
Abstract
South American camelids (SAC) have occupied a central role in the development of Andean societies. They are widely distributed in South America and since ancient times are an important factor in Andean economies and social and ritual life. The archaeological site Punta de Peña 4 (PP4) is located in Antofagasta de la Sierra Basin (Southern Puna of Argentina). PP4 is a rock overhang and presents a large occupational sequence (early Holocene until historic times). The aim of this contribution is to study parasite diversity and the relationship among parasites and SAC through holocenic times in the Southern Puna of Argentina. Besides, differences in parasite egg preservation in the site were discussed. A total of 65 coprolites assigned to SAC were rehydratated, homogenized, filtered, allowed to spontaneous sedimentation and examined for parasite remains. Given the morphology of the coprolites, the biogeographic origin and the presence of coccidian oocysts of Eimeria macusaniensis in some of them, a camelid origin of the coprolites is suggested. This is the first paleoparasitological study from PP4, an archaeological site of Antofagasta de la Sierra, a key region in northwestern Argentina and Central Andes. Eleven taxa of parasites were found belonging to Apicomplexa, Platyhelminthes, and Nematoda. Specific parasite species of SAC were found, such as E. macusaniensis and Lamanema chavezi/Nematodirus lamae. A remarkable finding was the presence of Moniezia sp. and Strongyloides sp. in coprolites from different levels of PP4, pointing out the presence of these genera in native artiodactyls of the region previous to the arrival of Hispanic societies. Results displayed a change in the number of parasite taxa present in SAC through time. The parasite-SAC relation through time and differences in parasite egg preservation between levels and sectors (covered and uncovered) of the site are discussed.
Introduction
Current South American camelids (SAC) include four native species, two domestic species, the llamas (Lama glama) and the alpacas (Vicugna pacos), and two wild species, vicuñas (Vicugna vicugna) and guanacos (Lama guanicoe). They are distributed along the Andes. Llamas, vicuñas and guanacos can be found in Argentina. SAC have occupied a central role in the development of Andean societies, both for ancient hunter-gatherers and for more recent pastoralists and farmers. In Prehispanic times, camelids were widely distributed and were the most important factor in Andean economies and social and ritual life (Mengoni Goñalons, 2008; Mengoni Goñalons and Yacobaccio, 2006). At present, alpacas and llamas are important livestock in some regions along the Andes (Bornstein, 2010).
The archaeozoological evidence places domestication of the alpaca from the vicuña in the wet Puna of Peru’s central Andes, 6000–7000 years ago, and domestication of the llama from the guanaco in the dry Puna in localities that today correspond to Peru, Chile, and Argentina (Mengoni Goñalons, 2008). The Puna of Argentina is a highland desert located between 22° and 27°S and at 3000–5000 masl. Evidence of SAC presence in Antofagasta de la Sierra Basin (Southern Puna of Argentina) is wide and comes from archaeological sites dated to the early Holocene (human occupations reaches to at least 10,200 years BP) to historical times (see Martínez et al., 2010, and references therein). SAC hunting was the main subsistence activity through the Holocene, even when agro-pastoral practices were also established (Urquiza, 2009). The zooarchaeology record of this area is dominated by SAC remains, while small mammals, reptiles, and birds are also presented in less frequency (Olivera and Grant, 2008; Urquiza and Aschero, 2014; Urquiza et al., 2013; Urquiza and Echeverría, 2018; and references there in). Taphonomic studies made on osseous material from the sites of the region highlighted the good preservation of the archaeofaunisitic record and differences on taphonomic processes that take place in the area (e.g. Marozzi et al., 2015; Urquiza, 2010, 2016).
SAC are generally affected by infectious and parasitic diseases. Endoparasites are considered a major health problem of camelids. Within parasites, some of them are specific parasites from SAC and others are shared with wild and domestic ruminants as sheep and goats, with which they share grazing and/or breeding areas (Aguirre and Cafrune, 2007; Beldomenico et al., 2003; Cafrune et al., 2009; Gonzalez-Rivas et al., 2019; Navone and Merino, 1989; among others). Some paleoparasitological studies in SAC were done, most of them from Patagonia (Argentina) (Amalfitano et al., 2017; Fugassa, 2007; Taglioretti et al., 2015, 2017). These studies allowed starting to know parasite diversity in the past and the relationship between parasites and SAC through the Holocene. One important aspect of paleoparasitological studies is the effect of taphonomic factors in the preservation of parasite remains. However, there are few studies performed on this topic at the moment.
