
Abstract
In this article, we examined the indicative value of a relationship between two non-pollen palynomorphs (NPPs), fungal HdV-10, related to the presence of Calluna vulgaris, and HdV-31A, which is testate amoeba – Archerella flavum. Both are frequently present on slides designated for pollen analysis, prepared from Sphagnum peat. We analysed three profiles from three extensive ombrotrophic peatlands in northern Poland, in which the content of testate amoebae (TA) was examined and TA-inferred depth to the water table (DWT) was reconstructed. The new analysis of palynological samples regarding NPPs revealed that strong increases in HdV-10 content were mostly simultaneous to HdV-31A declines. However, the relations between both types were not statistically significant. The rapid increases in HdV-10, despite the fact that this type of conidiospores is related to drier habitats on peatlands, were tentatively interpreted as an indicator of rapid rises in the water table level which, in consequence, might have stimulated the production of these spores by fungi. In addition, a negative correlation between HdV-31A and Arcella discoides and positive one, but weaker, in case of HdV-10 (attributed by some authors to species Trichocladium opacum (Corda) S Hughes) and A. discoides show a link between mutual fluctuations of both NPPs and hydrological instabilities on peatland. The Bagno Kusowo bog, the westernmost peatland subjected to the study, displayed an intriguing agreement between the presence of peat sections with strong increases in HdV-10 and cold climate events affecting Europe. Our study reveals that counting even a limited number of NPP types during the standard pollen analysis of Sphagnum peat may support the interpretation of results, especially, in cases when the investigation lacks testate amoeba analysis. When the reconstruction of TA-inferred DWT is provided, it may introduce additional information about the patterns of hydrological dynamics.
Introduction
Ombrotrophic peatlands, ecosystems dependent on precipitation, are sensitive to climatic changes and thus perceived as highly suitable for palaeoclimatic/environmental reconstructions (e.g. Chambers et al., 2012; Lamentowicz et al., 2009; Langdon et al., 2003; Väliranta et al., 2012). This is because of anoxic conditions and low pH that provide suitable conditions for the preservation of a broad set of subfossils (e.g. Chambers et al., 2012; Moore et al., 1991; Rydin and Jeglum, 2006). Among microfossils widely distributed in ombrotrophic peat, testate amoebae (TA) enable quantitative estimation of changes of the depth to the water table (DWT; Amesbury et al., 2016; Charman et al., 2007). However, there are numerous cases of research where the reconstruction of the water table changes on the basis of TA, because of limited financial resources or lack of suitable material, is impossible. Hence, other alternative indicators of hydrological changes are needed. Non-pollen palynomorphs (NPPs), a diverse group of microfossils, which consists of fungi, green algae, testate amoebae, invertebrates and others, and may withstand laboratory procedures for palynological analysis, might be useful in such cases (Miola, 2012; Shumilovskikh et al., 2015; van Geel, 1978). As the palynological analysis is one of the most widespread methods for retrieving information about past peatland ecosystem changes (Moore et al., 1991), the exploration of NPPs, which are present on palynological slides, seems to be a right direction to supplement the knowledge about past hydrological changes on peatlands, in cases when TA analysis is not possible.
In this study, we focused on the relationship between two NPP types frequently detected on slides designated to pollen analysis of Sphagnum peatlands (1) HdV-10 and (2) HdV-31A (Figure 1). HdV-10 is of a fungal origin and represents conidiospores (asexual propagules). Morphological inspection of this type enabled its attribution to species Trichocladium opacum (Corda) S. Hughes (Miola, 2012, where it is mentioned as Trigocladium opacum). However, to the best of our knowledge, there were no molecular studies focused on attribution of this NPP type to particular fungal species. T. opacum was reported from rotten wood, dead herbaceous stems and leaves, and soil (Goh and Hyde, 1999, and references therein). In palaeoecological studies, HdV-10 is linked with the presence of Ericaceae plants, mainly Calluna vulgaris (both roots and pollen; Shumilovskikh et al., 2015; van Geel, 1978; van Geel and Middeldorp, 1988) but also remains of Erica tetralix (roots; Bakker and van Smeerdijk, 1982). In general, HdV-10 is considered a good indicator of dry conditions in ombrotrophic peatlands (Kuhry, 1985; van Geel, 1978). HdV-31A is the testate amoeba Archerella flavum, known also as Amphitrema flavum (Miola, 2012). Its tests are among those which may resist chemical treatment of samples for pollen analysis and are easy to detect during the standard microscopic survey of slides (Payne et al., 2012). A recent study by Karpińska-Kołaczek et al. (2018) has revealed the intriguing, mutually exclusive optima of both NPPs, recorded in palynological data and interpreted as the signal of water table fluctuations. Unfortunately, the analysed profile was devoid of TA-inferred water table reconstruction, which prevented the validation of the inferences.
