
Abstract
A Late Pleistocene/Holocene paleoenvironmental record was obtained from the Rouer peatland (5°54′E, 49°45′N; 270 m a.s.l.), located in the Gutland area of southern Luxembourg. A total of six sediment samples were AMS radiocarbon-dated to obtain an age-depth model. XRF analyses and analyses of geochemical proxies of organic matter (TOC, TN, δ13C, δ15N) were conducted to identify major paleoenvironmental changes in the record. Pollen analysis reveals insights into the vegetation history throughout the last 14,000 cal. yr BP. The record offers unique insights into the evolution of local organic sediment/peat accumulation, as well as into the environmental history of the Gutland region and beyond. The accumulation of organic sediment and peat started at about 13,800 cal. yr BP before present. Until about 6000 cal. yr BP, periods of apparently stable climatic conditions had been interrupted repeatedly by pronounced episodes with increased input of minerogenic matter into the peat matrix (12,700–11,800 cal. yr BP; 11,500–11,300 cal. yr BP; 11,100–10,800 cal. yr BP; 9300 cal. yr BP; 8200 cal. yr BP), indicated by sudden increases of Ti/coh values. After 6000 cal. yr BP, environmental conditions stabilized. Between 4200 and 2800 cal. yr BP, during the Bronze Age, changes in the pollen spectrum indicate an increasing clearance of woodlands. Since the Roman period, an ongoing intensification of grassland farming and agriculture is evidenced. Lowest tree species abundances are witnessed during the Middle Ages. The Modern Era is characterized by enhanced sediment input due to soil erosion. In short, this record complements the Late Pleistocene/Holocene climatic history of the Gutland area and demonstrates that fen peat deposits can be valuable high-resolution paleoclimate archives.
Introduction
Peatlands are important natural archives of past climatic change due to their characteristic feature to deposit a record of their own development in the form of peat (Barber, 2007; de Jong et al., 2010). Biological, geochemical and mineral proxy-indicators can be derived from peat cores to offer a multi-proxy approach for a better understanding of how local and regional ecosystems responded to past climatic and environmental changes (Diaconu et al., 2020; Strobel et al., 2019). In Luxembourg, only a few peatlands remain, as most of them have been degraded by drainage and reclamation for land use (Frankard et al., 2017). Some fen remnants are very small in size and include fragments of their former sedge-dominated vegetation, which have established after a long history of traditional agricultural practices.
Well-dated paleoenvironmental investigations in Luxembourg are rare and mainly cover the Late-Holocene. An overview of paleobotanical research efforts on the Holocene in Luxembourg is given by Gaudin (2019). Slotboom (1963) published the first results of palynological research, based on sediment cores from Gutland mardel depressions. Couteaux (1969) summarized the data of several palynological studies in Luxembourg, the majority of which were not 14C dated. Approximately 95% of the reported investigations covered time periods beginning in the Subatlantic Period (ca. 2500 cal. yr BP until today; e.g. “Stockem,” Gilot, 1968; “Clairefontaine, Kaarlsbierg,” Couteaux, 1969; “Beaufort Birkenbach,” Gilot, 1968). Others are presumed to date back to the Subboreal (ca. 5600–2500 cal. yr BP) (e.g. “Saint-Marie-Chevigny, Fange”; Gilot et al., 1965) or the Atlantic Period (ca. 9200–5600 cal. yr BP; e.g. “Muno Amerois” or “Breidfeld,” Couteaux, 1969).
From the mardel depressions “Rulles, Sur-le-Haut” and “Rodenbourg Bretzbaesch,” macro-remains were 14C dated and revealed ages that correlate with the Subatlantic period (Gilot, 1968). In a table, Couteaux (1969) listed further locations with older 14C-dated ages (e.g. “Berdorf”: 7720 14C yr BC ± 290), but without going into detail and without reference to literature. In conclusion, a lot of palynological work was conducted in Luxembourg throughout the 1960s, but dated cores and samples are sparse, or age-depth models are often not well-constrained. As such, older diagrams are of limited use for paleoenvironmental comparisons.
Guiot and Couteaux (1992) developed a quantitative climate reconstruction, based on a sediment core from an abandoned loop of the Sûre River. Unfortunately, the adopted chronology is based only upon pollen-stratigraphic comparisons with nearby 14C-dated sites previously reported by Couteaux (1970). The period supposedly covered is 2000–15,000 cal. yr BP.
Van Mourik and Slotboom (2018) compiled palynological information from Riezebos and Slotboom (1978), Poeteray et al. (1984), and their own investigations, based on 14C-dated alluvial, colluvial and peat deposits, including mardel depressions. Only the main features of the Holocene vegetation history are available from these investigations, particularly Roudbaach and Boeckenwiesen valley mires, which date back to the Early Holocene. However, age control for these investigations is still limited and does not exceed three datings per site.
Data with higher time-stratigraphic resolution is offered by the multiproxy studies of Dabkowski et al. (2015) and Granai et al. (2020), who attempted to reconstruct climate and vegetation changes in association with local land snail successions derived from Direndall tufa sequences. Their malacological analyses were undertaken in parallel with δ13C and δ18O isotope studies from the same profile.
Here, we present a new, well-dated peat/organic sediment record covering the last 15,000 cal. yr BP. We show high-resolution geochemical and pollen data to enable a more robust reconstruction of the paleoenvironmental history of the area and to provide insights into the evolution of the peatland since the last glacial termination. Our study evolved from a cooperative project between the Municipal Association for Nature Conservation (SICONA, Syndicat intercommunal pour la conservation de la nature, Luxembourg) and the universities of Cologne and Münster (Germany). To contribute to ongoing ecological research and to refine paleoecological data in Luxembourg, Rouer fen was selected as a target for sediment core retrieval because previous ecological research pointed to a large peat depth available here (Hollenbach et al., 2014). In Luxembourg, peat archives are very rare and paleoclimate research is often hampered by discontinuities in sediment accumulation. The aim of our research was to obtain a continuous paleoclimate record to contribute to the regional Late Pleistocene-Holocene climate and landscape history. The analysis of paleobotanical and geochemical patterns was also to explore how environmental changes affect in situ peat characteristics as a pre-requisite for the management and protection of Rouer peatland.
Study area
Geographical setting, regional climate, and vegetation
The peatland is located near the village of Niederpallen in the commune of Redange (5°54′E, 49°45′N) (Figure 1). It covers a small area of about 1 ha and is surrounded by extensive and intensive grassland and agricultural fields.

Regional setting of the study area in the southwest of Luxembourg.
The Rouer peatland is part of the hilly Gutland landscape, which is typical for the south of Luxembourg. The Gutland is characterized by weathered calcareous geological substrates. Trias formations like red sandstone, shell limestone, Keuper, and also Lias formations like Luxembourger sandstone, overlaid by loess, clay, and limestone, are typical for this area (Hollenbach et al., 2014; Schneider, 2019). The geological substrate at the study site is Keuper.
The climate in this region is characterized by precipitation that totals about 750–800 mm per year and a mean annual temperature of between 8.5 and 9°C. The elevation is 270 m a.s.l. Hollenbach et al. (2014) measured pH values of 5.8–6.3 in the uppermost peat layers.
The peatland’s vegetation is mainly composed of Carex acutiformis and Scirpus sylvaticus communities. Additionally, there are high abundances of Schoenoplectus tabernaemontani and the endangered species Triglochin palustre in the central area of the study site. The surrounding area is dominated by a high coverage of Phragmites australis and Juncus inflexus (Hollenbach et al., 2014; Schneider, 2019). The study site is extensively used by horses for grazing (in summer) and has been mowed in alternating sections every year in autumn/winter with a Caterpillar mower since 2016. Detailed species lists and vegetation descriptions can be found in Schneider (2019). The nearby streams are characterized as streams of colline type of the Gutland landscape (Administration du Cadastre et de la Topographie, 2018).
Nature conservation aspects
Because of the very small number and area of peatlands in Luxembourg and their poor habitat condition (European Environment Agency, 2019; MDDI, 2017), it is particularly important to preserve the few remaining peatlands and improve their condition. Within the framework of SICONA’s LIFE-Grassland project, the land adjacent to the previously investigated area was purchased. It is currently undergoing restoration in order to be able to better preserve the study area and to enable its extension. The sealing of drainage ditches to restore water saturation and the removal of topsoil (according to geoelectrical investigations) was carried out in 2019. A transfer of species-rich mowed material is planned for this summer, as well as the reintroduction of endangered species which will take place in autumn. This study area is particularly important for Luxembourg, not only as it is the only such well-preserved climate archive, but also due to its good base cation supply, in contrast to otherwise rather acidic peatlands in Luxembourg, and the occurrence of one of only five populations of the critically endangered species Triglochin palustre in Luxembourg (Schneider, 2019).
