
Abstract
Despite the increasing interest in spring-fed fens’ sediments as a palaeoenvironmental archive, their potential as high-precision climatic records is rarely used to its full extent. Here, we present a detailed early to mid-Holocene record of environmental changes in the Turtul hanging spring-fed fen, northeastern Poland, to test whether the well preserved and accurately subsampled alternating peat and tufa sediments allow us to recognise short-duration local and regional climatic changes. Our reconstructions are based on a loss-on-ignition analysis of the sediment, carbon and oxygen stable isotope values of tufa, and biotic proxies: plant macrofossil and malacological analyses. A detailed environmental reconstruction was possible due to the combination of detailed sampling (1 cm intervals) and high sediment accumulation rates. The two sediment sequences collected from the fen revealed the congruent palaeoenvironmental history; however, the temporal shift associated with the distance of the coring site from the water outflow was also apparent, stressing the need for careful selection of the coring site in palaeoenvironmental research. Peat deposition at Turtul started at 10,300 cal yr BP; however, tufa precipitation did not begin before 9250 cal yr BP. During the most active tufa deposition, from 7850 to 6100 cal yr BP, a high sedimentation rate was observed (0.11–0.25 cm yr−1). We found several abrupt and short-duration declines in carbonate deposition corresponding to climatic fluctuations recorded in geological archives from the southeastern Baltic region. The carbonate drops at 8200–8130, 7900–7850, 7400–7300, 6600–6500, 5980–5870 and 5400 cal yr BP were associated with climate cooling or drying, whereas those at 10,200–9250, 8450–8340 and 6160–6100 cal yr BP can be attributed to drying with no cooling. We conclude that the evident sensitivity of the Turtul fen to climatic shifts that emerge from our high-resolution (one sample every 17 years on average) reconstruction complements the still rare high-resolution palaeoenvironmental records in the area.
Keywords
Introduction
High-resolution records are of prime importance in palaeoenvironmental studies. Among the terrestrial archives in the intermediate climatic zone, laminated lake sediments (Poraj-Górska et al., 2017) and tree rings (Ljungqvist et al., 2020), each with yearly or even seasonal time controls, are of the greatest interest. High-resolution records can also be derived from deposits in which the annual time control is absent. Here, the high sediment accumulation rate enables detailed palaeoenvironmental reconstructions. Among such archives are spring-fed fen deposits characterised by intensive CaCO3 deposition (Dobrowolski et al., 2016; Gałka et al., 2018).
Calcareous spring-fed fens are a unique type of marsh ecosystem, in which the continuous supply of carbonate-enriched groundwater is of prime importance and ensures CaCO3 precipitation and the presence of specific fauna and flora (Grootjans et al., 2015; Succow, 1988). The supply of the groundwater is either descensive, at the hanging spring-fed fens located at slopes in river valleys (Osadowski et al., 2009) or ascensive, when the confined groundwater finds its way to the surface through a hydrogeologic window in impermeable sediments or through a tectonic fault (Dobrowolski et al., 2016). Calcareous spring-fed fens are usually small (<1 ha) and restricted to areas in direct proximity to the active groundwater outflow (Dobrowolski, 2011). Carbonates and Ca2+ ions supplied at the fens are derived from the dissolution of carbonate rocks occurring in the aquifer bedrock; therefore, the geographic location of the spring-fed fens is widespread across the globe, including different continents (Bedford and Godwin, 2003; Hájková et al., 2012), wherever carbonate rocks occur. In east-central Europe, spring-fed fens are known from different locations, including young and old glacial areas (Wołejko et al., 2018), karstic regions (Dobrowolski et al., 2016) and mountains (Hájková et al., 2015). Precipitation of CaCO3, most commonly referred to as tufa (Grootjans et al., 2015; Pedley, 2009), is controlled by two major variables at the fen. First, it is sensitive to largely climatically driven changes in the groundwater level and groundwater circulation intensity, as it controls the chemical dissolution of carbonate rocks and the supply of Ca2+-supersaturated waters in the spring (Capezzuoli et al., 2014; Wołejko et al., 2019). Second, if the water supply is sufficient, temperature is also important because the greater the difference between the groundwater and surface air temperature is, the more intensive the loss of groundwater CO2 and eventually CaCO3 precipitation (Andrews and Brasier, 2005; Pedley, 2009). Tufa precipitation is most intensive in conditions of active water supply at the fen and high temperatures during spring and summer. CaCO3 deposition is further enhanced by CO2 assimilation by micro- and macroflora, which become tufa-coated and commonly preserve macrophyte morphologies (Capezzuoli et al., 2014; Pedley, 2009). When, for instance, in response to decreased effective precipitation, groundwater outflow becomes insufficient to sustain carbonate precipitation, and/or temperature drops significantly, tufa formation ceases, and peat deposits occur in the centre of the fen (Grootjans et al., 2006). The outcome of the combined influence of changes in the water supply and temperature is the formation of alternating layers of peat and carbonate deposits, which is characteristic of spring-fed fen sedimentary deposits (Dobrowolski, 2011; Succow, 1988). The sensitive reaction to climatic changes makes interbedded peat and calcareous tufa deposits a valuable archive of environmental changes throughout the Holocene (Hájek et al., 2016; Limondin-Lozouet et al., 2013; Šolcová et al., 2018). In palaeoenvironmental studies, the sedimentological record is supported by the analyses of biotic (plant macrofossils and molluscs) and abiotic (sediment geochemistry, including the records of C and O stable isotopes) proxies, a combination that assures more detailed palaeoclimatic and palaeoecological reconstructions (Dabkowski et al., 2019; Dobrowolski et al., 2019; Gałka et al., 2021).
Although fens usually cover small areas, they are characterised by the natural spatial variability of the ecological conditions controlled by the proximity to the water outflow and the topography of the fen (Hájek et al., 2006). On the top of the fen mound, where groundwater emerges at the surface, small ponds commonly occur. In contrast, the slopes of the fen are usually swampy but with no standing water. This has a profound influence on the ecological conditions and influences the habitat of fauna and flora and the sedimentary environment, which are eventually recorded in the deposits (Wołejko et al., 2019). Therefore, proper selection of the coring site is crucial.
Despite the increasing interest in spring-fed fens’ sediments as a palaeoenvironmental archive (Blaus et al., 2020; Dobrowolski et al., 2019; Jamrichová et al., 2018), their potential as high-resolution climatic records has rarely been used to their full extent. In numerous fens, where the high sediment accumulation rate, characteristic of the actively precipitating CaCO3 sites, is observed (i.e. 5–10 yr cm−1, Dobrowolski et al., 2016, 2019; Gałka et al., 2018, 2021; Limondin-Lozouet et al., 2013), a sampling resolution on the order of a decade or less is achievable. However, this potential is lost, and the resolution decreases to 100 years or more when 5- to 10-cm-thick sediment samples are used (Hájková et al., 2012; Jamrichová et al., 2018). Some of the spring-fed fens’ deposits, although covering long time frames, for example, the entire Holocene, are characterised by a low sediment accumulation rate on the order of 60–100 yr cm−1, and the resolution decreases to 350–550 years per sample (Dabkowski et al., 2019; Hájek et al., 2016; Hájková et al., 2015). The existing high-resolution records of the spring-fed fens (10–20 years per sediment sample), allowing for the recognition of even short-duration climatic shifts, are mostly restricted to the upper Holocene and cover the period of the last ca. 4000 cal yr BP or less (Gałka et al., 2018, 2021; Jamrichová et al., 2014). To date, in central Europe, the high-resolution spring-fed fen sediment records of the lower Holocene (25–40 years per sediment sample) were studied by Šolcová et al. (2018) in southern Slovakia and Dobrowolski et al. (2019) in northern Poland (Spurgle site).
Of the two genetic types, in palaeoenvironmental studies, cupola spring-fed fens are favoured over the hanging type (Dobrowolski et al., 2019), which is believed to be prone to erosion and easily changes from peat accumulating to eroding systems (Wołejko et al., 2019). Additionally, it was observed (Hájková et al., 2012) that fens located on slopes may undergo CaCO3 dissolution in their higher located sites and precipitation in the lower sites. This may strongly influence the palaeoenvironmental record, thereby causing discontinuity of the sedimentary record (Hájková et al., 2015). However, under moderate water flow and when the fen does not occur on steep slopes, sedimentation can be continuous (Wołejko et al., 2019), and the sediment sequence can be successfully used in palaeoenvironmental research.
This study investigated the environmental history of the Turtul hanging spring-fed fen (northeastern Poland) by employing high-resolution geochemical (loss on ignition and C and O stable isotopes), plant macrofossil and malacological analyses. We aimed to complement the still-lacking high-resolution palaeoenvironmental records and test whether the well-preserved and accurately subsampled (1 cm intervals) alternating peat and tufa sediments allow us to recognise the short-duration local and regional climatic changes. The spatial variability of environmental conditions at the fen is tested by analyses of two sediment cores – from the central part and the slope of the spring-fed fen cupola.
Study site
The Turtul (Tu, 54°13′22.5″ N, 22°48′9.96″ E, 197 m a.s.l.) groundwater-fed fen investigated in the present study is located in northeastern Poland in the mosaic landscape of Suwalski Landscape Park in the Turtul settlement (Figure 1). The site is included in the network of protected areas Natura 2000.

(a) Location of the Turtul spring-fed fen and sites cited in the text across central Europe: Bobolice, Ogartowo, Spurgle (Dobrowolski et al., 2019); Komarów (Dobrowolski et al., 2016); Jaczno (Makri et al., 2021); Purwin (Apolinarska and Gałka, 2017); Kojle (Gałka, 2014; Gałka et al., 2014, 2015); Szurpiły (Kinder et al., 2020); Żabińskie (Zander et al., 2021); Kurajnovas (Heikkilä and Seppä, 2010); Laihalampi (Heikkilä and Seppä, 2003). (b and c) Digital elevation models based on LiDAR data with location of the Turtul spring-fed fen, and cores Tu 1 and Tu 2.
The landscape of the area was formed during the Pomeranian Phase (ca. 16–17 10Be/36Cl kyr BP; Dzierżek and Zreda, 2010; Rinterknecht et al., 2006) of the Weichselian glaciation (Marks, 2012). The activity of the ice sheet and its subglacial waters resulted in an uneven topography characterised by numerous morainic and kame hills, with maximum heights of 270 m a.s.l., separated by depressions (150–170 m a.s.l.) where wetlands and lakes occur. Glacial sands, gravels and tills are the dominant types of sediments in the area (Krzywicki, 1990; Laskowicz, 2012).
The Turtul fen is of the hanging groundwater-fed fen type developed on the eastern slopes of the Czarna Hańcza River valley. Water supply to the fen is either direct, from meteoric waters, but mainly indirect, from the outflow of descending groundwaters of the local near-surface aquifer located in the slopes of the Czarna Hańcza River valley cutting Quaternary unconsolidated deposits (Wołejko et al., 2018), and from the surface runoff from the slopes of the river valley (Figure 1). The surface of the fen is swampy and has small, several cm deep water pools scattered on the surface.