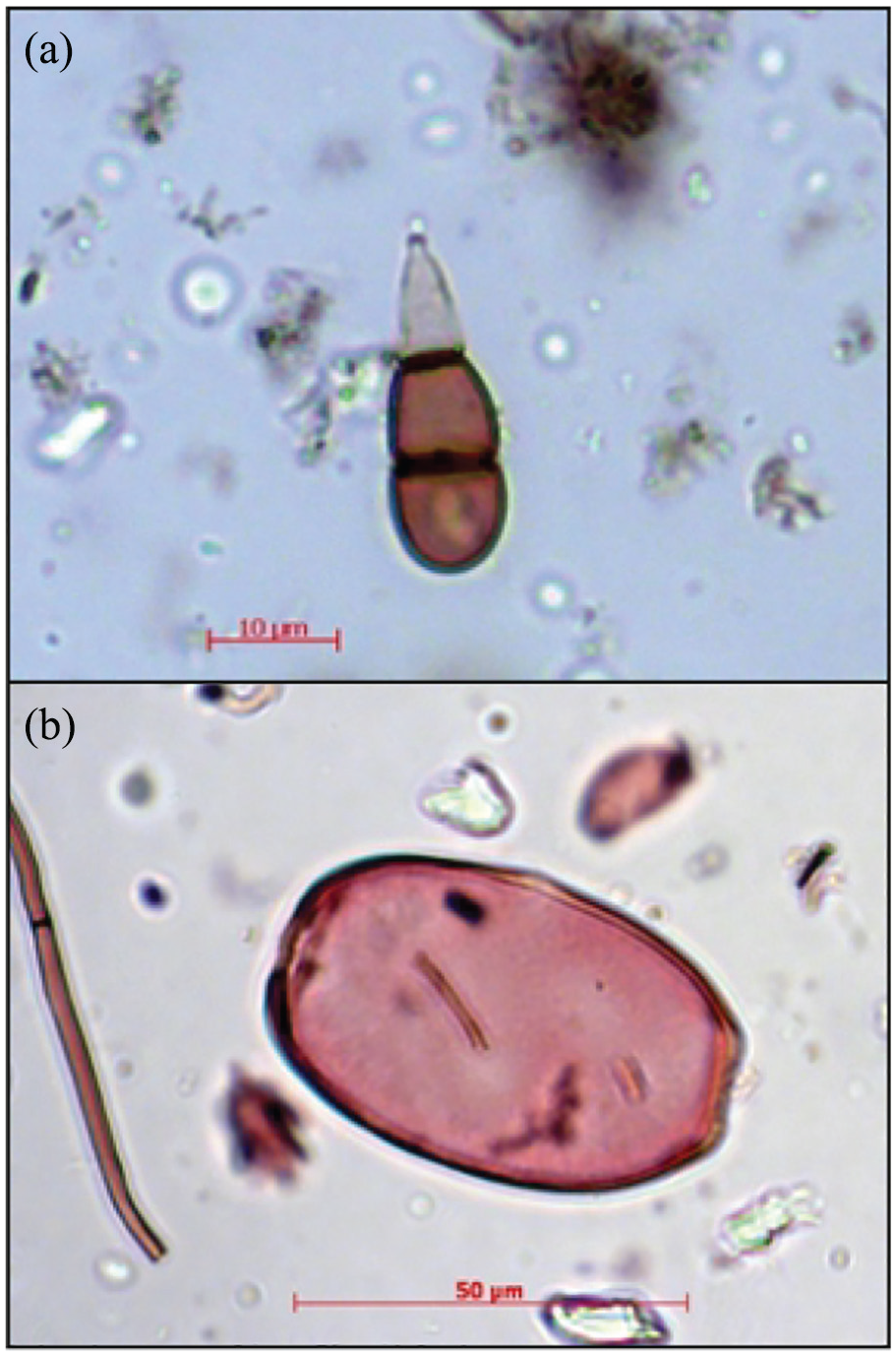
Non-pollen palynomorphs (NPP): (a) HdV-10, (b) HdV-31A Archerella flavum.
We hypothesise that the mutually exclusive extrema of HdV-31A and HdV-10 reflect dry conditions when HdV-10 increases with a simultaneous decline in HdV-31A and wet when reversed. To investigate this, we juxtaposed the newly retrieved NPP data with the already existing TA-inferred quantitative reconstructions of the DWT (Gałka et al., 2015, 2017; Lamentowicz et al., 2015). As the testate amoebae are widespread in peatlands and particular taxa are linked with the specific peatland microforms (Charman et al., 2000; Tolonen et al., 1992; Warner, 1990), we combined our results considering HdV-10 and HdV-31A interrelationship with the presence of TA species identified during TA analysis (a separate analysis) to determine the in-depth characteristics of the habitat. We also examined whether climate affected the distribution patterns of both NPP types in peat.
Material and methods
In 2011, along a west–east transect, three profiles from three large Sphagnum peatlands in northern Poland were collected: (1) Bagno Kusowo (BK, 318.82 ha), (2) Gązwa (Gz, profile GzII, 204 ha) and (3) Mechacz Wielki (MW, 146.72 ha; Figure 2; Table 1). The profiles have a thickness of 800, 835 and 600 cm, respectively. For better time control, we used absolute chronologies (Gałka et al., 2015, 2017; Lamentowicz et al., 2015), refined by Kołaczek et al. (2019). In the case of Gz, in which 14C date inversions have been detected, the model with the highest agreement index and the lowest number of excluded dates was selected (Supplementary Data 1, available online).