Material and methods
This investigation is based on a peat core, which was retrieved in February 2018 from the Rouer peatland (05°54′E/49°45′N) in SW Luxembourg. To select a suitable coring site, areas with thick peat accumulation and little disturbance by grazing in the peatland’s central section were chosen. Before coring, we applied Earth Resistance Tomography (ERT) to reveal detailed information about peat depth and the underlying valley shape structure. Three peat long cores were recovered by using percussion coring equipment. The peat was retrieved in PVC liners (100 cm, 50 mm in diameter), sealed and stored. The main core (Ro1A) was taken to a depth of 400 cm. The parallel core drilling was placed close to the main core drilling. There was one parallel core (Ro1C) taken from a depth of 350 cm.
Due to the high water content, the retrieved peat material was compacted by the drilling procedure. Therefore, some segments contain less sample material than 100 cm. Given that the 100 cm segments were taken out of the same borehole, contamination is possible. The contamination occurs because upper material is falling into the borehole (Pitkänen et al., 2011). To identify possible measurement errors due to contamination, the parallel core was taken with a 50 cm offset.
In the Palynology Laboratory of the Institute of Geography Education (University of Cologne), the cores were split into two core halves, photographed and described sedimentologically. One core half was subsampled at 5 cm intervals for geochemical analyses. To obtain qualitative element counts for major and trace elements, the other core half was analyzed at 2 mm resolution using an ITRAX X-ray fluorescence (XRF) core scanner (Cox Analytical Systems; Croudace et al., 2006) at GEOPOLAR (University of Bremen). XRF scanning was performed with a molybdenum (Mo) tube at 40 kV and 10 mA, using an exposure time of 5 s per measurement.
For stable isotope and element content analyses, subsamples of 5 cm were cut. The subsamples were freeze-dried and ground with a ball mill (Tungsten Carbide cups, MM400, RETSCH GmbH, Haan, Germany).
The total carbon (TC) and total nitrogen (TN) contents and δ13C and δ15N stable isotope composition were measured with an elemental analyzer (Eurovector EA 3000, HEKAtech, Wegberg, Germany) coupled to an isotope-ratio mass spectrometer (EA-IRMS; Nu Horizon, Nu Instruments, Wrexham, UK). Since the isotopic composition of total carbon would be highly sensitive to the presence of 13C-enriched CaCO3 in the samples, isotopic composition was measured within the peat-dominated, CaCO3-free core sections only. To this end, the samples were weighed into tin cartouches at a mass between 3.5 and 4.5 mg. A scale with a measurement uncertainty of d = 0.001 mg was used. While the content of C and N was measured in mass percent, we recalculated the TC/TN ratio as a ratio of molar mass.
To calibrate the δ13C and δ15N stable isotopic signatures of the samples, certified reference materials (IAEA 600) and certified commercial standards (wheat flour, organic rich sediment; IVA Analysentechnik, Meerbusch, Germany) were used. Results are reported based on the commonly used reference scales V-PDB for carbon and AIR for nitrogen, and expressed in the common δ-notation in units per mill (O’Leary, 1981):

Sample preparation for pollen analysis and macrofossil screening was carried out in the laboratory of the Institute of Geography Education (University of Cologne). The laboratory work followed the standard preparation protocol for pollen analysis, based on the recommendations of Faegri and Iversen (1989). For microfossil and macrofossil analysis, 100 samples of 1 cm³ volume were taken from the core segments at 4 cm intervals over the whole core.
The counting of pollen grains was performed using a transmitted light microscope with a 40× objective lens (ocular 10×/18). A total of 300 pollen grains per sample was counted. Pollen of trees, shrubs, herbs and grass pollen were included in the total. For identification, key illustrations of Beug (2004), Reille (1998), Moore et al. (1991), and a reference collection were used. The classification of fossil Betula pollen to species level was not possible due to its often moderate preservation in the samples. The low concentration of Betula pollen within the Younger Dryas sample did not allow the separation of Betula nana, but it is likely, according to Couteaux (1969), that Betula nana might have occurred in the Late Glacial steppe vegetation of the Gutland.
To offer chronological reference points for the scheme of vegetation changes within the late Pleistocene and the Holocene, Blytt-Sernander chronozones were added to the pollen diagrams. The timing of the chronozones is according to Bittmann et al. (2018) and Litt et al. (2001).
The plant and zoological macro-remains from the pollen samples were sieved in size fractions of 125–250 µm, 250 µm–2 mm, and >2 mm per sample to record the countable contents. The sample material was counted with a reflected light microscope (ocular 10×/21B). The identification of botanical remains was performed using Grosse-Brauckmann (1972, 1974, 1992) and Cappers and Bekker (2013).
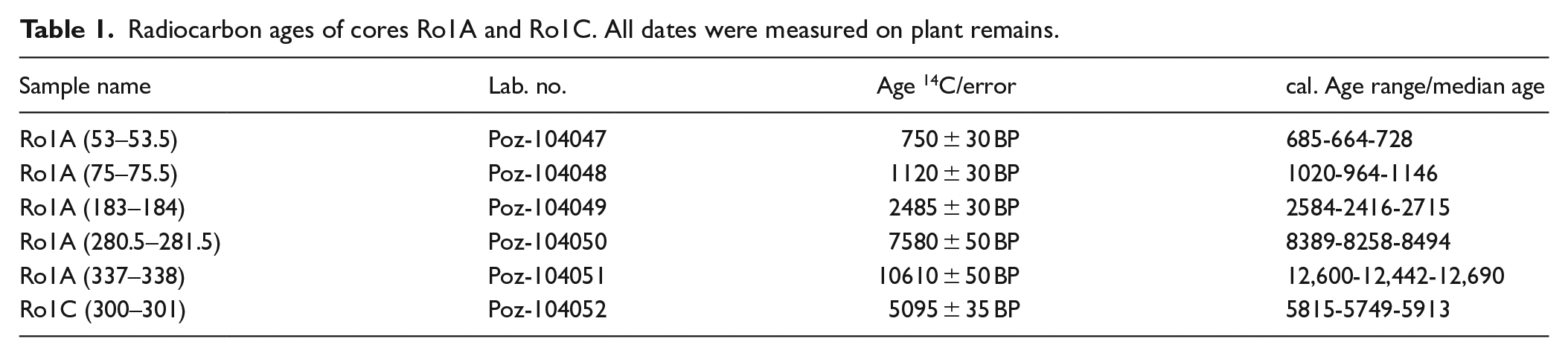
For age control, a total of six peat samples (five samples from core Ro1A, one sample from core Ro1C) were AMS (accelerator mass spectrometry) radiocarbon-dated at Poznan Radiocarbon Laboratory (Table 1). For radiocarbon dating, plant macrofossil residues were picked from the submitted sample material. No wood fragments were selected here. This is due to the fact that peat from fen woodlands can be contaminated with younger plant material by falling branches and tree trunks, as well as by the roots of the trees. All radiocarbon dates were calibrated using CALIB 7.0.4 and the IntCal13 data set for Northern Hemisphere calibration (Reimer et al., 2013). Due to the compaction of the peat material by the drilling procedure, core depth was adjusted by multiplying the compacted length by a correction factor, assuming linear compaction (core length/compacted length). The MCAgeDepth software package (Higuera et al., 2009) was used to develop the age-depth model based on five radiocarbon dates. The software was modified to calculate values at 1 mm resolution. All reported ages are calibrated ages unless stated otherwise.
Radiocarbon ages of cores Ro1A and Ro1C. All dates were measured on plant remains.
Northern Hemisphere temperature reconstruction data (Vinther et al., 2009), NGRIP oxygen isotope data (Svensson et al., 2008) and the sum of glacier advances (Wanner et al., 2011) were provided by the World Data Center for Paleoclimatology, NOAA/NCDC Paleoclimatology Program, Boulder, Colorado from their website (NOAA, 2019).
Results
Peat characteristics and chronology
The sedimentary deposits of the Ro1A sequence consist of four lithological units (Figure 2). Unit A is characterized by a Pleistocene solifluction soil composed of marl conglomerates. The accumulation of organic sediments starts in unit B, which is characterized by a silty-clayey gyttja with variable levels of embedded sand and organic matter. The initial deposition of peat is observed with the accumulation of a well-decomposed sedge peat in unit C1. Homogeneous alder forest peat with interspersed Cyperaceae rhizomes and wood fragments dominate unit C2. Wood fragments diminish rapidly at the transition to unit C3, which is characterized by sedge peat. Unit D, at the top of the sequence, is a strongly decomposed organic soil with a higher content of fine sand.
