
Abstract
Mast seeding – the pronounced inter-annual variability and synchrony in seed production – can have profound local ecological impacts. We explore the potential for generating long (centuries to millennia) proxy records of tree mast seeding, from pollen deposited in lake sediments. This proxy record uses pollen recovered from annually-banded (varved) sediments from a core collected from Lake Ohau, South Island, New Zealand to reconstruct masting events for two genera of Southern Beech, Fuscospora spp. and Lophozonia spp. We find average mast frequencies inferred from Fuscospora pollen from a 43-year sediment core collected from Lake Ohau comparable to seed fall expected for the catchment using a differential-temperature (ΔT) statistical model for the period 1974–2016. In contrast, Lophozonia pollen mast frequency in the 1974–2016 timeseries was consistently lower than that predicted by the ΔT model, although the patterns of variability were broadly similar. We explore this approach in a second 32-year pollen timeseries from Lake Ohau, spanning the pre-instrumental period 1833–1864. During this interval, average air temperature was ~1°C cooler than the late 20th century, and interannual variability of air temperature was subdued, such that mast frequency predicted by the ΔT model is the lowest in 200 years. We find mast frequency in our pollen records reflects this pattern for Fuscospora, with a minima of mast frequency from 1850, compared to the 1974–2016 record, but not for Lophozonia. This paper demonstrates that a centuries-long pollen record from the Lake Ohau sedimentary sequence has the potential to form a valuable proxy for Fuscospora masting that would supplement existing seedfall records. Long records of this type could significantly enhance our understanding of the environmental drivers of mast seeding.
Introduction
Mast seeding is the pronounced inter-annual variability and synchrony in seed production exhibited by many long-lived plant species around the world (Kelly, 1994). The local ecological impacts associated with these pulses of enhanced seedfall resources can be profound and commonly include rapid population increases in seed predators followed by cascading effects on other species (Bieber and Ruf, 2005; Curran and Leighton, 2000; Dingemanse et al., 2004; McShea, 2000; Ruscoe et al., 2004). Famously, these include the series of interactions that link mast seeding by Quercus to outbreaks of Lyme’s disease in eastern North American forest ecosystems (Jones et al., 1998). Given the influence of mast seeding on ecosystem dynamics, there is an imperative to understand the environmental cues that precede mast seeding events. Novel methods to extract long historical mast records could yield insights into past mast seeding behavior and provide the necessary time series data to resolve climate-masting relationships that would improve prediction of future mast behavior (Isagi et al., 1997; Piovesan and Adams, 2001; Schauber et al., 2002; Szabó, 2012).
In the Northern Hemisphere, decadal-scale seed fall data sets have been extended to multi-century masting records (Ascoli et al., 2017; Szabó, 2012), by collating archival records (Hilton and Packham, 2003; Övergaard et al., 2007), and by using tree ring records for species with a strong relationship between reduced wood growth and enhanced seed production (Drobyshev et al., 2010, 2014; Eis et al., 1965). These historical compilations are, however, out of reach for many countries including New Zealand, where data on mast seeding is urgently needed to develop better predictive models for managing a chronic conservation issue. In New Zealand forests, mast years lead to rapid increases of non-native mouse populations, followed by increases in non-native mustelids, which in turn predate highly vulnerable native bird and reptile fauna (Choquenot, 2006; Fitzgerald et al., 2004; Ruscoe et al., 2004; Veale et al., 2015). In this context, understanding the environmental cues that lead to mast years is crucial for conservation planning, both on annual timescales for predator control, and decadal timescales to anticipate likely changes with increasing climate variability and warming. The empirical datasets of seed fall in New Zealand used to derive relationships between environment and mast seeding are limited to the past ~40 years (Allen et al., 2014; Kelly et al., 2013), which has been too short a time to conclusively resolve relationships between climate and mast seeding. New Zealand is a country with a relatively short written history, and European settlers did not identify economic value (and thus reason to record variability) in any native seed crop. As a consequence, there are no archival records of seedfall suitable to assemble multi-century mast records for any plant species in New Zealand. Having longer records which cover the modern instrumental period would significantly strengthen our understanding of the environmental drivers of mast seeding.