Despite the numerous zooarchaeological researches done for several decades in the Southern Puna, there are few paleoparasitological studies. Therefore, this study aims to: (1) study the gastrointestinal parasite fauna in SAC from the Punta de Peña 4 archaeological site, Antofagasta de la Sierra, (2) discuss host-parasite relationships through the studied period (middle Holocene to late-Holocene), and (3) evaluate differences in parasite egg preservation between levels and sectors (covered and uncovered) of the site.
Study site, material, and methods
Environmental description and archaeological site
The Puna is a high desert biome located above 3500 masl and characterized by an arid and cold climate, intense solar radiation due to altitude, high thermal amplitude day/night, a marked seasonality with poor summer rains, and low atmospheric pressure (Bruniard, 1999). Vegetation is scarce and mainly xerophytic with the presence of peat-bogs (vegas) on the banks of small permanent rivers. In terms of precipitation distribution, the Argentinean Puna exhibits a latitudinal gradient that determines two sub-regions: the Dry or Northern Puna and the Salt or Southern Puna. While both have similar vegetation compositions, the Salt Puna is dominated by salt-lakes and saline soils (Cabrera, 1976).
Antofagasta de la Sierra is located in the Northwestern corner of Catamarca Province (Argentina), within the Salt Puna. The locality includes a diversity of landforms and a mosaic of resources and microenvironments. Also, it belongs to an endorheic basin from the rivers Toconquis-Calalaste-Punilla that debouch in the Antofagasta Lake, surrounded by two volcanoes and a lava field. One of the tributaries of Punilla river is Las Pitas river, where in their margins is located the archaeological site Punta de Peña 4 (Figure 1). The area presents extreme aridity (andino-puneño arid clime) and precipitations of summer regime lower than 100–150 annual mm. The mean annual temperature is 9.5°C with great thermal amplitude during the day and the stations, and low atmospheric pressure. The vegetation of the region belongs to the Puna Province of the Andean Domain, where shrub steppe dominates, and there is presence of halophilic, herbaceous steppes and wetlands areas (vegas) (Cabrera, 1976).

Archaeological sites from Antofagasta de la Sierra (Catamarca, Argentina). Location of Punta de Peña 4 (PP4) archaeological site is highlighted with a star. Modified from Urquiza and Echeverría (2018).
The archaeological site Punta de Peña 4 (henceforth, PP4) is located at the edge of an ignimbrite outcrop at 3.650 masl, in the middle course of the Las Pitas river (26°1′40.26″S; 67°20′33.17″W). Is a rock overhang which exhibits rock art painting and engravings of diverse chronology and styles (Urquiza and Aschero, 2014, and references therein). The site presents a large occupational sequence that includes early Holocene (ca. 8900–8300 years14C BP), late-Holocene (ca. 4500–3200 years14C BP) until historic times (500 years 14CBP), with the last occupation as a corral in the twentieth century in the covered sector of the shelter. Since about 40 years up to date, the site has not been occupied by humans or animals. Furthermore, the stratigraphy of the site remained undisturbed. Thus, the possibility of modern contamination of samples can be dismissed. In the site, humans made wide use of camelids as a source of meat, skin, coats, and bones.
Initially, PP4 was inhabited by archaic groups which were hunters of wild camelids, and prey processing possibly took place in an open-air sector, and area that through the stratigraphic sequence has been a dump. Later, agropastoral groups raised llamas and hunted vicuñas; this group would have developed the prey processing within the rock overhang. The zooarchaeological assemblages showed that preys arrived almost complete at the site. Most recently, during the Historical period, PP4 was used as a seasonal corral (Urquiza and Aschero, 2014).
Among wild camelid remains, guanacos predominated only in early stratigraphic levels. The vicuñas were the most exploited camelids throughout the stratigraphic record, and were more relevant in late levels, where the proportion between wild and domestic camelids was similar, with a high rate of juvenile individuals (Urquiza and Aschero, 2014).