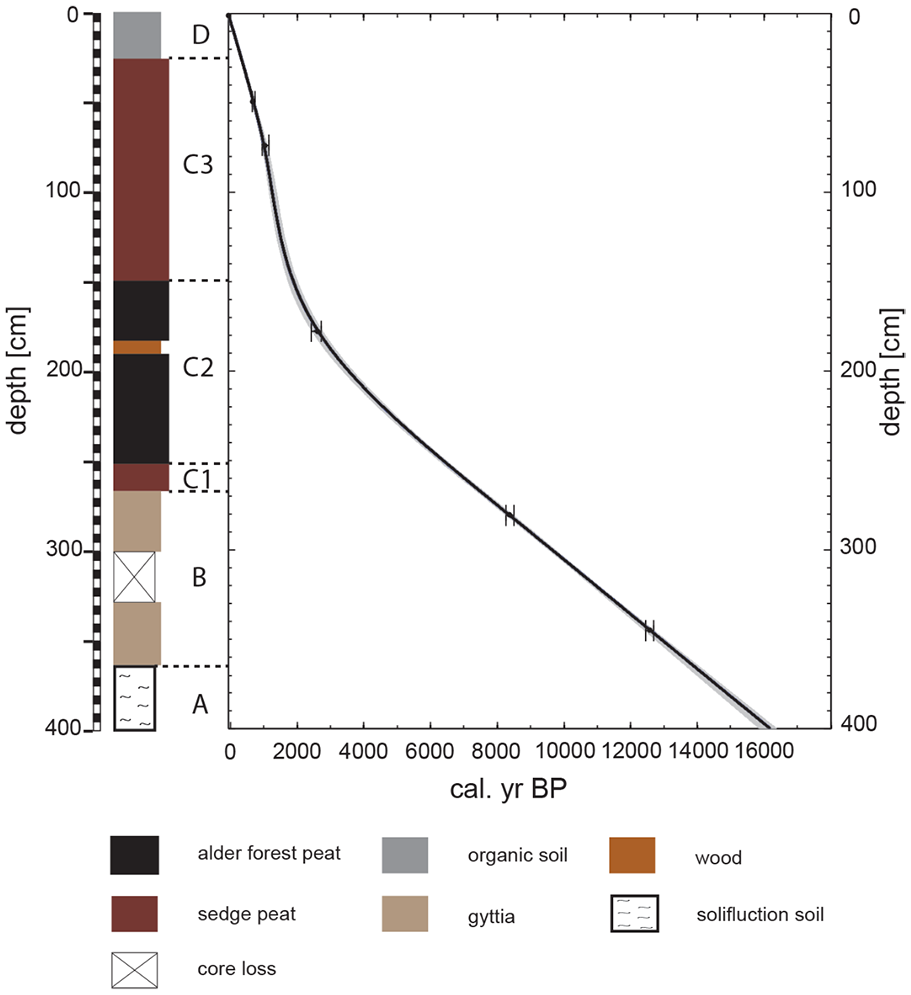
Stratigraphy, stratigraphic zones and chronology of core Ro1A of Rouer peatland.
The developed age-depth model (Figure 2) is based on five AMS radiocarbon dates and shows a constant peat accumulation rate within units B, C1, and C2 with a sample resolution of 20–65 years per centimeter. In units C3 and D, the accumulation rate is significantly higher (9–20 years per centimeter) with peak values at around 1100–1300 cal. yr BP.
Inorganic geochemistry
The results of the XRF-measured Ti/coh and Fe/Ti ratios are shown in Figure 3. As titanium (Ti) is considered to be immobile in peat, it can be used to reflect variations in allochthonous sediment input (Muller et al. 2008; Rothwell and Croudace 2015). To better illustrate effects arising from the sediment matrix, we used the ratio of Ti and the coherent (coh) scatter peaks of Mo (Ohlendorf et al., 2014). Iron (Fe) was normalized to Ti to better reflect the variations in autochthonous peatland dynamics.
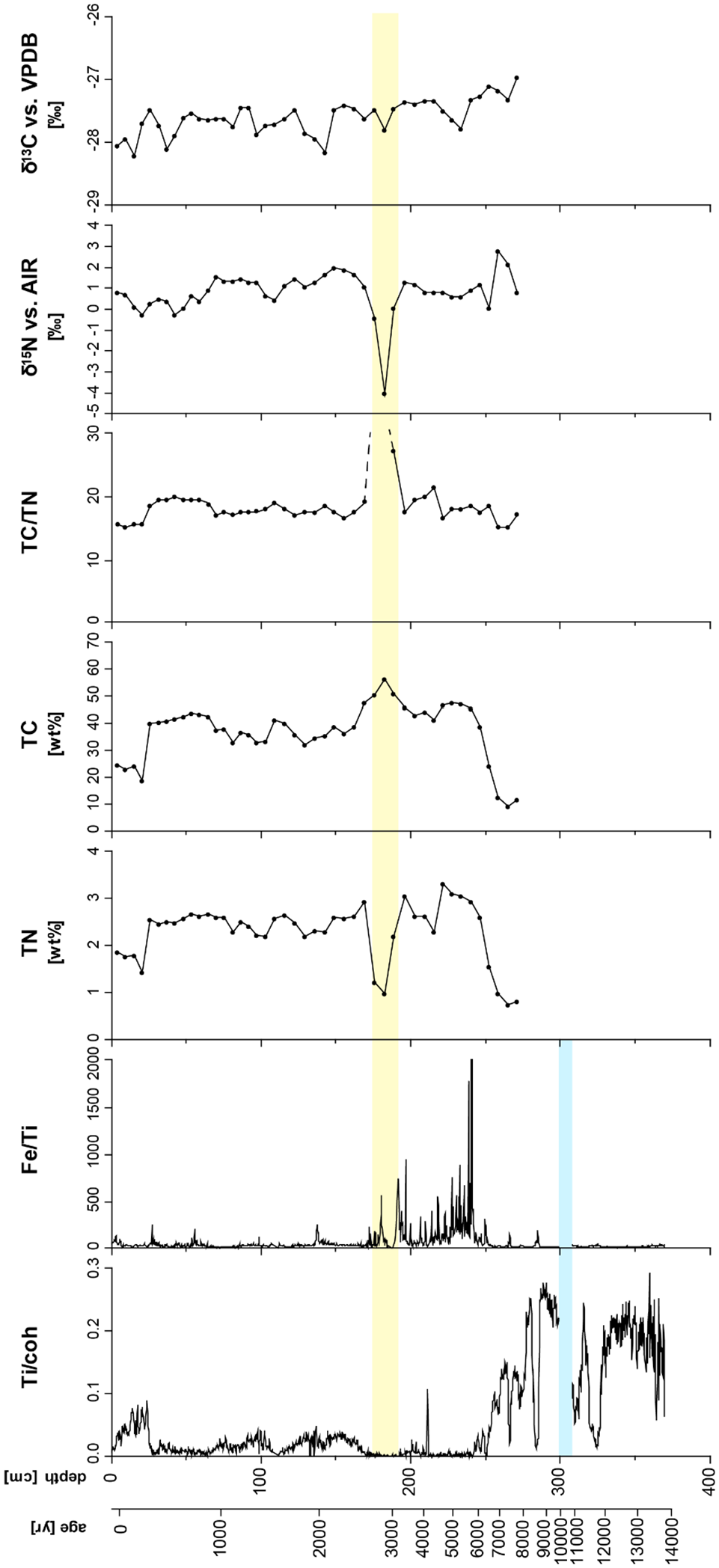
XRF-measured element ratios, total nitrogen/total carbon contents and stable isotope values for core Ro1A of Rouer peatland.
Further measured element variations were strongly influenced by allochthonous input of clastic sediment into the peat matrix, which is a typical feature for minerotrophic peatlands. Most notably in units A and B, before a homogenous peat body started to accumulate, the sediment was significantly characterized by mineral input.
Especially during the Late Pleistocene and Early to Middle Holocene sections of the core, the Ti/coh ratio was at its highest levels, which points to periods of high allochthonous sediment input. Here, low values, which were interpreted as periods of low allochthonous sediment input, were only observed at a depth of 340–330 cm (~11,800–11,500 cal. yr BP) and 285 cm (~8700–8500 cal. yr BP), with some shorter spells at 315–310 cm (~11,200–11,000 cal. yr BP), 265 cm (~7400 cal. yr BP) and 250 cm (~6400 cal. yr BP; Figure 3). After 6000 cal. yr BP, the Ti/coh ratio remained at consistently low levels until 2700 cal. yr BP. Another significant peak occurs only at 215 cm (~4200 cal. yr BP).
