
Abstract
Coprolites are one of the sources of evidence most commonly used in paleoparasitological studies. However, the zoological origin identification is not always possible. Parasites of the genus Eimeria (Apicomplexa: Eimeriidae) are obligate protozoan which have evolved to exhibit a great diversity of host range, and present strict host specificity. Usually, the identification of Eimeria spp. can be done by morphological characteristics such as size and shape of the oocysts. Moreover, molecular diagnostic methods such as PCR have proven to be very useful methodologies for an accurate taxonomic identification. In recent years, molecular techniques have started to be applied in paleoparasitological studies. Here, we provide the first integrative approach that combine light microscopic studies and ancient DNA (aDNA) in order to detect and identify Eimeria spp. on artiodactyl coprolites recovered from an archeological context from northwestern Patagonia, Argentina. The main objective is to discuss the advantages of complementing these two approaches as a promising tool to identify the zoological origin of coprolites. Furthermore, we provide paleoparasitological data about the presence of Eimeria spp. in coprolites of South American ungulates. Light microscopic studies allowed the identification of oocysts attributed to Eimeria spp. in 13 of the 40 coprolites. Amplification and sequencing of the aDNA ITS1 detected the presence of Eimeria spp. in 5 of the 10 samples, and allowed the taxonomic identification of these sequences as belonging to Eimeria arloingi. Furthermore, the results obtained from cytochrome b gene (Cytb) analysis confirmed the identity of the host as Capra hircus. Results displayed the presence of coccidiosis in goats from historical times. This study highlights that both techniques are complementary and promotes the use of the identification of Eimeria spp. in ancient samples in order to identify the zoological origin.
Introduction
Coprolites (or paleofeces) are desiccated or mineralized feces recovered from archeological and paleontological sites around the world, and have a great potential for paleoecological studies. In recent years, coprolite studies are becoming increasingly multidisciplinary and can provide multiple lines of evidences (Chame and Sianto, 2014; Wood and Wilmshurst, 2016). Coprolites are the source of information most commonly used in paleoparasitological studies, together with mummied remains (Camacho et al., 2018). Parasites remains recovered from these contexts are studied from an ecological approach with a focus in the study of the evolution of parasite–host–environment relationships, as well as the origin and evolution of infectious diseases within a paleoepidemiological perspective (Araújo et al., 2003; Reinhard et al., 2013). However, a difficulty presented by coprolites is that it is not always possible to identify their zoological origin.
Parasites of the genus Eimeria (Apicomplexa: Eimeriidae) are known as an important cause of intestinal infection in animals around the world. Eimeria spp. are obligate protozoan parasites which have evolved to exhibit a great diversity of host range including mammals, fish, birds, amphibians, and reptiles, but present strict host specificity. This genus is composed by more than 1200 species and this number continues to grow (Chapman et al., 2013; López-Osorio et al., 2020). These protozoa are responsible for the so-called “coccidiosis,” which represent a very important disease in vertebrate animals (Mohamaden et al., 2018). Traditionally, the identification of Eimeria spp. has been made on the basis of the morphological characteristics of the sporulated oocysts and host specificity. However, morphological techniques show relatively low sensitivity and practical limitations due to the similar morphologies and overlapping sizes of the oocyst stage. Molecular diagnostic methods such as the polymerase chain reaction (PCR) have proven useful methodologies for the identification and classification of Eimeria species. Commonly used gene targets are ribosomal genes including the 18S rDNA, internal transcriber spacer (ITS) sequences 1 and 2, as well as genes contained within the mitochondrial genome, such as cytochrome c oxidase subunit I (COI) or cytochrome b gene (Al-Habsi et al., 2017; Blake et al., 2020; Ogedengbe et al., 2015; Silva et al., 2017).