Paleoparasitological study
A total of 65 coprolites assigned to SAC from PP4 were examined for parasites remains. Coprolites were obtained from different microsectors within covered and uncovered sectors of the site (Table 1). The external observation of faeces (color, texture, inclusions, and measurements) was conducted according to Chame (2003). Coprolites were fully processed by rehydration in a 0.5% water solution of tris-sodium phosphate solution for 72 h (Callen and Cameron, 1960). Subsequently, following the methodology proposed by Bouchet et al. (2003), samples were homogenized with the aid of a glass rod, filtered and allowed to spontaneous sedimentation. The sediment was recovered with a pipette and a total of 20 slides for each sample were observed. All slide observations were conducted with the addition of one drop of glycerin. Eggs and oocysts of the parasites were measured and photographed at a magnification of 400×. The measurements were all taken from well-preserved eggs.
Parasite species found in camelid coprolites from PP4 site.

UGA: Center for Applied Isotope Studies (CAIS), University of Georgia, EE.UU. Calibrated ±2 sigmas using Oxcal 4.1.
For this level only the covered sector was dated. Tentatively is used the same dating for the uncovered sector.
Egg preservation
In order to evaluate the state of preservation of the eggs and oocysts, a modified scale of the categories proposed by Kumm et al. (2010) was employed. The categories used in this study were:
(1) Pristine = undamaged egg and, if applicable, polar plugs or operculum intact.
(2) Poor = deformed egg.
(3) Fractured = egg with a crack or tear.
(4) Fragmentary = egg broken in two or more pieces.
Results
The studied coprolites were oval, compact, and hard and exhibited light-brown coloration. The surfaces were smooth. The measures of the coprolites were 10.52 ± 1.96 mm in length × 8.11 ± 1.27 mm in width (n = 54).
A total of 29% of the samples (19/65) were positive for parasite remains. Eleven taxa of parasites were found (Table 1). Three taxa belong to Apicomplexa, one to Platyhelminthes, and seven to Nematoda.