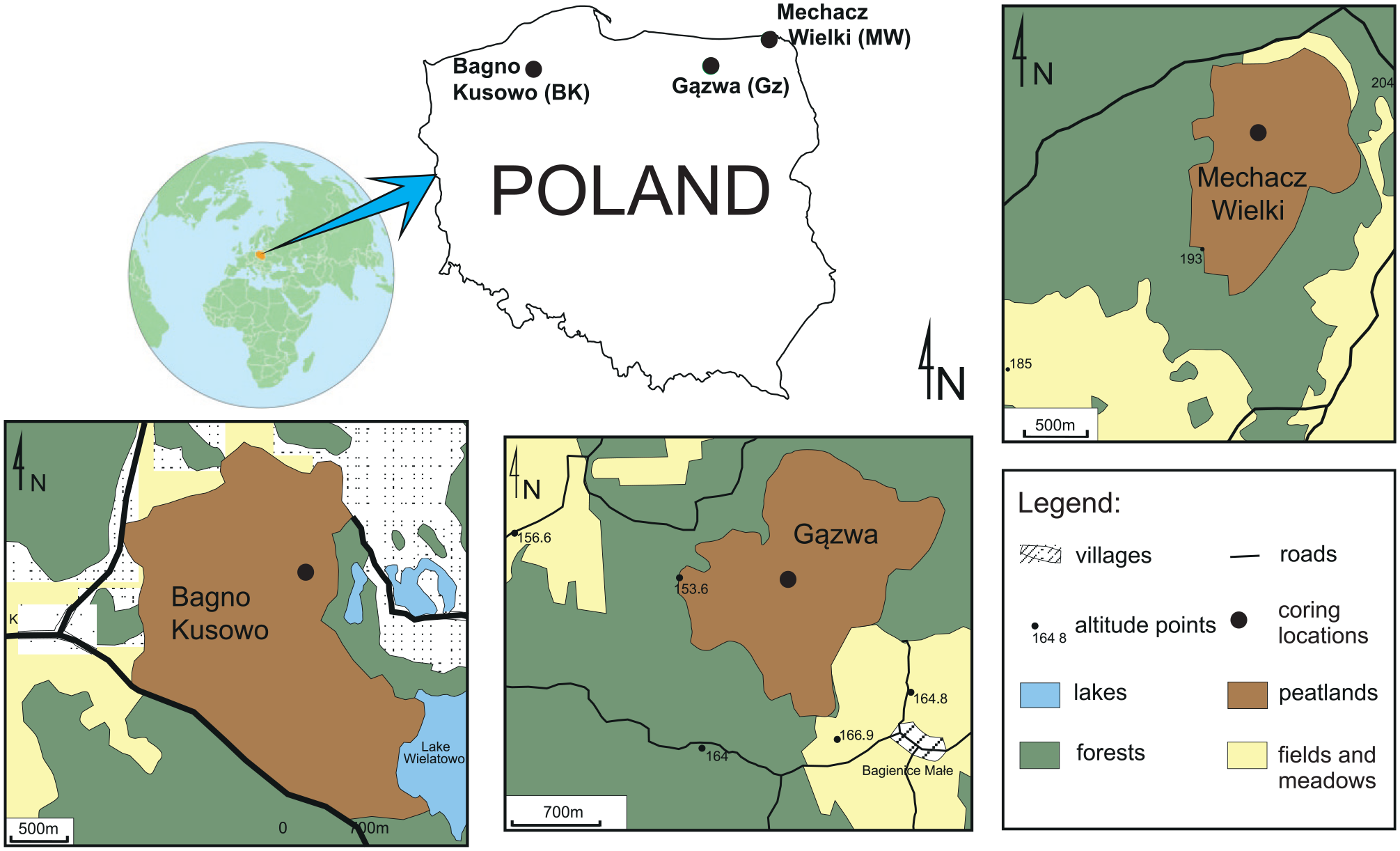
Location of the sites, with their local settings (Kołaczek et al., 2019; Marcisz et al., 2017; modified).
Site settings.

The NPP composition was determined in the samples subjected to standard laboratory procedures including acetolysis, the same ones as those designated to palynological analysis in previous studies (Gałka et al., 2015, 2017; Lamentowicz et al., 2015). As the Lycopodium tablets were added to samples (Stockmarr, 1971) and counted during both pollen and NPP investigations, it was possible to combine NPP and pollen data. The NPP and pollen percentages were calculated based on total pollen sum, so then values of some types and pollen taxa may exceed 100%. This calculation enabled avoiding the bias of the impact of NPP types occurring in prominent abundances on percentage values of other NPPs. For the purpose of the presented study, we selected only HdV-10 and HdV-31A from NPPs and the most frequent Ericaceae pollen taxa (as an indicator of suitable habitat for the HdV-10 occurrence).
The samples for TA analysis were taken from the same depth horizons as those for pollen and NPPs and were prepared according to the standard laboratory procedures, that is, stirring in distilled water and sieving through the mesh of ø 200-μm size (Gałka et al., 2015, 2017; Lamentowicz et al., 2015). The presence of Ericaceae plant tissues (as a potential marker of the conditions suitable for the HdV-10 presence) and of unidentified organic matter (as a potential marker of dry periods) were determined in each 1-cm slice of the profile and their values were rounded to 5% grade (Gałka et al., 2015, 2017; Lamentowicz et al., 2015).
