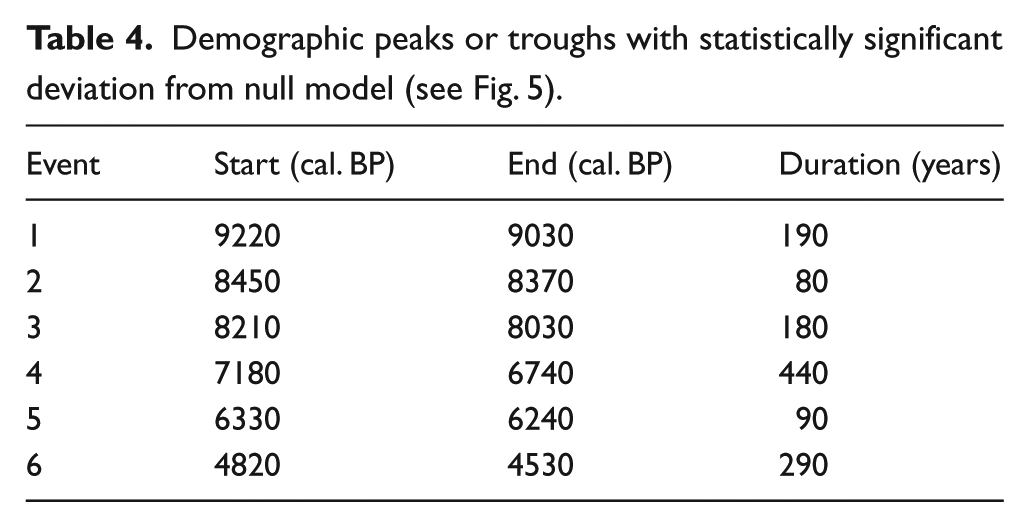
All of the radiocarbon dates are from archaeological contexts, with the majority based on samples of bone, charcoal and wood. Radiocarbon dates obtained from marine sources, such as shells, have been removed (and are not part of the above total) to avoid the complicated issues arising from unknown or poorly understood marine reservoir offsets. The probabilities from each calibrated date have been combined to produce an SPD (Figure 5). 1 The potential bias of oversampling particular site phases has been reduced by aggregating multiple uncalibrated radiocarbon dates from the same site that are within 100 years of each other and dividing by the number of dates that fall within this time ‘bin’ (see Timpson et al., 2014). Following this process, the probabilities of each bin are summed: in our case, 270 radiocarbon dates have been aggregated into 207 bins. Following previous works (Weninger et al., 2015; Williams, 2012), which show that normalized calibrated dates emphasize narrow artificial peaks in SPDs because of steepening portions of the radiocarbon calibration curve, we opted to use unnormalized dates prior to summation and calibrated via the IntCal13 curve (Reimer et al., 2013; see former applications of calibrated unnormalized radiocarbon dates in (Bevan et al., 2017; Palmisano et al., 2017; Roberts et al., 2018). A logistic null model representing the expected population increase has been fitted to the observed SPD in order to produce a 95% confidence envelope (composed of 1000 random SPDs) and statistically tested to see if the observed pattern significantly departs from this model (for the general approach see Shennan et al., 2013; Timpson et al., 2014; as specifically implemented in Bevan and Crema, 2018: modelTest, ‘uncalsample’). Deviations above and below the 95% confidence limits of the envelope, respectively, indicate periods of population growth and decline greater than expected according to a logistic model of population growth. However, it is important to recognize that a logistic model cannot strictly be considered as a realistic model for population growth, but rather as an elementary model useful for quantitatively testing population fluctuations (cf. Turchin, 2001). In this case, a logistic model was selected as a more suitable option as opposed to other possible null models (e.g. uniform, exponential), given the observed shape of radiocarbon date SPDs in our study area (see Figure 5).

Summed probability distribution (SPD) of unnormalized calibrated radiocarbon dates versus a fitted logistic null model (95% confidence grey envelope). Blue and red bands indicate that chronological ranges within the observed SPD deviate negatively and positively from the null model and correspond to four significant decreases in human population (events 1, 2, 3 and 6) and two significant increases (events 4 and 5).
Demographic peaks or troughs with statistically significant deviation from null model (see Fig. 5).