Coccidian oocysts were found in the samples. The most abundant oocyst observed present a thick wall, brown-reddish coloration, and a prominent micropyle and was found in coprolites no. 5, 8, 10, and 53 (Figure 2a). The measurements of the oocysts were 89.81 ± 5.1 µm in length × 64.09 ± 4.55 µm in width (82.5–112.5 µm × 57.5–82.5 µm, n = 66). These oocysts were compatible with E. macusaniensis (Apicomplexa, Eimeriidae). One coccidian oocyst (sp. 1) with four sporozoites was found in coprolite no. 2 (Figure 2b). The measurements were 37.5µm in length × 37.5 µm in width. One coccidian oocyst (sp. 2) with two sporozoites was found in coprolite no. 33 (Figure 2c). The measurements were 32.5 µm in length × 30 µm in width (n = 1).
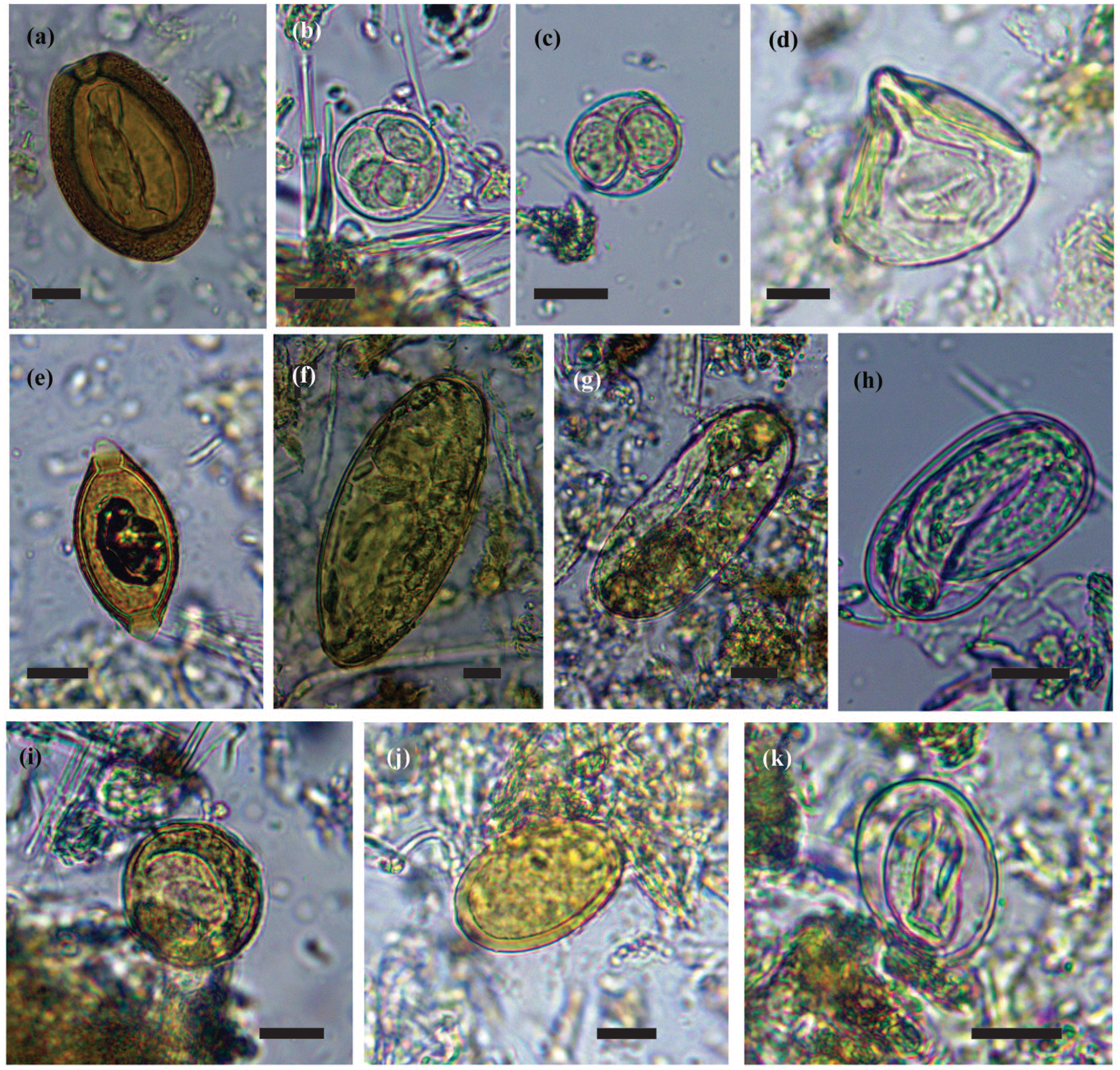
Photograph of parasite species recorded in PP4 archaeological site. (a) Eimeria macusaniensis, (b) coccidia sp., (c) coccidia sp., (d) Moniezia sp., (e) Trichuris sp., (f) Lamanema chavezi/Nematodirus lamae, (g) Strongylus-type egg, (h) Strongyloides sp., (i) Nematode undet. sp.1, (j) Nematode undet. sp.2, and (k) Nematode undet. sp.3. Scale bar 20 µm.
Anoplocephalid eggs were found in samples no. 2, 4, 8, 15, and 42. The eggs were tri- or quadrangular shaped with a piriform apparatus (Figure 2d). The measurements of the eggs were 66.9 ± 5.1 µm in length × 60.3 ± 6.8 µm in width (57.5–77.5 µm × 47.5–75 µm, n = 59). The eggs were attributed to Moniezia sp., possibly Moniezia benedeni or Moniezia expansa (Platyhelminthes, Cestoda, Cyclophyllidea. Anoplocephalidae). Both species are capable of parasitizing the same hosts but are very difficult their identification (Chilton et al., 2007).
Trichurid eggs were found in coprolites no. 4, 5, 6, 10, 16, 42, and 46 (Figure 2e). The eggs were barrel shaped, thick walled, and presented two polar plugs. The measurements were 69.7 ± 4.3 µm in length × 36.6 ± 3.5 µm in width (62.5–77.6 µm × 30–47.5 µm, n = 19). This egg was attributed to Trichuris sp. (Nematoda, Adenophorea, Trichurida, Trichuridae).
Large trichostrongylid eggs were found in coprolites no. 4, 45, and 48. The eggs were ovoid, thin-shelled, brownish, and with parallel sides (Figure 2f). Their measurements were 167.16 ± 6.8 µm in length × 80.83 ± 4.7 µm in width (158–175 µm × 75–87.5 µm, n = 6). The eggs were attributed to Lamanema chavezi or Nematodirus lamae (Nematoda, Strongylida, Trichostrongyloidea, Molineidae) due to the morphology of the eggs.