In recent years, molecular techniques have started to be applied in paleoparasitological studies. Nowadays, the additional benefits of combining paleogenetic studies with traditional microscopic studies are well known (e.g. Côté and Le Bailly, 2018; Kévin et al., 2021; Petrigh et al., 2021; Welker et al., 2014; Wood et al., 2013, 2019). However, so far, the molecular approach has not been used in the diagnosis of coccidian species from ancient samples. In the present work, we provide the first integrative approach of light microscopic studies and ancient DNA (aDNA) studies to detect and identify Eimeria spp. on artiodactyl coprolites recovered from an archeological context from northwestern Patagonia, Argentina. The main objective of this contribution is to discuss the advantages of complementing these two approaches as a promising tool to identify the zoological origin of coprolites.
Material and methods
Study site
The studied samples were recovered from the archeological site “Cueva Huenul 1” (CH1). This site is a large cave emplaced at 1000 m.a.s.l. in Neuquén Province, northwestern Patagonia (Argentina, 36°56′ S, 69°47′ W). It is currently located in a desert environment within the Monte phytogeographical Region, a scrubby formation typical of northern Patagonian deserts (Abraham et al., 2009; Páez et al., 2004). Preliminary pollen analysis of stratigraphic samples from CH1 show the presence of a Patagonian grass-shrub steppe integrated with shrubs (Schinus, Lycium), dwarf shrubs (Nassauvia, Ephedra), and grasses, which at present lie between 2000 and 2200 masl, suggesting colder than present local conditions since 13,800 years B.P. (Pompei et al., 2012), thus confirming tendencies observed on a broader scale (de Porras, 2017; Llano et al., 2020; Markgraf et al., 2009; Whitlock et al., 2006). Through the last 1500 years, however, pollen record indicates the presence of a Monte-Patagonian transition shrub steppe, suggesting that present environmental and climatic conditions prevailed (Pompei et al., 2012). The analysis of rodent assemblages from the site shows relatively stable ecological conditions since the Late Pleistocene with the presence of a mosaic of shrubby steppes and open rocky areas, consistent with the current Monte phytogeography (Fernández et al., 2012; Fernandez et al., 2021).
The site CH1 has been studied in the context of a regional archeological project (Barberena, 2015; Rughini et al., 2021) and provides a well preserved—but discontinuous– sedimentary sequence encompassing between ~16,000 and 300 cal years B.P. The site was excavated between 2010 and 2014 utilizing 10-cm artificial levels and discriminating the sedimentary layers. The excavations provided a 1.4-m sequence composed of four chrono-stratigraphic components. Component 1 encompasses the stratigraphic units VIII through V (artificial extraction levels 13 through 6), dated between 16,600 and 13,600 cal years B.P. The existence of rodent and anthropic disturbances on the top of these basal layers (VIII to V) makes it likely that the few lithic artifacts recovered from these layers migrated downwards. Hence, this component is interpreted as largely produced by natural agents (Barberena et al., 2015). Component two includes the stratigraphic unit IV (artificial extraction layers 5 and 4) and is deposited immediately above an erosive unconformity (hiatus 1). It presents the first well recorded human occupations at the site and is dated between 11,300 and 10,200 cal years B.P. Component 3 corresponds to an anthropic pit structure dated between 5600 and 5300 cal years B.P. Finally, the component 4 includes the stratigraphic units III, II and I (artificial extraction levels 3 through 1), dated between 1500 and 300 cal years B.P. It marks the lapse of most intense human occupation at the site and the region (Barberena et al., 2015).
Regarding the paleoparasitological analyses, well-preserved megafauna dung remains and wild mammals’ coprolites were recovered from chronologically bracketed late Pleistocene-early Holocene stratigraphic deposits. Rodents, camelid, felid and megamammal feces from CH1 were already studied for paleoparasitological purposes (Beltrame et al., 2012, 2016, 2020; Taglioretti, 2015; Tietze et al., 2019).
