
Abstract
Patterned rich fens have a diverse flora and are maintained by unidirectional inflowing water with high concentrations of base cations, along with high pH and limited nutrients. Rich fens are among the most threatened ecosystems in Europe, but are not uncommon across the western boreal forest zone of Canada. Utilizing 10 radiocarbon dated cores extracted from the wetland, we investigated the Holocene developmental history of a large patterned rich fen in northeastern Alberta (McClelland Wetland). Organic matter accumulation began around 11,457 cal yr BP as primary peat formation on recently deglaciated substrates. Over the 10,000+ years history of McClelland Wetland, the central portions of the wetland have been remarkably resistant to change, with little alteration in dominant species. The resistance to change is set against a background of fluctuations in regional Holocene climate and local varying water balance. The dominant bryophyte species (Hamatocaulis vernicosus and Scorpidium scorpioides) continued to play a foundational role on site for the duration of the Holocene, dependent on a persistent long-term ground water source. Accumulation rates of organic matter have remained steady once the dominant bryophyte layer was established, but with accumulation rates at paludifying marginal sites lower than those of the central moss-graminoid-dominated areas.
Keywords
Introduction
Hydrological inputs from precipitation and inflowing surface and ground waters regulate the mineral composition of peatland surface water, including pH, base cations, alkalinity, and inorganic nutrients, and provide the basis for differing peatland site-types. Peatlands are hydrologically divided into ombrogenous bogs and minerogenous fens. In bogs, the surface water and that of the upper peat column are derived only from precipitation, while fens receive additional inputs from surrounding mineral soils or water bodies. Based on direction of inflowing water, fens can be divided into soligenous peatlands, those that receive unidirectional inputs and usually have a patterning of raised strings and elongated pools (flarks – Andersson and Hesselmann, 1907), or those that are located in topogenous basins with inputs from the surrounding landscape, and have no patterning.
DuReitz (1949) recognized differences in plants occurring among various fen sites in Scandinavia, with some sites having a rich and unique flora, while others had fewer species (a poor flora), but with high fidelity. Witting (1947, 1949) was the first to recognize differences in pH and cation concentrations between the rich and poor categories, and Sjörs (1950) related the poor–rich floristic gradient to the pH and electrical conductivity of surface water (see also Sjors and Gunnarsson, 2002; Økland et al., 2001). The water chemistry of the poor–rich floristic gradient has been examined in numerous studies (reviewed by Gorham et al., 1987; Horton et al., 1979; Malmer, 1986). In addition to the differences in species diversity between poor and rich fens, poor fens are typically dominated by Sphagnum and ericaceous dwarf shrubs, while rich fens have abundant herbaceous and graminoid vascular plants and true mosses (Bryopsida). The partition between Sphagnum and true mosses represents the most distinct floristic gradient along the poor-rich continuum (Gorham and Janssens, 1992).
Rich fens, especially extreme-rich fens, are of specific interest as they harbor high species richness (Vitt et al., 1995), contain unique species assemblages (Janssen et al., 2016), and are important habitats for many rare and threatened species, for example, land snails and calcium-tolerant brown mosses (Horsáková et al., 2018; Jiménez-Alfaro et al., 2012). Rich fens are restricted to areas having base-rich groundwater that maintains basic pH and relatively poor availability of nutrients (phosphorus and nitrogen). In Europe, polluted areas with high land-use intensity, rich fens have lost many of their specialized plant species (Bergamini et al., 2009; Hájek et al., 2015; Kooijman, 2012) and are among the most threatened habitat types (Janssen et al., 2016).