Above 170 cm (~2700 cal. yr BP), Ti/coh values showed a high variability at medium levels, consistently remaining higher than those during the previous time span (6000–2700 cal. yr BP). The highest values of the period after 2700 cal. yr BP were observed at around 170–110 cm (~2700–1800 cal. yr BP), 100 cm (~1200 cal. yr BP), 70 cm (~1000 cal. yr BP) and above 25 cm (~300–100 cal. yr BP), which indicates that allochthonous sediment input had increased again in the youngest layers of the peat deposit.
Within the Late Pleistocene and Early to Middle Holocene sections of the core, higher Fe/Ti values corresponded to periods of low allochthonous sediment input, reflected by low Ti/coh values (Figure 6). After 6000 cal. yr BP, Fe has been actively accumulated by the growing peat body. The periodic retention and release of Fe depended on the stability of the water table. Peaks in the Fe/Ti ratio indicate autochthonous enrichment of Fe in the capillary fringe under more oxic conditions.
The mid-Holocene period between 6000 and 2700 cal. yr BP showed the highest variability and the highest levels of Fe/Ti in the record. After 2700 cal. yr BP, high values of Ti/coh and low values of Fe/Ti reflected higher contents of introduced minerogenic matter. Subsequently, higher levels of Fe/Ti ratios were only observed at around 1400 cal. yr BP and at around 900–800 cal. yr BP.
Organic geochemistry
The C/N ratios determined in core Ro1A ranged from 14.93 to 67.1. The maximum range and the average were, however, influenced by measurement of a wood fragment, which was incorporated in the peat at a depth of 145–160 cm (~2700–3100 cal. yr BP; Figure 3). Excluding this measurement, the maximum C/N ratio of Ro1A was 21.17 at a depth of 180 cm (~4300 cal. yr BP) and a mean value of 17.67. In the upper 20 cm (last 200 years) of the Ro1A core the C/N ratios were notably lower than at greater depth. At a depth of 25–60 cm (~200–800 cal. yr BP) the values of the C/N ratio range from 18.39 and 19.66, and are therefore higher than in the upper part of the Ro1A core.
The δ13C and δ15N signatures showed a trend of increasing values with depth (Figure 3). The δ13C values of the Ro1A core varied in a range of −28.21‰ and −26.96‰. The maximum value was observed at a depth of 225 cm (~7800 cal. yr BP), the deepest sample measured. The minimum value was observed close to the surface at a depth of 15 cm. The δ15N signatures showed a similar trend as the δ13C signatures, increasing with depth, though less pronounced. The range of the δ15N values of Ro1A varied between −4‰ and +2.82‰. Noticeably, due to the presence of wood, the minimum value is at a depth of 150 cm (~6500 cal. yr BP) where the C/N ratio is the highest.
Pollen analysis and selected peat macro-remains
The pollen stratigraphy within the Rouer sequence stretches from the late Pleistocene to the middle of the 20th century. Figures 4 and 5 show the results of the pollen percentages (see also Supplemental Figures S1 and S2). All samples of the peat/organic sediment record yielded sufficient pollen for counting, although the preservation of pollen grains was partially poor, especially within the mid-Holocene section (alder forest peat). Overall, the pollen spectrum reflects a tree-dominated vegetation structure with high percentages of Alnus, Pinus, Betula, Corylus, and Quercus. Open-land grasses and herbs reached higher percentages only during the late Pleistocene and within the past 2000 years (Figure 4).
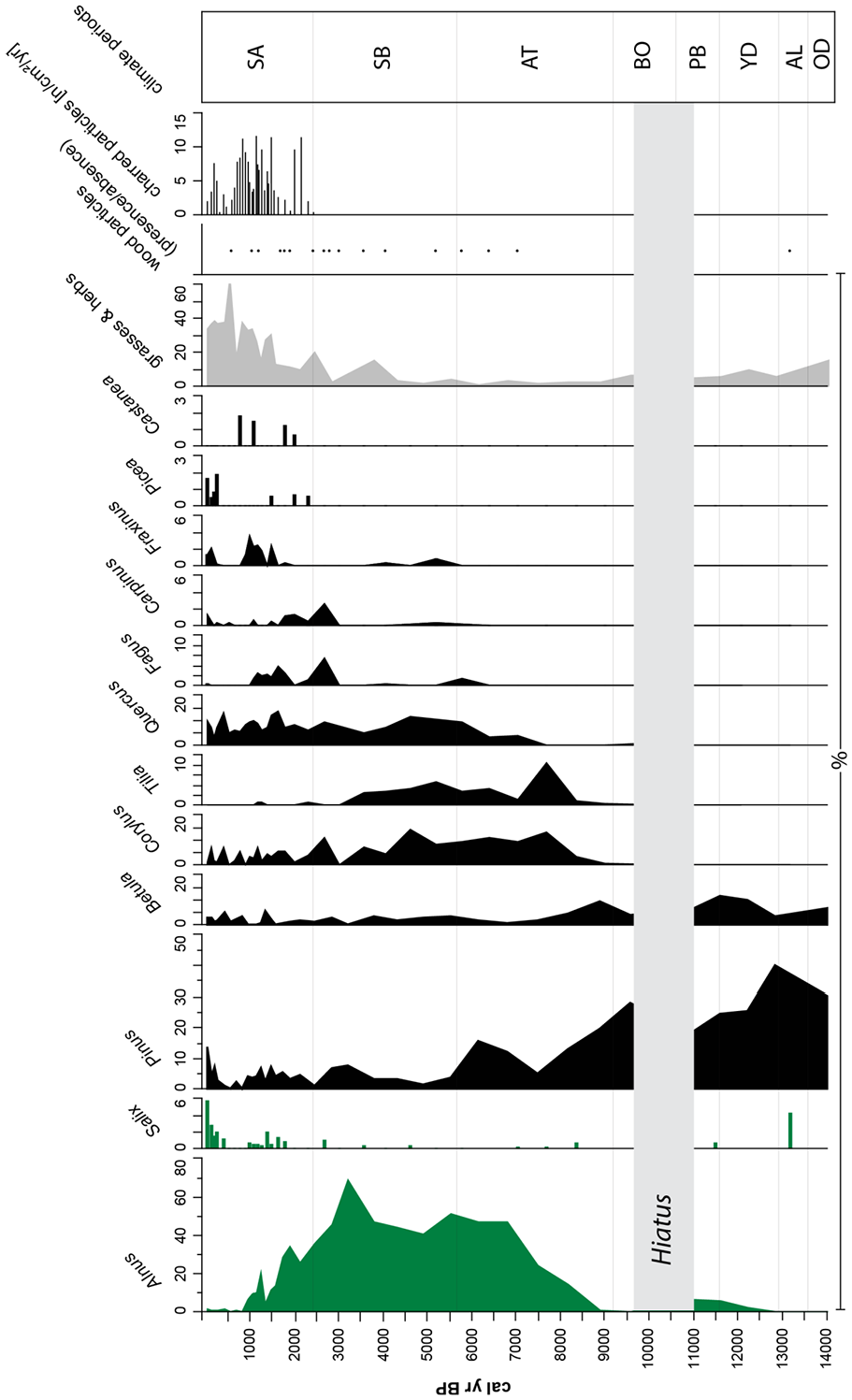
Tree species pollen percentage diagram of core Ro1A of Rouer peatland.

Pollen percentage diagram for the past 3000 years showing wetland types (green, excluded from the pollen sum), selected tree species (black) and selected open-land types (gray).
In total, 53 different pollen types were identified. Among the herbs, only Asteraceae and Brassicaceae reached percentages >20%, and only within the past 2000 years. None of the herbs appeared continuously throughout the record. Pollen of Cyperaceae, Lemnaceae, Alnus, Salix, Typha, and Caltha represent local wetland vegetation and were excluded from the pollen total. Wood particles, charred particles, Juncaceae diaspores, and Carex diaspores were counted in the sieved fractions (125–250 µm and 250 µm–2 mm) (Supplemental Figure S2).
Before the onset of the Holocene, the pollen spectrum is characterized by the dominance of Pinus pollen, which, as well as Betula, reached the highest percentages in the record. The high abundance of Salix suggests moderate climate conditions with pioneer trees during the Late Alleröd. Pollen of Poaceae, Asteraceae, Cichoriaceae, Brassicaceae, Apiaceae, and Caryophyllaceae point to a mainly open landscape.