Microscopic studies
Forty coprolites recovered from artificial levels 1, 3, and 4 (i.e. Components 2 and 4 of the chrono-stratigraphic sequence) from CH1 were studied. Examination of the external shape of feces was conducted according to Chame (2003). Samples were whole processed and were rehydrated in a 0.5% water solution of tris-sodium phosphate in a glass tube for at least 72 h (Callen and Cameron, 1960), followed by homogenization. After this period, samples were sieved through thrice-folded gauze for spontaneous sedimentation (Lutz, 1919) and preserved in 70% ethanol. Twenty slides per sample were observed, along with the addition of one drop of glycerin and examined at 10× and 40× by light microscopy (Zeiss® Primo Star). Coccidian oocysts were measured and photographed at ×40 magnifications and their dimensions and morphologies were compared with data from the literature. The macroscopic remains were separated and dried at room temperature for diet analysis.
aDNA extractions, PCR amplification, and sequencing
A subset of 10 samples belonging to the different studied levels from CH1 was submitted to molecular studies (Table 1). Sample processing was carried out under strict standard protocols.
Microscopic and paleogenetic results obtained to Eimeria spp. presence in a subset of 10 artiodactyl coprolites from Cueva Huenul 1 archeological site, Patagonia, Argentina.

ND: not detectable.
Confirmed by Sanger sequencing; cytb: confirmed by sequencing of cytochrome b gene.
Total genomic aDNA was extracted from the rehydrated sediment of each sample using ADN-Puriprep K1210 Kit (INBIO Highway, Argentina) following manufacturer’s instructions; a negative extraction control was included in order to discard reactive contamination with modern DNA. Obtained DNA was quantified in a DeNovix DS-11 FX Spectrophotometer (DeNovix Inc., Wilmington, USA). The whole processing was conducted applying specific protocols to minimize contamination risk with modern or exogenous DNA. A UV-cabinet was used.
For molecular Eimeria identification, samples were subjected in triplicates to real-time polymerase chain reaction (qPCR) with generic primers (ITS1-F-3’-GGGAAGTTGCGTAAATAGA-5’ and ITS1-R CTGCGTCCTTCATCGAT) that amplify all Eimeria species (Oliveira et al., 2011). The thermal cycling conditions were as follows: an initial denaturation of 3 min at 95°C, and 45 cycles of 94°C 25″, 50°C 30,” 72°C 40″. After amplification, a melting curve analysis to confirm specific amplification of the PCR product was performed. All qPCR reactions were carried out in a thermocycler QTower3 (Analytic Jena, Germany) in a final volume of 20 µl using EvaGreen as fluorescent intercalating dye (KAPA Fast, Biosystems, Woburn, USA). Negative and positive controls were included in each PCR run. To corroborate size and amplification specificity, PCR products were resolved on agarose gel. Amplified DNA fragments were purified using the Accuprep gel purification kit (Bioneer, South Korea) and directly sequenced (ABI 3500 Genetic Analyzer, Applied Biosystems, Foster City, CA, USA). Consensus sequences between replicates were built for each sample. Once available, sequences were submitted to the GenBank database (NCBI). The sequences similarities were determined by the Basic Local Alignment Search Tool (BLAST) (https://blast.ncbi.nlm.nih.gov/Blast.cgi) of the National Center for Biotechnology Information (NCBI).
Moreover, in order to identify the zoological origin of coprolites, a 358 bp of mitochondrial Cytochrome b (cytb) gene was amplified from DNA extractions with primers BM1 (5′CCCCTCAGAATGATATTTGTCCTCA3′) and BM2 (5′CCATCCAACATCTCAGCATGATGAAA3′) and sequenced (Soares et al., 2014). Additionally, samples were subjected to a specific Capra hircus PCR described by Santaclara et al. (2007) with primers that amplify a 192 bp PCR fragment.
Results
Macroscopic and microscopic studies
Coprolites were dark brown, cylindrical, usually pointed at one end and concave in the other extremity with smooth surfaces; they were in an excellent state of preservation (Figure 1). Average measurements of feces (N = 40) were 16.85 ± 3.31 mm long by 9.41 ± 1.25 mm wide. The rehydrated color was light-brown and had a domestic cattle-like odor. Macroscopic observations showed vegetal remains. The zoological origin was attributed to an herbivore, belonging to the Order Artiodactyla.

Morphological aspect of artiodactyl coprolites studied from the archeological site Cueva Huenul 1, Patagonia, Argentina.