The Epipalaeolithic and Neolithic periods have been defined according to Linstädter et al. (2018) in northern Morocco. As proxies for past population estimates from the SPDs of radiocarbon dates can be used as recently as ~3000 cal. yr BP in Morocco, that is, prior to the historical time period when archaeological chronologies rely more on specific evidence such as datable coins, written source and fine-ware pottery rather than on radiometric dating.
Chemical elements
A chemical elements analysis was carried out, using portable x-ray fluorescence (XRF) technique, on a sediment core retrieved in Ait Ichou swamp in the south of the Middle Atlas (Tabel et al., 2016). XRF analyses allowed the variation of more than 20 chemical elements to be measured over the past 25,000 years covered by the Ait Ichou core. However, only four chemical elements were used in this study (Figure 6). These elements are iron (Fe), lead (Pb), zinc (Zn) and copper (Cu). These elements were selected as indicators of human activity and a marked increase in their concentration in the sedimentary record is related to different human activities in the area.

X-ray measurements of iron (Fe), copper (Cu), lead (Pb) and zinc (Zn) in the Ait Ichou fossil record collected in the Middle Atlas (Tabel et al., 2016).
Results
The compiled fossil pollen dataset (Figure 1a) and archaeological radiocarbon dates (Figure 1b) allow us to reconstruct several anthropogenic markers (Figure 2), past plant taxonomic richness (Figure 3), natural composition of past ecosystems (Figure 4) as well as human demography changes (Figure 5) during the Holocene. The anthropogenic pollen markers allow us to identify the potential relationships between natural ecosystems and superimposed human disturbances. A spatial and temporal analysis of these pollen markers may provide information on the timing of human interference throughout the Holocene and the types of impacts. The proportions of the reconstructed anthropogenic markers correlate with the estimated human demography, which may provide information about the intensity of human impacts on the landscape.
The archaeo-demographic results (Figure 5) show some significant overall departures from the SPD of observed data from the envelope of the logistic model (p = 0.006), which indicates that the population did not grow logistically from 10,000 to 3000 BP in Morocco. The population was greater than that predicted by the logistic null model in the mid-Holocene between ~7200–6700 and ~6300–6200 cal. BP. In contrast, the population was significantly below the expected values in the early Holocene (~9200–9000, ~8450–8370 and ~8200–8000 cal. BP). Another marked decline in population occurs during 4800–4500 cal. BP. Generally, the level of population was lower in the early Holocene and started increasing substantially with the onset of the Neolithic in the 8th millennium cal. BP. The duration of these periods of decreasing and increasing human population differs and varies between ca. one and ca. four centuries (Figure 5).
The anthropogenic pollen markers API, ANH and CC show a marked increase (Figure 2) that matches the first significant expansion of human populations between 7180 and 6740 cal. BP (event 4 in Figure 5). This first increase in pollen indicators of human activity is recorded mainly in the lowland sites rather than in the Rif and Atlas Mountains. The arboreal pollen percentages decrease most significantly within sites located at a lower altitude. Human demographic changes (Figure 5) and the three APMs (API, ANH and CC) from the lowland sites are positively correlated (Table 5). Pollen taxonomic richness shows a significant correlation with SPDs (Table 5) with a marked increase between 7180 and 6740 and then between 6330 and 6240 when the human population also increased (events 4 and 5). This correlation is more significant in sites located in the lowlands than in the Rif and Atlas Mountains (Figure 3). Periods of decreasing human population (events 1, 2, 3 and 6, Figure 5) are not marked in the lowlands or in the mountains. In order to evaluate the impact of human demographic changes on natural ecosystems, we have reconstructed four groups of vegetation (Figure 4, Table 3), which are well defined in Morocco (Quézel and Médail, 2003) and pollen grains representing these plant species have been identified in the fossil records. These vegetation groups include montane conifers, deciduous trees, evergreen trees and shrubs and steppe plants (Figure 4). None of these groups shows a significant correlation with human demographic changes during the Holocene (Table 5) and none of the human population increases (event 4 and 5) or decreases (events 1, 2, 3 and 6) clearly reflect changes in any vegetation group.
Spearman’s correlation coefficients between the pollen percentages of different anthropogenic markers (ANH, API, CC, OJCV, RPI), vegetation groups and arboreal pollen (AP) from fossil pollen records and archaeo-demographic datasets (SPD) by region and elevation for the period 10,000–3000 cal. BP.
