
Abstract
This study examines the archeological seed assemblage of Anle, a middle Neolithic site located in the Lower Yangtze region, China. The Lower Yangtze is thought to be the origin of domesticated rice and most studies of this region to date have focused on rice domestication and cultivation within its paleoenvironmental setting. In contrast, we highlight here diverse uses of non-rice plant resources. In addition to large quantities of rice remains (carbonized grains and spikelet bases), we identify both foxtail and broomcorn millet, both AMS radiocarbon dated earlier than 5750 cal BP, demonstrating the dispersal of millet cultivation to the Lower Yangtze in the middle Neolithic, earlier than previously securely documented. While most wild species identified in macrobotanical assemblages are traditionally categorized as weeds or incidental intrusions among food residues, many can be exploited for food and medicinal purposes. By analyzing the ecological and functional implications of identified plants, we infer ecological niches of cultivation, gathering, and possible propagation of wild plants as food and medicine. Analyses of diversity and seasonality of plant resources identified show that residents of Anle created a complex seasonal sequence of temporally compatible crops, constructing niches for two crops (rice and millet) and actively structuring opportunities to exploit available wild plant resources in their immediate environment.
Introduction
The past two decades have witnessed a rapid development in paleoethnobotanical studies of the Lower Yangtze Delta in eastern China. A growing body of literature on paleoenvironment and crop domestication contributes many insights into the broad picture of past human societies in the Lower Yangtze Delta, especially during the Neolithic period. There is a growing consensus that rice (Oryza sativa) cultivation originated in the Lower Yangtze Delta at the beginning of Holocene, approximately 9000 cal BP (Liu et al., 2007; Zheng et al., 2016; Zuo et al., 2017). Paleoenvironmental studies also reveal that the Lower Yangtze Delta underwent various periods of environmental change throughout the Holocene that have contributed to the rise and fall of local Neolithic communities (Qin et al., 2011; Yu et al., 2000). While these new studies provide valuable insights into the process of rice domestication, as well as its interaction with paleoenvironmental dynamics in the Lower Yangtze, only a few case studies to date have been concerned with directly addressing subsistence or the diversity of plants used from either a macrobotanical or microfossil perspective (Qiu et al., 2016; Yao et al., 2016; Zheng et al., 2019). This lack of attention creates problems in reconstructing agricultural or subsistence practices because it is unclear to what extent non-rice plants were exploited and thus we have little insight into how overall agricultural regimes that included both rice and non-rice plants functioned during this critical period of early agriculture.
This study expands botanical studies of the Neolithic Lower Yangtze through macrobotanical analysis of Anle, a middle Neolithic site in the Lower Yangtze. This study shows that residents of Anle constructed niches for not only rice, but also for broomcorn and foxtail millets, which have been rarely identified in the macrobotanical record of sites in this region to date. We verify the early date of these millet remains with direct AMS radiocarbon dating. In addition, our data indicate that local residents, to a significant extent, managed their lives in accordance with the seasonality of exploited plants in order to take advantage of diverse botanical resources, both cultivated and wild, over an annual cycle. This study demonstrates how analysis of ecological and functional implications of a full suite of plant taxa, both domesticated and wild, can highlight how people incorporate plants with distinct ecological and nutritional characteristics to create a single functioning subsistence strategy.
Paleoenvironment and archeological background of Anle
Anle is a site located in northern Zhejiang province, along the lower reaches of the Yangtze River. The Lower Yangtze region is largely a floodplain encompassed by the Yangtze River in the north, low-elevation mountains in the west and the south, and the East China Sea to the east. Environmental features of the present day are quite different from those of the Holocene. The Lower Yangtze in the early Holocene was a flat marine terrace sitting on the deeply-incised paleo-valley and was subject to rapid sea level rise until 8000 cal BP (Wang et al., 2018; Zong et al., 2012b). Sea level rise, however, slowed down between 8000 and 7000 cal BP, followed by a period of warm and moist climate at around 7000–5500 cal BP (Tao et al., 2006; Zong et al., 2012b). By c. 7000 cal BP, the Lower Yangtze was still an open coastal area with shallow water. Due to the remarkedly reduced extent of sea level rise, sandy sediments with shells from previous marine transgressions accumulated, gradually forming Chenier ridges along the southern bank of the Yangtze River and the northern shore of Hangzhou Bay (Zong et al., 2011). These Chenier ridges functioned as a topographical barrier that not only transformed the Lower Yangtze into an enclosed wetland, but also contributed to the formation of the Tai Lake, once a coastal lagoon, as a freshwater marsh by c. 6000 cal BP (Tao et al., 2006). Nevertheless, inundations of brackish water still occurred at intervals throughout the Holocene, maintaining periods of low-level salinity in the Lower Yangtze region, and multiple Neolithic sites in the region show spatial and temporal variation in these hydrological successions (Chen et al., 2018; Qin et al., 2011; Zong et al., 2012a).
It is under these environmental changes that the people of Anle lived. The site is situated on the eastern edge of the Dipen Basin, southwest of the Tai Lake. Surrounding the site are low elevation hills that average 15 m high (Figure 1). The Zhejiang Institute of Cultural Relics and Archaeology (hereafter, the Zhejiang Institute) conducted four salvage excavations at Anle in 1996, 2001, 2013, and 2014 that yielded abundant lithics, pottery, jade, and wooden objects. The 2013 and 2014 excavations opened up an area of 4000 m2 in total. Eighty-eight burials and multiple mudbrick features, potentially associated with housing structures, were exposed, implying that Anle is a site composed of multiple independent settlement areas (Shen et al., 2014). Due to the salvage nature of the four excavations, neither AMS dating nor extensive analysis has been performed to render any deeper understanding of this site. Nevertheless, in their initial reports the excavators dated Anle from the late Majiabang period to the Songze period based on pottery typology (Shen et al., 2014). Conventional dates for the Majiabang and Songze period are c. 7500–5900 BP and c. 5900–5200 BP respectively (Itzstein-Davey et al., 2007). The Majiabang people and the Songze people were undoubtably rice consumers as both morphological study of rice grains and phytolith analysis suggest that rice cultivation intensified during this period (Fuller et al., 2007; Ma et al., 2016). On the other hand, however, it has been proposed that the persistent low salinity of the Lower Yangtze region may have posed threats to rice cultivation and thus limited the expansion of Majiabang and Songze people, whose settlements are frequently found in areas east of the Tai Lake (Zong et al., 2011, 2012a; Figure 1). From that perspective, Anle is unique because it is situated in a basin protected by hills whereas other Majiabang sites were normally positioned in the floodplains (Jiang, 2012).