Extreme-rich fens are dependent on a long-term constant ground water source with high cation content (especially Ca2+) and high alkalinity. In cases where these conditions remain constant over millennia, rich fens have undergone little internal change, with a high degree of species resilience; however, these situations are uncommon across the boreal realm (Kubiw et al., 1989; Yu et al., 2003, 2014; Zoltai and Johnson, 1985). In areas with topogenic fen basins or ground water with less alkalinity, autogenic changes within the peatlands provide conditions for succession to other fen site-types (Kuhry et al., 1993). Other paleorecords from boreal peatlands have shown recent increases of Sphagnum mosses and shifts toward ombrotrophic conditions during the 20th century, coinciding with warming and lengthening of the growing season (Loisel and Yu, 2013; Primeau and Garneau, 2021; Robitaille et al., 2021; van Bellen et al., 2018). The change from rich fen to Sphagnum-dominated poor fen and bog vegetation includes an ecosystem-scale shift, with the potential increase of carbon accumulation (Loisel and Bunsen, 2020).
Peatlands are abundant across the boreal plain of western Canada with approximately 23% of the area covered by fens and bogs (Halsey et al., 1998). Approximately two-thirds of these are fens and one-third are bogs. The distribution and composition of vegetation of fens are related to the pH, base cation contents, and alkalinity of surrounding hydrological inflows. Acidic poor fens are associated with low electrolyte concentrations in waters of areas with poorly weathered bedrock, a situation common on the siliceous Canadian and Fennoscandian shields (Tahvanainen et al., 2002; Vitt and Bayley, 1984). In western boreal Canada, an area largely of sedimentary limestones and shales, poor fens are restricted to topographic highs – for example, Swan Hills (Vitt et al., 1975), and Caribou Mountains (Horton et al., 1979), or to sandy outwash substrates – for example, Mariana Lakes area (Graham et al., 2016). Circumneutral (moderate-rich fens) and alkaline rich fens (extreme-rich fens) are abundant along the eastern foothills of the Rocky Mountains (Slack et al., 1980), and extreme-rich fens become less frequent eastward, except in local areas influenced by calcareous ground water discharge systems also high in base cation content and alkalinity (Vitt et al., 2022).
Peatlands across boreal western Canada have gradually increased in abundance on the landscape over the past 7000–8000 years (Halsey et al., 1998), and have accumulated organic matter to depths of between 200 and 250 cm, with less than 5% of these having depths greater than 450 cm (based on review of 371 sites – Vitt and Wieder, 2008). In general, over the course of the Holocene, peatlands of the region have undergone one of four developmental pathways. Firstly, at higher elevations and on drainage divides, peatlands initiated as Sphagnum-dominated poor fens and in the Late-Holocene diversified into bog-poor fen complexes (Nicholson and Vitt, 1990). Secondly, initiating as either marshes or shallow pools and early on developing into true moss-dominated rich fens (Bauer et al., 2003), sites expanded rapidly paludifying the surrounding uplands, and in some cases with consequent isolation from mineral waters and Sphagnum invasion, sites developed into bogs (Kuhry et al., 1992, 1993). Thirdly, situated on short-lived, Late Glacial lakes that dried in the Early Holocene, bogs developed and maintained ombrotrophy throughout the Late-Holocene (Bloise, 2007). Fourthly, in the foothills of the Canadian Rocky Mountains and at lower elevations farther east, peatlands initiated as true moss-dominated rich fens with high alkalinity and base cations. These rich fens, with a constant supply of calcareous water, have remained unchanged as wet, moss-graminoid rich fens paludifying surrounding landscapes as wooded or shrub-dominated rich fen site-types (Kubiw et al., 1989).
Patterned fens (or northern ribbed fens) are “very common in the Mid Boreal and High Boreal Regions” (Zoltai et al., 1988); however, few studies have investigated the developmental histories of these unique peatlands. Zoltai et al. (1988) presented an overview of one patterned fen located in north-central Alberta wherein they described rich fen species (Scorpidium scorpioides) continually present for the 175 cm length of a core 210 cm long and dated at ca. 7700 cal yr BP. They reported the long-term accumulation rate of 0.31 mm yr−1. This developmental pattern compares well with historical studies of non-patterned rich fens in other parts of Alberta (e.g. Bauer et al., 2003; Kubiw et al., 1989; Yu et al., 2003). From the limited studies available across the western boreal region of Canada, these unique peatlands appear to have remained largely unchanged for much of their Holocene development, in contrast to autogenic successional changes that have occurred in other peatlands.