Here, we explore the potential for generating a long (centuries – millennia) proxy record of masting for two genera of Nothofagaceae, Fuscospora spp. and Lophozonia spp. – which both exhibit strong mast seeding behavior (Schauber et al., 2002). This proxy record uses pollen recovered from annually banded (varved) sediments from sediment cores collected from Lake Ohau (44°16.782′S, 169°55.480′E), New Zealand. To the best of our knowledge, this approach has not previously been explored in detail: limnologists and paleoclimatologists who most frequently work with varved sediments have not recognized the great value of these lake-borne archives for this type of ecological research.
Methods and site setting
Three key assumptions that underpin the use of pollen abundance as a proxy for masting are; (1) that there is a strong correlation between the amount of pollen released and seed fall, (2) taphonomic effects do not obscure seasonal variability of pollen rain entering Lake Ohau, and (3) that pollen extracted from Lake Ohau sediments faithfully records the regional inter-annual variability in pollen production. We assess the validity of these assumptions. We describe two small experiments of pollen release compared to seedfall, and to pollen accumulation rates in Lake Ohau sediment. We then describe sediment core collection, development of an annually resolved age model, and analysis of pollen variability, from two multi-decadal timeseries (1974–2016 and 1833–1864), that were derived from Lake Ohau sediment cores. The 1833–1864 window represents an interval of NZ paleoclimate that was colder than present, (so allowing us to sample a contrasting temperature environment), but also some decades after the 1717 AD Alpine Fault earthquake that we know had a major impact on landscapes around Ohau (Roop et al., 2016; Wells et al., 1999, 2001).
Relationship between modern pollen rain and seed fall in central New Zealand
New Zealand Nothofagaceae are monoecious, self-incompatible, and have not been recorded to set seed apomictically (Wardle, 1984); therefore, mast seeding events are always preceded by periods of heavy flowering (Alley et al., 2001; Fitzgerald et al., 1996; Powell and King, 1997; Sweetapple, 2003). However, not all heavy flowering years result in mast seeding events because adverse weather can damage flowers and preclude successful pollination, and ovules can be aborted after pollination (Wardle, 1984). Consequently, while there is an imperfect correlation between flowering and seedfall in New Zealand Nothofagaceae (Wardle, 1984), a relationship between flowering, pollen release, and seed fall would not be unexpected, as such relationships have been observed in a number of tree species, including Fagus and Quercus (Bogdziewicz et al., 2017; Nussbaumer et al., 2020). There are no long-term combined records of flowering and seedfall because Nothofagaceae flowers are small, fragile, and compared to seedfall, relatively difficult to sample.
We partially address this shortcoming by collating four short opportunistic time-series of Fuscospora pollen rain from different locations around central New Zealand and compare these to seed fall counts from Fuscospora truncata sampled in the same region (Figure 1). Two of the four pollen rain datasets are from terrestrial locations (Wellington region (1994–1997) and Palmerston North City (2012–2014)) and two are from deep ocean sediment traps (Wairarapa Coast (2005–2009) and the Southern Ocean (2006–2009) (Figure 1). We have calculated average daily pollen rain for the 6 months after 1 July (i.e. including austral spring). The exception to this is the Palmerston North dataset, where only the spring August-December period was sampled. Seed fall counts came from the Orongorongo Valley (41.347354°S, 174.967354°E) where annual seed production is measured underneath target trees using circular traps (DoC, 2016; Tompkins et al., 2013). Interannual variation in seed production by Fuscospora is highly synchronized, even at distances of >500 km (Schauber et al., 2002): while the absolute seed fall values are likely to be site-specific, the year-to-year variation in mast seeding events at Orongorongo is likely to be representative of the pollen rain sites. Flowering and pollen release occurs from October to December, and seed falls the following February to September. For ease of comparison, we here label flowering according to the year in which seed falls in all plots.
Location map showing Lake Ohau (yellow star), sites used in seedfall comparison for Fuscospora (red square) and sites used for Fuscospora pollen rain compilations (solid blue square).