Two species of Strongylus-type egg were recovered. One egg was found in coprolite no. 1 (Figure 2g), the measurements were 107.5 µm in length × 45 µm in width (n = 1). Besides, Strongylus-type eggs were found in coprolites no. 18, 26, and 33. The eggs were thin shelled, slightly flattened poles and contained a developed larva. The measurements were 65 ± 4.6 µm in length × 33.9 ± 2 µm in width (57.5–75 µm × 30–37.5 µm, n = 16) (Figure 2h). Based on their morphology, these eggs were attributed to a Strongylus-type, possibly Strongyloides sp. (Nematoda, Rhabditida, Strongyloididae).
Two unidentified nematode eggs were found in coprolite no. 2 (Figure 2i). The eggs were ovoid, with a thin and brownish shell and were larvated. The measurements were 47.5 ± 0 µm in length × 45 ± 3.5 µm in width (47.5 µm × 42.5–47.5 µm, n = 2). Other unidentified nematode species was found in coprolites no. 3, 4, and 6 (Figure 2j). The eggs were elongated and presented a thick and brownish shell. The measurements were 60 ± 2.9 µm in length × 36.25 ± 4.3 µm in width (57.5–62.5 µm × 30–40 µm, n = 4). Finally, a third type of unidentified nematode egg was found in coprolite 61 (Figure 2k). The egg was ovoid, translucent, and larvated. The measurements of the egg were 40 µm in length × 30 µm in width (n = 1).
Concerning to the state of preservation of the eggs and oocysts, most of them (141/146) were classified as pristine eggs. The 1.4% of the eggs (2/146) was classified as poor preserved and 2% (3/146) as fractured. Regarding these last five eggs, one poor preserved egg was found in coprolite no. 33 belonging to an uncovered sector. In the other hand, three poor preserved or fractured eggs were found in coprolites no. 4 and no. 8, belonging to the covered sector. No fragmentary eggs or oocyst were found. Most of the eggs presented a good state of preservation. Therefore, no differences in egg preservation between covered and uncovered sectors of the site were evidenced.
Discussion
Given the morphology of the coprolites, the biogeographic origin and the presence of coccidian oocysts of E. macusaniensis in some of them, a camelid origin of the coprolites was confirmed. Among zooarchaeological remains of vertebrates found in PP4, SAC remains were the predominant, with llama remains (17.94%) followed by vicuña remains (15.85%), and lastly by guanaco remains (5.15%). The vicuña is present all along the chronological sequence (ca. 8970–540 BP), the llama is present in levels 1–4 (ca. 4060–540 BP) and, the guanaco is abundant in levels 5 and 6 (ca. 8970–4000 BP) (Urquiza and Aschero, 2014). The late occupation (ca. 760–460 BP) contains a level of herbivore dung which would belong to vicuñas and llamas. Llamas and vicuñas were predominant species of artiodactyls in the zooarchaeological record of PP4, being the most probable hosts of the parasitic fauna registered. However, it is not possible to confirm due to the similarity of the coprolite morphology among SAC.
At least five species of Eimeria have been reported in SAC: E. lamae, E. alpacae, E. punoensis, E. macusaniensis, and E. ivitaensis (Dubey, 2018). Eimeria macusaniensis is a specific parasite of SAC and is considered the most pathogenic. The host specificity of this coccidian is a useful indicator for camelid origin of faeces (Dubey, 2018). The life cycle of this species is direct with faecal-oral transmission. This eimeriid was previously registered in SAC from Argentina in current studies (e.g. Beldomenico et al., 2003; Borghi et al., 2004; Cafrune et al., 2009, 2014; Gonzalez-Rivas et al., 2019; Moreno et al., 2015). In ancient times, this coccidian species was recorded in SAC coprolites from Casa de Piedra 5 and 7 (CCP5 and 7) and Alero Destacamento Guardaparques (ADG), both archaeological sites from Patagonia Argentina. There was also recorded in humans and carnivore coprolites related to camelid consumption (Amalfitano et al., 2017; Beltrame et al., 2010; Fugassa, 2007; Fugassa et al., 2010; Taglioretti et al., 2015, 2017). These reports indicate the presence of this species since the early Holocene until the present in Patagonia Argentina. Its presence was also reported in human coprolites related to camelid consumption from northern Chile dated to Pre-Inca Hispanic Contact Period (de Souza et al., 2018) and from coprolites and intestinal contents of SAC from the archaeological site of Huanchaquito (Perú) (Le Bailly et al., 2020).