Here, we examine the Holocene development of a large patterned fen, with the following objectives: (1) Utilizing a set of radiocarbon-dated cores extracted throughout the peatland, determine the developmental history of the fen, (2) place the early history of the peatland into the post glacial chronology and peatland development, as presently known, of the area, (3) examine the rates of long-term organic matter accumulation and relate these to spatial development of the fen, and (4) explore whether changes in dominant species and overall vegetation have changed or remained constant over the life of the peatland.
Study area
The McClelland Wetland Complex is located at 57°27′01.81″N, 111°25′35.90″W, 83 km north of Fort McMurray, Alberta. The complex is approximately 10 km long and 5.0 km wide at its widest point, with an area of 3835 ha (Figure 1). The patterned area contained within the wetland is 6.6 km long, 2.5 km wide, and 720 ha in extent, with an elevational gradient from 300.4 to 292.8 m.

Google image of McClelland Lake Wetland (2005). Outline of vegetation zones surrounding the patterned fen and location of cores 1–10 (Black) and cores 11–13 (not used analyzed for macrofossils-Pink). L = Larix-dominated rich fens; P = areas of permafrrost/bog/fen swamp complexes, with permafrost-dominated peat plateaus that are actively thawing; G = moss/graminoid fens; and R = riparian swamp forests dominated by Picea mariana. N = area influenced by northern water sources (=northern patterned fen), S = area influenced by southern water sources (southern patterned fen), arrows denote direction of water flow. Core 1 from the patterned area of the northern patterned fen, Cores 2–6 from the southern patterned fen, Cores 7 to 8 and 11–12 from areas surrounding the southern basin, Cores 9–10 from surrounding areas of the northern fen, and Core 13 from the western moss-graminoid area. McClelland Lake on right of image, Athabasca River present beneath insets flowing to the NNE, Fort Hills Upland Complex adjoins the Wetland to the south with clear-cut upland forest evident to the SE of the Wetland. Inset map of Canada and Alberta * = location of study area.
Located on the northern slopes of the Fort Hills and associated with a fluvial-aeolian sand plain to the north, the McClelland watershed is 330 km2 in extent and consists of (1) McClelland Lake, (2) the McClelland Wetland complex located west of the lake (studied here), (3) a mosaic of upland forests of Picea glauca, Pinus banksiana, and Populus tremuloides, and (4) a number of large wetlands along the northeastern and southern margins of McClelland Lake. The Fort Hills Upland Complex is a thrust moraine of displaced blocks of pre-exiting sediments and bedrock that forms a prominent physiological feature rising about 50 m about the surrounding glacial till plain and provides calcareous groundwater discharge to the watershed. The McClelland Wetland complex is comprised of a number of wetland site-types. These various site-types are organized into complex patterns on the wetland landscape and together occur as areas that exhibit specific ecological and hydrological characteristics. These site-types can be described as follows.
Modern wetland vegetation
Northern patterned fen (136 ha – N in Figure 1). This area is characterized by moderate-rich fen chemistry and vegetation indicators (see Vitt and House (2021) for terminology and peatland classification), with well-organized strings and flarks. Water flows into the area from the northwest, flows eastward, and exits the wetland through one northern outlet (east of arrow at “1”) to McClelland Lake. This site-type grades sharply in chemistry and vegetation into the southern patterned fen to the south through a low water divide characterized by poorly organized strings and flarks (Vitt et al., 2022).
Southern patterned fen (576 ha – S in Figure 1). The area is a moss-dominated extreme-rich fen occupying the central portion of the complex. Characterized by large flarks to the east, with these becoming smaller in width and well-organized to the west along an elevational increase. At the western end of the patterned area, the flark/string pattern becomes oriented east/west indicating water flow from the southern boundary of the wetland complex (w in Figure 1) and directed along the main water track. Additionally, water flows eastward from a second source located to the south of the main water track (just NW of “8” in Figure 1). Strings are dominated by larch (Larix laricina) in the eastern portion of the area and by shrub birch (Betula glandulifera) to the west. Water exits the area to McClelland Lake mostly through one southeastern outlet (east of arrow at “2”).