A total of 32.5% of the samples (13/40) were positive for coccidian oocysts. All of them were identified as Eimeria sp. (Figure 2a–g) with the exception of one coccidian which could not be identified (Figure 2h). In each positive sample for Eimeria sp., only one eimeriid species was present.
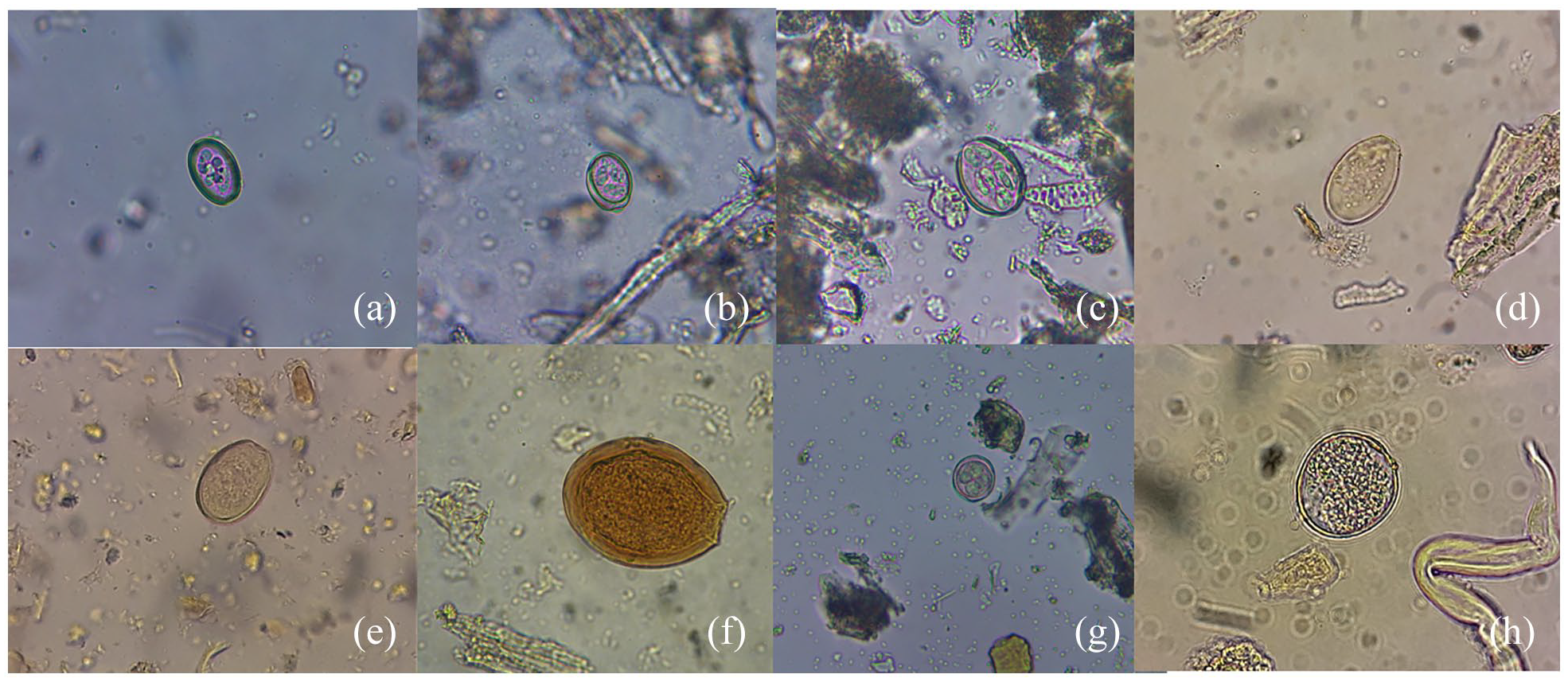
Coccidian oocysts (Coccidia, Apicomplexa) found in artiodactyl coprolites from the archeological site Cueva Huenul 1, Patagonia, Argentina. (a–e and g) attributed to Eimeria spp., (f) Eimeria macusaniensis, (h) unidentified oocyst.