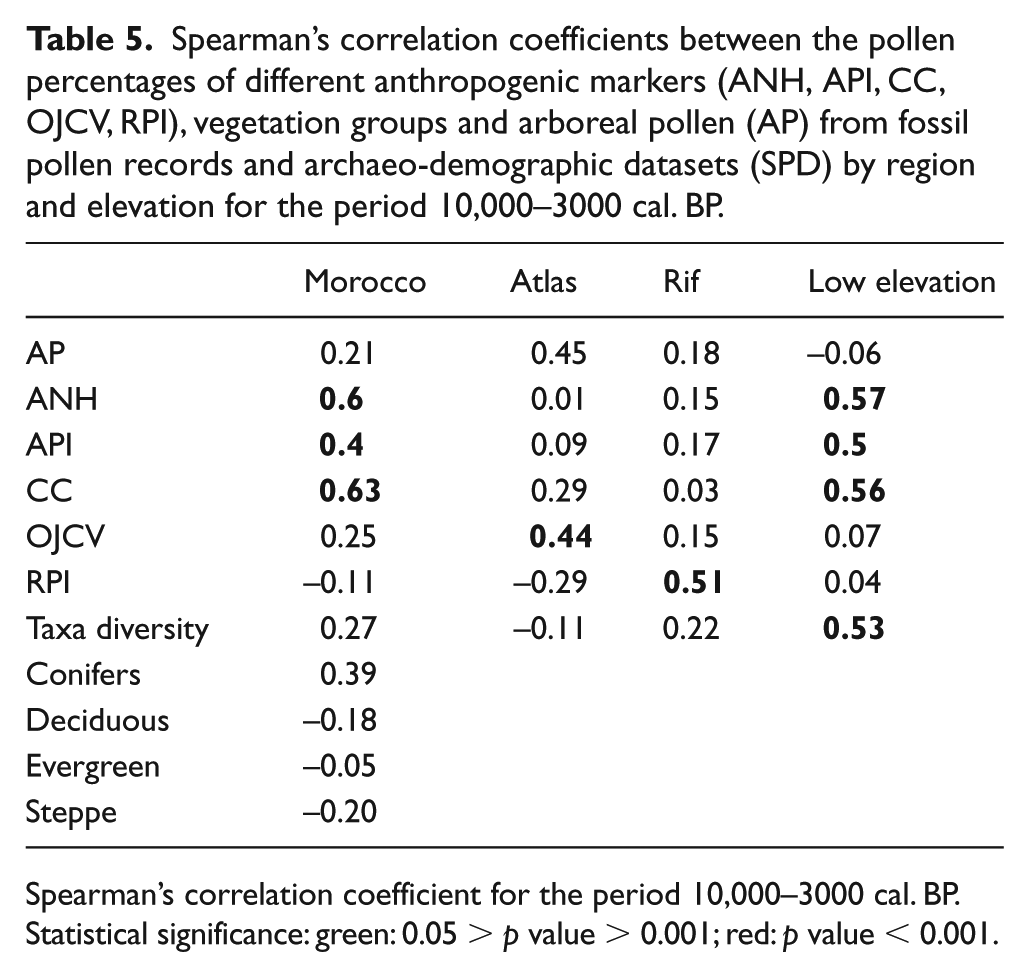
Spearman’s correlation coefficient for the period 10,000–3000 cal. BP.
Statistical significance: green: 0.05 > p value > 0.001; red: p value < 0.001.
Discussion
Modern challenges facing the Mediterranean region in terms of managing the impacts of both climate change and human population growth are intensifying. Human pressures on the Mediterranean biodiversity have steadily increased over the past century and have reached a critical threshold within the past few decades. Understanding of past relationships between human demography and ecosystem changes is paramount to managing ongoing landscape changes, and requires studies integrating longer timescales than in the past few decades.