Moss/graminoid fen (213 ha – G in Figure 1) with high water levels, no patterning, and relatively shallow peat depth (<1.5–2.0 m), occurs as a large homogenous region in the northwest portion of the wetland where it grades into a shrubby rich fen to the south. The northwestern moss/graminoid area is dominated by the moss, Scorpidium scorpioides and the graminoid, Carex lasiocarpa. The northwestern area is dissected by a sigmoid, linear, upland sand ridge. Water levels are highest along the southern edge of the ridge and drier along the northern side, suggesting water flow from the south and blockage along the ridge (pers. observation, D. Vitt).
Larix-dominated rich fens (662 ha – L in Figure 1) consist of areas of the wetland complex bordering the patterned fens to the west, north, and south and dominated by uniform open forest, grading to shrubby fen to the northwest. Mineral islands are present in the northern portion.
Permafrost/bog/fen/swamp complexes (1084 ha – P in Figure 1) Three extensive areas are located along the southern boundary of the wetland and also present to the northeast of the northern patterned area. Areas in this zone are a complex of bogs with and without permafrost, most of which has recently thawed and are currently regenerating with a variety of woody vegetation components. Peat plateaus are present to the southeast interspersed with Larix fen and extensive thaw in some areas. Graminoid-dominated areas, some with pooled water, indicate water movement through saturated peat.
Riparian swamp forests
Climate
The climate of the region is subhumid and continental (Little-Devito et al., 2019), with warm, short summers and long, cold winters. The annual mean monthly temperature 0.2;°Ca, ranging from −19.6°Ca in January to 16.8°Ca in July; annual precipitation is 429 mma, with 56% falling during the May-August growing season. Potential evaporation is 529 mmb and relative humidity 70%b (a 1919–1943 data from the Fort McMurray climate station, and 1944–2019 data Fort McMurray Airport and b from the Fort McMurray airport, 1953 to 2019).
Methods
Core extraction and analyses
We investigated development of McClelland Wetland through macrofossil profiles from 10 long cores (Figure 1, Table 1). In 2017, one long core was extracted using a 4 cm diameter MacCaulay peat corer 0.5 m in length (Core 5, Figure 1). Core material was carefully moved to 4 cm diameter, longitudinally split, PVC pipe and wrapped with plastic wrap. During the winter of 2017 three cores were extracted using a Sonic corer; however, these were allowed to dry out and were not useable for macrofossil analyses. Core logs from these three cores (Cores 11 to 13) were used to assist in elevational and depth profiles (Figure 1). In 2018, 2019, and 2020, nine 15 cm diameter cores were extracted using a sonic corer and shipped to Southern Illinois University (SIU) where they were stored at 2°C until analysis. These nine cores plus core 5 taken in 2017 were used for the macrofossil analyses and radiocarbon dates.
Elevations, peat depth, and locations for the 13 long cores extracted from the wetland in 2017 to 2020.