While these datasets are each too short to support formal testing of difference between mean pollen rain preceding mast and non-mast years, they are the first comparison of New Zealand beech pollen rain and seedfall we are aware of.
Pollen deposition into Lake Ohau
Lake Ohau (44.234°S, 169.854°E; 520 m asl) lies in the temperate Mackenzie Basin, South Island, New Zealand (mean winter air temperature 6.2°C) (Roop et al., 2016) (Figure 1). The lake is 54 km2, glacially formed, has a maximum depth of 129 m and the catchment is 924 km2, with a maximum catchment elevation of 2640 m asl. Contrasting seasonal catchment hydrology generates annual laminations in Lake Ohau, which allows an annually resolved multi-millennial chronology that is so far unique in the Southern Hemisphere (Roop et al., 2015, 2016). An 80 m core recovered from the lake in 2016 exhibits these annual laminations (Figure 2) extending for the last 17,000 years (Dunbar et al., 2017; Levy et al., 2018).

Age depth model of sediment core GC170427-3 (upper) and LOCH1 (lower) collected from Lake Ohau. Both cored intervals were sampled for pollen at 2.5 mm resolution. Core image is generated from a sagittal slice through the CT tomography capture.
Vegetation in the catchment is a mixture of low-producing tussock grasslands, alpine ecosystems in the upper reaches and above tree line, and forest. Forest communities are dominated either by Fuscospora cliffortioides (mountain beech) close to the lake or Lophozonia menziesii (silver beech) in the headwaters (Wardle and Guest, 1977).
We measured the timing of pollen entry into Lake Ohau, to establish the fidelity of the seasonal and inter-annual signal. A Rice University-type sediment trap (MacPherson, 1987) was moored continuously in Lake Ohau between late 2012 and late 2016. The trap was located in the southeast arm of the Lake. The intake for the trap is 1.5 m above the lake floor. The water depth at the trap site is 8 m and the maximum depth of the lake is 129 m. The site was selected to correspond with the location of previously collected sediment cores, providing an opportunity to relate observed deposition trends with those evident in the paleo record (Roop et al., 2016). The trap was emptied twice a year, during autumn and spring. One sample is missing from this timeseries: the August–November 2014 (i.e. austral spring) sample was lost during collection. The sample of the following austral spring (June-November 2015), may be an incomplete sample, because upon recovery the trap showed signs it had been tipped for an unknown portion of its deployment (See data, Supplemental Table A5).
Trap sediment (approx. 4 g) was processed for pollen, taken from a well-mixed subsample from each available trap cup. The bulk sediments were sieved through 6 µm Nybolt filter cloth first to get rid of clayey materials which is <6 µm and compose the major fraction of the sample. After this step, pollen extraction was carried out using standard laboratory techniques (10% hot HCl, KOH, heavy liquid density separation, and acetolysis of a known mass and volume of sample (Faegri et al., 1989)). A recorded proportion of pollen residue was mounted onto microscope slides and counted.
Lophozonia is represented by a single species in New Zealand (L. menziesii); Fuscospora has four species native to New Zealand (Heenan and Smissen, 2013), but their pollen is indistinguishable (Moar et al., 2011). Forests around Lake Ohau are dominated by just one species of Fuscospora – F. cliffortioides – and the nearest other Fuscospora species – F. fusca – is present as scattered individuals at least 30 km away over the Southern Alps, so it is highly likely that Fuscospora pollen is from F. cliffortioides. To be conservative though, pollen is referred to at a generic level in this study.
Lake Ohau sediment cores and age models
Gravity core GC170427-3 (Figure 2) was collected using an Alfred Wegener Institute-style gravity corer from a 6.8 m Naiad rigid inflatable boat. Core 6m1S was a Mackereth core collected at the LOCH1 drill site, of which the upper 1.5 m section was sampled for this study. Both cores consist of dark gray, millimeter-laminated lithogenic mud. The mm-scale laminations provide the basis for correlation between each core and the longer LOCH1 composite stratigraphy built on core GC170427-3, 6M1S and LOCH1 piston cores (Levy et al., 2018).