In the Turtul fen, tall sedge vegetation dominates. The top of the peat mound, in the spot where the sediment cores were collected, has sparse vegetation on swampy peat. Carex acutiformis, Equisetum palustre, E. fluviatile, Cardamine amara and Impatiens noli-tangere are the most common plant species there. In the weakly developed moss layer, brown moss species grow, mostly Plagiomnium elatum and Brachythecium rivulare. Towards the Czarna Hańcza River, mainly monodominant stands of Carex acutiformis occur. In the moss layer, only some specimens of Plagiomnium elatum, and Climacium dendroides are found. The richest vegetation occurs on the northern slope of the fen, where the ground is the softest and most swampy because of the active spring discharge. Carex paniculata dominates there. Other common plant species include Geranium robertianum, Angelica sylvestris, Equisetum palustre and Myosotis palustris. The moss layer is moderately developed and mainly consists of Plagiomnium elatum. Almost the whole surface of the fen is under the canopy of deciduous trees, including Alnus glutinosa, Betula sp. and Salix sp.
Both Atlantic and continental air masses influence the temperate climate of the study area; however, eastern air masses prevail. The area receives mean annual precipitation amounts of approximately 650–700 mm, with the maximum and minimum precipitation occurring in July and February, respectively. The mean annual air temperature is 6.5°C, and the monthly averages range between −5°C in January and 16 –17°C in July (Lorenc, 2005).
Material and methods
Field methods
Coring
Two sediment sequences, Tu 1 and Tu 2, were drilled using two 50 cm long, 9 and 5 cm in diameter, Instorf type peat corers. Tu 1 was retrieved at the highest point of the spring-fed fen (54°13′22.5″ N, 22°48′9.96″ E, 197 m a.s.l.), whereas Tu 2 about 10 m apart, at the NW slope of the fen. To obtain a continuous sediment record the sediment core segments were collected from two parallel holes, about 20 cm distant, with offset drive depths. Diameter of the Tu 1 core amounted to 9 cm between the surface and 3.5 m of depth, and 5 cm between 3.5 and 4 m. The 5 cm in diameter peat corer was used for drilling the 4 m long Tu 2 sediment sequence. The contact between the highly compacted decomposed peat present in the lowermost section of the Tu 2 sediment sequence and siliciclastic sediments was reached using a Dutch auger. The sediment cores’ segments were placed in U-shaped PVC gutters, wrapped in the plastic film and stored in the cold room until further subsampling.
Laboratory methods
Core cutting and subsampling of the sediments
In the laboratory, the cores were sliced into 1-cm thick fragments. Each sediment slice was subsampled for the analysis of plant macrofossils and molluscs (about 2/3 of the total volume, that is, approximately 20 cm3 in Tu 1 core between the surface and 350 cm, and approximately 7 cm3 in Tu 1 core between 350 and 400 cm, and the Tu 2 core), and the sample for geochemical and isotopic analysis (about one-third of the volume, that is, approximately 10 cm3 in Tu 1 core between the surface and 350 cm, and approximately 3 cm3 in Tu 1 core between 350 and 400 cm, and the Tu 2 core). Samples for plant macrofossil and malacological analysis were washed and sieved under a warm-water spray using a 0.20-mm mesh sieve. Samples for geochemical and isotope analyses were dried at 40°C and pulverised in an agate mortar (Pulverizette 2). The lithology of the sediments was described during sediment subsampling.
Radiocarbon dating
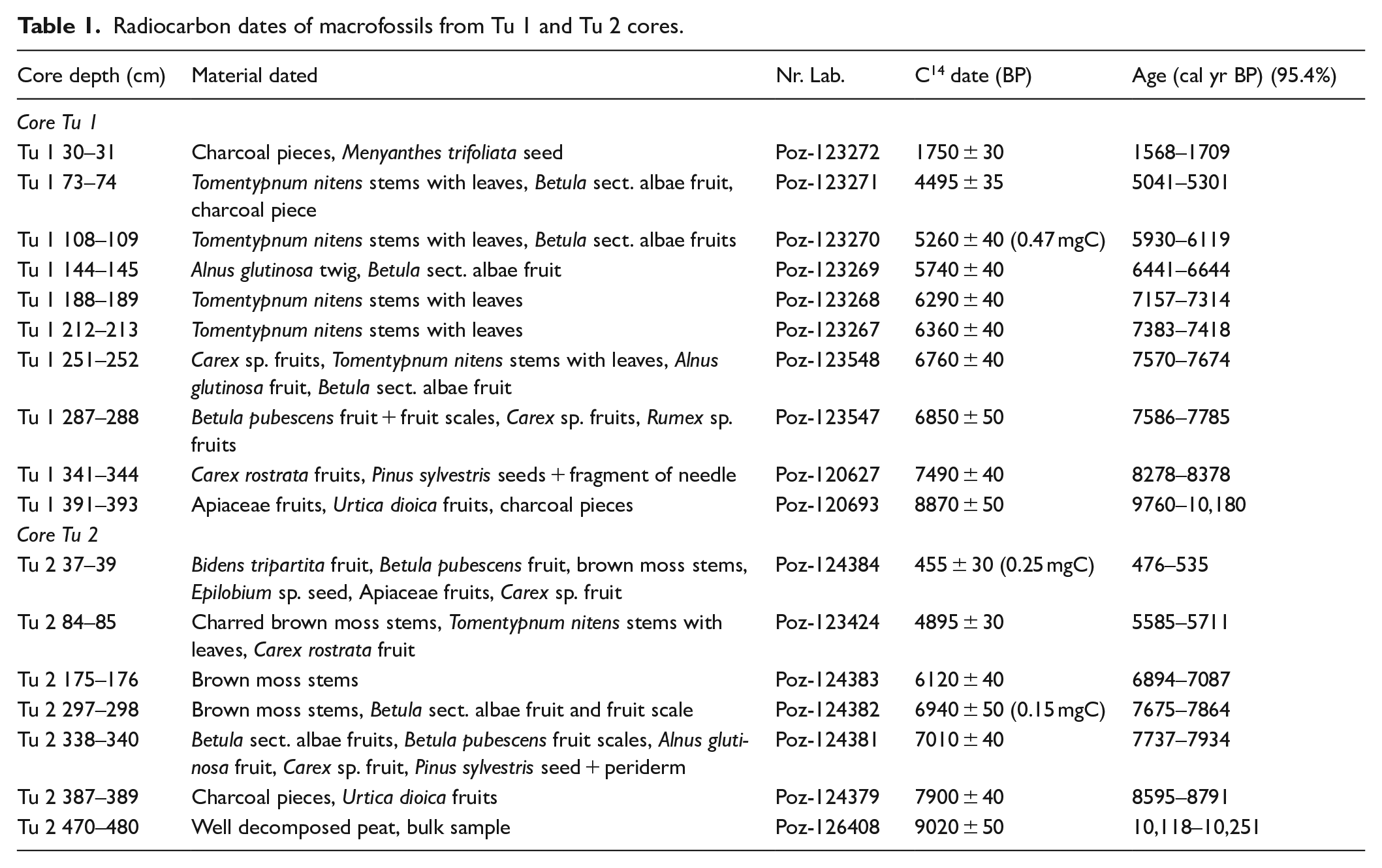
To establish the chronology of the studied sediment sequences, radiocarbon dating of plant macrofossils was performed in the Poznań Radiocarbon Laboratory. Ten samples from the core Tu 1 and seven from the core Tu 2 were used. The resulting conventional radiocarbon dates were calibrated with Bchron (Haslett and Parnell, 2008) applying the IntCal20 atmospheric curve (Reimer et al., 2020). The age-depth model was constructed using the rbacon package version 2.3.9.1 (Blaauw and Christen, 2019) in R ver. 3.5.1. (R Core Team, 2018). The rbacon package performs Bayesian age-depth modelling that includes the dating probability distribution. Given the substantially lower accumulation rates of peat comparing to carbonates on spring-fed fens, we set boundaries in rbacon between the main lithological units to force the software to model sharp changes in accumulation rates at these depths. A mean value for the sediment accumulation rate (SAR), expressed as cm year−1, was calculated based on the mean values of the probability distribution of the modelled age (μ) for each depth, for which a date was modelled. The applied formula was as follows: SARdepth = 1 cm/(μdepth – 0.5 cm – μ depth + 0.5 cm), where μdepth is the mean value of the modelled age for a given depth. For better readability of the following sections, μ values rounded to tens were selected as reflecting the modelled age, which was expressed as cal yr BP (Before Present).
Plant macrofossils analysis
Plant macrofossils were analysed at 1-cm intervals totalling 395 samples and 400 samples in Tu 1 and Tu 2 cores, respectively. Plant macrofossils were identified under a Nikon SMZ800 stereoscopic microscope at the magnification of 10–200× using the appropriate keys, including Velichkevich and Zastawniak (2006, 2008). The plant macrofossils have been summarised in the diagrams in absolute numbers. The volume percentages of the different vegetative remains were estimated to the nearest 5% or presence/absence. Numbers of seeds, fruits and bud scales were counted separately, employing a stereoscopic microscope. Macroscopic charcoal pieces (>1 mm) were also counted during plant macrofossils analysis, and their presence provides information on past local fire occurrence (Mooney and Tinner, 2011). The ecological requirements of several key plant species (Ellenberg et al., 1991; Zarzycki et al., 2002) were used to distinguish wet and dry phases in the peatland’s development. The nomenclature followed are Tutin et al. (1993) for vascular plants, Hill et al. (2006) for mosses, and Hájek et al. (2006) for fen mosses. We applied stratigraphically constrained cluster analysis (CONISS) to square root transformed macrofossils data assessing the number of significant zones by the broken stick model (Bennett, 1996). The analysis was performed with rioja package version 0.9–15.1 (Juggins, 2017) in R ver. 3.5.1. (R Core Team, 2018). The final zonation of macrofossil data relied on the combination of CONISS analysis and visual inspection of macrofossil diagrams.
Malacological analysis
After selecting the plant macrofossil remains, sediment samples from Tu 1 and Tu 2 sediment sequences were examined for mollusc shells. Complete mollusc shells and recognisable shell fragments were hand-picked and identified under a low-power binocular microscope (Zeiss Stemi 2000-C) using the reference collection of Institute of Geology AMU in Poznań as well as keys and atlases (Piechocki and Wawrzyniak-Wydrowska, 2016; Welter-Schultes, 2012). The mollusc nomenclature follows that of Welter-Schultes (2012). The mollusc remains are expressed as absolute numbers. To better illustrate the response of the mollusc population to paleoenvironmental changes, molluscs were divided into groups according to their environmental requirements (Alexandrowicz and Alexandrowicz, 2013), that is, land, hygrophilous and aquatic species. We applied stratigraphically constrained cluster analysis (CONISS) to square root transformed malacological data assessing the number of significant zones by the broken stick model (Bennett, 1996). The analysis was performed with rioja package version 0.9–15.1 (Juggins, 2017) in R ver. 3.5.1. (R Core Team, 2018).