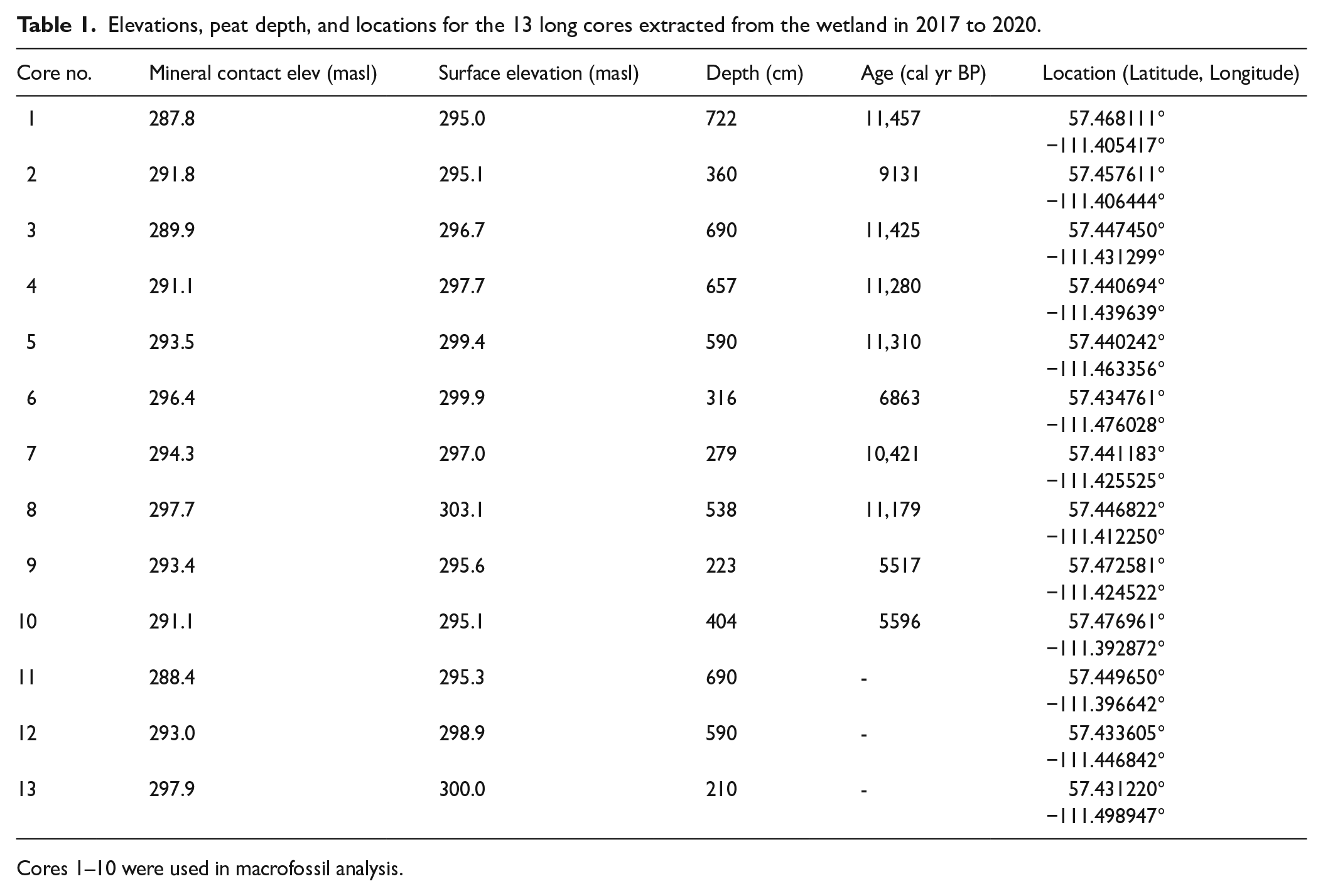
Cores 1–10 were used in macrofossil analysis.
Cores were sampled in 4 cm intervals by removing small (ca. 1–2 cm3) samples, placed in plastic bags, and all material identified in a three-step process. (1) Samples were removed by forceps, placed in a petri dish, and an estimate (to 5%) made of quantity of material identifiable by structure and unidentifiable material (debris); (2) samples were gently rinsed through a 0.5 mm sieve (tea strainer) and structural components estimated, (3) after structural percentages were estimated, percent of all bryophyte species were identified as percent of total bryophytes. Abundances of individual bryophyte species were quantified by multiplying % bryophytes found in step 2 by % in step 3. A list of all structural components and bryophyte macrofossils identified is given in Supplemental Table 1, available online and abundances of all macrofossils in each of the 10 cores are provided in Supplemental Figure 1, available online. Samples of bryophytes or wood were extracted for 37 AMS radiocarbon dates (Table 2) from points along the cores having structural changes and from the basal organic matter, and calibrated for calendar years BP (cal yr BP – Reimer et al., 2009).
Radiocarbon dates (as calibrated (or calendar) years before AD 1950 – cal yr BP) from McClelland Wetland.