The age models for both cores provides for annually resolvable accumulation. Age model development included (1) generating a single density-depth profile from CT scans of the core, (2) normalizing the density-depth profile to remove the effects of compaction, and (3) picking annual layers based on the density amplitude, spacing, and gradient while incorporating age independent age constraints, including 137Cs profiles, biostratigraphic horizons and 14C ages (Vandergoes et al., 2018). Further details on the age model generation are included in Supplemental Text.
The resulting age model assigned an age of 1974 to 355 mm depth in core GC170427-3, with an average sediment accumulation between 1974 and the 2016 core top of 8.2 mm/year. In core LOCH1, the interval 1150–1430 mm spans 1833–1864, with an average sedimentation accumulation rate of 8.75 mm/year (Figure 2).
Sediment core pollen sampling
We measured modern pollen inputs to Lake Ōhau, and then documented pollen variability from two annually-resolved multi-decadal sedimentary timeseries collected from Lake Ohau: between 1974–2016 and 1833–1864.
The two intervals, covering the nominal years 1974–2016 and 1833–1864, were continuously sampled at 2.5 mm resolution from a half-core of diameter ~ 60 mm, yielding sample volumes of ~3cm3. On average, these 2.5 mm samples represented ~4 months accumulation, and the maximum time assigned to a single 2.5 mm sample in the age model was 10.3 months. Given the duration of Spring pollen release is ~4 months (observed into Wellington pollen traps, Supplemental Data A1), it is possible that a small number of samples (<5%) could include pollen release from part of two consecutive Springs, which could have the effect of obscuring adjacent mast events in our pollen record.
Pollen extraction was carried out using standard laboratory techniques (10% hot HCl, KOH, heavy liquid density separation, and acetolysis (Faegri et al., 1989)). All core sample material collected was processed, and the pollen residue was mounted onto microscope slides and counted to the finest taxonomic level possible for the New Zealand pollen flora. Pollen abundance was calculated by counting a known proportion of weighed sample. During the course of our study, we were able to improve rates of pollen recovery, by adjusting our filtering technique. Average pollen abundance in the last batch of samples from core GC170427-3 processed (between 162.5 and 222.5 mm, year 1992 and 1998) was 11.3× that of the rest of the timeseries. We shifted entire pollen counts from GC170427-3 to the same magnitude scale prior to frequency analysis, by multiplying the pollen counts outside of this interval by 11.3, resulting in equivalent frequency distribution of the raw and shifted data sets (Supplemental Figure 1).
There are no records of beech seedfall from the Lake Ohau catchment, nor any long records of seedfall within 200 km of our site. Instead, we compare our modern (1974–2016) pollen record to an empirical statistical model predicting seedfall, from the temperature (“T”) difference between the two previous summers (mean monthly temperature for January to March; the ΔT model), which is the strongest climate-seedfall correlation for the New Zealand flora (r2 ~0.45) (Kelly et al., 2013). We applied a relationship between ΔT and log10 seedfall of 0.696*ΔT + 1.676, and followed Barron et al. (2016), where a threshold value of >180 seeds/m2 (a ΔT threshold value of 0.84) corresponded to mast years resulting in increased predator populations in New Zealand beech forests. For the 1974–2016 time interval, we use temperature data from the NIWA virtual climate station network, extracting average Jan–Feb–Mar temperature for a 45 × 45 km block around point 169.85° longitude,−44.23° latitude for years 1972–2019. For this time interval, the ΔT model predicts seedfall frequency is low during the 1980s and increases through the late 1990s to early 2000s (Figure 4d).