Geochemical analysis
Loss on ignition (LOI)
Samples for LOI analysis were taken at 1-cm intervals in Tu 1 core and 10-cm intervals in Tu 2 core. Pulverised sediment samples were combusted at 550°C for 4 h and subsequently at 950°C for 2 h to determine the geochemical composition of the sediments (Heiri et al., 2001). The weight loss at 550°C is presumed to represent the percentage of organic matter (OM). The carbonate (CO32−) content was calculated by multiplying the mass of CO2 evolved in the second step of the analysis (the weight loss at 950°C) by 1.36. Finally, the CaCO3 content was calculated by multiplying the CO32− content by 1.66. The residue represented mainly by organic silica, siliciclastic grains and various precipitates like iron oxides was calculated by subtracting CaCO3 and OM contents from the total sample. The results are presented as the percentage concentrations of organic matter, CaCO3 and residue.
Stable isotope analyses of carbonates
Stable carbon (δ13C) and oxygen (δ18O) isotopic compositions were measured in the bulk carbonate tufa after the LOI analysis which allowed to determine the CaCO3 share in the sediments. Samples for isotope analysis were taken at 2-cm intervals, wherever the CaCO3 content in the sediments exceeded 2% (agreed with the stable isotope laboratory as a threshold value of accurate analysis). The δ13C and δ18O values of the carbonates were measured using a Gas Bench II hooked up to a Finnigan MAT 253 gas source mass spectrometer (both Thermo Fischer). Details concerning the analytical setup are given in Spötl and Vennemann (2003). Isotope values are reported relative to VPDB based on a NBS-19 calibrated Carrara marble value of +2.02‰ (δ13C) and −1.76‰ (δ18O) (Fiebig et al., 2005). On average, replicated precision (1SD, multiple measurements of the Carrara standard) was better than ±0.06‰ for δ13C and better than ±0.08‰ for δ18O.
Results
Lithology, chronology and sediment accumulation rate
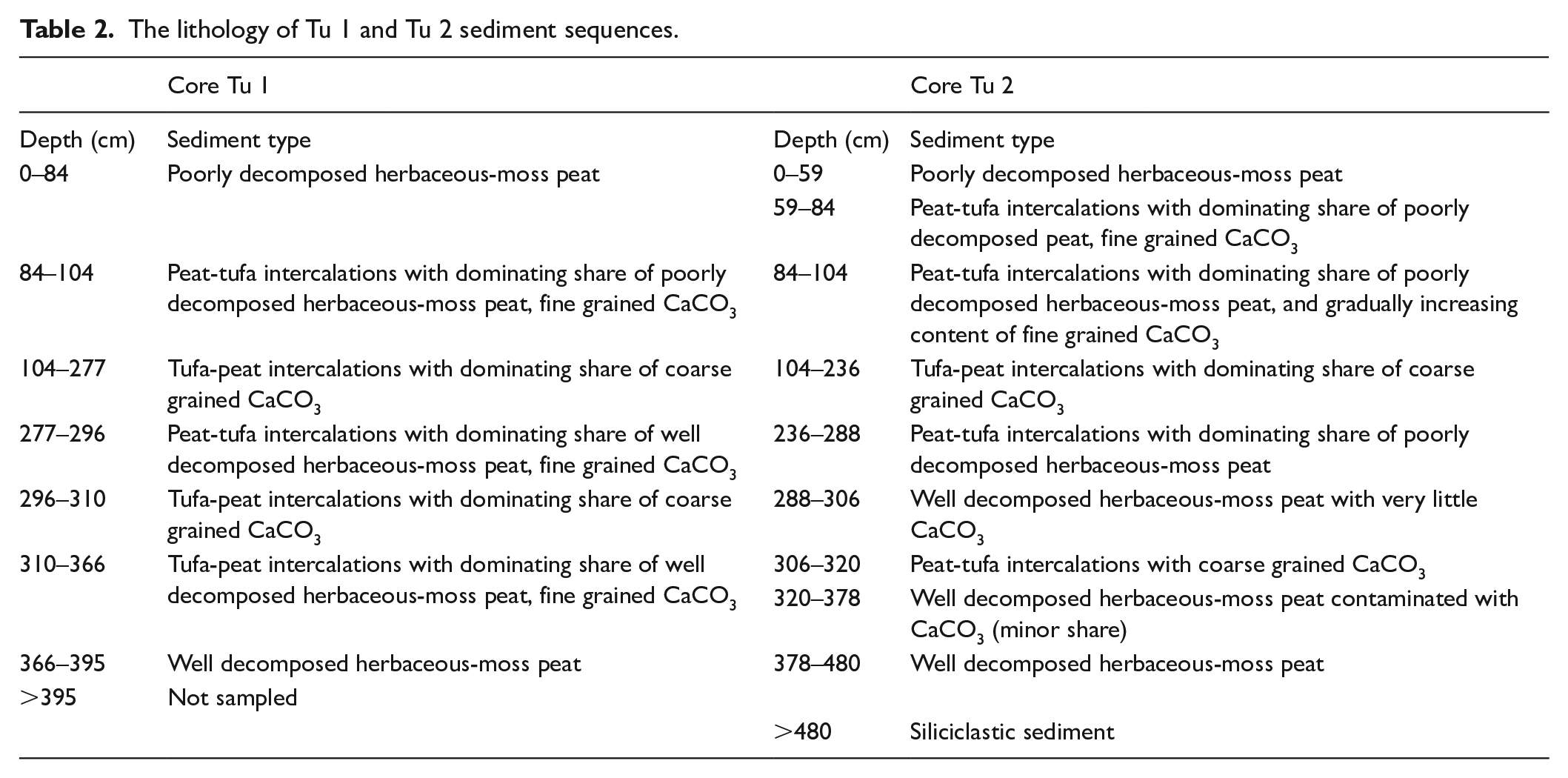
A detailed description of the lithology in the Tu 1 and Tu 2 sediment sequences is shown in Table 2, and the organic matter and CaCO3 contents are shown in Figure 2. Despite some differences in the depth where lithostratigraphic shifts occur, the sedimentary sequences share the same type and succession of sediments. The fine-grained clastic bedrock sediments are reached 480 cm below the surface in the Tu 2 core; however, the bottom of organic sediments are not reached in the Tu 1 core. The biogenic sediments in both sequences start with well-decomposed herbaceous moss peat (mean organic matter content of 84% in Tu 1). The peat is overlain by peat-tufa intercalations with varying proportions between the two sediment components. The grain size of the tufa changes from silty to coarse, with frequent fragments exceeding 1 cm in diameter. The significant enrichment in CaCO3 is noted from 366 to 84 cm in Tu 1 and between 378 and 59 cm in Tu 2; however, values fluctuate from 17% to 94%, with a mean of 73%, and from 22% to 90%, with a mean value of 72%, in Tu 1 and Tu 2, respectively. The highest percentage share of CaCO3 is observed between 277 and 104 cm in Tu 1 (37–94%, mean of 82%) and from 288 to 104 cm in Tu 2 (22–90%, mean of 81%). After 79 cm in Tu 1 and 59 cm in Tu 2, the shift to poorly decomposed herbaceous moss peat occurs with only a trace share (usually below 1% and in all samples below 5%) of CaCO3 and a dominance of organic matter, which is 79% on average.
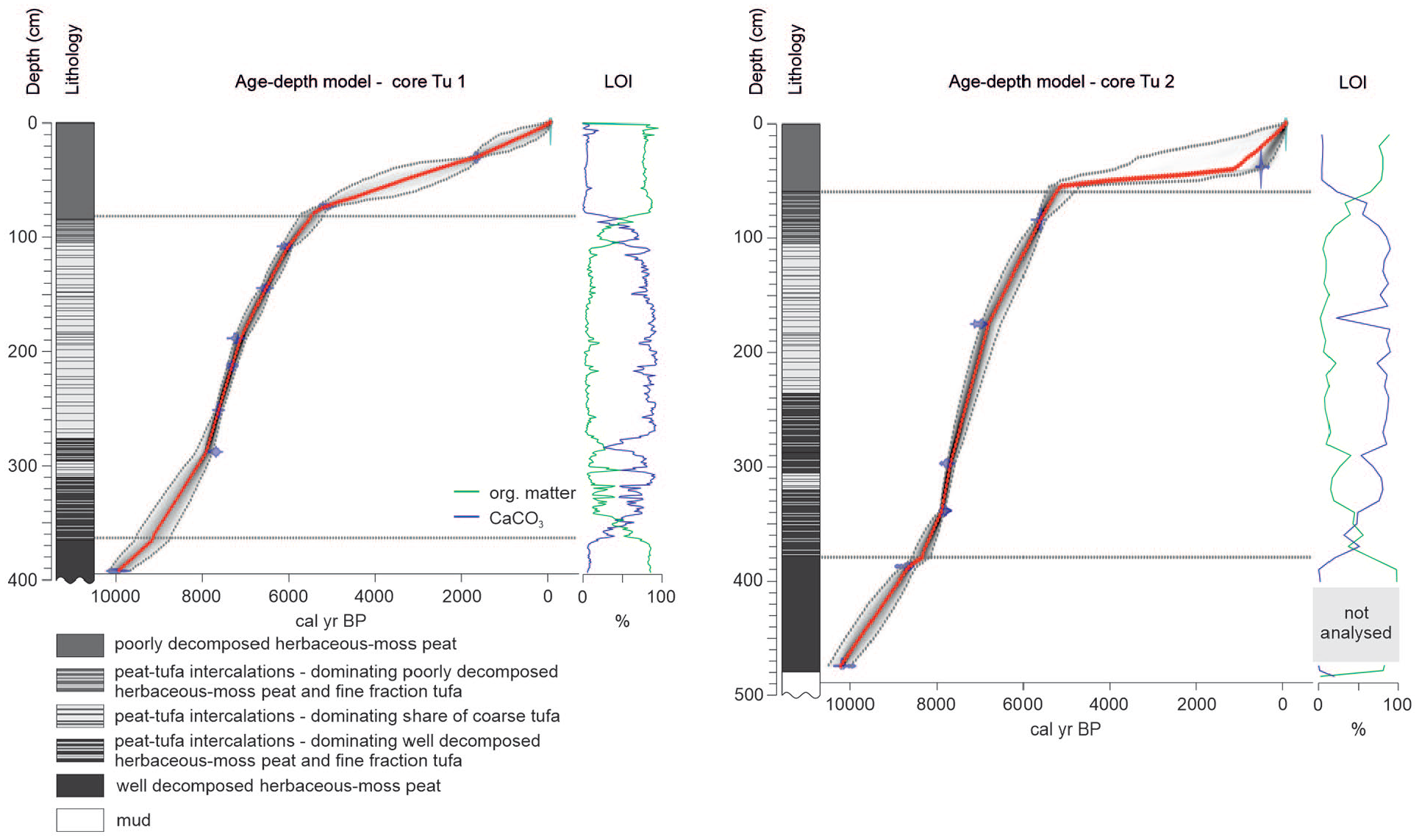
Age-depth models of the Tu 1 and Tu 2 cores presented against lithology of the sediment sequences. The age-depth models constructed using the rbacon R package are based on 10 (Tu 1) and 7 (Tu 2) AMS 14C dates. The calibrated ages are shown in transparent blue. Black stippled lines indicate 95% confidence intervals. The red curve shows a single ‘best’ model based on the weight mean age for each depth. The horizontal lines at the transition between tufa and peat deposits visible in the models mark the depths of boundaries we set to force the software to model rapid accumulation rate shifts (see Method section for rationale).