The Preboreal, Boreal and the beginning of the Atlantic still showed high abundances of Pinus, but its dominance steadily decreased until about 7500 cal. yr BP. Betula also decreases in abundance toward the end of the Atlantic. Very low values of Carpinus and Tilia (lower than 1%) occurred during the Boreal. Decreasing open landscape-related pollen types such as Poaceae, Asteraceae, Rubiaceae, Brassicaceae (disappearing in the middle of the zone), Apiaceae and Artemisia suggest a decrease of open landscape vegetation in favor of a boreal forest. Pine trees were declining toward the end of the Boreal, while birch fluctuated. Other trees (Carpinus, Tilia, and Quercus) remained at low values. Wet-tolerant plants like Typha, Lemnaceae, and Caltha showed slightly increasing values. With the beginning of the Atlantic period, Alnus pollen significantly increased in abundance. Furthermore, Tilia showed a remarkable peak in this period, whereas Corylus, Pinus, Betula, and Salix tended to decrease. The first appearance of Fagus pollen (< ca. 2%) and an increase of Poaceae can be seen toward the end of the Atlantic period. Cyperaceae fluctuated at lower, but steady levels. Furthermore, wet-tolerant plants such as Lemnaceae, Caltha, and Typha remained abundant. Herbs generally remained at lower values; only Apiaceae showed increasing values toward the Atlantic/Subboreal transition.
The beginning of the Subboreal is marked by a high dominance of Alnus pollen. In this record, only low contents of Fagus pollen prevailed in this period. Light-loving trees such as Corylus benefited from this by showing relatively high abundances. Betula, Tilia, and Quercus showed relatively constant values; low and fluctuating values with single peaks occurred for Carpinus and Fraxinus. Pinus had decreased during the mid-Subboreal, but had increased again to medium abundances at the transition to the Subatlantic. Cyperaceae and Poaceae still fluctuated at lower values, but showed a peak around 4500 cal. yr BP. Open-land vegetation taxa (Asteraceae, Apiaceae) and those related to arable lands (Chenopodiaceae, Artemisia) and Plantaginaceae showed single high values within this zone.
The end of the Subboreal and the beginning of the Subatlantic stood out with higher abundances of Fagus and Quercus, and a steady decrease of Alnus pollen (Figure 5). Furthermore, Picea, Salix, Carpinus, Fraxinus, and Castanea increased in abundance. Pinus showed fluctuating, mainly low percentages, and have only risen to higher values toward the top of the record within the past 400 years. Juniperus and Larix showed only one occurrence with a percentage lower than 1%. The dominating types during the Subatlantic period were Cyperaceae, Poaceae, Asteraceae, Chenopodiaceae, Brassicaceae (high values up to nearly 10%), Linum and Plantaginaceae, which points to the increasing establishment of an open, deforested landscape during that period.
Discussion
Geochemical indicators for peat formation and peat decomposition
The Ti/coh ratio clearly identified periods in which the peat was affected by the input of allochthonous minerogenic matter, originating from the catchment area. The input was likely triggered by rainfall events, especially under unstable climatic conditions, and depends on the density and characteristics of the vegetation cover. Hence, a stratigraphic analysis of the peat/organic sediment column allows inferences about the intensity of erosional processes, and therefore, can be linked to paleoclimatic changes. The overall effect depends on the productivity of the peatland ecosystem in relation to climatically and/or human-induced rates of weathering and erosion in the catchment area (Schittek et al., 2012).
Peaks in the Fe/Ti ratio are interpreted as indicators of changes in redox conditions due to water table fluctuations within the peatland (Schittek et al., 2016). High Fe/Ti ratios reveal an upward movement of Fe2+ from the anoxic peat to the upper aerated layers, followed by precipitation as Fe3+ oxide (Damman et al., 1992). Higher enrichment of Fe relative to Ti in the zone of water table fluctuations under oxic conditions is thus indicative of climatic conditions including episodic droughts, which affect the water saturation of the peatland (Shotyk, 1988).
The C/N ratio is a commonly used indicator for the decomposition of organic matter (Hornibrook et al., 2000; Krüger et al., 2015; Kuhry and Vitt, 1996). Because of higher loss of C relative to N during microbial decomposition, the C/N ratio becomes smaller in strongly decomposed peat.
The C/N ratios of the peat profile of the minerotrophic peatland “Rouer” vary in a range, which is considered typical for minerotrophic peatlands (Andersson et al., 2012), except for high ratios due to the wood fragment in Ro1A at 2700–3100 cal. yr BP (Figure 3). At this depth, exceptionally low δ15N values also occurred. Besides this, the C/N ratios showed a relatively uniform profile with depth, suggesting that there had been little variation in conditions for decomposition and litter inputs were of comparable quality (Jones et al., 2010). In the uppermost part of the core, the C/N values were notably smaller than at greater depth, indicating stronger decomposition due to past drainage or a higher input of minerogenic matter and nutrients, likely altering predominant vegetation (Gałka et al., 2019; Jones et al., 2010). Lower C/N ratios were also observed at the transition from the Middle Holocene to the Early Holocene, when minerogenic input had also increased. In the remaining part of the profile, C/N ratios did not consistently respond to changes in either Ti/coh or Fe/Ti ratios.
The δ13C signature varied in a narrow range around −27‰ and corresponded to δ13C values typically reported for peatland vegetation (Broder et al., 2012; O’Leary, 1981). More specifically, the measured δ13C signatures fell into the range of minerotrophic peatlands, which are generally somewhat lower than δ13C values of ombrotrophic peatlands (Hornibrook et al., 2000; Jones et al., 2010), an effect typically ascribed to a recycling of carbon from 13C-depleted methane (Jones et al., 2010; Nichols et al., 2009; Raghoebarsing et al., 2005), and/or differences in vegetation and decomposition (Biester et al., 2014; Gałka et al., 2019), but to a lesser extent to decomposition or diagenesis (Jones et al., 2010).
In general, an increasing trend with depth was observed for the δ13C profiles of the analyzed peat cores (Figure 3). The δ13C record may on the one hand be an indicator for changes in climate and vegetation (Gałka et al., 2019; Hornibrook et al., 2000; Jedrysek and Skrzypek, 2005). Drier and warmer conditions would lead to more negative δ13C values in bogs (Broder et al., 2012; Jedrysek and Skrzypek, 2005), where the C uptake of Sphagnum mosses depends on the thickness of water film on the mosses. The effect is therefore not easily applicable to wetlands without a dominance of Sphagnum mosses (Jones et al., 2010). On the other hand, the increase of δ13C values with depth here may be related to changes in decomposition. In most ombrotrophic peatlands, δ13C signatures are expected to show a uniform depth profile, or even slightly decreasing values, caused by limited decomposition and the accumulation of slowly decomposing substances depleted in 13C (e.g. lignin) (Alewell et al., 2011; Meyers, 1997). However, such residual enrichment of 13C-depleted, refractory compounds may not strictly apply to fens. In the presence of vascular plants with aerenchyma, oxygen is transported in the water-saturated peat and could enhance decomposition (Mainiero and Kazda, 2005). For example, this was described for a restored cutover peatland (Agethen et al., 2018), and also in an extreme case, of peatlands in Patagonia with a particularly high root density of cushion-forming plants with aerenchyma (Münchberger et al., 2019). Such enhanced decomposition, likely in the presence of oxygen, could then also lead to a breakdown of refractory compounds and residual enrichment of 13C, as experimentally validated for example, by Lehmann et al. (2002) for oxic versus anoxic decomposition, and as typically observed in aerated soils (Ågren et al., 1996; Nadelhoffer and Fry, 1988). Variations in δ13C may thus predominantly indicate differences in oxygen availability for decomposition. Indeed, we see congruent patterns of increasing values of δ13C around phases of high Fe/Ti ratios, indicating drier conditions (see e.g. 25 cm/330 cal. yr BP, 60 cm/840 cal. yr BP, 100–120 cm/1230–1400 cal. yr BP). It has to be noted, though, that patterns are in part shifted (see e.g. 130–150 cm/1530–1850 cal. yr BP) and the signals of anoxic versus oxic decomposition (Lehmann et al., 2002) are apparently obscured by other factors such as vegetation (Galka et al., 2019; Jones et al., 2010).