For the 1833–1864 time interval, we explored the same approach using a temperature record estimated from tree ring thickness, from sub-fossil tree stumps retrieved from Oroko Swamp (West Coast South Island), 100 km north of Lake Ohau (Cook et al., 2002), but found that record to be unsuitable, because a tree-ring proxy for paleo temperature has three key disadvantages in this context. First, the proxy-calibration itself for air temperature and tree ring width is imperfect (correlation coefficient r = 0.61 to Jan-March air temperature) (Cook et al., 2002); second, correlation has also been reported between tree ring thickness and mast events in other New Zealand tree species (Monks and Kelly, 2006); and third, while both Lake Ohau and Oroko Swamp lie in the same climate zone (Kidson, 2000), the 100 km distance, and Southern Alps mountain range between the two sites suggests we should expect divergence between their temperature records. It is perhaps not surprising then, that when the ΔT model is applied to this temperature dataset, a ΔT threshold value >0.84 predicts fewer mast events than would be expected compared to mast frequency from typical seedfall records. We do however use this temperature record to justify our selection of the 1833–1864 time period. The record does suggest that the interval of lowest year-to-year temperature variability, and therefore lowest seedfall frequency since 1800s occurred within our 33-year time series, around 1850–1870 (Supplemental Figure 2), but the temperature record is not suitable to predict quantitative mast frequency.
Analysis of mast frequency from pollen record
Mast frequency was estimated from the abundance of Fuscospora and Lophozonia pollen respectively, using a “CharAnalysis” model. CharAnalysis is an analytical tool, normally used for sediment-charcoal analysis (Higuera et al., 2010). In this capacity, it applies a decomposition method, separating background charcoal accumulation from charcoal “peaks,” which are considered to represent fire events. In this study, CharAnalysis was used to estimate and detrend background beech pollen accumulation and identify significant beech pollen peaks. Peaks in pollen abundance are here inferred to reflect beech masting years. We use the term “pollen mast” through-out this manuscript to describe a mast event inferred from pollen abundance. The parameterization of the CharAnalysis model is included in Supplemental text. All figures relating to CharAnalysis results were created using R v.40.00.5 with package ggplot2 (Wickham, 2016).
Results and discussion
Modern pollen rain
All four modern pollen rain datasets show greater Fuscospora pollen flux in years associated with Orongorongo Valley mast years than years of low seedfall (Table 1; Supplemental Appendix A1–A4). Across the four short datasets, the difference in Fuscospora pollen flux between the smallest mast and most prolific non-mast years ranged from a factor of 2.98 to 32.92: at least 2.98 times more pollen was measured during the spring preceding mast years than non-mast years.
Fuscospora pollen flux from four locations compared to Orongorongo Valley F. truncata seed fall. Gray bars highlight “full” and “partial” mast years following criteria of Wardle (1984).

For the three modern pollen rain data sets in which fewer non-mast than mast years were sampled, the pollen rain during the spring preceding mast years was between 2 and 3 times the average pollen rain. This is a similar range to that observed in European Fagus pollen rain studies between mast and non-mast years (Pidek et al., 2010; Simoleit et al., 2016). Although the ocean sediment traps appear to record a lower Fuscospora pollen flux than the terrestrial sites, it is worth noting that the ratio of mast to non-mast years is similar.
Pollen in Lake Ohau sediment trap
The highest concentrations of Fuscospora and Lophozonia pollen in the Lake Ohau sediment trap were observed in samples that were deployed during spring (September–November), coinciding with flowering (Figure 3, Supplemental Appendix A5). Fuscospora pollen concentrations in traps collected during the spring preceding mast years (2013/4 and 2015/16) were 7.9 times the average pollen concentrations in all other trap samples, while Lophozonia pollen concentrations during those two spring seasons were 43 times the background average.

Pollen rain in Lake Ohau sediment trap, 2013–2017. Red solid line: Fuscospora pollen concentration in trap sediment (grains/ml of sediment); Green dashed line: Lophozonia pollen concentration in trap sediment (grains/ml of sediment). Gray bars in sediment trap data denotes missing or compromised sample. Black arrows denote mast year, inferred by ΔT value >0.84, from Figure 4a.
Lake Ohau sediment results
Fuscospora and Lophozonia pollen extracted from Lake Ohau sediment exhibited a high degree of inter-annual variability for both the 1974–2016 and the 1833–1864 time series (Figure 4b and c, Supplemental Appendices A6–A9).