The results of the radiocarbon dating of the Tu 1 and Tu 2 cores are presented in Table 1, and the age-depth models are shown in Figure 2. The ages of all the radiocarbon dates in both sediment cores increase with depth, reaching 9760–10,800 cal yr BP and 10,118–10,251 cal yr BP at the bottoms of Tu 1 and Tu 2, respectively. Given that no outlier dates were found, we used all the samples to construct age-depth models with rbacon (Blaauw and Christen, 2019). A clear difference in accumulation rates between the peat intervals and the intercalating peat-tufa sediments, emerging from the preliminary inspection of our radiocarbon dates, justifies setting the boundaries in the rbacon models described in the methods section. The results of our Bayesian age-depth modelling reveal that in both sediment cores, the accumulation rate (SAR, Figure 2) was low in peat (0.1–0.6 mm year−1) and increased markedly in the peat-tufa intercalations (0.5–2.5 mm year−1). The highest SAR in the Tu 1 core, 1.1–1.4 mm year−1, was observed between 189 and 284 cm (ca. 7100–7870 cal yr BP), whereas in the Tu 2 core, the highest SAR of 1.7–2.5 mm year−1 was observed between 295 and 339 cm (ca. 7680 and 7910 cal yr BP). According to our models, the accumulation of intercalating peat-tufa sediments corresponds with the periods of 9180–5400 cal yr BP and 8350–5300 cal yr BP in the Tu 1 and Tu 2 sediment cores, respectively.
Radiocarbon dates of macrofossils from Tu 1 and Tu 2 cores.
Considering the high sediment accumulation rate in Turtul during the early and middle Holocene (Figure 2), enabling the high-resolution reconstruction of the environmental conditions during the deposition of the intercalating peat-tufa sediment, the plant macrofossil and malacological data presented here are restricted to samples older than 5000 cal yr BP.
Plant macrofossil analysis
Core Tu 1
Five zones in the local vegetation development were determined based on a visual inspection and CONISS analysis of macrofossil data of core Tu 1 (Figure 3). In the Tu 1-ma-1 zone (ca. 10,000–9150 cal yr BP; 395–364 cm), herbs, represented mainly by seeds of Apiaceae, are replaced by wood, accompanied by charcoal pieces (>1 mm), which have the greatest share of the sediment sequence. The share of Urtica dioica and sclerotia of fungi that increases concurrently was also noted. The Tu 1-ma-2 zone (9150–8830 cal yr BP; 364–343 cm) is dominated by Carex rostrata and wood pieces and frequent macrofossils of trees: Alnus glutinosa, Pinus sylvestris and Betula sect. albae. Within zone Tu 1-ma-3 (8830–7620 cal yr BP; 343–252 cm), abundant macrofossils of B. sect. albae and Betula pubescens occur. Stellaria palustris, which is characterised by the most continuous record, is accompanied by Rumex acetosella, U. dioica, C. rostrata, Carex paninculata/appropinquata, and Caltha palustris. In the upper part of the zone, Tu 1-ma-3 stems of brown mosses and Tomentypnum nitens occur for the first time. T. nitens becomes abundant and dominates in zone Tu 1-ma-4 (7620–6200 cal yr BP; 252–122 cm). Abundant seeds of Cladium mariscus occur at ca. 7000 cal yr BP. Zone Tu 1-ma-5 (6200–4620 cal yr BP; 122–67 cm) is dominated by herb rootlets and epidermis and is macrofossil-poor. A single seed of C. mariscus occurs at ca. 5540 cal yr BP.
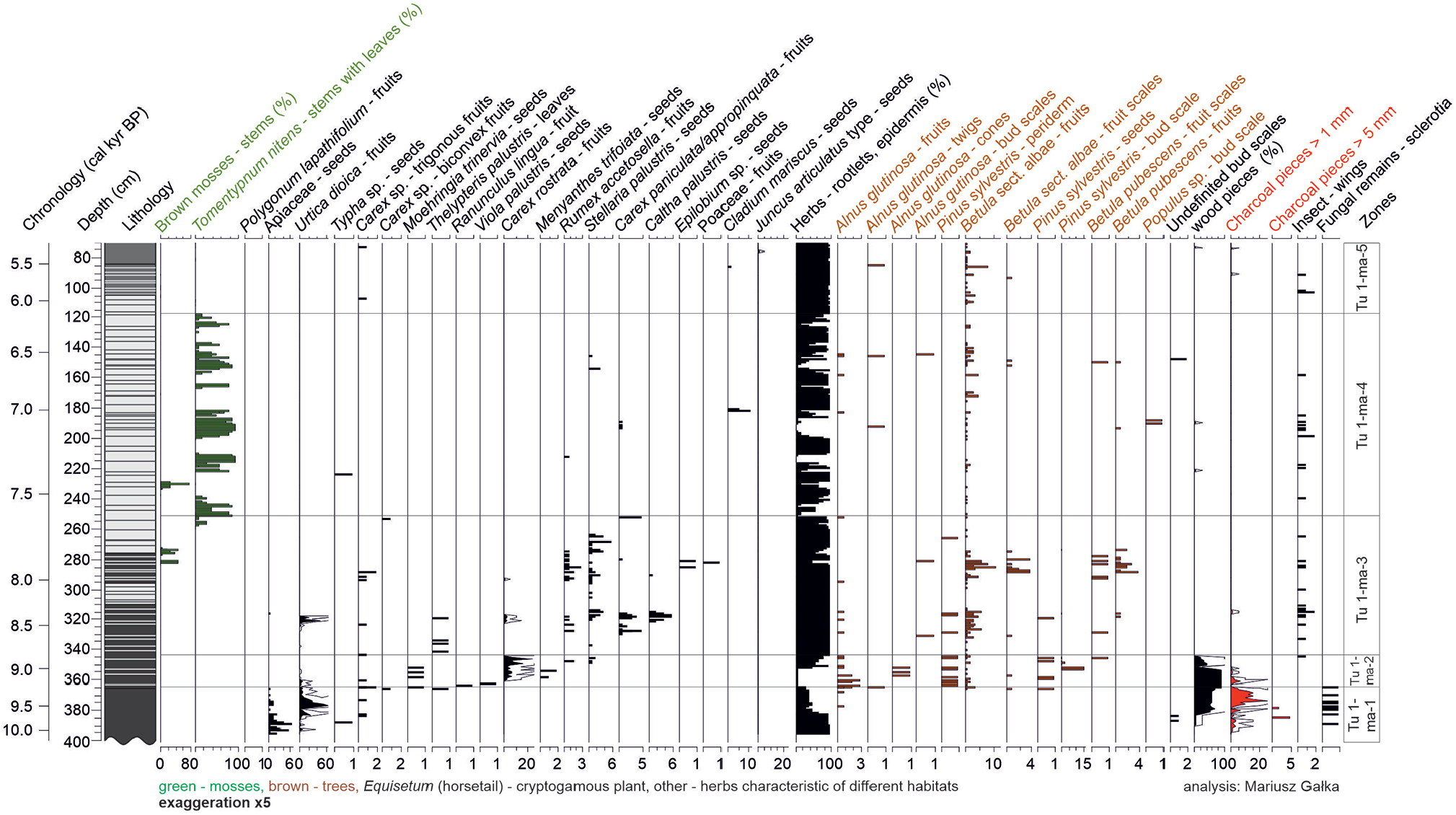
Local plant succession in Turtul spring-fed fen, core Tu 1, based on plant macrofossils presented along with sediment lithology. Taxa with (%) are estimated volume percentages, and all others are counts (with X-axis scale labels; note scale differences).
Core Tu 2
Five zones in the local vegetation development were determined based on a visual inspection and CONISS analysis of macrofossil data of core Tu 2 (Figure 4). Sparse plant macrofossils in zone Tu 2-ma-1 (8890–8360 cal yr BP; 400–380 cm) are dominated by fruits of U. dioica. Very abundant charcoal pieces occur here. The Tu 2-ma-2 zone (8360–7860 cal yr BP; 380–330 cm) is dominated by C. rostrata and frequent macrofossils of trees: A. glutinosa, P. sylvestris and B. sect. albae. S. palustris, C. paninculata/appropinquata, Menyanthes trifoliata, and C. palustris also occur in this zone. Abundant stems of brown mosses characterise zone Tu 2-ma-3 (7860–7550 cal yr BP; 330–277 cm). Single finds of T. nitens are also present. Fruits of B. sect. albae are the most abundant here. Tu 2-ma-4 (7550–7120 cal yr BP; 277–217 cm) is barren of plant macrofossils with only a single seed of Typha sp. present. In zone Tu 2-ma-5 (7120–5530 cal yr BP; 217–78 cm), plant macrofossils are rare; only occasional increases in brown mosses, T. nitens, and Equisetum sp., are observed. An abrupt increase in B. sect. albae and wood pieces from ca. 5650 to 5600 cal yr BP is concurrent with abundant charcoal pieces. The assemblage changes with the transition to peat, in which fossils of the Juncus articulatus type are abundant.

Local plant succession in Turtul spring-fed fen, core Tu 2, based on plant macrofossils presented along with sediment lithology. Taxa with (%) are estimated volume percentages, and all others are counts (with X-axis scale labels; note scale differences).
Malacological analysis
Core Tu 1
Five statistically significant mollusc zones were distinguished in core Tu 1 based on CONISS analysis (Figure 5). Zone Tu 1-moll-1 (9150–8360 cal yr BP; 364–315 cm) is characterised by a high diversity and abundance of aquatic mollusc species, including mainly Anisus leucostoma, Anisus vortex, Valvata cristata, Radix labiata, Pisidium casertanum and hygrophilous Succinea putris. In zone Tu 1-moll-2 (8360–7740 cal yr BP; 315–267 cm), the diversity and abundance of aquatic species declines; however, Galba truncatula becomes common, accompanied by Bathyomphalus contortus and hygrophilous species: Carychium minimum, Euconulus alderi and S. putris. In zone Tu 1-moll-3 (7740–7240 cal yr BP; 267–206 cm), the sparse mollusc assemblage is composed of hygrophilous (C. minimum and E. alderi) and land (V. pulchella and N. hammonis) snails. Zone Tu 1-moll-4 (7240–6850 cal yr BP; 206–169 cm) is dominated by aquatic Stagnicola palustris. The abundance of hygrophilous C. minimum and E. alderi increases. Within zone Tu 1-moll-5 (6850–5510 cal yr BP; 169–84 cm), a gradual change from a mollusc assemblage dominated by hygrophilous species, including the endemic and protected Vertigo genesii/geyeri, to land snails, namely, V. pulchella, N. hammonis, Vitrea crystallina and slugs, is observed. Protected Vertigo angustior occurs here.