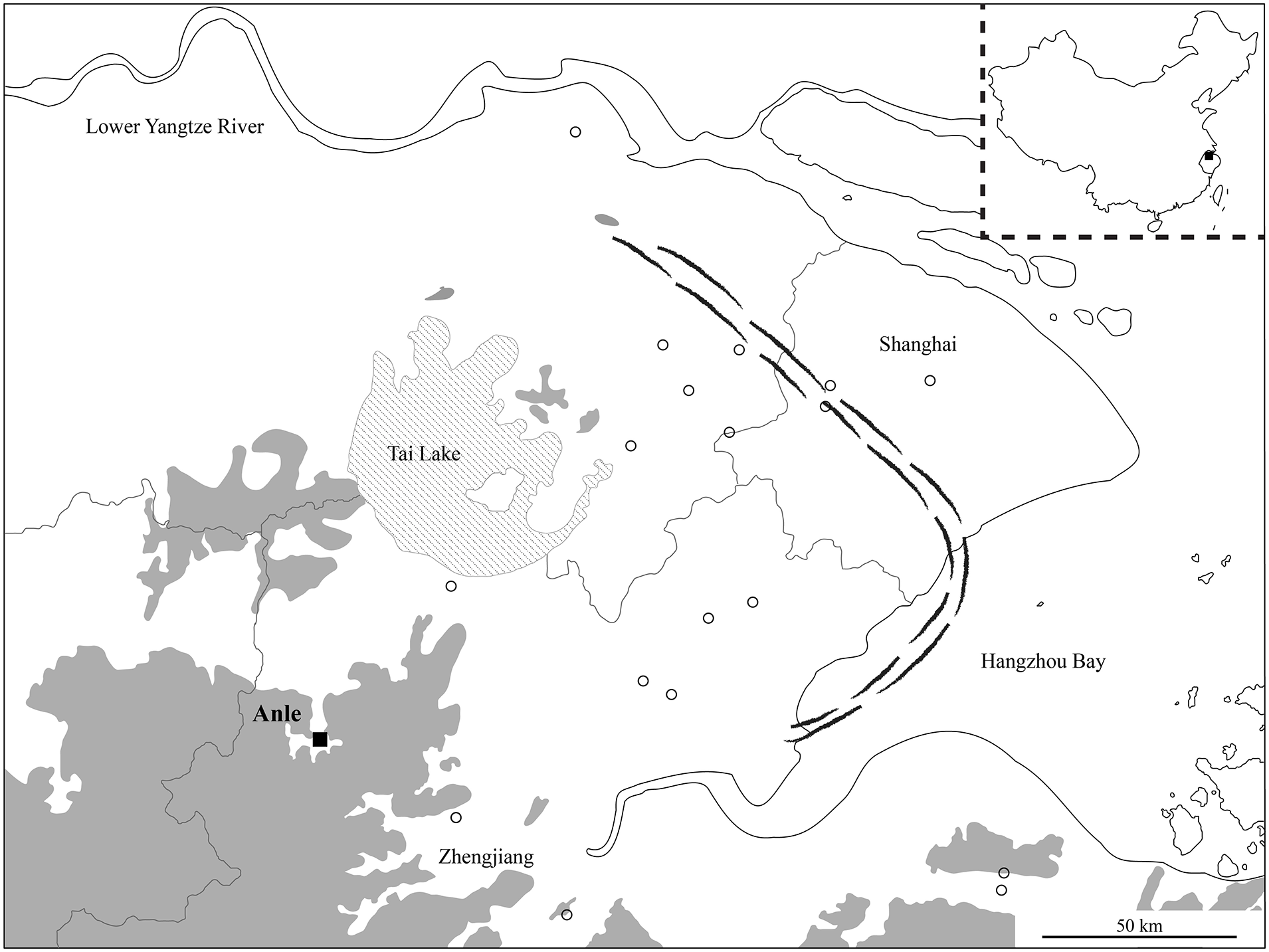
The location of Anle in the Lower Yangtze region. Square: Anle; hollow circles: distribution of major Neolithic sites in this region; dashed line: Chenier ridges in 6000 BP; gray shading: mountains and hills. Figure redrawn after Zong et al. (2012a).
Materials and methods
The soil samples analyzed in this study were taken in 2014 during the final excavation season. Given the salvage nature of the excavation, extensive sampling of the site was not conducted, but samples for potential future botanical analysis were taken from one particularly notable deposit. A total of 24 soil samples were taken from the top and bottom of Stratum 7 (S7) of the excavation unit TN5E2 (30.655335°N, 119.695390°E) (Figure 2). S7 was a refuse or a trash dump, rich in pottery sherds, wood materials, and animal bones (Shen et al., 2014). The excavators did not date the site or the individual strata using any direct dating method, but dated S7 to the Majiabang-Songze transitional period based on pottery from the layer. Ma et al. (2016), without specifying the dating method or the materials used, date S7 of TN5E2 to 6000–5700 cal BP. Since this date fits into the broad chronology of the site, we considered 6000–5700 BP a convincing date for this stratum, although we sought to confirm this date through AMS radiocarbon dating of individual millet remains, as detailed in the results.

Southern wall of TN5E2. S1: surface soil; S2: Shang and Zhou dynasties (c. 3500–2250 BP); S3 and S4: Songze (c. 5900–5200) layers in which mudbrick features and burials are found; S5 and S6: early Majiabang (c. 7500–5900) to Songze period, soil likely redeposited from other places; S7: dark deposit of early Majiabang to Songze period, containing rich deposits of pottery sherds, animal bones, and wooden materials; S8: natural deposits of sterile silt.
Flotation of these soil samples took place in winter 2019 using a modified wash-over technique (White and Shelton, 2014), recovering macrobotanical remains with a 0.45 mm mesh. Due to the high clay content of the soil, 250 ml of baking soda (sodium bicarbonate) was added to each sample during flotation as a flocculant (Pearsall, 2015). When some macrobotanical material did not float during the first flotation attempt, water was added again and the sample was refloated until the water became clean and no visible charcoal material remained in the sample bucket. Only botanical material from light fractions was collected, as there was no visible botanical material in the heavy fraction. Nevertheless, large pottery sherds were handpicked from the heavy fraction and stored with the light fractions for future study.
Sorting and identification took place in the laboratory of the Zhejiang Institute. A standard stereomicroscope was used in the sorting process. Since this laboratory does not use geological sieves in the sorting process, seeds and charcoal of different sizes were sorted together and charcoal was not sorted from the sample residue in order to prioritize seed collection and identification. The residue from sorted light fractions, which includes the charcoal, has been kept in separate bags with tags to allow future charcoal analysis. A modern seed collection of local plants and several reference publications available in the lab (Ishikawa, 1995; Liu et al., 2008; Nakayama et al., 2000) were used to identify seeds. We follow the taxonomy of the Flora of China (Wu et al., 2013) for all botanical names; notably, this means that we treat Amaranthaceae and Chenopodiaceae as separate families. During analysis, relative frequency was calculated based on the count of whole seeds. Relative frequency excluding rice (Oryza sativa) caryopses and spikelet bases is also calculated in order to understand the relative contribution of non-rice taxa compared to one another.