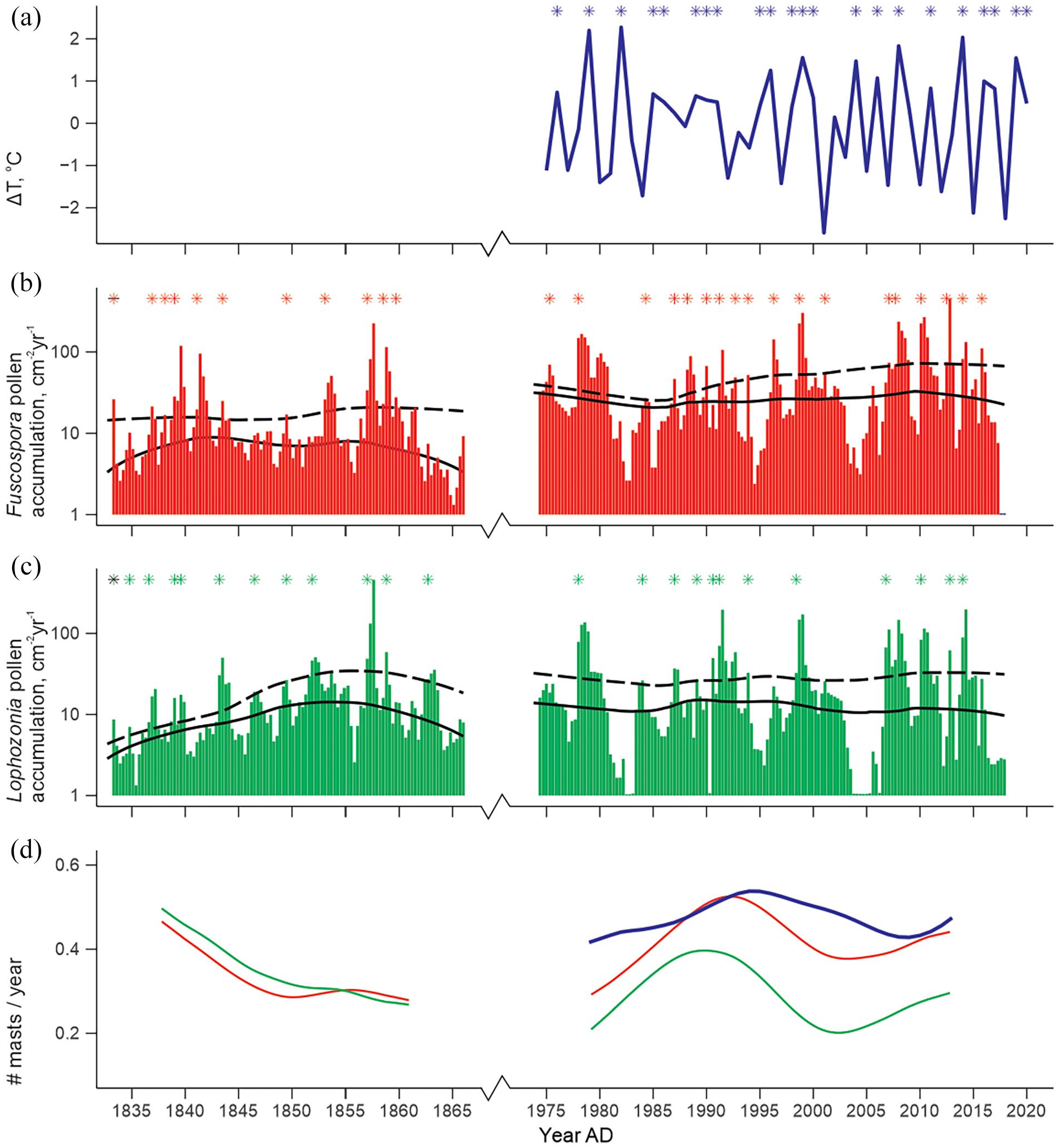
Masting frequency inferred from pollen, for interval: 1833–1864 (left column) and 1974–2016 (right column), compared to mast frequency inferred from ΔT climate statistic. (a) Calculated ΔT at Lake Ohau based on VCSN climate network data (right, dark blue). (b) Fuscospora pollen accumulation (red line) (c) Lophozonia menziesii pollen accumulation, green. Asterisks indicate inferred mast years, either by exceeding a ΔT threshold value >0.84 (a), or by CharAnalysis identification of peaks in pollen accumulation above background (b and c). In (b and c) solid line indicates background accumulation based on a LOWESS smoother; dashed line indicates 95% threshold of a Gaussian mixture model, used to identify significant peaks. Note the logarithmic scale in (b and c) (+1 to all accumulation values to improve legibility). (d) Mast frequency in number of masts per year, calculated over a 15-year window: Fuscospora (red), Lophozonia menziesii (green) and VCSN climate network ΔT (dark blue).