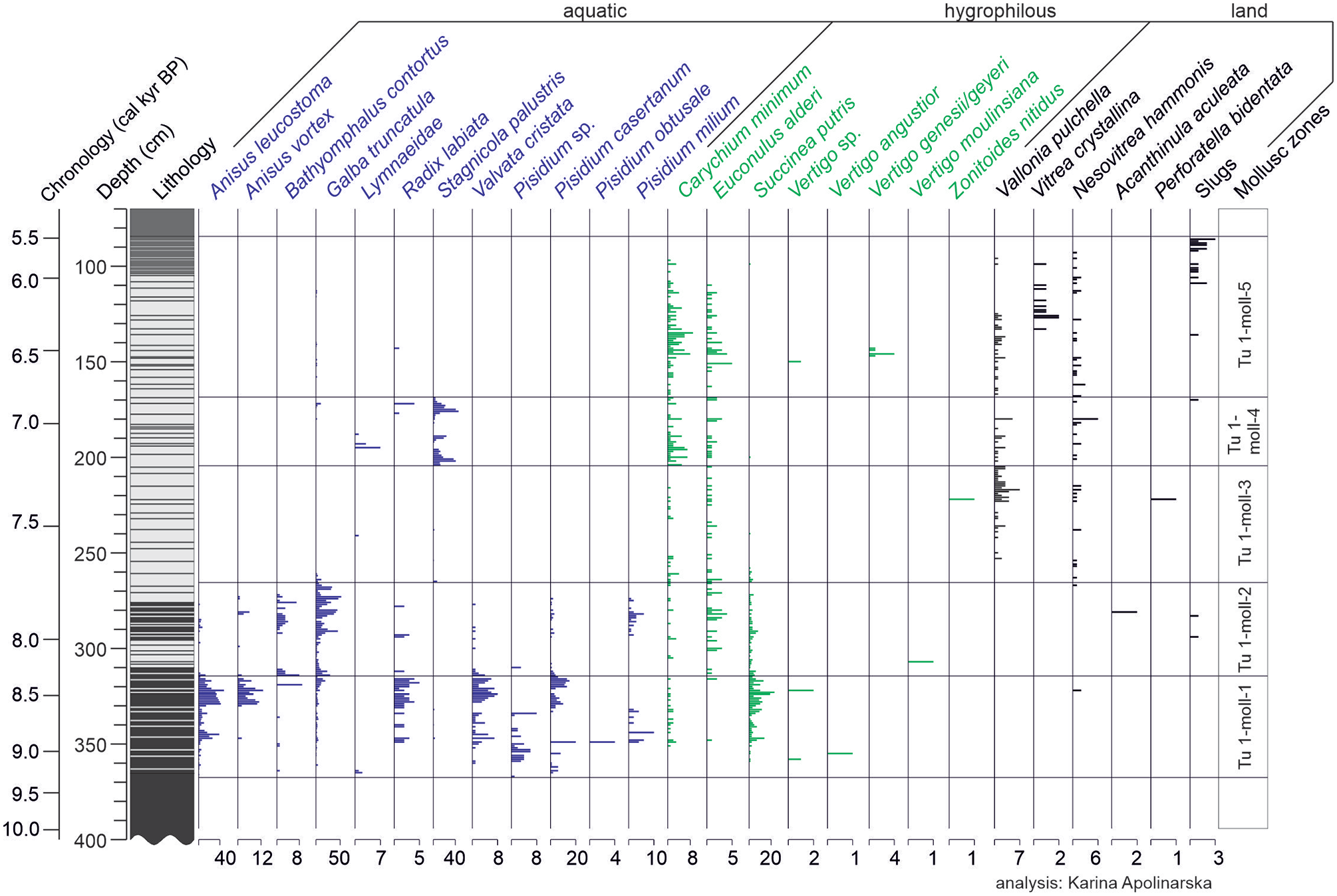
Local mollusc succession in Turtul spring-fed fen, core Tu 1, presented along with sediment lithology. Mollusc assemblages divided into aquatic, hygrophilous and land species. Taxa presented as shell counts (with X-axis scale labels; note scale differences).
Core Tu 2
Four statistically significant mollusc zones were distinguished in core Tu 2 (Figure 6). Zone Tu 2-moll-1 (8360–7810 cal yr BP; 380–320 cm) is characterised by a high diversity and abundance of aquatic mollusc species, including mainly A. leucostoma, G. truncatula, B. contortus, P. casertanum, V. cristata and hygrophilous S. putris. In zone Tu 2-moll-2 (7810–7250 cal yr BP; 320–235 cm), aquatic species are dominated by G. truncatula, and among hygrophilous species, E. alderi occurs. In zone Tu 2-moll-3 (7250–6640 cal yr BP; 235–162 cm), the sparse mollusc assemblage is composed of hygrophilous (C. minimum and E. alderi) and land (V. pulchella and N. hammonis) snails. Within zone Tu 2-moll-4 (6640–5470 cal yr BP; 162–74 cm), hygrophilous and land species gradually decline.

Local mollusc succession in Turtul spring-fed fen, core Tu 2, presented along with sediment lithology. Mollusc assemblages divided into aquatic, hygrophilous and land species. Taxa presented as shell counts (with X-axis scale labels; note scale differences).
Tufa carbon and oxygen stable isotope values
Within the peat-tufa intercalations in the Tu 1 sediment sequence, δ13C values vary between −11.6 and −6.79‰, and a gradual enrichment in 13C occurs (Figure 7). The δ18O values change between −10.39 and −8.33‰. Strong fluctuations between the samples characterise both isotope records. The correlation between C and O stable isotope records, which can inform about the rate of water exchange resulting from the intensity of the water supply at the fen, is positive and strong (r = 0.66) between ca. 9200 and 8440 cal BP but changes to weak and negative (r = −0.24) between 8840 and 7700 cal yr BP with an increased CaCO3 content in the sediments. A strong negative correlation (r = −0.54) is observed between ca. 7700 and 5510 cal yr BP during the major phase of tufa precipitation in Turtul.
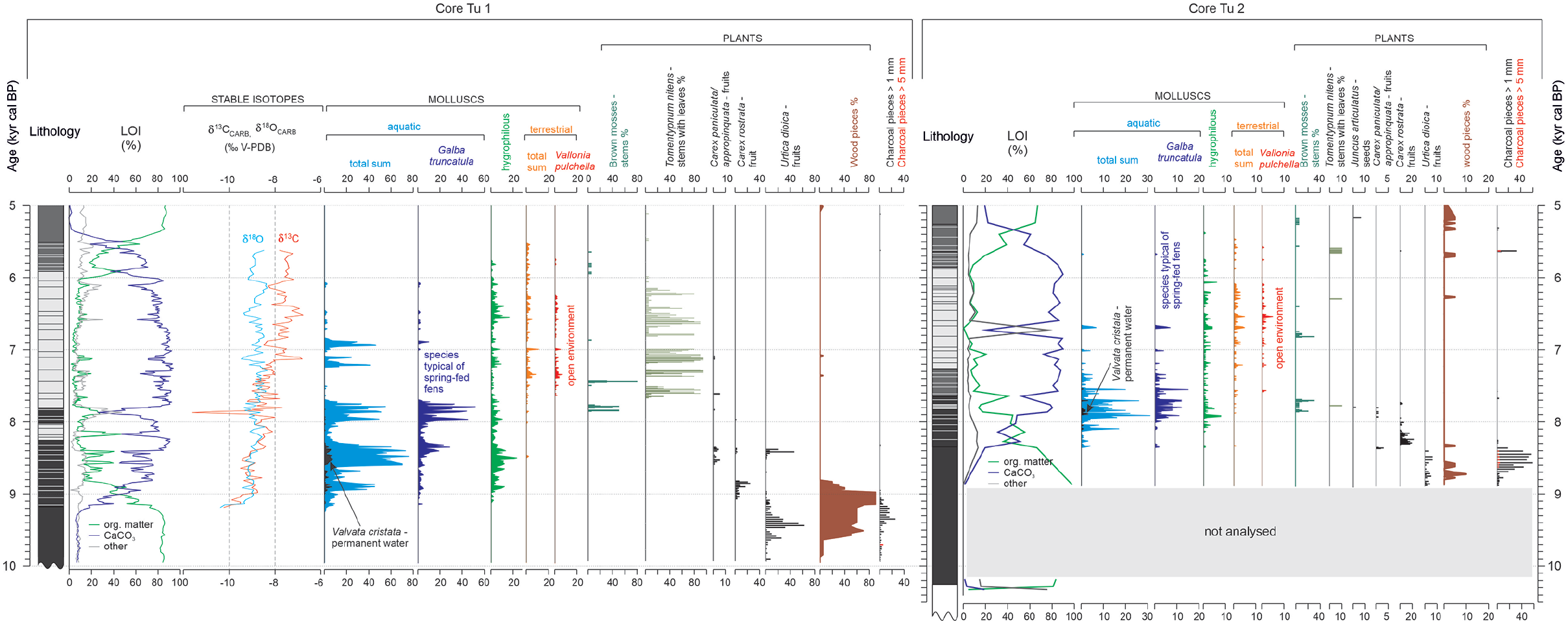
Compilation of selected data of the Tu 1 and Tu 2 sediment sequences: Lithology, LOI, C and O stable isotopes, molluscs and plant macrofossils.
Discussion
Spatial variation in the sediment records of Turtul fen
The Tu 1 and Tu 2 sediment sequences reveal congruent palaeoenvironmental histories, common trends in the geochemical records, and a similar succession of plants, mosses and molluscs (Figures 3–7). However, a detailed inspection of the records reveals differences in the timing of deposition between the two cores, including the start and termination of carbonate deposition (Figure 7). The observed shifts are partially covered by the confidence intervals (2sigma) of the ages derived from the models, which vary between 165 and 810 years in Tu 1 and from 110 to 518 years in Tu 2 (Figure 2).
The apparent differences between the two cores can be attributed to the well-recognised internal fen variability (cf. Gałka et al., 2017, 2018). The delayed start of CaCO3 deposition and concurrent occurrence of molluscs in the Tu 2 sediment sequence (8350 cal yr BP) compared to the Tu 1 core (9250 cal yr BP) (Figure 7) was probably due to the different locations of the two coring sites. Tu 1 was drilled in the highest point of the fen where water outflow was located and where CaCO3 sedimentation started first. As the water circulation was gradually unblocked, as recorded by the intensification of CaCO3 precipitation at ca. 8350 cal yr BP in Tu 1 (Figure 7), the spatial reach of the Ca2+-enriched spring water was more extensive and allowed for tufa precipitation and the occurrence of aquatic and hygrophilous mollusc species. The different locations of the two cores are also reflected in the subsequent temporal shifts in sediment geochemistry and biotic proxies. However, the general pattern of changes is very similar in Tu 1 and Tu 2. The discontinuous but frequent occurrence of Tomentypnum nites in Tu 1 and only sparse occurrence of this moss in Tu 2 (Figure 7) can indicate the difference in the substrate between the two spots. According to the habitat preferences of T. nitens (Nicholson and Gignac, 1995), the top of the fen must have been less swampy compared to its slopes. A similar change in the substrate is apparent today. Additionally, the poor preservation of the moss remains in the Tu 2 core prevent their accurate taxonomic classification.