To check the potential correlations between HdV-10 and HdV-31A, and between DWT table and TA taxa, a Pearson correlation coefficient was applied. These calculations and graphical output of the analysis were carried out using PAST software (Hammer et al., 2001). Only statistically significant results (p < 0.05) have been presented. NPP, pollen, plant macrofossil and DWT curves were drawn using the TILIA software (Grimm, 1992).
Results
The percentage values of HdV-10 revealed more distinct fluctuations (0–370% in BK, 0–311% in Gz and 0–295% in MW) in comparison with HdV-31A (0–24% in BK, 0–25% in Gz and 0–39% in MW; Figures 3–5). HdV-31A, even if less abundant, was more regular in analysed samples: (1) in BK, HdV-10 was identified in 78% of samples, whereas HdV-31A in 91%; (2) in Gz, HdV-10 was in 30% of samples, whereas HdV-31A in 85%; and (3) in MW, HdV-10 was found in 44% of samples, whereas HdV-31A in 93%. The maxima of HdV-10 percentages were most frequent in BK and were identified in the layers reflecting the age of ca. (1) 3250–3050, (2) 2950–2750, (3) 2650, (4) 2300, (5) 1900, (6) 1700–1300, (7) 900–800 and (8) 650–250 cal. BP. Some of these were simultaneous with TA-inferred wet or very wet events ca. 3100, 2650, 2300 and 1450 cal. BP. What is noticeable, the declines in DWT (wet stages) of the highest amplitude were always simultaneous to the rapid increase(s) in HdV-10 percentages. In Gz, very clear maxima of HdV-10 were identified in the layer representing the age ca. (1) 2650, (2) 2350, (3) 2100, (4) 2000 and (5) 250 cal. BP. All of them were simultaneous with low DWT characterising wet stages. Finally, the most incomprehensive signal was identified in MW, where HdV-10 percentage peaks were identified in the peat horizons reflecting periods ca. (1) 1080, (2) 970, (3) 940, (4) 900–850 (towards drier conditions), (5) 800–750 and (6) 700–650 cal. BP (towards wet conditions; Figures 3–5).
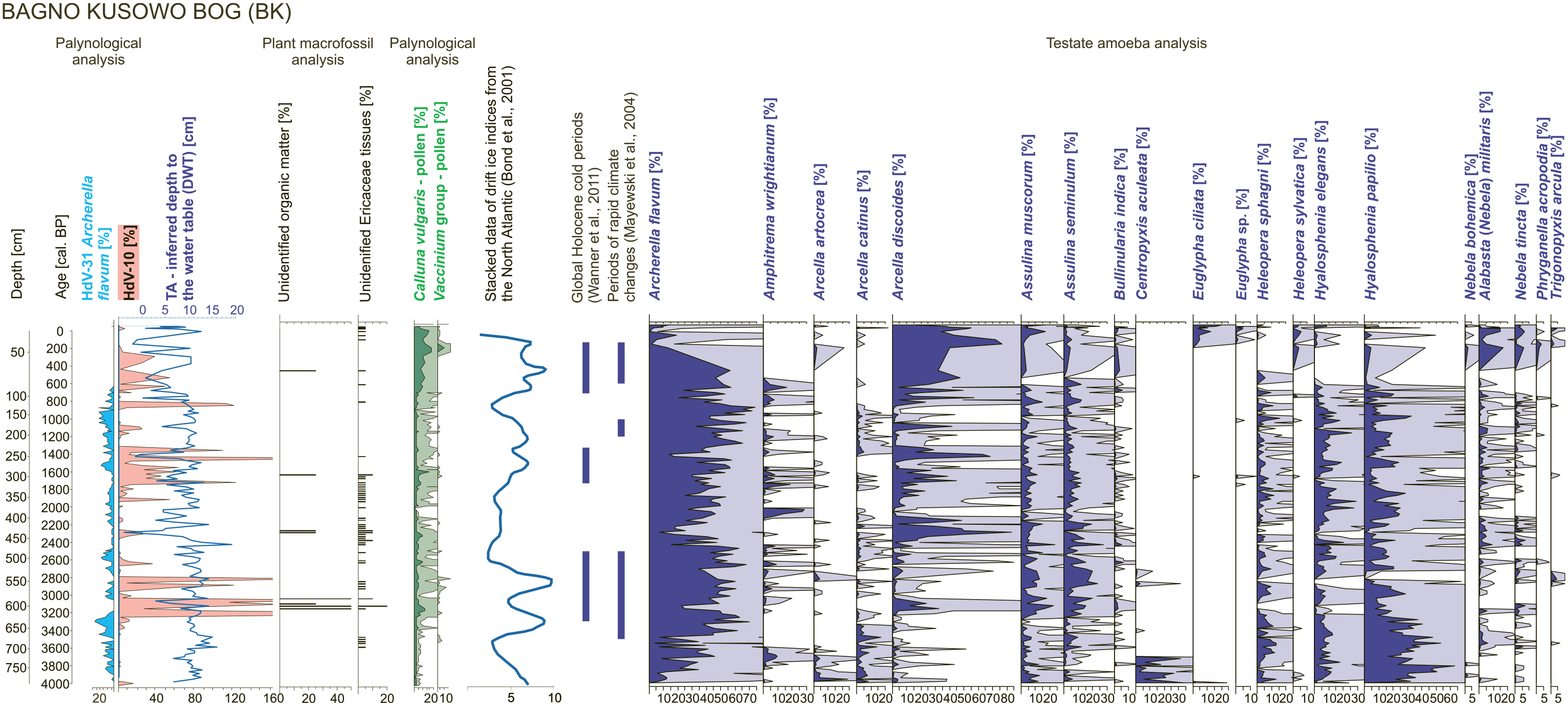
Bagno Kusowo bog (BK) – HdV-10 and HdV-31A percentages along testate amoebae (TA)-inferred DWT reconstruction, Ericaceae plant tissues content in peat (data from samples parallel to samples designated to TA analysis are presented) and selected TA (the most frequent and these which revealed correlations to HdV-10 and/or HdV-31A, with p < 0.05). TA-inferred DWT, plant macrofossils and TA data were published in the previous article (Lamentowicz et al., 2015), whereas palaeoclimate data follow Bond et al. (2011), Mayewski et al. (2004) and Wanner et al. (2011).