Cestodes of the family Anoplocephalidae are common parasites of reptiles, birds and mammals. Anoplocephalidae presents more than 165 species. The genus Moniezia is the most important because it has a cosmopolitan distribution and causes economic losses in domestic ruminants (Denegri et al., 1998; Schuster et al., 2000). Moniezia species are the most common intestinal tapeworms in ruminants. There are two species of Moniezia that parasitize cattle, goats, sheep, and camelids: M. expansa and M. benedini (Kaufmann, 1996).The life cycle requires oribatids as intermediate hosts and ruminants as definitive hosts. Infection of the latter hosts is initiated by ingesting the infected mites containing cysticercoids (Diop et al., 2015; Jyoti et al., 2014). In current studies, the presence of Moniezia parasitizing SAC has been associated with the presence of domestic ruminants in the area, probably sharing grazing areas (Beldomenico et al., 2003; Moreno et al., 2015). However, this is the first time that Moniezia eggs are found in ancient times from SAC coprolites. Therefore, this finding confirms the presence of Moniezia in SAC at least since the middle to late-Holocene, prior to European cattle arrival. This is an interesting contribution to the study of the biogeographic history of this anoplocephalid.
Species of Trichuris (Nematoda: Trichuridae) have a cosmopolitan distribution and parasitize a broad range of mammalian host, such as ruminants, marsupials, rodents, and primates, including humans (Anderson, 2000). The only species of the genera Trichuris recognized in SAC is Trichuris tenuis. The first report of this species in vicuñas from Argentina was in 1996 and is a species frequently registered in camelids. This suggests that this species is primarily a camelid parasite and is found in SAC (Cafrune et al., 1999). Since their first record, it has been recorded parasitizing llamas, vicuñas, and guanacos (Beldomenico et al., 2003). Besides T. tenius, another trichurid species have been registered in SAC, T. ovis, both species has similar egg sizes so, differentiation among them is impossible. In Argentina, eggs of Trichuris were found in camelid coprolites presents in archaeological contexts in the arid steppe of Patagonia (CCP7 and ADG sites; Amalfitano et al., 2017; Taglioretti et al., 2017) and the mountain area of Catamarca (Alero Los Viscos site; Fugassa et al., 2018). Also, were reported in domestic camelids from the mountain area of Peru in the Huanchaquito archaeological site (Le Bailly et al., 2020).
Most of the nematodes that are commonly found in SAC belong to order Strongyloidea, (Strongylidae and Trichostrongylidae). Several genera of strongylid-type nematodes affect SAC (Guerrero, 1960; Guerrero and Leguía, 1987; Leguía and Casas, 1998). In this study, different types of strongylid eggs were found. One of the eggs was attributed to L. chavezi/N. lamae, both specific nematodes of SAC (Cafrune et al., 2001). Both species have similar eggs and differentiation is very difficult. In previous paleoparasitological studies in Patagonia, N.lamae or L. Chavezi eggs were registered in coprolites from the sites CCP5 and CCP7, and in some samples from AGP, being the most representative species (Taglioretti et al., 2015).
Strongyloides is a genus containing about 50 species of obligate gastrointestinal parasites of mammals, birds, reptiles, and amphibians (Speare, 1989). This genus has been recorded parasitizing in wild SAC from Argentina in the provinces of Chubut and Mendoza (Karesh et al., 1998; Moreno et al., 2015) and in captive or domestic SAC in Chaco and Catamarca (Cardozo, 2019; Martinez et al., 2012). This genus is shared among wild and domestic ruminants in the region. At the moment, this species has not been found in ancient SAC. Their presence in Holocene samples contributes to the knowledge of the biogeographic history of Strongyloides and confirms its presence in prehispanic times.
The absence of capillarid eggs in the present study is striking. Regarding this point, Fugassa et al. (2018) neither found capillariid eggs in camelids from the archaeological site Alero Los Viscos from Catamarca. Nevertheless, in Holocene samples from Patagonia capillariid eggs have been found frequently in SAC (Amalfitano et al., 2017; Fugassa, 2007; Taglioretti et al., 2015, 2017). In current studies capillariid eggs have been frequently found, in llamas, alpacas and vicuñas (Bouts et al., 2003; Cafrune et al., 2004; Valenzuela et al., 1998; among others).