Accelerator mass spectrometer radiocarbon dating was conducted individually on four millet caryopses at the National Ocean Sciences Accelerator Mass Spectrometry (NOSAMS) facility at the Woods Hole Oceanographic Institution. Due to the small mass of these samples, an acid-base-acid pretreatment was applied to only one caryopsis (the Panicum miliaceum caryopsis from the top of S7); the other three were combusted without pre-treatment to avoid additional mass loss and improve precision. Laboratory standard organic carbon small-sample protocols were used for sample preparation and combustion. Samples were run on the NOSAMS Continuous-Flow AMS System and reported as standard radiocarbon dates, corrected for isotopic fractionation using unreported d13C values measured on the accelerator, then calibrated by the authors using OxCal 4.4.4 (Bronk Ramsey, 2009) and the IntCal20 calibration curve (Reimer et al., 2020).
Results
Across the 24 samples, a total of 48.5 L of sediments were floated, yielding 3.04 L of light fractions. The total count of identified complete seeds and plant parts is 15,719, including spikelet bases from rice, which number 12,719 (Table 1). Among the 2853 seeds that are not rice, at least 33 distinct taxa and 21 families were identified, indicating a rich and diverse composition of macrobotanical remains. The overall results show that plants that naturally grow in wetlands were common. Table 1 summarizes the macrobotanical results as well as potential uses and typical habitats for identified taxa.
Summarized macrobotanical results.
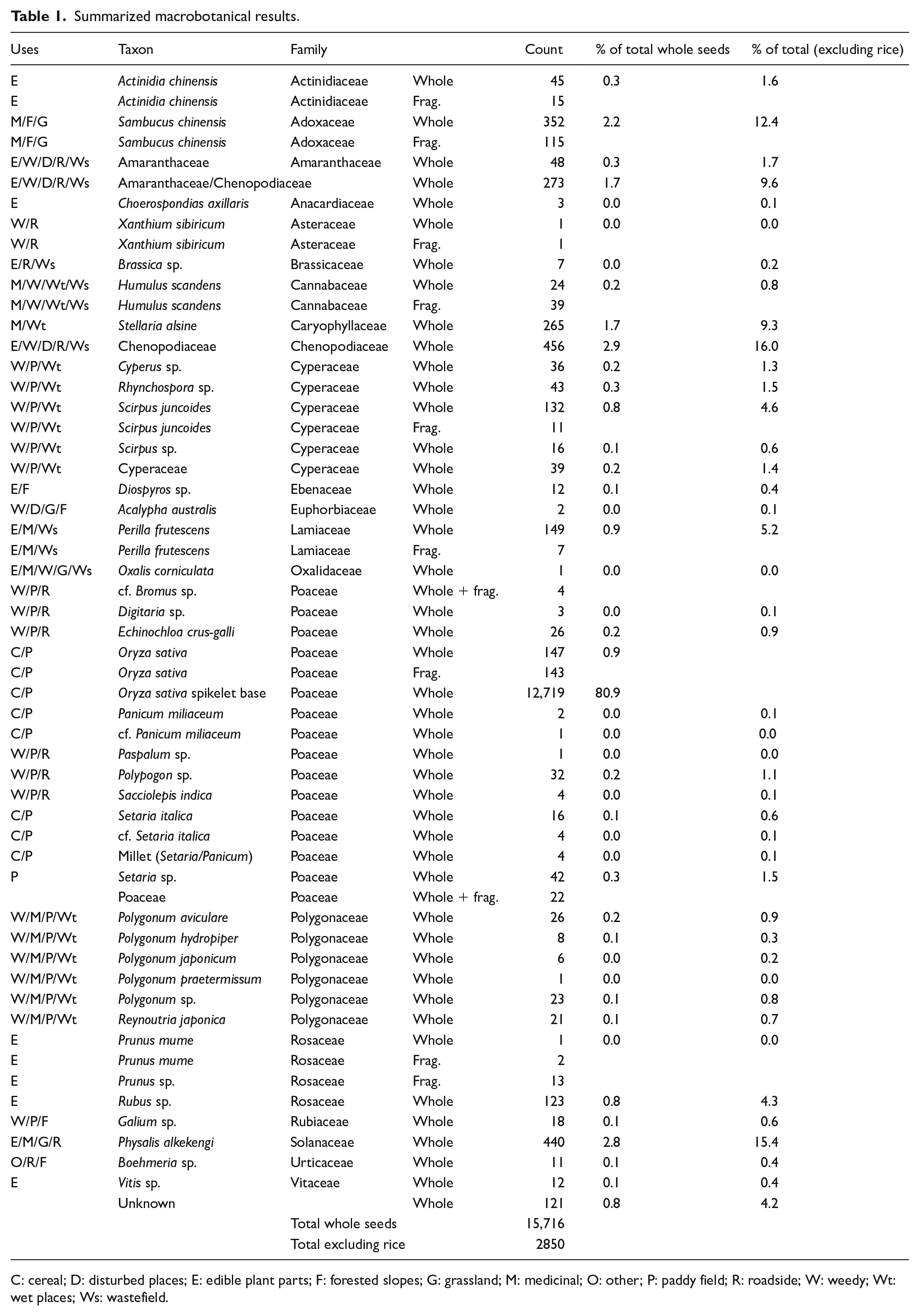
C: cereal; D: disturbed places; E: edible plant parts; F: forested slopes; G: grassland; M: medicinal; O: other; P: paddy field; R: roadside; W: weedy; Wt: wet places; Ws: wastefield.
Radiocarbon dating
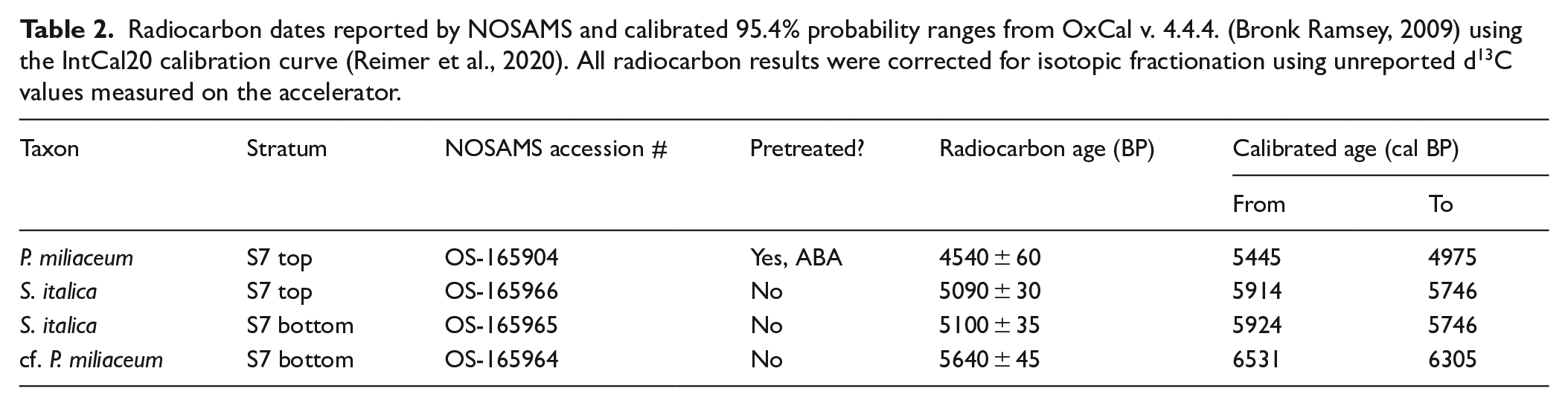
Four millet caryopses (two Setaria italica, one certain, and one probable Panicum miliaceum) were AMS radiocarbon dated. Results are presented in Table 2 and Figure 3; the caryopses analyzed are illustrated in Figure 4.