The most abundant oocyst observed was translucent, oval-shaped with a small micropyle. Most of them were sporulated and contained four sporocysts. The measurements were 28.3 ± 3.4 µm in length × 21.4 ± 3.5 µm in width (n = 21). These oocysts were observed in six samples and were attributed to Eimeria sp. (Figure 2a). Four oval-shaped oocysts, translucent, sporulated with four sporocysts and with small micropyle were found in another sample. The measurements were 33.1 ± 3.8 µm in length × 22.4 ± 0.3 µm in width (n = 4). These oocysts were assigned to Eimeria sp. (Figure 2b). One oocyst, translucent, oval-shaped, sporulated, with small micropyle and eight sporozoites was also observed. The measurements were 45.0 µm in length × 30.0 µm in width. The oocyst was attributed to Eimeria sp. (Figure 2c). In one sample were recorded three coccidian oocysts. They were yellowish, oval shaped, with micropyle and were not sporulated. The measurements (length by weight) were 37.5 × 25 µm, 40 × 30 µm, and 47 × 30 µm (Figure 2d). They were assigned to Eimeria sp. In addition, three yellowish, oval-shaped and not sporulated oocysts were also found, and were attributed to Eimeria sp. The measurements (length by weight) were 30 × 22.5 µm, 35 × 22.5 µm, and 37.5 × 25 µm (Figure 2e). One oocyst with a thick wall, brown-reddish coloration and a prominent micropyle was found in one coprolite. Measurements of the oocyst were 97.5 µm in length × 75.0 µm in width. The oocyst was compatible with Eimeria macusaniensis (Figure 2f). Two coccidian oocysts with four sporocysts, small micropyle and round shape were found in other sample. The measurements were 33.0 µm in length × 22.5 µm in width and 25.0 µm in length × 22.5 µm in width. They were attributed to Eimeria sp. (Figure 2g). One translucent oocyst, not sporulated, with small micropyle and oval shaped was found. The measurements were 40.0 µm in length × 28.0 µm in width. This coccidian oocyst could not be identified (Figure 2h).
Paleogenetic studies
The presence of Eimeria spp. was studied by qPCR in 10 samples; positive samples (1, 4, 7, 8, and 9) were sequenced and BLAST analyzed (Table 1). Samples 1 and 7 showed 96%, 75%, and 98% homology with Eimeria arloingi respectively. Samples 4, 8, and 9 showed an identity range of 83%–97% with the following Eimeria species: E. arloingi, E. parva and E. ovina, respectively.
An informative sequence of the cytb gene amplified from three of the analyzed coprolites was identified by BLAST analysis as the domestic goat, Capra hircus (Artiodactyla: Bovidae), with 99.19% homology. All analyzed samples showed amplification product with Capra hircus specific primers (Table 1).
Discussion
In the present study, the diagnosis based on morphometric and morphologic analyses of coprolites samples, in addition to the odor and rehydrated color, allowed the primary identification of the taxonomic group of the depositing species. This first approach (described in results) made it possible to assign the coprolites to Artiodactyla. In the study area, this Order is represented by camelids, specifically guanaco (Lama guanicoe) present in this region throughout the Holocene until recent times. This species is one of the most represented in the zooarchaeological record in the region (Rindel et al., 2021). More recently, from historical times to the present, several artiodactyls species were present in the South American extensive Phylogeographical regions of Monte, Espinal, Patagonian desert, and other Andean landscape configurations; not only with the presence of guanacos but also with the introduced goats and sheep. These species arrived to South America after the arrival of Europeans in the 15th century (Lanari et al., 2003). Deer remains were not observed in the site in recent times nor in the past (Barberena et al., 2015), although were registered in archeological sites throughout Patagonia, most of them located in or near Andean Forest ecosystems (Fernández et al., 2015). Based on this first approach, the coprolites could have been deposited by goats, deer or camelids. However, the differentiation of these artiodactyl feces is readily not possible due to their similar shape and size.