AMS=DirectAMS, 11822 North Creek Parkway North, Suite 107, Bothell, WA 98011, USA. All calibrations were based on intCal109 calibration dataset (Reimer et al., 2009).
Long-term organic matter accumulation (LTAR)
We regressed peat depth against age using calibrated radiocarbon dates from 10 cores extracted from both the patterned areas and the surrounding wooded areas spanning the entire date range of 11,457 cal yr BP. Additionally, we assessed long-term accumulation rates for individual cores using calibrated radiocarbon dates and core depth profiles. Long-term accumulation rates were calculated for the entire length of each core, given as “depth.”
Results
Core 1-Northern patterned fen (Figure 2)
Basal date 11,457 cal yr BP, depth 722 cm, mineral contact at 287.8 m elev.
LTAR (long-term accumulation rate) = 0.63 mm yr−1
This core came from a flark located in the center of the northern patterned area (Figure 1). The organic material at 730 cm depth (dated at 11,457 cal yr BP) contained abundant woody twigs with lesser amounts of graminoids, and no bryophytes. Just above this organic stratum, a 24 cm wide zone (with a duration of 231 years) of sandy mineral sediments occurred. These zones suggest a moist post-glacial landscape with at first shrubs, succeeding to a wet meadow. The first peat-forming community (at 11,226 cal yr BP) was dominated by graminoids with shrubs and few bryophytes, suggesting a wet meadow environment formed on wet sand. High percentages of debris suggest relatively high rates of decomposition and limited peat accumulation. About 1000 year later, eutrophic bryophytes became dominant (Calliergon giganteum and Drepanocladus aduncus) indicating a marsh or young rich fen community. At about 10,000 years, Hamatocaulis vernicosus became dominant, but with some remaining shrub presence. This species is a key indicator of moderate-rich fen conditions as found at the site today and indicates succession to rich fen habitats. At about 8500 years cal yr BP shrubs decrease and Scorpidium scorpioides is abundant for a short period, along with H. vernicosus. This latter species remains dominant throughout the core until the present time. The continued abundance of H. vernicosus indicates a stable water regime with little variation and little change for the past 9000–10,000 years (Figure 2).