Radiocarbon dates reported by NOSAMS and calibrated 95.4% probability ranges from OxCal v. 4.4.4. (Bronk Ramsey, 2009) using the IntCal20 calibration curve (Reimer et al., 2020). All radiocarbon results were corrected for isotopic fractionation using unreported d13C values measured on the accelerator.
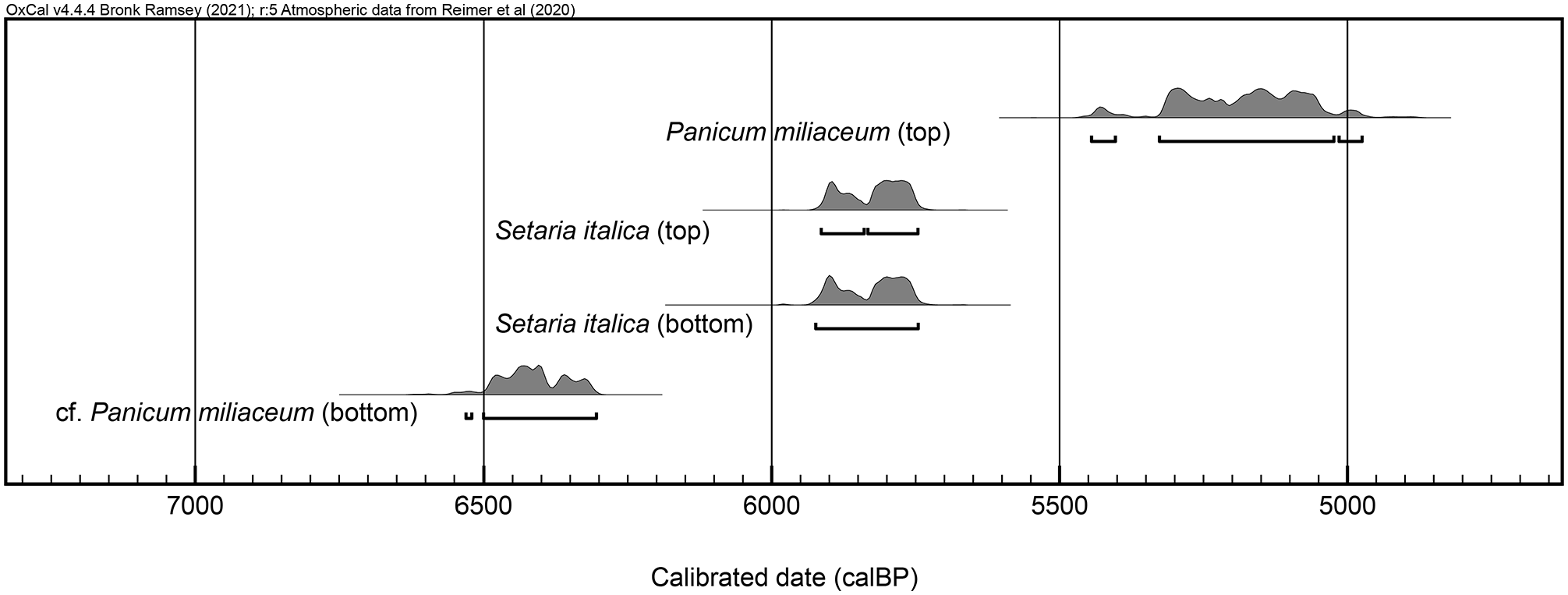
Calibrated radiocarbon probability distributions for millet caryopses dated. “Top” refers to seeds from the upper portion of S7; “bottom” refers to seeds from the lower portion of S7. Calibration performed in OxCal v. 4.4.4. (Bronk Ramsey, 2009) using the IntCal20 calibration curve (Reimer et al., 2020).

Millet caryopses radiocarbon dated. Ventral, lateral, and dorsal (left to right) views are given for each except b, for which only ventral and dorsal images are available. (a and c) come from samples near the top of S7 and (b and d) from samples near the bottom of S7.
While it is possible that some residual material was incorporated into the deposit and that some later material was mixed into the top of the deposit, we note the similar ages provided by the two foxtail millet (Setaria italica) caryopses from the top and bottom of S7, and the chronostratigraphic continuity of dates identified as supporting a late Majiabang (conventionally, 7500–5900 BP) to Songze (conventionally, 5900–5200 BP) date for S7. All dates fall into the late Majiabang, early Songze, or late Songze; the two foxtail millets (c. 5920–5750 cal BP) closely match the original late Majiabang to early Songze date range of 6000–5700 BP attributed to this stratum by Ma et al. (2016).
Crops: Rice and millets
We expected that rice (Oryza sativa) would be the predominant species at Anle, as the Lower Yangtze is recognized to be one of the locations of origin of rice domestication (Crawford, 2016). Spikelet bases from rice make up 81% of the total count of identified remains in these samples. It has been argued that rice domestication accelerated during the transition between the Majiabang and Songze period, the period to which multiple Neolithic paddy fields found in this region date (Fuller et al., 2007; Qiu et al., 2014) and to which our samples date. Rice was clearly the main staple cultivated and consumed by the residents of Anle. It is no surprise to find weedy species that are commonly found in paddy fields, such as barnyard grass (Echinochloa crus-galli), other weedy grasses (Setaria sp., Digitaria sp., Sacciolepis indica, and Polypogon sp.), and sedges (Scirpus spp. including S. juncoides, Cyperus sp., and Rhynchospora sp.). Barnyard grass is thought to be the wild progenitor of Japanese millet (E. esculenta), which was domesticated in Japan before 4000 BP (Crawford, 2011). Evidence of both rice and E. crus-galli phytoliths was found on lithic processing tools from an early Neolithic site in the Lower Yangtze, leading the researchers to conclude that wild E. crus-galli was a food source at around 10,000 BP (Yang et al., 2015). Thus it is possible that barnyard grass was also consumed to some extent in Anle, although our data do not indicate whether it was cultivated.