Morocco is within the Mediterranean floristic area, which is considered a hotspot of biodiversity (Myers et al., 2000) with approximately 22% of endemic vascular plants (Rankou et al., 2013). Owing to the geographical expansion of the human population and related activities, several species have become extinct over the past century and many species are endangered or in threat of extinction today (IUCN, 2018). Forest cover in Morocco has been decreasing steadily and substantially over the past century (Kaplan et al., 2009) and all ecosystems, from the seashore to the highest elevation mountains, have been impacted by expanding human activities and increasing exploitation of ecosystem resources. For example, cedar forest cover in the Rif Mountains has decreased by about 75% over the past 50 years, from more than 45k ha to ca. 10k ha today (Cheddadi et al., 2017). Simultaneously, the human population has more than quintupled between 1900 and 2014, from ca. 6 to ca. 34 million inhabitants. In this context, today more than ever, it is important to analyse and understand past impacts of human demographic change on different ecosystem types. The Neolithic is a very interesting period for exploring the impact of early human expansions and regressions, on landscapes and the increasingly complex ways in which such activities are superimposed on climate change records. This may help to evaluate the ecosystem’s capacity for adaptation and resilience to the combined effects of natural and human-induced changes.
Evaluation of human demography during the Neolithic can be performed using radiocarbon dates available from archaeological sites and ad hoc statistical tools (Crema et al., 2016; Gamble et al., 2004; Palmisano et al., 2017; Shennan et al., 2013; Williams, 2012). However, one should be cautious as archaeological sites are often not exhaustively studied and there may be important differences in the number of dates available at each site. This is clearly the case for Morocco, where the number of 14C dates used in this study could certainly be improved with additional archaeological sites and more 14C dates per site. Other additional potential biases related to 14C date measurements and their calibration may also introduce some errors in estimating the human demographic trends using SPDs (Shennan et al., 2013), as well as the duration of the expansion/regression of human populations (Manning et al., 2014). Used as a demographic proxy, SPDs may reflect only a local (or regional) expansion of the human population rather than representing a spatially large spread (Shennan et al., 2013).
There is a large literature focused on human expansion and related activities in the Mediterranean basin during the Neolithic (summarized in Shennan, 2018). However, the timing of expansion, and the type and intensity of the impacts are not synchronous and probably not comparable throughout the Mediterranean region. Today, the human population in Morocco is composed mainly of Berbers (autochthonous population) and Arabs. Genetic analyses of North-Western African populations reveal that lineages of different Berber groups may date back to at least the last glacial period and that the omnipresence of a certain mitochondrial DNA motif suggests a continuous presence of these populations in Morocco over more than 20,000 years (Rando et al., 1998). All recent genetic studies agree that the spread of human populations in North Africa originated from the East–West (Bentayebi et al., 2014), rather than originating from sub-Saharan populations (Desanges, 1981). The genetic contribution of sub-Saharan populations to the modern North African populations seems to be minor (Bosch et al., 1997; Brakez et al., 2001). Recent archaeological findings in northern Morocco indicate the absence of Saharan influences during the early Neolithic until 6.0 cal. ka BP (Linstädter et al., 2018). Thus, even if the timing, continuity and degree of expansion of the migrating original populations are still under scientific debate, the Eastern origin of the modern North African populations is now genetically proven. Rando et al. (1998) state that the modern dominating lineages arrived in North Africa during the Mesolithic and Neolithic in waves, while Arredi et al. (2004) propose that the Neolithic transition in North Africa was accompanied by demic diffusion (see Cavalli-Sforza et al., 1993). The marked variations in 14C date SPDs (Figure 5) and the discontinuous occurrences of the fossil anthropogenic markers (Figure 2) suggest that the human population in Morocco did not increase steadily during the Holocene, but involved marked periods of ‘booms’ and ‘busts’ that impacted upon the landscape intermittently. These past demographic variations (Figure 5) and discontinuous landscape changes could probably have resulted from waves of immigrating populations rather than demic diffusion into Morocco during the Holocene.