Eimeria spp. are differentiated according to their morphology and size. However, morphological techniques have been described as having relatively low sensitivity (Chapman et al., 2013). In the present work, light microscopic studies allowed the identification of oocysts attributed to Eimeria spp. However, the identification of the species was not possible in any case. Only one sample presented one oocyst identified as E. macusaniensis. This coccidian is a specific parasite of South American camelids. The host specificity of this parasite is a useful indicator for the camelid origin of feces (Dubey, 2018). However, the finding of a single oocyst of E. macusaniensis, together with the paleogenetic results, suggests that its presence is due to contamination.
Amplifications of aDNA ITS1 used in this study displayed the presence of Eimeria spp. in 5 of the 10 studied samples. Some of them were previously identified by light microscopy as positive and other as negative. Sequencing analysis allowed the identification as E. arloingi with 99% homology. Since this specie is a specific parasite of the goat, the zoological origin of the coprolites can be confidently attributed to this artiodactyl even in the case of a lack of host confirmation of the origin of the samples. To date, 17 Eimeria species have been reported in goats with nine of major epidemiological and veterinary importance. E. arloingi, E. ninakohlyakimovae, E. christenseni, and E. caprina are considered to be the most pathogenic species. As was previously mentioned, molecular diagnostic methods such as PCR have proven useful for the species identification and classification of Eimeria (Ogedengbe et al., 2015). However, this is the first report in which aDNA was used to identify Eimeria spp. in ancient samples from archeological contexts. Eimeria arloingi has a worldwide distribution (Chapman et al., 2013). This coccidian was reported previously from Argentina and others countries from South America (e.g. Macedo et al., 2019). This is the first time that was it registered in fossil assemblages. On the other hand, the results obtained from the cytochrome gene analysis confirmed the identity of the host as C. hircus.
BLAST analyses of samples 4, 8, and 9 displayed the presence of E. arloingi/ ovina/parva. Currently, 11 Eimeria species are recognized as causing sheep infections (Andrews, 2013). Eimeria parva and E. ovina are specific species of sheep (Chartier and Paraud, 2012). Sheep and goats harbor their own species of Eimeria and there is no cross-infection (McDougald, 1979). Therefore, according to the specificity of Eimeria and the paleomolecular results, the records presented here can be attributed to E. arloingi.
Coccidiosis (Eimeriosis sensu stricto) is a protozoan infection of small ruminants worldwide caused by parasites of the genus Eimeria, which develop and propagate in the small and the large intestines of animals and affect young age particularly. Coccidiosis is of great economic importance because of the losses due to clinical disease (diarrhea), subclinical infections (poor weight gain in particular) and occasionally death (Chartier and Paraud, 2012). Our results displayed the presence of coccidiosis in goats from historical times. It is plausible that the presence of this infection in Patagonia is as old as the arrival of goats from Europe.
Comparing the two methods utilized for Eimeria spp. detection, three samples were negative with both methods (samples 3, 5, and 6), three samples (1, 8, and 9) were positive with both methods and four (2, 4, 7, and 10) were positive by either one of the techniques, showing that they can be complementary used. Discrepancies between microscopic and paleogenetic analyses found in our results are in accordance with previous studies where both methodologies were compared. Parasite remains and aDNA or other biomolecules, are not homogenously distributed and/or preserved within sediment samples (Côté et al., 2016). In the case of aDNA, PCR inhibition in environmental and ancient samples is also a common problem (Schrader et al., 2012). Our results combining with previous findings (e.g. Kévin et al., 2021; Wood et al., 2013), suggests that an approach combining microscopy and genetic techniques for the study of palaeoparasitological samples provides the most robust results under diverse contextual circumstances.
The authenticity of the sequences assigned to C. hircus and E. arloingi is supported with the number of positive replicates, here determined by two different PCRs (cytb and C. hircus specific) and by the absence of amplification in DNA extraction and PCR negative controls. In addition, DNA of E. arloingui was never amplified in our laboratory, thus minimizing the risk of contamination with modern DNA. The good preservation of aDNA obtained in this study is in accordance with the origin of the samples, which belong to recent times. Unlike other works with aDNA, the sequenced fragments in this study were longer and show less DNA damage. This may be due to the arid setting of the archeological site and the recent chronology of these samples, since the goat was first introduced in South America near the end of the XVI century (Bandieri, 2014).