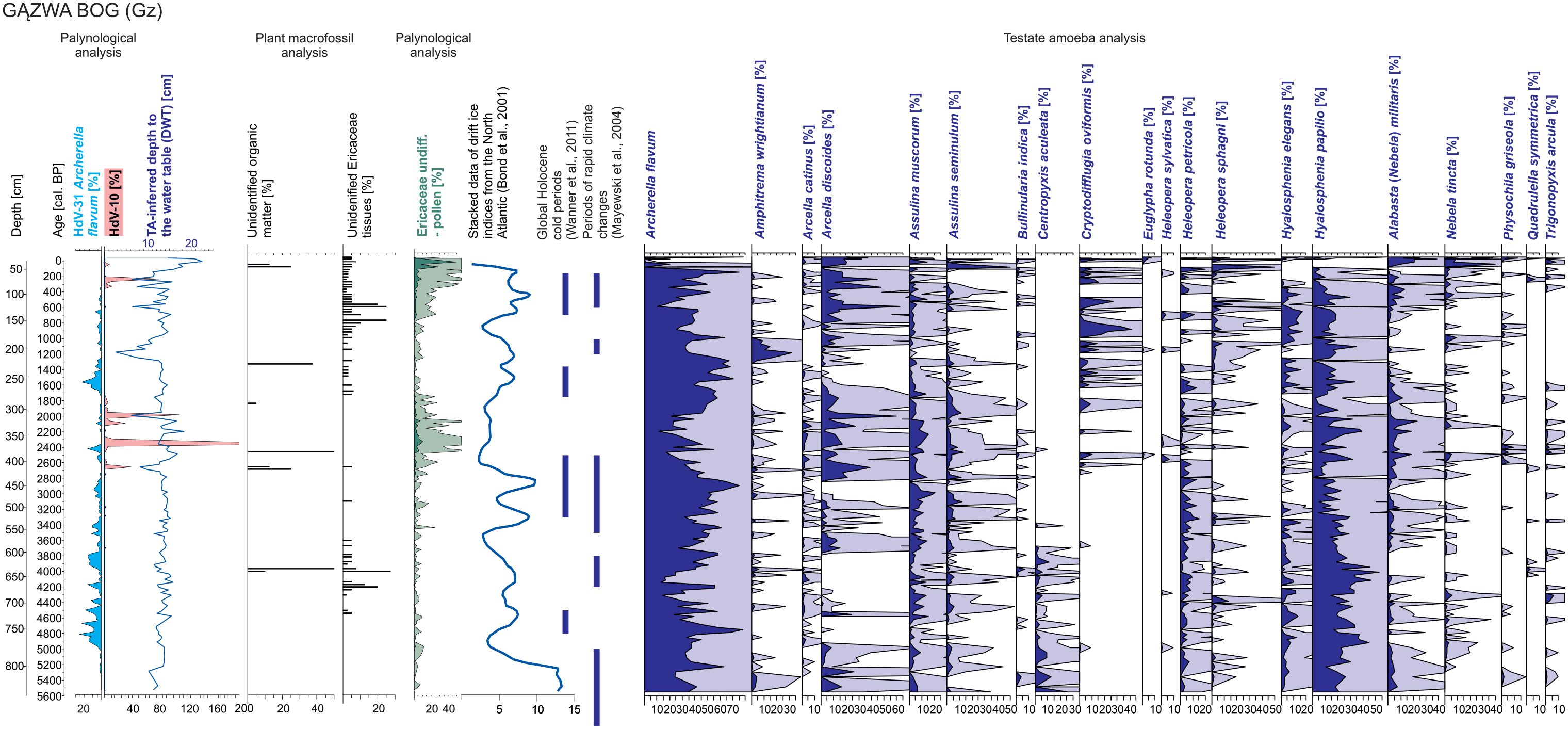
Gązwa bog (Gz) – HdV-10 and HdV-31A percentages along testate amoebae (TA)-inferred DWT reconstruction, Ericaceae plant tissues content in peat (data from samples parallel to samples designated to TA analysis are presented) and selected TA (the most frequent and these which revealed correlations to HdV-10 and/or HdV-31A, with p < 0.05). TA-inferred DWT, plant macrofossils and TA data were published in the previous article (Gałka et al., 2015), whereas palaeoclimate data follow Bond et al. (2011), Mayewski et al. (2004) and Wanner et al. (2011).