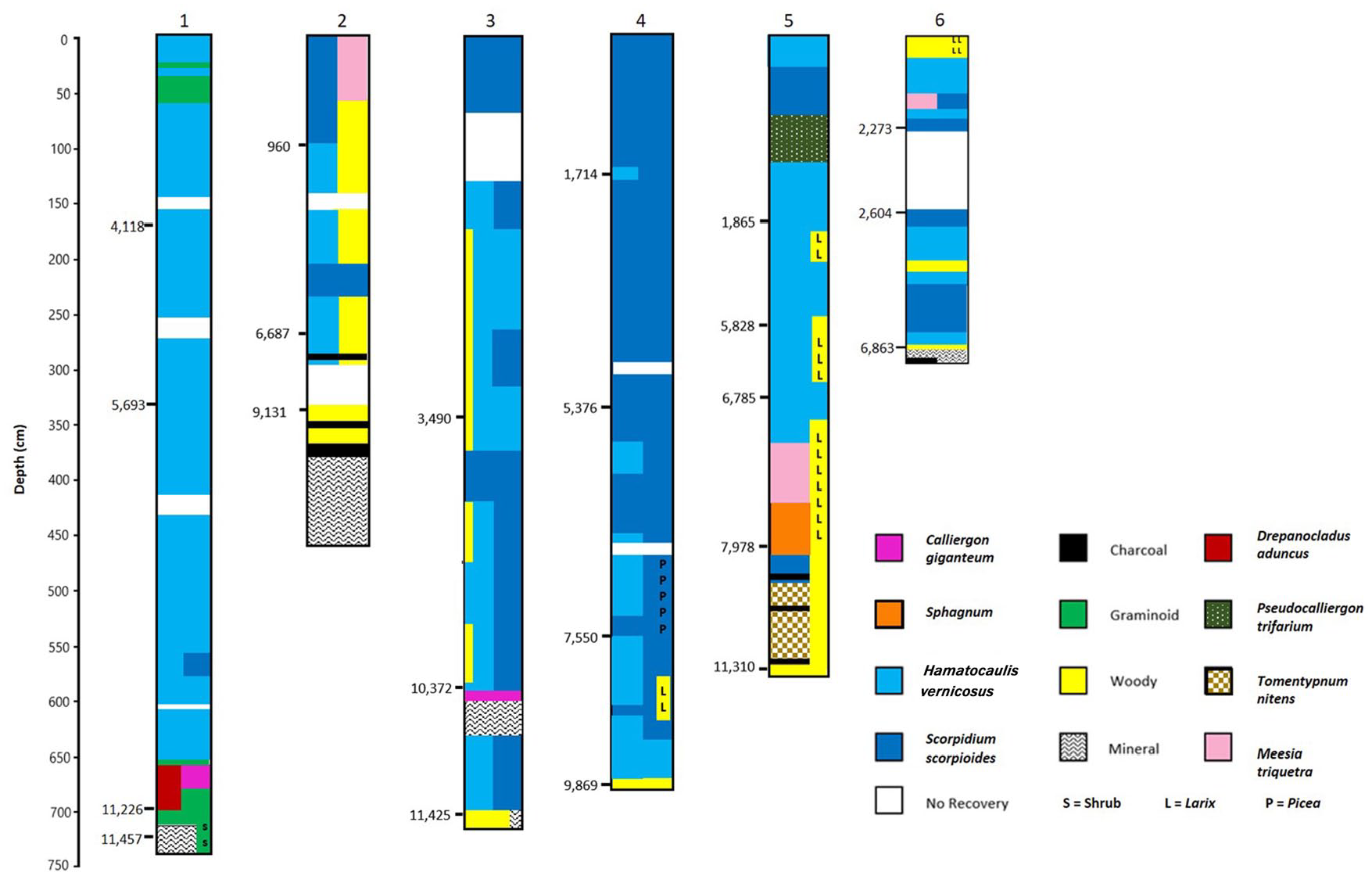
Summarized profiles of core lithologies for Cores 1–6. Dominant macrofossil components color coded. Radiocarbon dates shown on left of profiles (cal yr BP). Profiles based on data in Supplemental Figure 1, available online.
Core 2-Southern patterned fen (Figure 2)
Basal date 9131 cal yr BP, depth 352 cm, mineral contact at 291.8 m elevation, LTAR = 0.39 mm yr−1
This core was extracted from a medium-sized flark located in the southern portion of the southern patterned area, and just west of the very large flarks bordering McClelland Lake (Figure 1). The core is 360 cm long, with an additional 90 cm of sand mixed with small amounts of wood. At 430–440 cm and 342–368 cm and at the peat-mineral interface (360 cm) several charcoal layers are present. Just above the peat-mineral interface, ectomycorrhizal roots and wood are present. No bryophytes occur in the core until 300 cm (but note that there was no recovery between 304 and 338 cm). The peat/mineral interface is at 291.8 m elevation, 4 m above the surrounding mineral surface of the basin. Peat initiation began at 9131 cal yr BP, 2326 years later than at Core 1. These data indicate the presence of a mineral rise (4 m) above the surrounding basin that for 2000 years contained upland woody vegetation. At the peat/mineral interface, the lack of bryophytes and very few graminoids, coupled with wood and ectomycorrhizal roots indicate a wooded landscape. At about 7500 cal yr BP Hamatocaulis vernicosus along with graminoids increase. Wood and twigs continue to be abundant in the core until about 500 cal yr BP. At about 5000 cal yr BP, Scorpidium scorpioides and Pseudocalliergon trifarium increased in abundance, decreasing a short time afterward, replaced by Hamatocaulis vernicosus. At 960 cal yr BP, S. scorpioides becomes dominant and woody materials decrease to the surface (Figure 2). Organic matter accumulation rates were relatively low for the first 8000 years at 0.30 mm yr−1, increasing to 1.04 mm yr−1 in the last 960 years correlating to the increase in S. scorpioides.