A surprising find is the significant presence of additional crops, including both foxtail millet (Seteria italica) and, to an apparently lesser extent, common millet (Panicum miliaceum), although counts are relatively low (together, all millets comprise 2.5% of non-rice remains). The recognized region of origin of millet domestication is in northern and central China, where early Neolithic sites (around 10,000 BP) yield carbonized foxtail and common millet grains (Diao and Jia, 2017; Lu et al., 2009; Zhao, 2019). Recent paleoethnobotanical studies show that a dualistic regime of rice and millet started to emerge in the Middle Yangtze c. 5800 cal BP and sites south to the Lower Yangtze around 4000 cal BP (Deng et al., 2018; Nasu et al., 2007, 2012). The number of millet grains found in the Lower Yangtze to date remains extremely low. Zhao and Jiang (2016) reported foxtail millet grains found in a Neolithic layer of the site of Shangshan, which was dated c. 6500 BP. This “earliest” presence of millet in the Lower Yangtze is, however, questionable due to the lack of direct dating (Deng et al., 2018). Two additional sites in the Lower Yangtze reported merely one millet grain found at each site, dated to 4500–4300 BP and 2770–2255 BP (He et al., 2017). Therefore, the discovery of more than 20 definitely domesticated millet grains at Anle, with both species AMS radiocarbon dated earlier than 5700 BP, opens up a new discourse on the dispersal of millet from central China to the Lower Yangtze during the Middle Neolithic. The appearance of millet might also explain the occurrence of cf. Bromus sp., a grass that normally grows in temperate regions and mountains in the tropics (Wu et al., 2013). Today farmers still grow millets in small quantities in hilly areas in northern China, where the climate is colder and more arid (Song et al., 2013). Considering the growth requirement of millets, it is likely that residents of Anle utilized the low elevation hills adjacent to their settlement for millet fields. Weeds such as Bromus sp. could have been collected when harvesting millets and later removed and deposited as waste during crop processing (An et al., 2022; Harvey and Fuller, 2005; Reddy, 1997)
Weeds, edible, and medicinal plants
Only a few species outside of the grasses and sedges discussed above can be regarded as weeds. The first set of weedy species includes those in the Polygonaceae family (Polygonum aviculare, P. hydropiper, P. praetermissum, P. japonicum, and Reynoutria japonica). Though some sources suggest that most of them (excluding P. praetermissum) are used in medicine (Guo, 2009; Wu et al., 2013), the fact that they grow in wet places introduces threats to paddy field management as they may compete with rice for water and other resources. Among the identified species in the Polygonaceae, P. hydropiper perhaps has the highest requirements for water availability and nutrient conditions in porous soil, and is tolerant of low soil temperature (Sultan et al., 1998). Traditional paddy-field agricultural practices including plowing before seeding, maintaining a shallow water level, and manuring, create an ideal growing environment for P. hydropiper, even at times outside the rice growth season (i.e. in winter to early spring). The presence of P. hydropiper also suggests that the pH level of paddy fields was slightly acidic, as it normally grows in soils with a pH level close to 6.0 (Sultan et al., 1998). Paddy fields in southern China were commonly mildly acidic by the 1980s, suggesting that traditionally cultivated paddy fields were a suitable habitat for P. hydropiper (Guo et al., 2018). It is likely that these Polygonaceae weeds, especially P. hydropiper, were intentionally removed alongside other weedy plants during paddy field management. Galium arguably could have been used for medicinal purposes, as in Native American traditions (Moerman, 1986). However, there is a lack of compelling Chinese ethnographic evidence that plants of this genus were used in medicine. In fact, some Galium spp. are toxic to pigs (Wu et al., 2013). Though pig remains are frequently found in Majiabang and Songze period sites, they did not contribute significantly to the diet until 5500 BP (Dong and Yuan, 2020; Pan and Yuan, 2018). It is thus unlikely that removing Galium spp. was a specific strategy used in pig husbandry, but more likely a general land-preparation technique used in managing farmlands. Another common weed is Xanthium sibiricum, which grows in lowlands and roadsides. The hooked projections on its bur makes it easily dispersed by animals or humans, possibly the reason why it entered the archeological record. There are three other weedy species, Humulus scandens, Oxalis corniculata, and Acalypha australis, that grow in wastelands and streamsides. Though they arguably can be used in medicine or, in the case of Oxalis, as occasional foodstuffs, their relatively frequency is less than 1%. Thus, it is hard to conclude that residents of Anle intentionally collected them for medicinal or other purposes.
Weedy species that have edible plant parts, in contrast, make significant contributions to the macrobotanical assemblage. Chenopodiaceae have the highest relative frequency, which is likely due to their high seed yield per plant (Stevens, 1932). Amaranthaceae, the seeds of which are sometimes indistinguishable from those of the Chenopodiaceae, also contributes meaningfully to the total count. Species in these two families are strongly weedy and usually grow in waste places and fields created through anthropogenic efforts (Spengler, 2019; Wu et al., 2013). As waste places and fields are likely located close to farmlands or settlement areas, their successful weedy character might motivate people to utilize these nearby resources for their edible leaves and medicinal stems. Several species in these families were tolerated weeds or even propagated deliberately for their greens in other parts of the world, especially North America (Sullivan and Forste, 2014).
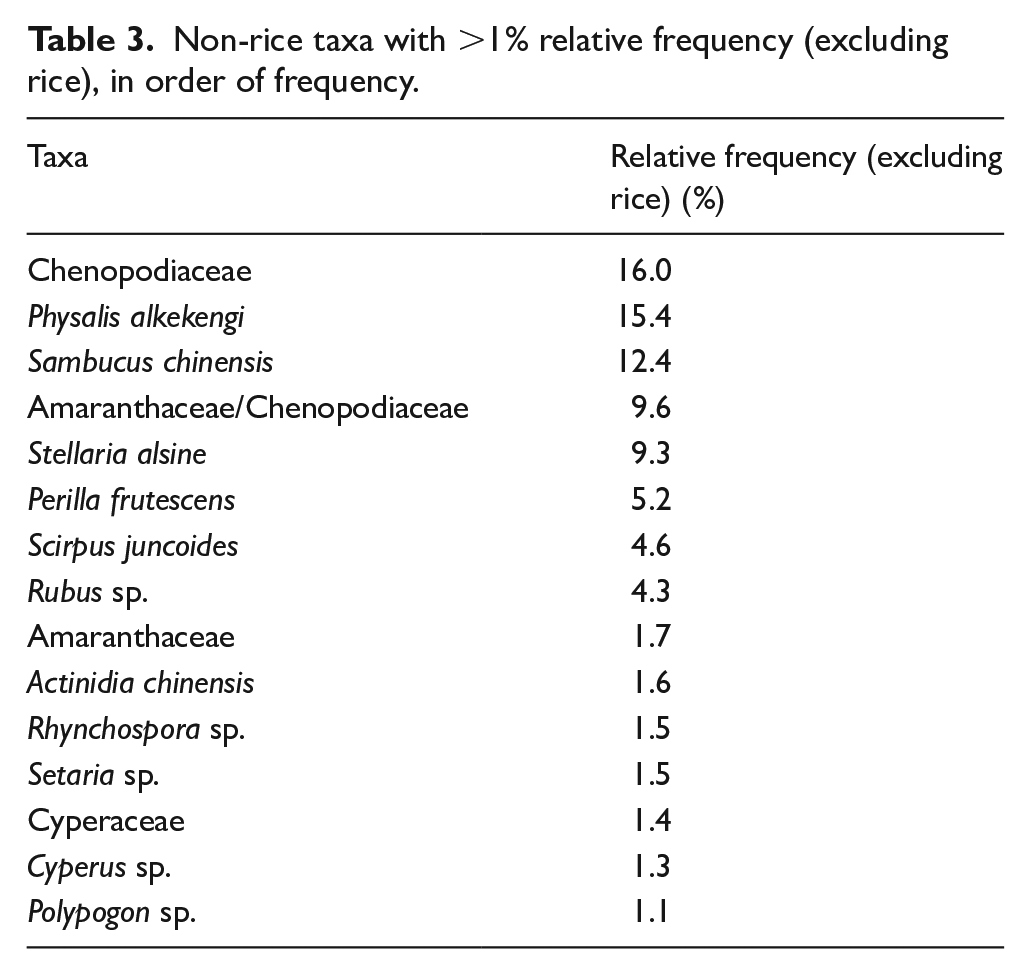
Other plants that grow close to settlement areas include Sambucus chinensis (elderberry), Stellaria alsine (chickweed), and Perilla frutescens (shiso). Whereas S. chinensis and S. alsine most often grow in natural habitats (grasslands and wet places), P. frutescens grows in waste areas and cultivated gardens. Though the origin of P. frutescens cultivation is still unclear, P. frutescens is frequently used in medicinal and culinary traditions in East Asia. Being a multipurpose plant, P. frutescens produces edible leaves while its seeds are used in flavoring and oil production (Nitta et al., 2003). Similar to P. frutescens, S. chinensis and S. alsine are traditional medicine for tending wounds (Guo, 2009). Though there is a lack of strong evidence to suggest that local residents were aware of the medicinal potential of these plants and thus cultivated or collected them, the high relative frequency of the three species makes it clear that the local residents had already started to utilize these plants for one or more purposes. Whether this early phase of utilizing herbs marks some of the first archeological evidence for traditional medicine is a possibility open to future debate. As these five taxa (Amaranthaceae, Chenopodiaceae, P. frutescens, S. chinensis, and S. alsine) are five of the six most common non-rice taxa in the assemblage and together account for 56% of the total non-rice seed count, these results imply a moderate level of exploitation of these weedy but useful taxa (Table 3).