Mechacz Wielki bog (MW) – HdV-10 and HdV-31A percentages along testate amoebae (TA)-inferred DWT reconstruction, Ericaceae plant tissues content in peat (data from samples parallel to samples designated to TA analysis are presented) and selected TA (the most frequent and these which revealed correlations to HdV-10 and/or HdV-31A, with p < 0.05). TA-inferred DWT, plant macrofossils and TA data were published in the previous article (Gałka et al., 2017), whereas palaeoclimate data follow Bond et al. (2011), Mayewski et al. (2004) and Wanner et al. (2011).
Pearson’s correlation did not reveal any statistically significant correlations (p < 0.05) between HdV-10 and HdV-31A (Figure 6; Supplementary Data 2, available online). When HdV-10 is present, it often displays strong fluctuations and most of the peaks are linked with declines in HdV-31A percentages (Figures 3–5). Both NPP types revealed unclear relationships with DWT: (1) in BK, only in the case of HdV-31A, low but positive correlation to DWT (r = 0.25) was recorded, which contradicts the A. flavum ecology (Lamentowicz and Mitchell, 2005); (2) in Gz, HdV-10 revealed low and negative correlation to DWT, which contradicts its indicative potential (Kuhry, 1985; van Geel, 1978); and (3) in MW only, a positive weak correlation between HdV-10 and DWT (r = 0.31) was identified. The results of Pearson’s correlation between NPP and TA data revealed a positive correlation between HdV-31A (A. flavum) percentages calculated from pollen samples and TA analysis for each site (r = 0.52 in BK, r = 0.34 in Gz, r = 0.36 in MW; p < 0.05). Moreover, all sites revealed the statistically significant (p < 0.05) positive correlations between HdV-31A and Hyalosphenia papilio (the strongest in Gz), and in case of BK also between Hyalosphenia elegans and Heleopera sphagni. Each site recorded a negative correlation between HdV-31A and Arcella discoides (r = −0.43 in BK; r = −0.4 in Gz, r = −0.41 in MW, p < 0.05; Figure 6). Weak negative correlations (p < 0.05) were also identified between HdV-31A and (1) Assulina muscorum (r = −0.35), Assulina seminulum (r = −0.29) and Bullinularia indica (r = −0.32) in BK, (2) Alabasta (Nebela) militaris (r = −0.23) in Gz, and (3) Nebela tincta (r = −0.32), Heleopera sylvatica and Physohila griseola in MW. The strongest correlations between HdV-10 and TA were registered in the MW profile, in which this NPP positively correlated with A. muscorum, A. seminulum, Bulinularia indica, Difflugia pulex, Nebela tincta and Trigonopyxis arcula. Statistically significant (p < 0.05), negative, but weak, correlations were detected only in BK in case of Arcella catinus, H. sphagni, H. elegans and H. papilio.

Pearson’s correlations between HdV-10, HdV-31A and testate amoebae (derived from independent TA analysis; Gałka et al., 2015, 2017; Lamentowicz et al., 2015).
Discussion
Relationships between HdV-10 and HdV-31A as the indicator of water table changes
The highest abundance of HdV-10 was connected with the presence of C. vulgaris or other representatives of the Ericaceae family (Figures 3–5; Gałka et al., 2015, 2017; Lamentowicz et al., 2015). However, in our data, pollen and/or macrofossils were widely distributed within peat whereas HdV-10 was not. Our study did not reveal the statistically significant negative correlation between HdV-10 and HdV-31A. However, when the rapid increases in HdV-10 occur, the declines in HdV-31A values are recorded. Strong positive correlations between HdV-10 and Bulinularia indica, Assulina seminulum, Assulina muscorum, Difflugia pulex, Nebela tincta and Trigonopyxis arcula were revealed only in MW. In other sites, the correlations between HdV-10 and TA, even if statistically significant (p < 0.05), were very weak (Figure 6). Only in the MW site, HdV-10 revealed the positive correlation with TA-inferred DWT, that is, it indicated low water table, as it was preliminarily assumed in this study. D. pulex was identified only in one sample so its interpretative strength for HdV-10 presence is very low. Other taxa, were more common and they are associated with dry and acidic habitat, as well as low environmental variability (Lamentowicz and Mitchell, 2005; Mieczan, 2009; Sullivan and Booth, 2011). Lamentowicz and Mitchell (2005) classified Trigonopyxis arcula, Bulinularia indica and Nebela tincta as the most xerophilous among taxa identified in Sphagnum peatlands of northern Poland. On the other hand, the MW site was characterised by the most stable hydrological conditions and the highest peat accumulation rate among analysed sites (Gałka et al., 2017). This site revealed an unclear relationship between the extrema of HdV-10 and declines in DWT (Figure 2). In contrast, in the Gz profile, in which the vertical dislocations in peat were identified (Kołaczek et al., 2019), the maxima of HdV-10 were clearly related to declines in DWT. Different species of fungi, when the availability of oxygen is limited, may react by changing the type of reproduction (Carlile et al., 2001). Hence, it is possible that fungus producing the HdV-10 conidia, if subjected to stress caused by oxygen depletion, may react by increased productivity of them. This was observed in the case of Beauveria bassiana that increased the production of conidia under low oxygen atmosphere (Garza-López et al., 2012). On the other hand, fungus Metarhizium anisopliae increased conidia production in a culture exposed to oxygen-enriched atmospheres (Tlecutil-Beristain et al., 2010). Moreover, it has been proven that oxidising states favour cytodifferentiation in different genera of fungi (Abrashev et al., 2008; Belozerskaya and Gessler, 2006; Gocheva et al., 2009; Miller et al., 2004). So then, as there is no detailed study on the reaction of fungi producing HdV-10 conidia to oxygen stress it is difficult to interpret the strong increases in this NPP frequency as a reaction to the increase in the water table.