Regarding the host-parasite relationship through time, in the oldest occupation (8970 ± 6 years BP) there was found one egg of an unidentified nematode (five analyzed samples). Later, in the late-Holocene (ca. 4560 ± 60–4100 ± 160 years BP) there were identified four parasite taxa (21 analyzed samples). During this time, the covered sector delimited a housing space whose first occupation (4560 ± 60 BP) displayed a predominance of guanacos hunting. Around the 990 ± 40 and 960 ± 40 years BP (21 analyzed samples), six parasite taxa were registered. During this stage, were observed areas of activities (such as processing of fibers and lithics) in the covered sector and discard concentrations in the uncovered sector. From this period up to the end of the sequence, the proportion of llama bone remains is comparable to wild camelids (vicuñas) (Urquiza and Aschero, 2014). Finally, in the most recent period of PP4 (540 ± 40–470 ± 50 years BP), seven parasite taxa were found (18 analyzed samples). In this period, the site was a settlement of camelid herders, and bone remains of domestic (llama) and wild camelids (vicuña) were found. A microhistological study of camelid dung from this period showed that camelid used high Andean wetlands (vegas) and dry areas (tolar and grassland) as foraging areas (Mosca Torres et al., 2018). Despite there were not found differences in the state of preservation of the eggs between covered and uncovered sectors of the site, the number of parasite taxa seems to increase through time until historic times. This finding is an interesting point to be discussed. As was previously mentioned, in prehispanic times, llamas and alpacas were the most important factor in Andean economies and were important livestock of some regions (Mengoni Goñalons, 2008; Mengoni Goñalons and Yacobaccio, 2006). One possible reason for the increase of parasite taxa in more recent times could be related to the domestication of SAC. The presence of domestic camelids (L. glama) in the site was observed from 4000 years B.P. to recent times (Urquiza and Aschero, 2014). According to the OIE (World Organization for Animal Health, 2019), a major constraint to increasing global livestock productivity in current times is loss due to parasite diseases. The more intensive breeding of SAC in recent times could increase overcrowding and, consequently increased the chance of infection. However, it is an observation that needs further studies and is an interesting point to be studied in future works.
Urquiza (2016) found differences in preservation features of bones and teeth from SAC in PP4, which were probably related to the overhang hydrogeologic dynamic, where the most affected were the oldest remains. Nevertheless, in regard to the state of preservation of parasite remains, most of the eggs presented excellent preservation, and no differences between covered and uncovered sectors of the site were found. One or more variables may influence the preservation of zooarchaeological remains in sites of the Puna. The results suggest that taphonomic factors act differently on the diverse zooarchaeological remains from PP4.
Conclusions
This is the first paleoparasitological study from SAC coprolites recovered from PP4, an archaeological site of Antofagasta de la Sierra region in the northwest of Argentina. SAC samples presented a parasitic fauna that agrees with previous modern and paleoparasitological studies in the area. The present study displayed that, the parasitic diversity of camelids in the past was similar to that of current times. Specific parasite species of SAC were found, such as E. macusaniensis and L. chavezi/N. lamae. A remarkable finding was the presence of Moniezia sp. and Strongyloides sp. in coprolites from different levels of PP4, pointing out the presence of these genera in native artiodactyls of the regions previous to the arrival of Hispanic societies. This is the first time that Moniezia and Strongyloides eggs are found in ancient times from SAC coprolites, resulting in an interesting contribution to the study of the biogeographic history of these species. In addition, another remarkable aspect to be mentioned is that none of the parasitic species found are considered zoonotic species; therefore its presence in the environment was not a risk to the health of ancient human populations of the area. Besides, results displayed a change in the number of parasite taxa present in SAC through time. Further studies are necessary to continue with the development of these interesting findings.
Footnotes
Acknowledgements
We are very grateful for comments from the anonymous reviewers who help to improve the manuscript.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was supported by grants FONCyT-PICT 0316 (M.O. Beltrame), FONCyT-PICT 1160 (S. Urquiza), PIUNT G503 Universidad Nacional de Tucumán (C. Aschero), and PIP-CONICET 577 (C. Aschero).