The discrepancy in the termination of CaCO3 deposition in the two cores, ca. 5500 versus 5300 cal yr BP, in Tu 1 and Tu 2 (Figure 7), respectively, may also be related to changes in the chemical composition of the water flowing on the fen. When water emerging at the fen flows down on the fen surface, it loses CO2 through outgassing and absorption by autotrophs. As a consequence, the water pH increases, and CaCO3 can precipitate at some distance from the source (Capezzuoli et al., 2014). Therefore, it is likely that the location from which the Tu 2 sediment core was collected favoured the temporary persistence of carbonate deposition, even after it stopped at the centre of the fen location. The importance of the above mechanism for CaCO3 precipitation is evidenced by the mean sediment accumulation rate values (Figure 2). In core Tu 2, the highest SAR values of 0.17–0.25 cm yr−1 were observed between ca. 7680 and 7910 cal yr BP (295–339 cm) and were noticeably higher than those observed between ca. 7100 and 7870 cal yr BP (189–284 cm) in the Tu 1 core: 0.11–0.14 cm yr−1. Considering the above discussion, careful selection of the coring site is crucial in terms of accurate palaeoenvironmental research of spring-fed fens. In studies where one sediment sequence is analysed, the coring should be done in the central and in commonly the highest, part of the fen close to the outflow where water is not subjected to geochemical modifications observed along the increasing distance from the source.
The early and middle Holocene ecosystem changes in the Turtul spring-fed fen are a response to regional and local climate changes. Given the central location of the Tu 1 core in the spring-fed fen in Turtul, high-resolution LOI analysis, and C and O stable isotope analysis performed exclusively for this core, the climatic reconstruction is based mainly on the data from the Tu 1 sediment sequence supplemented with the data from the Tu 2 core.
Start of peat deposition (480–368 cm; 10,300–9250 cal yr BP)
Deposition of peat in Turtul started at ca. 10,300 cal yr BP (Tu 2) and continued until 9250 cal yr BP (Tu 1), when CaCO3 precipitation was initiated (Figure 7, Table 2). The highly decomposed peat underlying the peat-tufa intercalations typically occurs at the bottom of the spring-fed fen sediments and records accumulation associated with an active layer of permafrost that thawed seasonally during the progressive warming of the early Holocene (Dobrowolski et al., 2019). The deposition of peat in Turtul started as a response to a major environmental transformation in the region that was forced by the final decay of the Scandinavian Ice Sheet and change in the atmospheric circulation in the Baltic region (Lauterbach et al., 2011). The replacement of cold and dry air from the northeast by warm and moist westerlies accelerated the spread of deciduous trees in the region (Gałka et al., 2015), with pioneer Betula-Pinus forests noted since ca. 11,250 cal yr BP (Gałka et al., 2014).
The lithology of Tu 1 and Tu 2 sediment sequences.
The open environment at Turtul that was dominated by herbs (Apiaceae taxa) persisted until ca. 9600 cal yr BP (380 cm) when trees entered the fen, as indicated by the increase in the wood pieces (Figure 3). The site became more fertile and favoured the spread of Urtica dioica. The substantial share of charcoal pieces (>1 mm), which increased as trees entered the fen, indicates drier conditions at the site and/or its surroundings, favouring local fire events. This is supported by the presence of fungal remains (sclerotia) and large charcoal pieces (>5 mm), the share of which was the highest after 9400 cal yr BP (Figure 8) when the cold and dry Erdalen Event, Bond event 6 (Bond et al., 2001), occurred. The increase in the intensity of fire events in the early Holocene was recorded at many sites in Central and Eastern Europe (Feurdean et al., 2020).
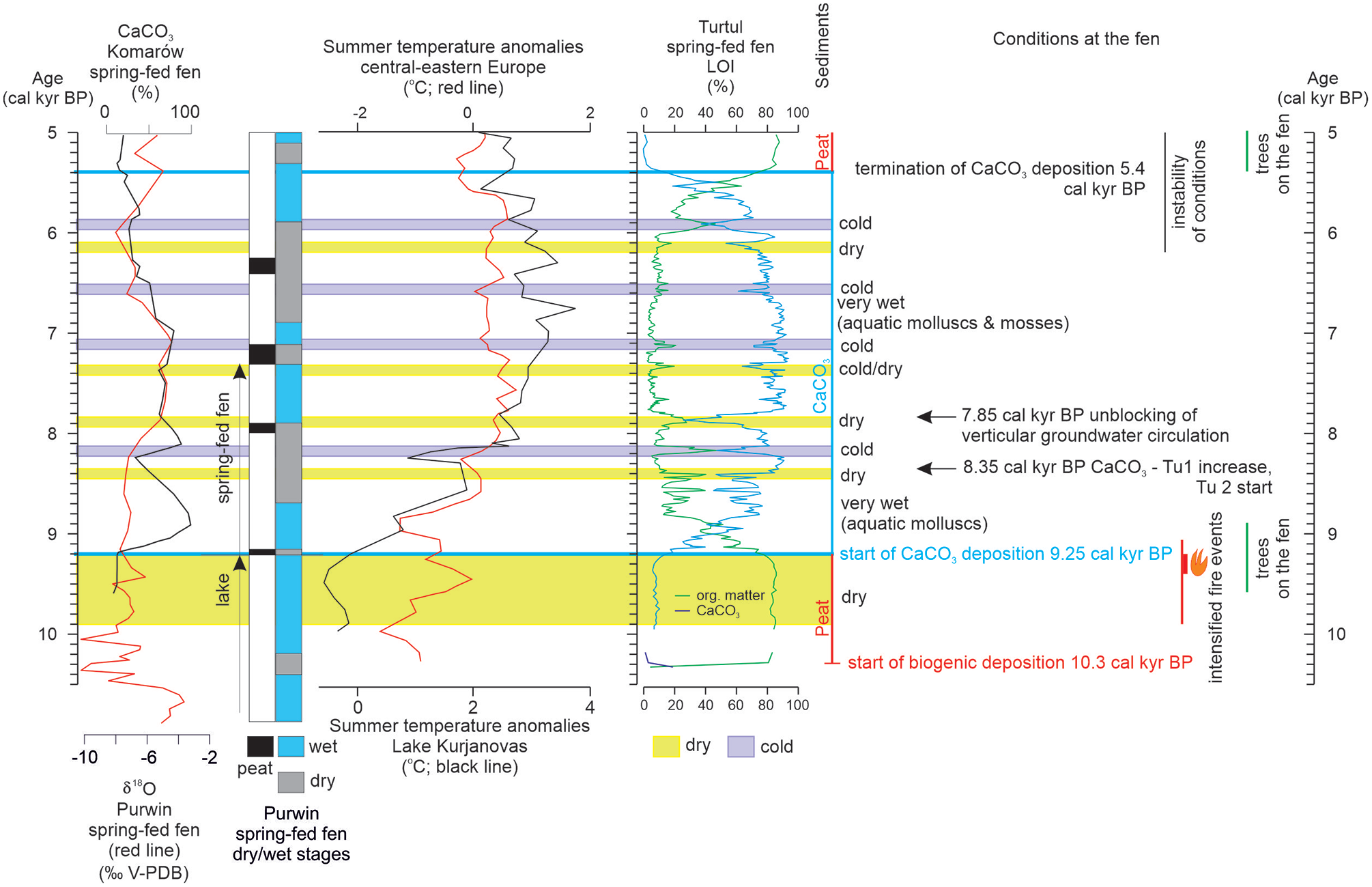
The major stages of the Turtul spring-fed fen development and regional climatic changes presented against CaCO3 and organic matter contents in Tu 1 and Tu 2 (only between ca. 10,300 and 10,200 cal yr BP) cores, compared with the local record of Purwin spring-fed fen (Apolinarska and Gałka, 2017), the regional record of Komarów spring-fed fen (Dobrowolski et al., 2016), pollen-based summer temperature anomalies in Lake Kurjanovas (Heikkilä and Seppä, 2010) and central-eastern Europe (Davis et al., 2003).
Carbonate spring fed-fen (368–79 cm, 9250–5540 cal yr BP)
The first increase in the CaCO3 content in the Tu 1 sediments at ca. 9250 cal yr BP (368 cm) indicates the start of enhanced groundwater circulation, as suggested by Dobrowolski et al. (2019) for northern Poland. Based on the highly variable CaCO3 content (Tu 1; Figure 7), it is suggested that at this stage, only shallow water circulation was active in the area and was highly susceptible to even small shifts in the water supply in the spring. The negative δ13C and δ18O values of the carbonates (Figure 7), which are typical of the authigenic CaCO3 precipitated at the spring-fed fens (Andrews et al., 1997; Gałka et al., 2021), exclude the detrital origin of CaCO3, the supply of which was documented in the Purwin spring-fed fen (Apolinarska and Gałka, 2017) located 10 km northeast of the Turtul site. Sedimentary and palaeobotanical records from Kojle-Perty Lake (located in the vicinity of Purwin Lake) (Gałka et al., 2015) and the Purwin spring-fed fen (Apolinarska and Gałka, 2017) indicate an abrupt shift to a wet habitat at ca. 9400 cal yr BP and 9200 cal yr BP (Figure 8), respectively. The nature of the change to humid conditions was regional, as the timing of the onset of CaCO3 deposition at Turtul was concurrent with observations at Komarów (Figure 1) and other cupola spring-fed fens (Dobrowolski et al., 2002, 2005, 2016) located on uplands in eastern Poland. In Lake Jaczno (Figure 1), 9200 cal yr BP marks a change in the sedimentation type and a shift to higher productivity in the lake (Makri et al., 2021). The observed environmental changes were related to prominent warming in the Baltic Sea basin (Borzenkova et al., 2015; Heikkilä and Seppä, 2010) and central-eastern Europe (Davis et al., 2003) (Figure 8). In the Turtul spring-fed fen, the combined increase in humidity and warming enhanced CaCO3 precipitation (Figure 7) and stimulated the development of a diverse and abundant mollusc assemblage (Figure 5).
Very wet conditions at the Turtul fen between 9250 and 8450 cal yr BP (368–320 cm) are indicated by the highest share of aquatic mollusc species (Figure 5), characteristic mainly of small and shallow but permanent water basins (Anisus leucostoma, Valvata cristata, Anisus vortex, Radix labiata and Pisidium casertanum). Similarly, plant macrofossils indicate a gradual increase in the water level (Figure 3). The remains of Alnus glutinosa, U. dioica and charcoals (>1 mm) were gradually replaced with Carex rostrata, which is the plant that usually occurs in a very wet habitat such as lake shores and hollows in peatlands (Ellenberg et al., 1991; Zarzycki et al., 2002). The presence of the water pools at the fen is confirmed by a strong positive correlation (r = 0.66) between δ13C and δ18O values, indicating the influence of the residence time effects, including evaporation, on the C and O stable isotope records (Andrews, 2006). The share of wood pieces gradually decreased; however, the presence and occurrence of macroremains of A. glutinosa, Betula sect. albae and Pinus sylvestris indicate that trees grew on the fen surface.