The SPD data suggest that human demography fluctuated during the Holocene with two periods of noticeable population increase and four others with noticeable population decrease (Figure 5). The human population increased substantially with the onset of the Atlantic Neolithic around 7400/7300 cal. BP. In agreement with the SPD data, pollen markers of cultures and crops and farming increased in sites located at low elevation. The correlation between the anthropogenic markers and SPD (Table 4) suggests that the human impact was not only local or regional, but probably took place over a larger area. However, this positive correlation is based on only five lowland records, which includes two Holocene archaeological sites (Figure 1a, Table 1) and three pollen records that encompass the second half of the Holocene (younger than 6500 cal. BP). To confirm that human impact in the lowlands was spatially more extended would require additional Holocene data from off-site contexts, such as lake sediments at low elevation. The pollen records available in the Rif and Atlas Mountains are more numerous, well dated and many of them encompass the entire Holocene (Table 1). In these montane records we do not observe a significant correlation between the APMs and SPDs (Table 4), which supports the interpretation that human imprints were probably restricted to the lowland areas during the early Neolithic. Archaeological findings in the Moroccan coastal areas and lowlands confirm the presence of cultivated landscapes as early as ca. 7000 cal. BP (Ballouche and Marinval, 2003; Linstädter et al., 2016; López-Sáez et al., 2013; López-Sáez and López-Merino, 2008; Morales et al., 2013; Zapata et al., 2013). At higher elevations, several palaeoecological studies indicate late-Holocene human impacts on ecosystems in the Atlas and the Rif Mountains (Abel-Schaad et al., 2018; Campbell et al., 2017; Cheddadi et al., 2015; Lamb et al., 1991; Reille, 1977; Zielhofer et al., 2017). The low correlation between forest ecosystems (Figure 4) and SPDs, which occurs mainly in the mountain areas (Table 4), suggests a lower level of human impact at higher elevations. Inhabitants of the Moroccan mountains may have included populations of hunter-gatherers rather than farmers, which could have delayed the expansion of cultivation and food production ‘technologies’ (Bosch et al., 1997) and, therefore, may explain the absence or low level of human imprints during the early Neolithic in the Rif and Atlas Mountains. Prior to the first period of significant human expansion (ca. 7400 cal. BP), there are high values of OJCV (>10%) that are dominated by Olea pollen percentages, which may be interpreted as related to early domestication of the olive tree in Morocco. However, these high Olea occurrences are recorded during a time span of a significantly lower human population (Figure 2). The increase in Olea pollen percentages during the early Holocene in Morocco likely corresponds to the spread of wild stands (oleaster) under a warmer early Holocene climate (Cheddadi et al., 1998) rather than to early domestication of the olive tree (see Langgut et al., this volume).
Reconstructed pollen taxonomic richness (Figure 3) is not well correlated with human demographic changes either in the lowlands or in the mountainous sites (Table 4, Figure 3), which suggests that either human demographic fluctuations had a minor impact on the structure and composition of the ecosystems or that such ecosystems are highly resilient. This reflects the characteristics of modern Mediterranean ecosystems, which are considered highly resilient to human disturbances because of their high ecological diversity (Lavorel, 1999; Pausas et al., 2008).
After the first major increase in the human population (7180–6740 cal. BP) we observe a quasi-steady decreasing trend, which reached a noticeable SPD minimum between ca. 4820 and ca. 4530 cal. BP (Figure 5). The anthropogenic markers (API, ANH and CC) also decreased in the lowlands and remained low in the Rif and Atlas Mountains (Figure 2) during this time, which is coherent with the reconstructed decreasing trend in the human population. Within the Rif Mountain, archaeological sites of cereals and anthropogenic herbs decreased between 6700 and 6000 cal. BP, indicating reduced grazing and cultivation activities (Linstädter et al., 2016). The SPD data indicate that human population remained low until 4000 cal. BP, which probably marks the end of the Neolithic in Morocco.
Unlike other parts of the Mediterranean, the metal ages (Bronze, Copper, Iron) are not well dated in Morocco. X-ray fluorescence measurements of the fossil record in the southern part of the Middle Atlas (Tabel et al., 2016) show that iron (Fe), lead (Pb), copper (Cu) and zinc (Zn) started to increase significantly after 4000 cal. BP (Figure 6), which seems to be earlier than in other parts of the Mediterranean (Van Der Plicht et al., 2009), and coherent with earlier archaeological studies in Morocco (Ballouche and Marinval, 2003; Daugas et al., 1998). Chemical elements (Fe, Cu, Zn and Pb) are often associated with human activities during the metal ages (i.e. the Bronze and Iron Ages) and probably mark the beginning of an ‘industrial’ period dedicated to their extraction. The SPD data cover the period between 10,000 and 3000 cal. BP, which does not allow exploration of human demographic changes during the Iron Age. In the northern part of the Middle Atlas, an increase in lead concentration (Pb) in a fossil record (Nour El Bait et al., 2014) started around 2000 cal. BP, which corresponds to the beginning of the Roman presence in Morocco. In the Rif Mountains, the geochemical content of several records show changes similar to those in the Middle Atlas after 2000 cal. BP and are clearly related to Roman industrial activities, which started to impact upon mountain ecosystems, such as through the degradation of the Atlas cedar forests (Cheddadi et al., 2015). The impact of Roman activities in Morocco seems to have been more critical for forest ecosystems with a decrease in arboreal pollen percentages (Figure 2), particularly those of the deciduous and evergreen trees (Figure 4).