Non-rice taxa with >1% relative frequency (excluding rice), in order of frequency.
Fruits and potential cultigens
Some of the species identified are fruit-producing plants (Physalis alkekengi, Actinidia chinensis, Rubus sp., Vitis sp., Prunus mume, and Choerospondias axillaris) and potential domesticates (Brassica sp., Diospyros sp., and Boehmeria sp). P. alkekengi (ground cherry or Chinese lantern) is used in treating coughs in traditional medicine and it is widely cultivated today for its distinctly flavored fruits (Wen et al., 2019), The high relative frequency of P. alkekengi (second only to Chenopodiaceae) suggests that residents of Anle actively consumed its fruits, perhaps contributing to its later cultivation. Other fruits disposed of at Anle come from plants that grow at higher elevations (forested slopes and hillsides), and so P. alkekengi may have been routinely harvested alongside other plants in those habitats. Actinidia chinensis (kiwi fruit), Rubus (bramble), and Vitis (grape) are commonly found in Neolithic sites alongside the Yangtze River, though the exact process of their cultivation is still debated (Liu et al., 2008). Modern domesticated kiwi fruits grow in sparse secondary forests. However, Actinidia spp. today are recognized as wild fruits by many communities in China (Liu et al., 2019). The genus Rubus is well known for its great diversity and wide range of distribution. There are nearly 400 species in China, with the majority of them found south of the Yangtze River (Lu, 1983; Wu et al., 2013). Given the current poor understanding of historical Rubus use, it is hard to identify any specific local practice, although such fruits have a long tradition of consumption in Asia and worldwide; the seeds, dried fruits, and leaves may have been used medicinally (Wu et al., 2013). Similarly, Vitis is well recognized and was frequently recorded by the ancient Chinese. However, its cultivation is considered to have begun only much later when V. vinifera was introduced, possibly due to the poor taste of Chinese wild species (Jiang et al., 2009).
Besides these fruits, the presence of Prunus mume (Chinese plum) and Choerospondias axillaris (lepsi) suggests a certain degree of human modification of primary forests. Prunus mume and its close relative Prunus persica (peach) grow in sparse forest and along streams, with peach having been domesticated as early as 7500 BP (Zheng et al., 2014), though strong evidence for a broader system of arboriculture in South China only arises in the sixth millennium BP (Fuller and Stevens, 2019). Recent studies show that the fast-growing Choerospondias spp. are highly adaptive in terms of topography and temperature, thus making Choerospondias an ideal choice in constructing secondary broad-leaved forests (He et al., 2004; Wang et al., 2020). Its sour-tasting, fleshy mesocarps were consumed, as its endocarps are found in many Neolithic sites in the Lower Yangtze (Pan et al., 2013). If residents of Anle managed plum trees for fruit in the late spring or early summer, then some clearance of original forests might have been required. In that case, Choerospondias is an ideal substitute as its large number of deciduous leaves increases soil fertility while it also provides edible fruit (He et al., 2004). Most Brassica spp. (mustards) have edible leaves while Diospyros spp. yield persimmon fruits. Both become domesticated later in history (Liu et al., 2008). Boehmeria might have been collected by residents of Anle for its fiber. The domesticated Boehmeria nivea yields ramie, a high-quality fiber. However, the low count of seeds of these three plants in the assemblage is insufficient to make claims about their intentional cultivation or even deliberate gathering, rather only their potential for use and eventual cultivation.
Discussion
Seasonality
The macrobotanical results demonstrate that the residents of Anle made use of a diverse spectrum of food sources, with rice being the predominant crop. Studying the seasonality of plants and fruits identified at Anle provides insights into how its residents managed their labor over an annual cycle (Figure 5). September and October are often the busiest time of the year for food collection, as many fruit-bearing plants reach maturity or are in the middle of their fruit-producing season. Herbs such as S. chinensis are still available until September. Physalis alkekengi and Perilla frutescens might have been collected with ease alongside other regular activities as they grow in disturbed places, whereas collection of A. chinensis and other tree crops, as well as some herbs, are not found in anthropogenic habitats (streamside, hillside, and even mountain slopes). Foxtail and common millets today are sowed in spring and harvested in summer or sowed in summer and harvested in autumn, depending on temperature and precipitation in different regions (An et al., 2022; Song et al., 2013). In either way, millet harvesting, and perhaps processing as well, is finished by late September or early October. Keeping this schedule is essential to the success of a double-crop regime with rice, as rice is harvested in late October to early November. The International Rice Research Institute (IRRI) estimates that 25 person-days/ha is needed for cutting and gathering rice, 15 person-days/ha for threshing, and about 8 person-days/ha for cleaning paddy fields (IRRI Rice Knowledge Bank, 2021). Though it is hard to estimate the exact scale of rice production at Anle, a communal effort was surely necessary given the labor intensiveness of rice harvesting and processing.