It is not possible to perform a reliable quantitative reconstruction of TA-inferred DWT, calculated on the basis of TA from pollen slides, because of the loss of most of the taxa during chemical treatment (acetolysis and the addition of HCl and KOH) of samples (Payne et al., 2012). However, our study revealed that the values of A. flavum (type HdV-31A) counted during NPP analysis are correlated with the values of A. flavum in samples subjected to TA analysis (r = 0.52 in KB, r = 0.34 in Gz and r = 0.36 in MW, p < 0.05). The same applies to another mixotrophic species H. papilio. Hence, the presence of HdV-31A may also provide information about the conditions for the occurrence of mixotrophs, a group related to carbon fixation in peatlands (Jassey et al., 2015). All sites revealed a negative correlation between HdV-31A and A. discoides (TA analysis). This taxon (A. discoides type) was recognised as having broad ecological tolerance (Lamentowicz and Mitchell, 2005) and both as a species and type, in studies from Sphagnum peatlands in Europe, was related to rather wet habitats (Lamentowicz and Mitchell, 2005; Mazei and Tsyganov, 2007; Mieczan, 2009; Tolonen et al., 1992). Lamentowicz et al. (2009) interpreted the past dominance of A. discoides in peatlands as the effect of abrupt fluctuations of the water table, whereas A. flavum was rather associated with relatively stable water table conditions. Sullivan and Booth (2011) reported that A. discoides is more abundant under highly variable conditions. As the HdV-10 revealed statistically significant but weak correlations with A. discoides in two from three sites (Figure 3), the mutually exclusive extrema in HdV-10 and HdV-31A values in pollen spectra should rather be interpreted as a result of abrupt fluctuations in the water table.
Autogenic processes (movements of microforms – hummocks and hollows) should always be taken into consideration when the relationship between HdV-31A and HdV-10 is interpreted. The places of coring are not always evenly covered by C. vulgaris in space and time, so the lack of HdV-10 might be an effect of the lack of these plants. Hence, if HdV-10 and HdV-31A are used for the interpretation of the potential impact of climate, only sections of profiles with the regular presence of both NPP types should be selected. At the same time the examination of plant tissues, in terms of occurrence of C. vulgaris rootlets, should be considered.
The TA-inferred reconstruction of DWT reflects the averaged pattern per time-interval spanned by the sample, so the relationships between HdV-10 and HdV-31A may introduce additional qualitative information about the dynamics of the water table fluctuations within such time-interval. The value of NPPs, as a proxy providing supplementary information to TA-inferred DWT, was shown in the palaeoecological reconstruction from the Linje mire (central-northern Poland; Marcisz et al., 2015). In this case, the presence of algae Botryococcus sp. was recorded during the driest stage in peatland history. This was interpreted as the occurrence of short periods of inundations within longer and generally dry time-intervals (identified in TA-inferred DWT). However, there are also examples, as in the case of Puścizna Mała and Puścizna Krauszowska bogs (southern Poland), when it is difficult to unequivocally interpret the HdV-10 versus HdV-31A relationship (Kołaczek et al., 2018).
HdV-31A and HdV-10 versus global/regional climate and environmental changes
Among three analysed peatlands, only BK revealed an agreement between the periods of fluctuations of water table recorded in HdV-31A and HdV-10, and cold climate events (Mayewski et al., 2004; Wanner et al., 2011). The series of rapid increases in HdV-10 were identified in the layers concurrent to ca. 3500–2500, 1750–1350 and 700–150 cal. BP cold periods (Wanner et al., 2011) and periods of rapid climate changes ca. 3500–2500, 1200–1000 and 600–150 cal. BP (Mayewski et al., 2004). The analogous rapid fluctuations in abundance of HdV-10 were not synchronous to these climatic periods in the Gz and MW peatlands. Only a peat layer reflecting the period of 2700–2600 cal. BP in Gz was characterised by the increase in HdV-10 abundance, parallel to the increase in the water table reconstructed from TA. This period was coincident to Homeric solar minimum that took place ca. 2750–2550 cal. BP (Słowiński et al., 2016; Van Geel et al., 1999); however, it must be stressed that in GZ, the most prominent increases in abundance of HdV-10 appeared later, from ca. 2400 to 1950 cal. BP. Between 2700 and 2600 cal. BP, an increase in HdV-10 with a simultaneous increase in the water table was also identified in BK. Słowiński et al. (2016), on the example of the Homeric solar minimum, showed that westward sites recorded rather wet conditions (TA-inferred DWT), whereas eastward sites rather dry conditions during this period. Nonetheless, in the Lake Czarne bog (north-eastern Poland), where HdV-10 versus HdV-31A relationship was investigated, fluctuations in the water table (dry-wet shifts) were identified at that time (Karpińska-Kołaczek et al., 2018). However, the Lake Czarne bog, being much smaller (ca. 5 ha) in comparison with the BK, Gz and MW bogs, was more prone to climatic changes and strongly dependent on the water level changes of the adjacent lake.