Signatures of δ15N in fens are not straightforward to interpret because N can enter fens through several sources in different organic and inorganic forms. Moreover, the processes in the N cycle leading to fractionation are highly complex (Jones et al., 2010). The δ15N values in bogs typically scatter around zero ‰ (Broder et al., 2012; Jones et al., 2010). This is also observed in the minerotrophic peat profile of the Rouer peatland. Yet, the N isotope signatures vary in a relatively wide range of −4‰ to 3.22‰ (Figure 2). Similar ranges were observed in bogs by Broder et al. (2012) and in forest soils of the temperate zone by Novák et al. (2003), while limnic sediments may be more enriched in 15N (Lehmann et al., 2002). The variability of the values should at least partly be related to the high variability of the δ15N values of different plant species (Krüger et al., 2015), but Liu et al. (2020) also reported that the initial 15N may be diagenetically altered, limiting its interpretation. There was a slight trend of increasing values with depth observable in the Rouer peat profile. Higher δ15N values may indicate stronger decomposed organic matter, as repeated cycling and mineralization of N leads to a preferential loss of 14N over 15N (Dijkstra et al., 2008; Nadelhoffer and Fry, 1988). Such enrichment of 15N would therefore be expected for decomposition under oxic conditions, while under anoxic conditions, a 15N depletion would be expected (Lehmann et al., 2002). Krüger et al. (2015) suggest that like δ13C values, δ15N values increase with depth in degraded peatlands due to higher microbial activity under aerobic conditions; conditions that might also apply to fens with intense decomposition in the presence of vascular vegetation with aerenchyma. This suggests that the organic matter is increasingly decomposed with depth, presumably with the availability of oxygen, confirming our interpretation of observed patterns in 13C (Krüger et al., 2015; Nadelhoffer and Fry, 1988). Smaller variations in the profile were indeed not related to other indicators of drought or wetness, thus the exact interpretation of distinct fluctuations may be limited due to diagenetic processes (Liu et al., 2020).
Late Pleistocene and Holocene environmental changes
Figure 6 summarizes the Late Pleistocene/Holocene environmental history of the Rouer fen in comparison with the paleoclimatic evidence from Greenland ice cores (Svensson et al., 2008; Vinther et al., 2009) and the sum of Holocene glacier advances (Wanner et al., 2011). The accumulation of organic-rich sediment, which is not solifluidally reworked, can be identified in the deposits of the Rouer fen at about 13,800 cal. yr BP during the Bølling interstadial. The Late Pleistocene was a period of dramatic climatic changes, which is also reflected in the oldest sediment layers of Rouer fen profile. During the Bølling period, solifluction and runoff within the Pall and Näerdenerbaach valley floors diminished, so that organically-enriched floodplain sediments started to accumulate at Rouer fen, interspersed by less organic-rich debris, as reflected by the oscillating Ti/coh values. During the Bølling and Alleröd interstadials (between 13,800 and 12,500 cal. yr BP), several short phases of significantly low Ti/coh values were identified, which points to a denser vegetation cover. According to Guiot and Couteaux (1992), the climate became increasingly warmer after 13,000 cal. yr BP and moisture was nearly as high as the present day. Higher abundances of grasses at this time indicate a landscape with an open character (Slotboom and van Mourik, 2015). At Rouer fen, pollen spectra show high scores of Pinus, Betula, and herbal/grass pollen. Apart from the cold-tolerant tree species (Pinus, Betula, Salix), deciduous arboreal pollen taxa are still very rare due to the long lag time for their arrival from their glacial refugia (Litt and Stebich, 1999). Pinus is the dominant pollen species, which shows that Pinus trees successfully colonized the open bare ground, where soils had yet to develop.
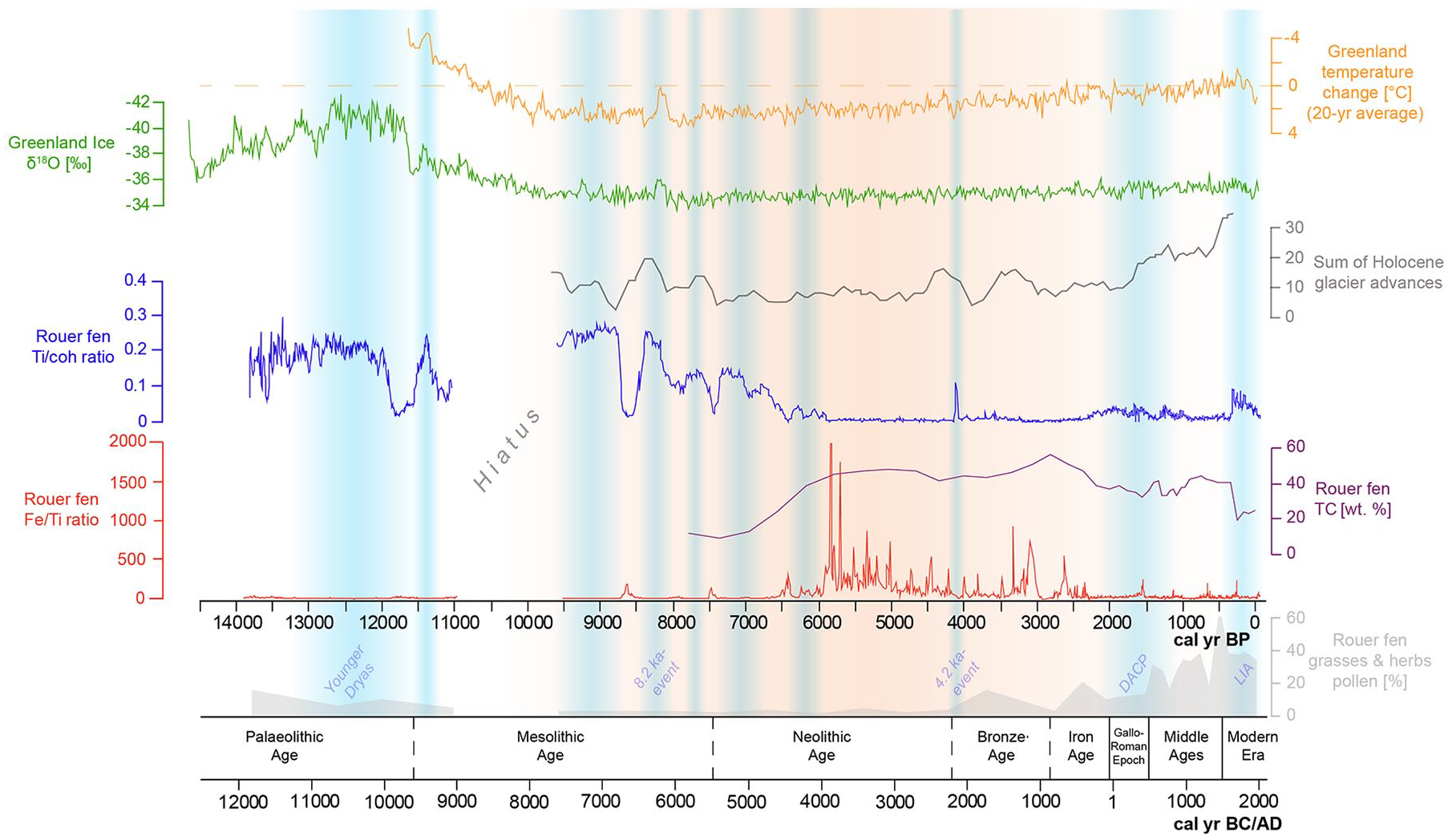
The late Pleistocene/Holocene Ti/coh, Fi/Ti, TC/TN ratio, and grasses/herbs pollen sequences of Rouer peatland compared with NGRIP oxygen isotope data (Svensson et al., 2008), Northern Hemisphere temperature reconstruction data (Vinther et al., 2009) and the sum of glacier advances (Wanner et al., 2011) in comparison to the archaeological chronology (Paulke, 2019).
At around 12,700 cal. yr BP, this period of warmer climate ended due to a sudden southward shift of the oceanic polar front, returning to a position similar to that of the Last Glacial Maximum (Anderson et al., 2007). During this 1200 year-long cold period, the Younger Dryas, the climate was markedly drier due to a renewed strengthening of the continental high-pressure cells (Sirocko et al., 2016). The Younger Dryas cold event is clearly depicted in several paleoenvironmental records of the Eifel/Ardennes area (e.g. Brauer et al., 2008; Guiot and Couteaux, 1992; Sirocko, 2010; Sirocko et al., 2016). The mean July temperature depression of 7–8°C had a strong impact on the environment and led to a return of vegetation characteristic for subarctic conditions (Atkinson et al., 1987; Brauer et al., 2008; Lowe et al., 1994). Due to the harsher conditions, trees should have disappeared from the higher lying areas with harsher climate conditions and might have survived only in more protected valley floors. The pollen spectra of Rouer fen correspondingly show a decrease in Pinus, which was also evidenced by Guiot and Couteaux (1992) in the east of Luxembourg. The timing of the fluctuations in the Ti/coh values match the range and the oscillations of the Younger Dryas, as recorded by the Greenland ice δ18O isotope values. A drier and colder climate presumably led to an increased allochthonous sediment input into the fen area, which might have been a result of an overall reduced vegetation cover within the catchment area of Pall and Näerdenerbaach creeks. The end of the Younger Dryas is marked by a sudden temperature increase of about 8°C within only 5 years, at around 11,500 cal. yr BP, according to temperature reconstructions based on isotope data (Alley et al., 1993; Sirocko, 2010). The Rouer fen records this event apparently earlier at around 11,800 cal. yr BP, which might be due to insufficient age control in this section of the core. However, the variation pattern of the Ti/coh compares well to the Greenland isotope values.