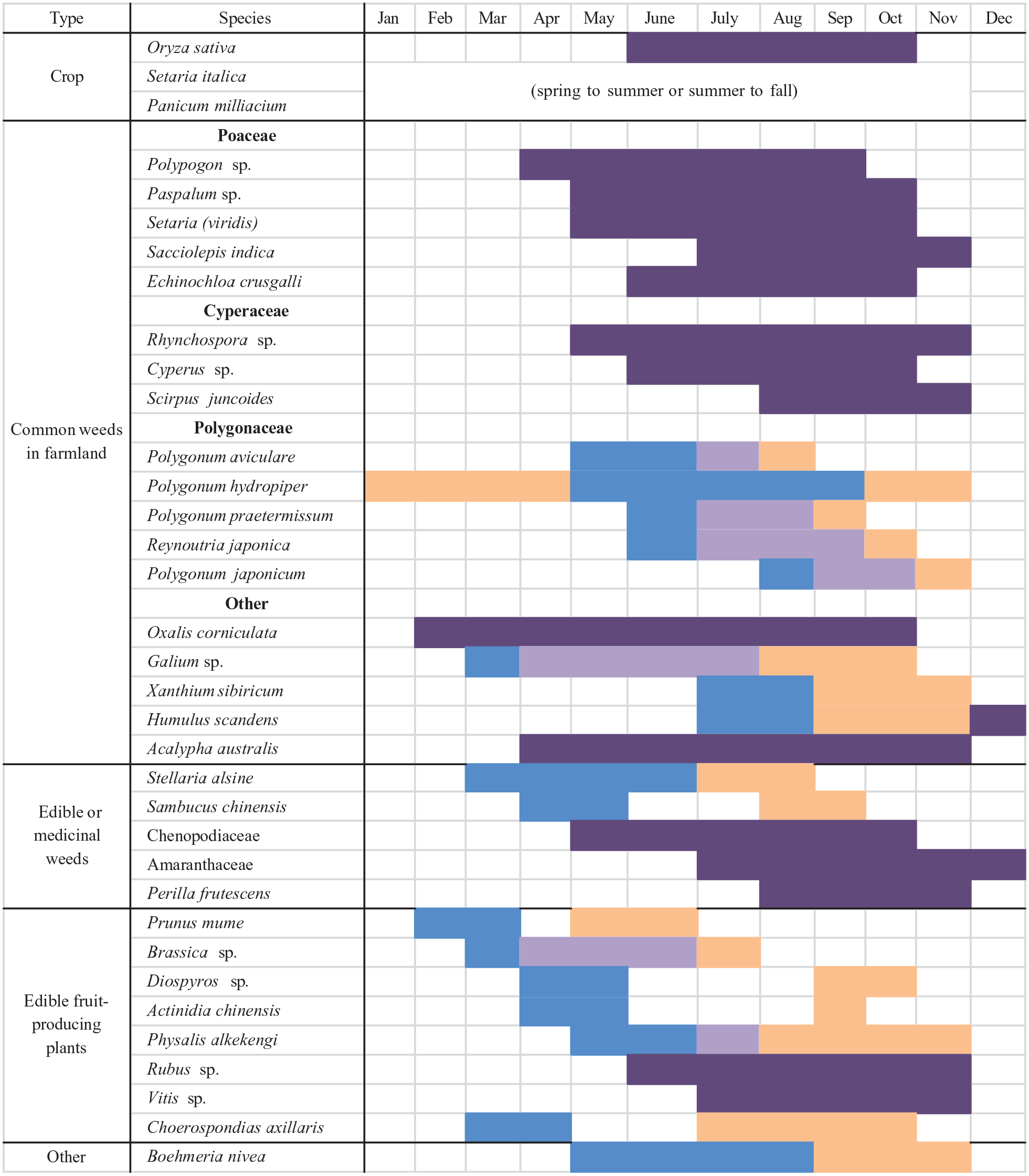
Seasonality of plant taxa, both cultivated and gathered, discussed in the text. Blue: flower; Orange: fruit; Light purple: flower and fruit overlaps; Dark purple: flower and fruit simultaneously. The weedy category is divided by botanical family into three subcategories since each subcategory is discussed separately in the text. Weeds from Poaceae and Cyperaceae are discussed with crops, while weeds from the Polygonaceae and other families are discussed with the edible and medical species. Seasonality is gathered from Wu et al. (2013) and conversations with contemporary farmers in Zhejiang.
Starting in November, there were few food resources available until April. Only P. hydropiper is able to fruit in the winter, probably due to its tolerance to low soil temperature, which makes it one of the few weeds living in paddy fields even in winter (Sultan et al., 1998). The local residents might have paid closer attention to weeds such as P. hydropiper when they began to prepare paddy fields by removing weeds before sowing in May. If millets were sowed in spring and harvested in summer, then residents of Anle could have started millet production in March or April when foraging was less intense and rice production was not yet in process. Plants with edible leaves (Amaranthaceae, Chenopodiaceae, Perilla frutescens, Brassica spp.) become available in the spring and early summer, alongside Prunus mume fruits. The major focus of labor during this period, however, likely centered on paddy fields, as multiple management tasks are required from May/June to August: leveling fields, removing weeds, manipulating water level, manuring, etc. During that time, several fruits (Rubus spp., Vitis spp., and Choerospondias axillaris), herbs (Stellaria alsine), and plants with edible leaves discussed above are available for opportunistic or intentional gathering. Whether local residents intentionally planted or selectively saved C. axillaris trees, they produce a large number of fruits over a long period (July to October). When lepsi fruits mature, they drop to the ground, making gathering an easy task.
Niche construction
Human-plant interactions are not limited to the seasonality of plants and its influence on the rhythm of the human experience. Niche construction theory (NCT) recognizes humans as potent niche constructors who have enormous ability to alter ecosystems and environments through transgenerational learning and application of knowledge (Laland and O’Brien, 2010). It is clear that the residents of Anle created two anthropogenic agricultural niches for rice and millets by constructing specific resource catchments (crop fields) that facilitated the growth of these crops in this region. Rice is an advantageous crop to grow in the Lower Yangtze where sufficient water is available, making it a logical target of initial exploitation as proposed by NCT (Smith, 2011). This perspective on desired characteristics provides insight into millet adoption in the Lower Yangtze as well. Though more labor is required to implement a double-cropping regime of rice and millet, incorporating millet into the diet provides two benefits. The first is that it increases options for storable food. The macrobotanical results suggest that only rice and millets of all the food plants cultivated or collected at Anle could be easily stored for periods of months. Historically, drying persimmon (Diospyros kaki) fruits is a common practice (Luo and Wang, 2008), a practice potentially employed on other fleshy fruits (e.g. Prunus mume). The low count of Diospyros sp. seeds in this assemblage, however, is insufficient to make any specific claim to fruit preservation. The second benefit of adding millet to the diet is that it reduces risks of food scarcity through ecological, spatial, and temporal diversification (Marston, 2011). Many other Neolithic sites in the Lower Yangtze yield acorn remains that imply acorn consumption and storage (Pan and Yuan, 2018; Qin et al., 2011). Environmental studies, however, show that there was a general decline in grasslands, wetlands, and tree nuts across the region around 6000 BP (Fuller and Qin, 2010). Though the absence of acorn at Anle is more likely an effect of sampling one particular site than representing a total lack of oaks in the local area, it is still likely that a regional decline in acorn availability would have influenced food production decisions at Anle. Cultivating millet diversifies food production without introducing competition with rice agriculture for resources such as water and arable land. The short growth cycle of millet is temporally offset from that of rice, avoiding severe labor and resource conflicts with rice production. Anle residents may have utilized the topographical advantage they had in that the low-elevation hills near their settlement meet the growth requirement of millets, an opportunity unavailable to residents of the more numerous contemporary sites located in the floodplains around the Tai Lake.