CharAnalysis results from the 1974 to 2016 Fuscospora timeseries show average pollen mast frequency to be comparable to that predicted by the ΔT model (Figure 4d). Both Fuscospora pollen mast frequency, and mast frequency predicted by ΔT, increased from 0.3 to 0.4 masts/year around 1980, peaked at 0.5 masts/year in the early-mid 1990s, then declined during the 2000s to around 0.4 masts/year. In contrast, Lophozonia pollen mast frequency in the 1974–2016 time series was consistently lower than that predicted by the ΔT model (Figure 4d). However, the pattern of change in mast frequency, was broadly similar to that of the ΔT model; with an increase in frequency through the 1980s, peak frequency in the early 1990s, and a low during the 2000s.
In the 1833–1864 time series, there were strong similarities between Fuscospora and Lophozonia pollen mast frequency (Figure 4d), with pollen mast frequencies of 0.4–0.5 masts/year observed in the early 1840s, declining to below 0.3 masts/year in the period 1850–1860. This pattern, of relatively lower frequency after 1850, are generally consistent with relative seed fall we might expect from Oroko air temperature record.
Uncertainties and implications
Results thus far suggest multi-centennial proxy records of pollen mast will provide insights into masting variability that are complementary to seedfall records. However, longer timeseries – at least twice as long as have been generated here, both of sedimentary records of pollen accumulation, and proximal records of air temperature – and more robust reference datasets of seedfall, would increase confidence.
Testing of our pollen-mast proxy is hampered by the lack of proximal seedfall records. Although the ΔT model resulted in the strongest climate-seedfall correlation tested across New Zealand taxa (Kelly et al., 2013), typical r2 values are <0.5. Due to its imperfect ability to predict seedfall, it is not an ideal target against which to test variability of the pollen dataset. When assessing the value of this approach for the pre-instrumental record, significant further uncertainty arises from our generation of a ΔT statistic from a distal, tree ring-derived paleotemperature record, itself underpinned by an imperfect correlation between climate and annual tree ring widths.
Notwithstanding the considerations described above, of the two genera investigated, Fuscospora appears to demonstrate stronger potential as a pollen-based proxy for masting than Lophozonia in this setting. This is because changes in Fuscospora pollen abundance extracted from Lake Ohau sediment are consistently closer to seedfall predicted by the ΔT model, in both the 1974–2016 record, but also the 1833–1864 record. While Lophozonia shows strong alignment in the older timeseries, we observe a relatively poor match in the 1974–2016 record. This different pattern between the genera is surprising, as in our short timeseries of pollen entering the lake: Lophozonia pollen was 43× more frequent during mast years than non-mast, while Fuscospora pollen was only 7.9× more frequent during mast years. However, Fuscospora pollen is approximately twice as frequent as Lophozonia in the 1974–2016 record, but the two types have similar frequencies in the 1833–1864 record. The reasons for this difference in pollen rain are not clear. Longer pollen records (in preparation) from the lake do not show any compelling evidence of Lophozonia retreat during the 20th century. However, it may reflect changing pollen source areas over the last 150 years, perhaps arising from temperature-driven shifts in wind direction or intensity (Ahmed et al., 2013; Fowler et al., 2012).