Despite the value of coprolites in archeological contexts, they are not routinely collected or considered in many archeological studies (Shillito et al., 2020). This interdisciplinary study of coprolites broadens the archeological analysis at two levels. Firstly, as already discussed, by complementing osteological approaches, these results enhance the taxonomic resolution in the study of fossil assemblages, hence increasing the ecological evidence on the composition of past environments. In the present study, the paleoparasitological study of coprolites records the presence of goats in historical times, contributing to assess the interactions between native and introduced fauna in Patagonia. In addition, the paleoparasitological determinations provide a tool to assess the formation processes of archeological contexts by informing on the vertical migration of materials. The coprolites assigned to C. hircus appear in artificial levels 1, 3, and 4 of the stratigraphic sequence from CH1. While the presence in level one is consistent with the post-Hispanic introduction of the goat, level three is bracketed between 300 and 1500 cal years B.P., and level four between 10,200 and 11,300 cal years B.P. (details in Barberena, 2015; Llano et al., 2019). Accordingly, the paleoparasitological results provide firm evidence of the vertical redeposition within the sequence of the site. This is not unexpected considering that, despite the long temporal range spanning the Holocene, the site has a relatively shallow sequence (Barberena, 2015). Coupled with redundant human occupations and with the use of the shelter by goats, the displacement of these small and spherical coprolites is expected. Importantly, our results provide a direct proxy of the extent of the vertical displacement of materials, which may be highly usual in sheltered and repeatedly occupied spaces (Farrand, 2001; Hilton, 2003; Nielsen, 1991). Indeed, we have inferred the vertical migration of small lithic artifacts recovered from Pleistocene layers of the site (Rughini et al., 2021).
Conclusions
The coprolites studied in the present work have strong similarities to those of deer, goat and camelid previously described from archeological and paleontological sites from the southern cone of South America (e.g. Beltrame et al., 2017; Petrigh et al., 2021; Tietze et al., 2021). This suggests that it is necessary to apply different methodological tools to identify the correct host origin of the coprolites.
This work offers the first integrative approach complementing paleogenetic and light microscopic approaches to study Eimeria spp. as a tool to identify the zoological origin of coprolites. Light microscopic studies allowed the identification of oocysts attributed to Eimeria spp. However, the identification of the species was not possible. In addition, the targeted aDNA approach offered some advantages over microscopy yielding results to higher level of taxonomic identification. Sequencing allowed the identification of E. arloingi. Based on the host specificity of this coccidian, the identification of the zoological origin of coprolites was attributed to C. hircus. Thus, this study highlights that both techniques are complementary and promotes the use of the identification of Eimeria spp. in ancient samples in order to identify the zoological origin. Besides, the complementarity of both methodologies is more evident in those samples that tested positive with just one methodology. The use of both methodologies decreases the possibility of false-negative results. Moreover, the study of cytb genes confirmed the zoological origin of coprolites as belonging to domestic goat. Results showed the potential of aDNA profiling to identify artiodactyl coprolites. From the perspective of the study formation of the archeological record, the multidisciplinary study of coprolites offers an additional proxy that informs about taphonomic processes and archeological integrity.
Finally, results displayed the presence of coccidiosis in goats from historical times and suggests that possibly the presence of this infection in Patagonia is as old as the arrival of goats from Europe. Further application of these approaches to coprolites will continue to provide new insights.
Footnotes
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was funded by the projects PICT 2016-0062, PICT 2016-0316 (Agencia Nacional de Promoción Científica y Tecnológica) and PIP 2015-301 (Consejo Nacional de Investigaciones Científicas y Técnicas) from Argentina. The authorities on cultural heritage from Neuquén Province: Carlos Cides and Claudia Della Negra. Two anonymous reviewers for their time and improve the manuscript.