Another, and more ambiguous, niche includes the resources that grow in close affinity to human disturbance: Sambucus chinensis, Stellaria alsine, Perilla frutescens, Physalis alkekengi, Rubus spp., Vitis spp., Amaranthaceae, and Chenopodiaceae. Many plants in this niche tend to grow near human settlements, thus making them easy targets for foraging. Unlike rice and millets, which received significant selective pressure from humans during cultivation, these exploited plants would have received anthropogenic pressures through a much slower process of long-term exploitation that would ultimately influence their growth pattern and distributions. Herbs such as Perilla frutescens that today grow in waste fields and gardens provide one example. Anle residents created, originally inadvertently but later perhaps deliberately, a resource catchment of “wild” herbs and shrubs near their settlement that facilitated exploitation. Such a process might have eventually broadened to include selective curation or even propagation of fruit-bearing trees (i.e. Actinidia chinensis, Prunus mume, and Choerospondias axillaris) in local forests, as has been observed in semi-managed forest systems of the Southeastern United States (Gremillion, 1993; VanDerwarker et al., 2013). Zhang et al. (2019) suggested a similar practice of creating nearby vegetation catchments at another Neolithic site in the region. The fact that many plants in this niche (alongside Brassica spp., Diospyros spp., and Boehmeria spp.) either become domesticates later in history or are still used in culinary and medicinal traditions today attests to the coevolution of plants and human through multi-generational efforts such as those described here.
Diversification and adoption of new crops in a period of social and environmental change
The diversity of plant resources, both cultivated and gathered, at Anle gives insight into how its residents diversified their subsistence base, reducing the risk inherent in agricultural production (Marston, 2011) and facilitating the introduction of new crops (Boivin et al., 2012; Marston, 2021; van der Veen, 2010). By identifying these subsistence choices, we gain valuable insights into how residents of Anle adopted strategies in response to both environmental uncertainties and cultural change.
The S7 deposit of Anle dates to the Majiabang-Songze transitional period, a period of significant social change in the Lower Yangtze. Growth in regional population and village size date to this period (Liu and Chen, 2012) and social stratification has been suggested to first emerge around 5800 BP based on material differences in grave goods observed in Songze burials (Zhou et al., 2010). Moreover, the material culture of Majiabang indicates that craft specialization was highly developed, as elaborate jade ornaments have been found at various sites, including Anle (Zhejiang Institute, 2014). The Majiabang-Songze period also witnessed either direct or indirect cultural contacts between northern China and the Lower Yangtze in the sixth millennium BP, resulting in new styles of jade objects produced in the Lower Yangtze region (Liu and Chen, 2012). These cultural contacts may also explain the dispersal of millet to Anle and, presumably, the entire Lower Yangtze region where suitable locations for cultivation existed. While adapting local agricultural practices to include millet cultivation is one example of agricultural change, other innovations, including better paddy field construction, water management, and agrarian tools, made intensification of rice production possible during the same period (Fuller et al., 2008; Ma et al., 2016; Zhuang, 2018). The result of such intensification is indeed evident at Anle based on the extremely high count and percentage of rice remains, which suggest intensive cultivation.
It is also clear, however, that environmental change played a role in agricultural decision making at Anle. Middle Neolithic climatic amelioration created a more habitable space even though low-level salinity and periodic inundations of sea water were still in place in the Lower Yangtze region in this period (Chen et al., 2018). As human societies have the capacity to use collective memory to respond to historic risks even if not immediately evident in the present environment (e.g. DiNapoli and Morrison, 2017; Rosen and Rivera-Collazo, 2012), the Anle community, who likely had a memory of the previous unstable climate, may have found that the mountains and hills provided more stability and security for food production. The hilly environment around Anle provided more arboreal resources for exploitation than did the immediate area around Tai Lake (Zhuang, 2018). The higher elevation, inland position of the site protected rice production from inundations of brackish water closer to Tai Lake. The macrobotanical results of this study show that at least two crop niches were intentionally created and a diverse range of gathered and potentially encouraged (perhaps even incipiently cultivated) wild plants also formed a meaningful part of the diet. It would be much more difficult to maintain such subsistence without the presence of low-elevation hills, which fulfilled the necessary requirements for the dualistic rice-millet regime and also offered ready access to seasonal exploitation of diverse wild fruit and herb resources. It is also possible that regional demographic increases and incipient social stratification created new regional stresses on agricultural production, leading to the settlement of Anle adjacent to the mountains and hills bordering the Lower Yangtze region, although data for this causal chain are only circumstantial to date and should be addressed further in future work in the region.
Conclusions
The macrobotanical record of Anle in uniquely rich and varied among published remains from middle Neolithic sites in the Lower Yangtze region. Although rice was the primary cultigen at Anle, as at other sites in the region, we find securely dated early evidence for millet cultivation, though perhaps at an experimental level, with most labor still devoted to rice cultivation. The discovery of meaningful numbers of millet remains at Anle argues for the adoption of northern crops in the Lower Yangtze region at an earlier date than previously documented. Future studies that find more evidence of millet in this region at similarly early dates will clarify the route and timing of millet dispersal, and whether the migration of people or new cultural interactions enabled millet dispersal. Beside millets, the discovery of multiple plants that have a long history in traditional medicine and in later cultivation enriches our understanding of how they were exploited alongside intensive crop production. The diverse subsistence methods documented at Anle help us to understand how people actively constructed their regional landscape through the creation of new ecological niches to promote food availability within an established seasonal system of rice cultivation. More macrobotanical work in this region in the future will help to fill in this broader picture of subsistence created by the emphasis on rice in current published studies. Future work on non-rice plants in the archaeobotanical record of the Middle Neolithic Lower Yangtze region may also challenge the status of “wild” plants, which may have been subject to long-term anthropogenic pressure sufficient to meet the criteria of low-level food production (Smith, 2001), cultivation, or even early domestication. Ultimately, future archaeobotanical studies at other sites in the region should be able to reconstruct a comprehensive botanical record for the Lower Yangtze region, thus strengthening our understanding of how subsistence strategies, not only at Anle but also at contemporary neighboring sites, managed agricultural uncertainty in the context of changing social and environmental landscapes.
Footnotes
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The authors gratefully acknowledge the support of the Henry Luce Foundation, via a grant to Boston University’s Asian Archaeology Program, for financing the radiocarbon dating described here.