Short drying (8450–8340 cal yr BP; 320–314 cm) within the generally wet stage is recorded as a decrease in CaCO3 content and a decline or retreat of most of the aquatic mollusc species (Figures 5 and 7). However, small water ponds must have occurred at the site, as indicated by the dominance of Galba truncatula and the presence of Bathyopmphalus contortus (Figure 5). The fen became overgrown by U. dioica, C. rostrata, C. paniculata, Stellaria palustris and Caltha palustris (Figure 3).
Between 8340 and 7850 cal yr BP (314–281 cm), the conditions at the fen remained wet and favoured CaCO3 precipitation. The supply of the fen with groundwater was intensified as the correlation between δ13C and δ18O records changed to weakly negative (r = -0.24). A high (up to 91%) CaCO3 content (Figure 7) can be linked with gradual regional warming (Borzenkova et al., 2015; Davis et al., 2003; Heikkilä and Seppä, 2010). The strongly increased CaCO3 content in Tu 1 was concurrent with the onset of tufa deposition in the Tu 2 core (Figure 7). Small water ponds at the fen are indicated by the mollusc assemblage composed of G. truncatula accompanied by Carychium minimum, Euconulus alderi and Succinea putris. A single shell of Vertigo moulinsiana, a species characteristic of alkaline fens, is also present. The wet stage between 8400 and 7700 cal yr BP is also observed in the Bobolice cupola spring-fed fen (Dobrowolski et al., 2019) (Figure 1).
The widespread 8200 cal yr BP cooling event in Turtul is recorded as a distinct CaCO3 decrease (from ca. 70% to 80% to a minimum of 47% at 8170 cal yr BP at 304 cm) between 8200 and 8130 cal yr BP (302–305 cm), a negative shift on the generally increasing δ18O curve (Figure 7) and strongly decreasing shares of plant macrofossils (Figure 3) and mollusc shells (Figures 5 and 7). The 8200 event is well recognised in central-eastern Europe (Figure 8; Davis et al., 2003), including the Baltic region, for example, Lake Rouge in Estonia (Veski et al., 2004); Lake Flarken in southern Sweden (Seppä et al., 2005); Lakes Eini and Malmuta in eastern Latvia; Lake Arapisto in Finland (Sarmaja-Korjonen and Seppä, 2007); and in Poland (Szeroczyńska and Zawisza, 2011). Among the central European cupola spring fed-fens, it was recorded in Komarów (Dobrowolski et al., 2016) and Bobolice and Ogartowo (Dobrowolski et al., 2019) (Figure 1).
The next short, from 7900 to 7850 cal yr BP (286–281 cm), but strong decrease in the CaCO3 content had its maximum at ca. 7870 cal yr BP (Figure 7) and marks a regional climatic change. In the Purwin spring-fed fen, a thin peat layer was dated to 7900 cal yr BP (Apolinarska and Gałka, 2017). Regionally, it coincides with dry and/or cold conditions in the Ogartowo and Spurgle spring-fed fens in northern Poland (Dobrowolski et al., 2019) and cold spells in Lake Kurjanovas, eastern Latvia (Heikkilä and Seppä, 2010) and Lake Laihalampi, south-central Finland (Heikkilä and Seppä, 2003) (Figure 1). In the Turtul spring-fed fen, the share of fruits and fruit scales of Betula sect. albae and Betula pubescens increase markedly (Figure 3). Trees must have occurred close to the fen; however, they most likely did not grow over its surface, as no wood remains were found. The abundance of molluscs declined except for hygrophilous E. alderi (Figure 5), a snail species inhabiting humid sites with leaf litter (Welter-Schultes, 2012).
A significant increase in CaCO3 at 7850 cal yr BP (Figure 7) likely records an onset of deeper water circulation in Turtul, concurrent with the unblocking of vertical groundwater circulation in the Spurgle spring-fed fen (Dobrowolski et al., 2019) (Figure 1). Increased recharge is confirmed by a shift to a more negative correlation between C and O stable isotope values (r = −0.45). The regional increase in humidity was observed from 8000 to 7900 cal yr BP in higher lake levels in lakes Kojle-Perty (Gałka, 2014) and Purwin (Gałka and Apolinarska, 2014) and in the Purwin spring-fed fen (Apolinarska and Gałka, 2017) (Figure 1). Since ca. 8000 cal yr BP, a shift to high lake levels was noted in northern Poland (Michczyńska et al., 2013). Increased humidity is recorded in frequent event (flood) layers from 8500 to 7000 cal yr BP in laminated sediments in Lake Jaczno (Makri et al., 2021) and Lake Szurpiły (Kinder et al., 2020). Repeated episodes of enhanced erosion, attributed to the increased runoff, were also recorded in Lake Żabińskie between 8000 and 7500 cal yr BP (Zander et al., 2021).
The culminating developmental stage of Turtul calcareous fen functioning occurred from 7850 to 6700 cal yr BP, when the sediment accumulation rate was the highest (Figure 2). The coring site was overgrown with Tomentypnum nitens and brown mosses (Figures 3, 4 and 7), which are species occurring at sites with CaCO3-rich substrate (Ochyra, 1992). Seeds of Cladium mariscus, a plant indicative of a warm climate with summer temperatures exceeding 15.5°C and preferring sites with alkaline substrates at the eastern limits of its distribution in Europe (cf. Gałka and Tobolski, 2012; Gałka et al., 2021 and references therein), occurred at ca. 7000 cal yr BP (Figure 3). Along with T. nitens, the mollusc assemblage became dominated by hygrophilous species C. minimum and E. fulvus, which typically occurred at fens, and land species Nesovitrea hammonis (Figure 5). The site must have remained open, that is, there was no canopy of trees, as Vallonia pulchella, land snail species characteristic of open environments gradually became more frequent (Figure 5).
The most intensive CaCO3 deposition in Tu (7850–6700 cal yr BP) is consistent with the maximum number of sites with tufa deposition observed across Europe between 7500 and 7000 cal yr BP (Dabkowski, 2020). This matches the regional change to a warm and humid climate. High water levels were observed in Lake Purwin (8000–7900 to 7100–6900 cal yr BP; Gałka and Apolinarska, 2014), lakes Kojle and Perty (Gałka, 2014), and other lakes in northern Poland (Michczyńska et al., 2013). Wet and warm conditions indicated by a high CaCO3 content are known from cupola spring-fed fens in Komarów (8100–6800 cal yr BP; Dobrowolski et al., 2016), Bobolice (7700–6400 cal yr BP; Dobrowolski et al., 2019) and Ogartowo (7700–6000 cal yr BP; Dobrowolski et al., 2019). In both Latvia and southern Finland, the warmest climate was recorded between 8200 and 5700 cal yr BP, with a peak from 7950–6750 cal yr BP (Heikkilä and Seppä, 2003, 2010). Of importance for CaCO3 precipitation must have been the intensive leaching of CaCO3 material (Dabkowski, 2020) dispersed in the sediments deposited by the Scandinavian ice sheet. It was triggered by the extensive development of the forests in the area (Gałka et al., 2014), which enhanced soil respiration and the release of CO2 and led to intensified CaCO3 dissolution in the background sediments.
Approximately 50 years (7350–7300 cal yr BP, 220–213 cm) of cooling manifested in decreased CaCO3 contents and a concurrent shift to lower δ18O values (Figure 7). A concurrent decrease in pollen-inferred temperature was observed in south-central Sweden (Seppä et al., 2005). In Turtul, the environment became drier and more open, as indicated by the absence of aquatic and hygrophilous mollusc species and the increased share of V. pulchella (Figures 5 and 7), which dominated the mollusc assemblage from ca. 7390 to 7230 cal yr BP (223–205 cm). Abundant T. nitens also points to a dryer habitat (Figure 7). Locally, this environmental shift was recorded as decreased water levels in Lake Purwin (7300 cal yr BP; Gałka and Apolinarska, 2014) and a dry stage in the Purwin spring-fed fen record (7300–7100 cal yr BP; Apolinarska and Gałka, 2017). Regionally, a short (ca. 7350 cal yr BP) decrease in CaCO3 contents in the sediments occurred in the Komarów cupola spring-fed fen (Dobrowolski et al., 2016).
Another cold spell during ca. 7150–7050 cal yr BP is manifested by a coinciding decrease in CaCO3 contents and a drop in δ18O values (Figures 7 and 8), contemporaneous with decreased temperatures in central-eastern Europe (Davis et al., 2003). At the same time, abundant T. nitens (Figure 7), preferring a drier, albeit open and calcium-rich habitat; the presence of Carex paninculata; and increased tufa δ13C values (12C that was preferentially assimilated by autotrophs affected the δ13C value of the ambient dissolved inorganic carbon to a greater extent) point to a decreased water level.
Following the short cooling event, very intensive CaCO3 deposition continued until ca. 6700 cal yr BP (158 cm). Although the composition of the mollusc assemblage remained unchanged, the hygrophilous snails C. minimum and E. alderi became more frequent (Figure 5). A negative correlation between the presence of the aquatic snail Stagnicola palustris (Figure 5), a species occurring mostly in permanent waters, swamps and desiccating reservoirs, and T. nitens moss (Figure 3) indicates temporal occurrence and disappearance of the shallow water pool at the coring site.
Gradual environmental change at the fen began ca. 6700 cal yr BP (158 cm) with a shift to lower CaCO3 contents (from an average of 88% to an average of 78%) in the sediments (Figure 7). T. nitens, although still abundant, was less numerous than before (Figure 3). Small water pools must have disappeared from the fen surface, at least at the coring site, since aquatic mollusc species retreated, and the fen was inhabited by hygrophilous and land snails (Figures 5 and 7). The change observed in Turtul is concurrent with a shift to a dry stage at the Purwin spring-fed fen (Apolinarska and Gałka, 2017) and fits shallow-water stands in lakes and peat formation in northern Poland during ca. 6800–6500 cal yr BP (Michczyńska et al., 2013). Żurek et al. (2002) describe the period between 6700 and 6000 cal yr BP as the driest on the mires of eastern Poland. This finding is confirmed by the decreasing trend in CaCO3 precipitation and the retreat of Phragmites australis and thus drier conditions in the Komarów spring-fed fen, which began ca. 6800 cal yr BP and continued until 5300 cal yr BP, with a distinct drying at ca. 6750 cal yr BP (Dobrowolski et al., 2016). At the Turtul spring-fed fen, the change was relatively moderate as the decrease in CaCO3 deposition was lower, but to a limited extent. It is possible that the influence of the dry continental climate was greater in eastern Poland than at sites located proximal to the Baltic Sea.
Cooling between 6600 and 6500 cal yr BP at Tu 1 is recorded in decreased CaCO3 contents in the sediments and lower δ18O values (Figure 7), which matches the reduced summer temperatures in central-eastern Europe (Davis et al., 2003). The environment became more open, as indicated by the increase in V. pulchella.