The studied sites revealed that the westernmost peatland revealed the strongest synchronicity between potential water table fluctuations and cold climatic periods (sensu Wanner et al., 2011) as well as periods of rapid climate changes (sensu Mayewski et al., 2004). The BK peatland, situated in north-western Poland, was more influenced by the impact of oceanic air masses. The previous studies on presented peatlands revealed the stronger impact of continental climate on Gz and MW sites (Marcisz et al., 2017). Nonetheless, in western Poland, further studies on NPPs and TA are needed to explore the potentially variable response of big peatlands to events of climate cooling located along the climatic gradient in Central Europe.
Another problem which arises, when the potential linkages between the water table changes and periods of climate changes in the past are investigated, are absolute chronologies. In the case of the GzII profile, the peat layer reflecting the age of ca. 3100–700 cal. BP (480–140 cm) was characterised by 14C date inversions (Kołaczek et al., 2019). Hence, even if the most statistically reliable age-depth model is selected as a basis for absolute chronology, it is still probable that the assignment of the environmental event to time-frames is inaccurate. Therefore, the synchronicities, leads and lags between environmental processes and climate events might also be the effect of the quality and/or choice of absolute chronologies.
Conclusion
The mutually exclusive extrema of percentage values of NPP types HdV-10 and HdV-31A, identified in peat cores collected from three vast peatlands in northern Poland, might be explained by the following mechanisms:
Fungus producing conidiospore type HdV-10, as most of the terrestrial fungi, probably cannot thrive when inundated, and this kind of stress may stimulate the production of the propagules. Such temporal increases in the water table level, if they do not contribute to the retreat of C. vulgaris, fail to provide conditions for the expansion of A. flavum and/or other TA mixotrophic species. Hence, in some cases, it is possible to detect simultaneous rapid increases in HdV-10 values and increases in water table level (reconstructed from testate amoebae). However, it must be stressed that until accurate taxonomic affinity of HdV-10 and its reaction to the oxygen depletion is recognised, this interpretation of HdV-10 during periods of the generally high water table is tentative.
The negative correlations between HdV-31A (pollen analysis) and A. discoides (testate amoeba analysis) and mostly positive, but weaker, between HdV-10 (pollen slides) and A. discoides (testate amoeba analysis) show that fluctuations in the frequency of HdV-10 and HdV-31A (pollen analysis) indicate rather fluctuations in the water table on peatland.
The Bagno Kusowo peatland, the only within this study, revealed an agreement between the presence of events with strong increases in HdV-10 and climate cold periods (sensu Wanner et al., 2011) and periods of rapid climate changes (sensu Mayewski et al., 2004). This site, as the only representing the area of north-western Poland, was more influenced by oceanic air masses, which might have affected the record. Nonetheless, further research on other sites influenced by oceanic climate are needed to verify this pattern.
Our study reveals that parallel counting of easy to identify NPP types HdV-10 and HdV-31A in pollen slides may introduce additional qualitative information about the patterns of the past water table changes.
Supplemental Material
Supplementary_Data1 – Supplemental material for Do the relationships between testate amoebae and fungi reflect the variability of past water table fluctuations in the ombrotrophic peatlands of Central Europe?
Supplemental material, Supplementary_Data1 for Do the relationships between testate amoebae and fungi reflect the variability of past water table fluctuations in the ombrotrophic peatlands of Central Europe? by Piotr Kołaczek, Monika Karpińska-Kołaczek, Mariusz Gałka, Grażyna Miotk-Szpiganowicz, Milena Obremska and Mariusz Lamentowicz in The Holocene
Supplemental Material
Supplementary_Data_2_poprawiony – Supplemental material for Do the relationships between testate amoebae and fungi reflect the variability of past water table fluctuations in the ombrotrophic peatlands of Central Europe?
Supplemental material, Supplementary_Data_2_poprawiony for Do the relationships between testate amoebae and fungi reflect the variability of past water table fluctuations in the ombrotrophic peatlands of Central Europe? by Piotr Kołaczek, Monika Karpińska-Kołaczek, Mariusz Gałka, Grażyna Miotk-Szpiganowicz, Milena Obremska and Mariusz Lamentowicz in The Holocene
Footnotes
Authors’ note
The data already published, that is, TA, TA-inferred DWT and plant macrofossils (Gałka et al., 2015, 2017; Lamentowicz et al., 2015) were retrieved within the frame of the previous NSC project NN305 062 240 (PI: Mariusz Gałka).
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship and/or publication of this article: The present research was supported by the National Science Centre (NCS; Poland; Grant No. UMO-2014/13/B/ST10/02091; PI: Monika Karpińska-Kołaczek) which covered the expenses connected with conducting non-pollen palynomorph analysis presented in this manuscript, preparation of absolute chronologies (Kołaczek et al., 2019) and the entire cost of preparation of this article.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.