The onset of the Holocene is marked by a rapid shift of temperature, right at the end of the Younger Dryas. According to Anderson et al. (2007), the mean annual temperature increased by 7°C within, at most, 50 years. This quick shift was attributed to changes in the North Atlantic drift system and an increase in sea surface temperatures, when the Gulf stream warmed abruptly (Alley et al., 1993; Knorr and Lohmann, 2007). The Rouer fen record documents this shift as a sudden decrease of Ti/coh values between 11,900 and 11,500 cal. yr BP. This period of low Ti/coh values correlates well with a contemporaneous increase in δ18O values of Greenland ice cores and can be assigned to the Friesland phase (Bos et al., 2005), which marks the beginning of the Holocene warming.
The Early Holocene experienced a series of important environmental changes. Steppe species declined and land became available for the expansion of an open woodland composed of Pinus, Betula and a little Corylus (Guiot and Couteaux, 1992). Following our Rouer fen pollen record, Alnus also became more abundant and might have colonized wet depressions and river valleys. Compared to the Late Pleistocene, especially during the warm Friesland phase, there was obviously a reduced fluvial discharge through rivers and brooks. River channels in the Rhine-Meuse area started to fill with organic sediment and peat, which significantly slowed down water run-off and resulted in a process of aggradation (Crombé, 2018).
After the warm beginning of the Holocene, a cold episode occurred from ca. 11,500–11,300 cal. yr BP, which led to a sudden increase of Ti/coh values again. This episode correlates with the Preboreal oscillation, which is characterized by a mean temperature decline of about 1–2°C for the subsequent 100–150 years (Björck et al., 1997; van der Plicht et al., 2004). Magny et al. (2007) presume wetter conditions in west-central Europe in response to the Preboreal oscillation, as inferred from higher lake water levels reconstructed for several European lakes between 11,300–11,150 cal. yr BP.
After this first prolonged Holocene cooling episode, reduced Ti/coh values indicate a return to a state with less input of allochthonous sediment at Rouer fen. This coincides with a warm episode, following the cold Preboreal oscillation (Fiłoc et al., 2016; Magny et al., 2007). However, this short warming was followed by the next abrupt climate cooling starting from ca. 11,100 cal. yr BP and lasting until 10,800 cal. yr BP (Bohncke and Hoek, 2007). At this point, a hiatus appears in the Rouer fen record. Crombé (2018) reported that a sedimentary hiatus in peat growth was observed in several, mostly smaller and medium-sized valleys of Rhine-Meuse area during that period. The main cause for this hiatus is likely climate oscillations during the Preboreal and a resulting instability of the environment with high amplitudes in fluvial discharge (Fiłoc et al., 2016). Paleoenvironmental information from the Rouer fen record is evident again at ca. 9600 cal. yr BP, when the pollen record shows noticeably high percentages of Pinus.
The abrupt isotope shifts in Greenland ice cores at around 9300 cal. yr BP and 8200 cal. yr BP represent the most sudden climatic shifts during the Holocene (Figure 6; Alley et al., 1993; Rasmussen et al., 2007). These are likely driven by catastrophic meltwater releases into the North Atlantic Ocean, which disrupted the thermohaline circulation and its attendant northward transport of warm water and air masses (Törnqvist et al., 2004; Yu et al., 2010). Sudden increases in Ti/coh values indicate a strong impact on allochthonous sedimentation in the Rouer record, associated with these sudden cooling events. The climate deteriorations at 9300 and 8200 cal. yr BP are part of a repeating pattern of longer-term anomalies during the Holocene, seemingly related to fluctuation in solar output. This pattern manifested as “Bond events” later on again at 5900, 4200, 2800, 1400 cal. yr BP, and most recently, during the Little Ice Age (Rohling and Pälike, 2005).
Lake sediment studies in eastern France also point to cooler temperatures and higher precipitation during the Bond events, due to a strengthened Atlantic westerly jet stream that resulted in more storm tracks across mid-latitude Europe (Anderson et al., 2007). The 8200 cal. yr BP cold anomaly affected all of central Europe (von Grafenstein et al., 1998) and lasted for about 120 years, according to the precisely-dated evidence from Lake Holzmaar in the nearby Eifel Mountains (Prasad and Baier, 2014). The pollen evidence from the Eifel maar lakes further shows that the Early Holocene forest structure in that area, mainly composed of Pinus and Corylus, did not recover after the 8200 cal. yr BP event. Instead, the development of a mixed oak forest started (Sirocko et al., 2016). The Rouer fen record shows a continuous increase of Alnus and Tilia after this event, as these species might have dominated the wetter valley grounds in the local catchment areas and have spread due to the warming after the 8200 cal. yr BP event.
However, environmental conditions remained relatively unstable until about 6000 cal. yr BP, as the Rouer fen record remained enriched with allochthonous titanium. Yet between 7000 and 6000 cal. yr BP, sediment input decreases steadily and the TOC content rises to levels above 30%. Dabkowski et al. (2015) suggest relative humidity stability and maximal forest development for the early Middle Holocene, inferred from mollusc fauna at Direndall tufa. The reconstruction of drier conditions between around 7100 and 6500 cal. yr BP by Granai et al. (2020), based on further malacological studies from the same site, is consistent with a general decrease of Ti/coh values, generally less negative values of δ13C, and lower values of C/N ratios. Guiot and Couteaux (1992), based on pollen data from the Echternach area, suggest warmer and wetter summers during the Atlantic period until 6000 cal. yr BP and a climatic change toward drier summers after 6000 cal. yr BP. At Rouer fen, by 6000 cal. yr BP, the alder forest had apparently fully developed and alder dominates the pollen spectrum for the next 3000 years. Pinus decreases, and deciduous trees, including Fagus, Carpinus, and Fraxinus, were fully established. From 6000 cal. yr BP onwards, temperatures in all seasons in central Europe had evened out to values within ± 1 K from modern values (Davis et al., 2003). The prevailing climate after 6000 cal. yr BP is characterized by short-term, periodic dry events, as reflected by the frequency of shifts in the Fe/Ti ratio, still low C/N ratios, but values of δ13C turning more negative. Sedimentation at Rouer fen had shifted from clastic-detrital deposition to an autochthonous sedimentation in the form of peat.
Climate remains relatively stable until about 4200 cal. yr BP. Afterwards, the frequency of Fe/Ti shifts reduces. Due to the evidence of tree line retreat in the Alps, a transition to a climate cooling with an increase in precipitation can be determined for the period after about 5000 cal. yr BP (Wick and Tinner, 1997). This is related to an intensification of the Gulf Stream, which at least from about 4000 cal. yr BP, also had an impact on increased cyclonic activity and a more meridional circulation pattern across Europe (Frenzel, 1998).
The Rouer fen record shows a marked peak of Ti/coh at 4200 cal. yr BP, which evidences a prominent climatic anomaly (e.g. Weiss, 2016). Dörfler et al. (1998) and Löhr and Neyses-Eiden (2001) mention an extreme weather event at the same time, which had led to the embedding of numerous tree trunks in abandoned meanders of Mosel and Kyll rivers in the nearby Eifel Mountains. A disastrous flood event was also evidenced in the sediments of Ulmener Maar Lake (Sirocko, 2010).
Between 4200 and 2800 cal. yr BP, during the Bronze Age, no further climate anomalies occurred. Conditions seem to have been rather stable, which is also reflected by more stable values of C/N and δ13C. Increasing percentages of grasses and herbs in the Rouer pollen spectrum indicate that Bronze Age people had extended the clearance of woodlands. The decrease of Tilia during that period might be related to the fact that the sites where Tilia grew often turned out to be suitable as farmland, and therefore, forest was preferentially cleared at these sites (Behre, 2007). In the Rouer pollen record, Tilia continuously decreases from 5500 cal. yr BP on, and especially between 3500 and 2900 cal. yr BP, abundance reduces to percentages below 1%. Alnus pollen levels, starting at 3800 cal. yr BP, rise to the highest in the record, reaching nearly 70% at around 3100 cal. yr BP.