The final stage of the carbonate fen in Turtul, between ca. 6200 and 5400 cal yr BP, was characterised by the significant instability of the conditions reflected in the overall decrease but simultaneously highly fluctuating CaCO3 contents in the sediments (Figure 7). In central-eastern Europe, the temperatures gradually decreased at that time (Davis et al., 2003; Heikkilä and Seppä, 2003).
The strong decrease in the CaCO3 content in the sediments during ca. 6160–6100 cal yr BP (115–119 cm) was simultaneous with a major change in vegetation, that is, finish of the abundant occurrences of T. nitens (Figure 7), which can be regarded as the termination of the most alkaline stage of the fen history. Changes in sedimentological and ecological conditions at the fen must have resulted from drying, which enhanced the evaporative loss of 16O and increased δ18O values. Decreased δ13C values indicate a smaller influence of autotrophs on the ambient DIC, which was probably related to the abovementioned retreat of T. nitens. The subsequent abrupt but short-duration increase in the CaCO3 content in Turtul, with the highest content at ca. 6050 cal yr BP (Figure 7), indicates an improvement in temperature-humidity conditions. This was concurrent with a similar change observed in the Komarów spring-fed fen between 6200 and 6000 cal yr BP.
A major decrease in CaCO3 accumulation (Figure 7) from ca. 5980 to 5870 cal yr BP (102–107 cm) records a regional climatic change. This matches the local cooling recorded in the sediments of Lake Purwin at 5900 cal BP (Gałka and Apolinarska, 2014). Regionally, low lake levels in Poland persisted from 6000 to 5900 cal yr BP (Michczyńska et al., 2013). Major cooling and a decrease in humidity were observed at ca. 5900 cal yr BP in the Komarów spring-fed fen (Dobrowolski et al., 2016), and since ca. 6000 cal yr BP, CaCO3 has decreased in the Orgatowo spring-fed fen (Dobrowolski et al., 2019). Cooling and drying responsible for the observed changes match Bond event 4, which culminated at ca. 5900 cal yr BP (Bond et al., 2001).
The final stage of the calcareous spring-fed fen functioning in Turtul (5870–5600 cal yr BP), which was recorded in increased CaCO3 contents and macrofossils of brown mosses (Figure 7), coincides with increased summer temperatures across central-eastern Europe (Davis et al., 2003) (Figure 8). The concurrent increase in humidity was recorded in Lake Purwin and the Purwin spring-fed fen since ca. 5850 cal yr BP (Apolinarska and Gałka, 2017; Gałka and Apolinarska, 2014) and on the peatland located next to Lake Perty after 5900 cal yr BP (Gałka, 2014). Enhanced tufa precipitation was observed in the Komarów spring-fed fen (Dobrowolski et al., 2019).
Termination of the carbonate spring-fed fen functioning (79 cm, 5540 cal yr BP)
Termination of the Turtul carbonate spring-fed fen functioning was dated to ca. 5500–5400 cal yr BP when the CaCO3 content decreased abruptly (Figure 7). After 5400 cal yr BP (79 cm), only a trace share (usually below 1% and in all samples below 5%) of CaCO3 was measured.
The mechanisms underlying cessation of tufa precipitation can vary (Goudie et al., 1993) but are broadly related to climatic or anthropogenic causes (Dabkowski, 2020). In the study area, the first traces of human influences on the natural ecosystems involve burning of woodlands during 7650–6150 cal yr BP (Madeja et al., 2004), which preceded the decline in the Tu calcareous fen and the development of the pastoral economy dated to 6150 and 5300 cal yr BP (Gałka et al., 2014). Both resulted in the clearing of the land and a subsequent increase in nutrient supply from soils. The increased nutrient supply had a negative impact on calcareous fen functioning, which is warranted by the low nutrient levels (Grootjans et al., 2006). A natural transformation could have involved the overgrowth of the fen surface by trees, as indicated by wood fragments occurring in the deposits since ca. 5300 cal yr BP and plant macrofossils present in the sediments during the transition from the peat-tufa intercalations to the overlying peat, that is, Betula sect. albae fruit scales and single Alnus glutinosa fruit (Figures 3 and 4). Forestation of the fen would have led to increased shadowing and eventual cooling of the fen surface. As temperature is among the most critical variables controlling CaCO3 deposition (Grootjans et al., 2006), its role in the cessation of tufa precipitation in the Turtul fen was most likely vital. In addition, the termination of CaCO3 deposition ca. 5500–5400 cal yr BP is concurrent with the culmination of the long-term cooling trend observed in the eastern Baltic area since ca. 6300 cal yr BP (Heikkilä and Seppä, 2010) and the major cooling in east-central Europe ca. 5500–5200 cal yr BP (Davis et al., 2003) (Figure 8).
The termination of CaCO3 deposition was likely not caused by decreased effective precipitation because the distinct cooling since ca. 5600 cal yr BP was associated with a shift to high water levels in northern Poland (Michczyńska et al., 2013). Similarly, since ca. 5500 cal yr BP, increasing water levels promoted the spread of plant communities demanding a higher water table in the Gązwa bog (Gałka and Lamentowicz, 2014). A short decrease (ca. 5300–5200 cal yr BP) in the water level in Lake Purwin (Gałka and Apolinarska, 2014) and the Purwin spring-fed fen (Apolinarska and Gałka, 2017) were observed, but this change was not recorded in the nearby Lakes Kojle and Perty (Gałka et al., 2014). Moreover, we regard the eventual disability of the ascending water from reaching the surface at cupola spring-fed fens (Grootjans et al., 2015) being a consequence of increasing thickness of accumulated peat and tufa deposits, as less likely at the hanging fens supplied with descending groundwater from the local near-surface aquifer located in the slopes of the river valley cutting Quaternary unconsolidated deposits. At present, small water pools occur continuously at the Turtul fen surface (2-year personal observation); therefore, water shortage was probably less likely to cause the cessation of CaCO3 deposition. However, there could also have been a gradual diminishing of Ca2+ concentration in the groundwater as the carbonate rock detritus in the sediments deposited by the Scandinavian Ice Sheet was progressively dissolved by groundwater during the early and mid-Holocene. At present, the concentration of Ca2+ in the groundwater emerging at the Turtul fen (mean 63 mg l−1; Apolinarska et al. unpublished) is in the lower limit of values measured in extremely rich fens from the Carpathian Flysch Zone, where CaCO3 precipitation was observed, and vegetation characteristic of extremely rich fens occur (Hájek et al., 2002). However, in a majority of the fens located in European lowlands with similar Ca2+ concentrations, CaCO3 is not precipitated (Grootjans et al., 2015).
The termination of tufa deposition in different sedimentary environments across Europe during the Holocene was not uniform and could have been caused by various factors, as reviewed by Dabkowski (2020). In contrast to the Turtul spring-fed fen sediment record, at numerous sites, CaCO3 precipitation initiated in the early or mid-Holocene and continued in the upper Holocene: Komarów (Dobrowolski et al., 2016), Ogartowo, Spurgle (Dobrowolski et al., 2019), and a cupola spring-fed fen in Romincka Forest (Apolinarska et al., unpublished data), which are all located in northern and eastern Poland. Therefore, changes in the climatic conditions at ca. 5500 cal yr BP, which might have been of crucial importance at the Turtul fen, did not stop CaCO3 precipitation at other locations. In the Komarów spring-fed fen, a change to the lowest CaCO3 content in the sediment sequence began at 5400 and lasted only until ca. 4800 cal yr BP (Dobrowolski et al., 2016), whereas in Bobolice, a short cold and dry episode was observed from ca. 5650 to 5600 (Dobrowolski et al., 2019). Finally, there is no consensus on the Holocene phase when temperatures were the highest and therefore promoted the precipitation of carbonates in terrestrial environments. The long-agreed maximum air temperatures across central Europe between 8000 and 5500 cal yr BP (Davis et al., 2003) and globally between 8000 and 4000 cal yr BP for northern latitudes between 30° and 60° based on various proxies (Kaufman et al., 2020) are challenged by reinterpreted pollen data, which indicate a warm climate between 5500 and 2000 cal yr BP (Marsicek et al., 2018). The declining temperatures two millennia ago coincide with the second stage of tufa decline in Europe (Dabkowski, 2020).
Conclusions
A combination of a high sampling resolution and high sediment accumulation rate provided the opportunity to assess Turtul’s palaeoenvironment in detail, thereby providing a perfect opportunity to complement the still infrequent high-resolution records of the early and mid-Holocene and verify the accuracy of the well-preserved and minutely subsampled hanging spring-fed fen sediments and their record of the short-duration local and regional climate shifts of the early and mid-Holocene in the southeastern Baltic region.
(1) In this paper, so far, the most detailed record from spring-fed fens, recording climatic changes in the southeastern Baltic region and ecological history of the site from the early and mid-Holocene (10,300–5400 cal yr BP) is presented. A high-resolution reconstruction was achieved by the combination of detailed sampling (1-cm sampling interval) and a high sediment accumulation rate at the fen. The peat-tufa intercalations accumulated with the highest rates, 0.11–0.25 cm yr−1, between ca. 7900 and 7100 cal yr BP.
(2) The genesis of the Turtul spring-fed fen is soligenous and related to an outflow of spring waters on the slope of the Czarna Hańcza River valley with no lacustrine phase present. Major stages of fen development include (i) the start of biogenic deposition at 10,300 cal yr BP, (ii) the start of CaCO3 precipitation at 9250 cal yr BP, (iii) the main stage of fen functioning with the most intensive tufa accumulation between 7850 and 6700 cal yr BP and (iv) the termination of tufa precipitation at 5400 cal yr BP.
(3) Analyses of the two cores from the fen revealed a very similar sequence of ecological and geochemical conditions but with an apparent temporal shift associated with the distance of the coring site from the water outflow. The spatiotemporal differences between Tu 1 and Tu 2 records stress the importance of an accurate selection of the coring site for palaeoenvironmental studies.
(4) The ecosystem of the Turtul fen was susceptible to short shifts to cold and/or dry spells revealed by previous studies from the southern Baltic region. The climatic shifts at Turtul were recorded mainly in the decreased CaCO3 content and in changes in biological and isotopic proxies, which occurred from 8200–8130, 7900–7850, 7400–7300, 6600–6500, 5980–5870 and at 5400 cal yr BP. Dry stages with no associated cooling were also recorded from 10,200–9250, 8450–8340 and 6160–6100 cal yr BP. This, and the instant reaction of the fen, mainly by changes in CaCO3 precipitation, to hydrological and temperature changes indicates the sensitivity of the well-preserved and continuous record of hanging spring-fed fens in palaeoclimatic studies.
(5) The reason for the termination of tufa deposition at Turtul at 5450 cal yr BP is not clear. However, based on the available data, we suggest that cooling and/or the immigration of trees at the fen surface were the two most important drivers.
Footnotes
Acknowledgements
We greatly appreciate the work of two anonymous Reviewers who carefully revised our manuscript.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The work was financed by Polish National Science Centre grant No. 2018/29/B/ST10/00120.