It is interesting to note that taxonomic diversity, as detected by pollen records, (Figure 3) was not altered during the Neolithic nor the Bronze/Iron Ages and not even during the Roman period. Today, areas rich in endemic species are threatened by the wide range of human activities, particularly in areas identified as biodiversity hotspots such as the Mediterranean (e.g. Cincotta et al., 2000). Pollen taxonomic richness in Morocco actually shows a relatively steady increase throughout the Holocene and increases more over so throughout the past 3000 years during the metal ages. Elsewhere, several palaeoecological studies have also shown that the last 1000 years of the Holocene are marked by an increase in pollen taxa richness (e.g. Birks and Line, 1992; Lotter, 1998), which is paradoxical to the modern negative impacts of human activities on ecosystems and their species richness, but perhaps consistent with the well-known (but debated) intermediate-disturbance hypothesis (e.g. Fox, 2013). Unlike during the modern industrial era, human activities during earlier periods of the Holocene, which mainly involved livestock grazing and cultivation, were excellent means for the dispersal of seeds, propagation of domesticated plants and the dispersal of ruderal plants that are often subservient to crops.
Our study suggests that there are major differences between past and modern human activities, such as modern artificial reduction of species ranges through the industrial exploitation of forest resources (e.g. Pearson and Dawson, 2003), mono-specific plantations over large areas (Brockerhoff et al., 2008), the introduction of invasive and alien plant species which strongly and negatively disturb the ecosystem composition (Thuiller et al., 2005), the widespread use of herbicides and pesticides and the abruptness of ongoing climate change (http://www.ipcc.ch/) which restricts species ranges. Modern human activities are causing rapid, novel and substantial changes to Earth’s ecosystems (Nolan et al., 2018; Vitousek et al., 1997) that we have not observed in our Holocene records in Morocco.
Conclusion
The past 10,000 years represent an informative time span encompassing the spectrum of natural to anthropogenic forcing, which includes a period of natural climatic changes, with negligible human impact in the early Holocene, followed by a period of interplay between natural and anthropogenic impacts with the expansion of human populations.
The archaeological and environmental data used in this study indicate that prior to 7400 cal. BP, human populations had a limited impact on the lowland landscape and mountain ecosystems. The earliest significant expansion of the human population in Morocco during the Holocene took place around 7000 cal. BP and it is marked in the fossil pollen record by an increase in farming indicators, particularly crop pollen markers. This time span is a few 100 years later than the beginning of the Neolithic period in Morocco and ends around 4000 cal. BP when iron, copper and zinc content started to increase in sedimentary records. Geochemical elements were extracted to make metal tools, which marks the end of the Neolithic period and probably the beginning of a prehistoric metallurgical industry era. We observe that several anthropogenic indicators increase when the human population increases. Likewise, natural ecosystem changes, including forest species, are negatively or positively impacted by an increase or a decrease in human population size, respectively, during the Holocene.
The correlations we have performed between 14C date SPDs, as a proxy for population change, and the fossil pollen data suggest the following:
Early expansion of human populations around 7000 cal. BP took place mainly in the lowlands, and if there was a spread towards the mountain areas then it was either minor or the spreading populations had a negligible impact on the natural ecosystems. However, early Holocene anthropogenic evidence is derived from very few lowland sites. To confirm whether there was a more extensive vegetation change, additional data from archaeological off-site contexts, such as lake sediments, are still needed.
The principal human activity detected in the lowland records involved grazing and farming until ca. 4000 cal. BP.
Plant domestication seems not to have taken place before the early expansion of human populations in Morocco, which is recorded during the Neolithic around 7000 cal. BP.
The conclusions drawn in this study have the potential to be clarified through integration of additional archaeological sites with more radiocarbon dates and new fossil pollen records from lower elevation areas.
Footnotes
Acknowledgements
This work stems from a workshop dedicated to the Leverhulme Trust-funded project ‘Changing the face of the Mediterranean: land cover and population since the advent of farming’ organized by J. Woodbridge, R. Fyfe and N. Roberts in Mallorca in September 2017.
Funding
The author(s) received no financial support for the research, authorship and/or publication of this article.