Several paleoclimate records suggest that temperatures cooled further at the Subboreal-Subatlantic transition at around 2800 cal. yr BP (van Geel et al., 1996; Wanner et al., 2008; Zolitschka et al., 2003). Pare et al. (2010) remarked that the change toward a more cool-humid climate at 2800–2700 cal. yr BP led to a subsistence crisis and provoked migration in several regions during the early Hallstatt culture. The Rouer pollen record shows a contemporaneous decrease of grasses and herbs, and a strong increase of Fagus, Carpinus, Quercus, and Corylus, which indicates that open land was abandoned and forest recovered in the surroundings of Rouer fen. By the 2nd century BC, during the late Iron Age, settlement conditions in central Europe had stabilized again. City-like, fortified hilltop settlements of large dimensions were created all over the area of the Celtic culture.
During that period of cultural bloom, the transformation of the alder forest into an open sedge reed can be demonstrated. The first signs of clearing can be inferred from the appearance of larger numbers of charred particles at around 2200–2100 cal. yr BP. The appearance of Juncaceae seeds and Carex seeds indicates a more open vegetation. The presence of oospores from Characeae point to the existence of sun-exposed pools within the fen. Dabkowski et al. (2015) link the environmental change during the episode of Roman occupation to anthropogenic forest clearing.
The Roman period is known for a significant impact on vegetation through deforestation and intensification of agriculture. In the pollen diagram (Figure 5), clearing of the forest is shown by a reduction in tree pollen percentage of Alnus in particular, and an increase of Cyperaceae and Poaceae. Fagus also declines sharply, but, like Quercus, recovers by around 1800 cal. yr BP, when Cyperaceae and Poaceae again decrease in abundance.
The cold spell of the Migration Period, after the collapse of the Roman Empire, led to a decrease in population density, especially in rural areas (Kausch, 2006). Tree pollen, mainly Fagus and Alnus, show increasing values. Numerous types of herbs and shrubs such as Asteraceae, Cichoriaceae, and Brassicaceae show increased values, which indicates an intensive spread of these species on the former arable land.
The increased solar activity during the Middle Ages led to a positive North Atlantic Oscillation, which supplied mild and humid airmasses to Middle Europe (Sirocko, 2010). The recurred warming, known as the Medieval Climate Optimum, resulted in the highest population density in Middle Europe up to that point in time (Grupe, 1986). The population growth provoked a notable extension of the cultivated landscape (Bork et al., 1998). The Rouer peat archive witnessed these changes, containing the highest Poaceae pollen percentages during that period in the whole record, and the lowest overall tree species abundances. Alnus nearly disappears from the record, presumably due to the transformation of the remaining alder forest stands into wet grasslands. The warm period is further expressed in the occurrence of charred particles, which show by far the highest accumulation rates between 800 and 1400 AD. This could be a possible sign of slash-and-burn agriculture.
The spreading of Quercus between 1500 and 1700 cal. AD may be a result of its use for housebuilding, coppicing practice, and for tanneries. Furthermore, oak stands were widely used for forest pasturing practices during the Middle Ages and into the Modern Era, especially for feeding pigs acorns (Sudhaus and Friedmann, 2015). The increase in Brassicaceae is indicative of the expansion of arable land since the Middle Ages.
The Little Ice Age marks the most recent climatic shift to cold and humid climates: winter temperatures were lower and there was a sharp increase in precipitation, which led to severe soil erosion (Slotboom and van Mourik, 2015). Increased Ti/coh values is evidence of enhanced sediment input into the Rouer peat layers. In these upper peat layers, presumably stronger decomposition obscured concurrent responses in δ13C or C/N. After 1800 cal. AD, large areas of heathland and grassland in the Gutland were reforested, mainly with Pinus and Picea. Sylviculture, grassland farming, and agriculture characterize the youngest Modern Era.
Conclusions
The investigated peatland represents an exceptional fen ecosystem in Luxembourg. Peatlands with such well-preserved peat deposits are extremely rare in Luxembourg and its surrounding landscapes. Therefore, it represents an important climate archive of supraregional importance, which offers unique insights into the origins of local peat accumulation, as well as into the environmental history of the Gutland region and beyond.
With the application of XRF analyses, it was possible to identify periods of intensified erosion at a high temporal resolution. The Ti/coh ratio is a useful indicator of fluctuations in the input of detrital minerogenic matter, which is triggered by climatic and environmental changes. The Ti/coh value fluctuations match the range and the oscillation of cold spells as recorded by the Greenland ice δ18O isotope values. The Fe/Ti ratio reflects fluctuations of in situ redox conditions, and therefore serves as an indicator for periodic dry events. Values of δ13C seemed to reflect conditions of decomposition, mainly oxic versus anoxic and thus congruent with Fe/Ti ratios, while values of C/N and values of δ15N appeared to be modified by other factors related to diagenesis or vegetation.
Verified by pollen analysis, the Rouer peatland record evidences prominent Late Pleistocene and Holocene climate oscillations. The accumulation of organically-enriched sediment started at about 13,800 cal. yr BP, when fluvial discharge within the valley floor had diminished. Until about 6000 cal. yr BP, climate had kept warming, but had been interrupted repeatedly by pronounced episodes with increased input of minerogenic matter into the peat matrix (12,700–11,800 cal. yr BP; 11,500–11,300 cal. yr BP; 11,100–10,800 cal. yr BP; 9300 cal. yr BP; 8200 cal. yr BP), indicated by sudden increases in Ti/coh values.
After 6000 cal. yr BP, an alder forest developed and peat formation started. Between 4200 and 2800 cal. yr BP, during the Bronze Age, changes in the pollen spectrum indicate an increasing clearance of woodlands. During the late Iron Age (2200–2100 cal. yr BP), the alder forest was cleared and sedges started to dominate the local peatland vegetation. The Roman period is marked by ongoing deforestation and intensification of agriculture. Lowest tree species abundancies are witnessed during the Middle Ages. The Modern Era is characterized by enhanced sediment input due to soil erosion. Pollen types from then on point to a landscape dominated by sylviculture, grassland farming, and agriculture.
In general, our study demonstrates that fen peat deposits can be valuable high resolution paleoclimate archives, which offer excellent opportunities to investigate the timing and character of climatic shifts in the Luxembourg Gutland and beyond. Without doubt, Rouer peatland represents a wetland habitat with a high conservation value. The exact knowledge of the evolution of the peatland and the effects of environmental changes on peat accumulation provides valuable information for species and biotope protection, as well as renaturation and management measures. Now that we know about the peat thickness and quality, this central area will certainly be preserved untouched and management by grazing and mowing will be maintained. The adjacent area not belonging to the study area has already been renaturalized in the meantime by SICONA. For this purpose, the soil that had been piled up there many decades ago was removed and irrigation measures carried out. The reintroduction of typical plants has also been initiated.
Supplemental Material
sj-bmp-1-hol-10.1177_0959683621994645 – Supplemental material for A 14,000 year peatland record of environmental change in the southern Gutland region, Luxembourg
Supplemental material, sj-bmp-1-hol-10.1177_0959683621994645 for A 14,000 year peatland record of environmental change in the southern Gutland region, Luxembourg by Karsten Schittek, Lelaina Teichert, Katrin Geiger, Klaus-Holger Knorr and Simone Schneider in The Holocene
Supplemental Material
sj-bmp-2-hol-10.1177_0959683621994645 – Supplemental material for A 14,000 year peatland record of environmental change in the southern Gutland region, Luxembourg
Supplemental material, sj-bmp-2-hol-10.1177_0959683621994645 for A 14,000 year peatland record of environmental change in the southern Gutland region, Luxembourg by Karsten Schittek, Lelaina Teichert, Katrin Geiger, Klaus-Holger Knorr and Simone Schneider in The Holocene
Footnotes
Acknowledgements
The authors give their sincere thanks to Bertil Mächtle, Sebastian Kock, and Fanny Schaul for great help during fieldwork. We also thank Sabine Stahl and Christian Ohlendorf for support and help with the calibration of the XRF results. We further acknowledge the support of Sinja Kraus, Hannah Siep, and Ahrabi Ananthan in the preparation of the pollen slides and help in the laboratory. Analyses of C and N contents and of the stable isotopic composition of the peat were carried out at the laboratory of the Institute of Landscape Ecology, University of Münster, with the help of Ulrike Berning-Mader. We also thank Caroline Grounds for the English proofreading. The authors thank the two anonymous reviewers of the manuscript for their helpful comments and recommendations, as well as Associate Editor Prof. Frank Chambers.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Funding by SICONA, the municipality of Redange, and the Department for the Environment of the Ministry of the Environment, Climate and Sustainable Development (MECDD).
Data accessibility
Multiproxy data from the Rouer peatland will be made available via the National Oceanic and Atmospheric Administration (NOAA) Paleoclimate database.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.