The pollen masting approach described here will be of particular value in interpreting changes in mast frequency on decadal and multi-decadal scales of any anemophilous plants. For example, a multi decadal record would establish the uniqueness or otherwise of the changes in mast frequency observed in New Zealand beech forests since 1970 (Tompkins et al., 2013). Another intriguing application would be to extract Dacrydium cupressinum (Podocarpaceae) pollen from this record, which appears to have an environmental cue to masting different from other New Zealand plants, but for which only limited seedfall records exist (Norton and Kelly, 1988).
Conclusions
We explore the potential for generating long (centuries to millennia) proxy records of tree mast seeding from pollen captured in annually laminated lake sediments, from Lake Ohau.
We have observed a strong inter annual variability in the flux of Fuscospora pollen at five sites in the New Zealand region. The pollen flux during the spring preceding mast years was between 2 and 3 time the average pollen rain. This strong interannual variability of pollen rain was also observed in an incomplete 3-year sediment trap timeseries moored in Lake Ohau. Fuscospora pollen concentrations in the spring preceding seed mast years was 7.9 times the average pollen concentrations, while Lophozonia pollen concentrations preceding a mast year were 43 times the average.
Within an annually resolved sediment record from Lake Ohau, we observe a strong correspondence between Fuscospora pollen mast frequency, and mast frequency predicted from a temperature difference (ΔT) model, from two multi-decadal timeseries (1974–2016 and 1833–1864). In contrast, pollen mast frequency inferred from Lophozonia pollen is lower than would be predicted by temperature difference (ΔT) model for 1974–2016 timeseries, but comparable for the older 1833–1864 series. The lower pollen mast frequency inferred from Lophozonia pollen abundance may be due to changing transport pathways of Lophozonia pollen, due to changing wind patterns over the last two centuries.
A centuries-long pollen record from the Lake Ohau sedimentary sequence has the potential to form a valuable proxy for Fuscospora masting that would supplement existing seedfall records. Such a record could provide a unique mechanism to extend the modern empirical datasets back in time, to allow the investigation of correlation between past climate change and masting frequency.
Supplemental Material
sj-docx-2-hol-10.1177_09596836221114292 – Supplemental material for Toward a novel multi-century archive of tree mast using pollen from lake sediments
Supplemental material, sj-docx-2-hol-10.1177_09596836221114292 for Toward a novel multi-century archive of tree mast using pollen from lake sediments by Joseph G Prebble, Gavin Dunbar, Valerie van den Bos, Xun Li, Marcus Vandergoes, Sarah Richardson, Huw Horgan, Katherine A Holt, Jamie Howarth and Richard Levy in The Holocene
Research Data
sj-xlsx-1-hol-10.1177_09596836221114292 – for Toward a novel multi-century archive of tree mast using pollen from lake sediments
sj-xlsx-1-hol-10.1177_09596836221114292 for Toward a novel multi-century archive of tree mast using pollen from lake sediments by Joseph G Prebble, Gavin Dunbar, Valerie van den Bos, Xun Li, Marcus Vandergoes, Sarah Richardson, Huw Horgan, Katherine A Holt, Jamie Howarth and Richard Levy in The Holocene
Footnotes
Acknowledgements
GNS Science contribution to this project was supported by the Global Change Through Time Programme via the New Zealand Strategic Science Investment Fund (SSIF). SJR was supported by SSIF provided to Manaaki Whenua- Landcare Research. We thank two anonymous reviewers and the journal editor for comments that have improved this paper.
Author contributions
Conception, design and drafting: JP, MV, GD; acquisition of data: XL, VVdB (Ohau pollen), GD, MV, HH, RL (Ohau cores, traps and age models), KH (Palmerston North pollen trap); analysis and interpretation of data: all authors.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship and/or publication of this article: GNS Science contribution to this project was supported by the Global Change Through Time Programme via the New Zealand Strategic Science Investment Fund (SSIF). SJR was supported by SSIF provided to Manaaki Whenua- Landcare Research.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.