Core 3-Southern patterned fen (Figure 2)
Basal date 11,425 cal yr BP, depth 718 cm, mineral contact at 289.5 m elevation, LTAR = 0.60 mm yr−1
This site is situated near the middle of the southern patterned area, about 1.9 km west of Core 2 (with the mineral contact 2 m lower in elevation) and 0.9 km east of Core 4 (with the mineral contact 1.2 m lower in elevation) in an area of medium-sized flarks (Figure 2). Initiation of organic matter accumulation began at 686 cm depth and 11,425 cal yr BP on wet sand. Woody vegetation and graminoids were present just before and after initiation of peat with continued presence until 610 cm depth. At 674 cm, both Scorpidium scorpioides and Hamatocaulis vernicosus are present and remain frequent until 606 cm. At 610 cm, continuing until 586 cm, sand dominates the core with no organic matter present until at 582 cm and dated at 10,372 cal yr BP. Calliergon giganteum with ectomycorrhizal roots dominate this transition, indicating a zone with woody vegetation. Evidence of shrubs and woody vegetation remain present until 180 cm. At about 86 cm (but with no recovery between 86 and 152 cm), S. scorpioides becomes dominant with no evidence of woody vegetation. Between 560 and 150 cm, H. vernicosus and S. scorpioides are variably dominant with S. scorpioides dominant for a period between 416 and 368 cm depth (at about 4000 cal yr BP for 500 years). Organic matter accumulation during the first 5700 years was relatively slow at 0.17 mm yr−1, and relatively fast in the last 4600 years (at 1.04 mm yr−1). Overall, this site developed from a shrub-dominated (perhaps with scattered trees) area to an open, moss-dominated, wet flark. The 20 cm mineral band at 586–610 cm depth is worth noting as a unique mineral incursion (Figure 2).
Core 4-Southern patterned fen (Figure 2)
Basal date 11,280 cal yr BP, depth 657 cm, mineral contact at 291.1 m elevation, LTAR = 0.58 mm yr−1
The site of this core is 0.9 km SW of Core 3 (with the mineral contact 1.6 m higher in elevation) and 1.1 km east of Core 5 (with the mineral contact 2.4 m lower in elevation), in a flark on the south side of the patterned area (Figure 1). Organic matter initiation began at 11,280 cal yr. BP and 657 cm depth. Picea needles, wood, and abundant twigs are present in the core at this depth, with bryophytes (Hamatocaulis vernicosus and Scorpidium scorpioides) evident at 648 cm. Hamatocaulis vernicosus and then together with S. scorpioides dominate the core until 528 cm (and 8372 cal yr BP), with brief evidence of Picea (at 576–580 cm) and Larix (at 568–572 cm). At 8372 cal yr BP an abrupt shift occurs with bryophytes decreasing and Picea and woody components increasing, remaining conspicuous in the core until 468 cm (about 7400 cal yr BP). From this depth until the present, S. scorpioides is the dominant component in the core, with several occurrences of co-dominance with H. vernicosus (Figure 2). The rate of peat accumulation in the first 5054 years was 0.63 mm yr−1, compared to a slightly lower rate of 0.54 mm yr−1 for the last 6226 years. In summary, this site paludified quickly at 11,280 cal yr BP to a moderate-rich fen, underwent a dry treed phase beginning at 8372 cal yr BP lasting until about 7400 cal yr BP, after which moss-dominated wet conditions prevail until the surface.
Core 5-Southern patterned fen (Figure 2)
Basal date 11,310 cal yr BP, depth 590 cm, mineral contact at 293.5 m elevation, LTAR = 0.52 mm yr−1
This core came from the northern edge of the patterned area, 1.1 km northwest, with the mineral contact 2.4 m higher in elevation than Core 4 and 1.24 km east, with the mineral contact 2.5 m lower in elevation than Core 6 (Figure 1). Peat formation began at 11,310 cal yr BP (at 549 cm depth) associated with abundant macrofossils of Tomentypnum nitens, Picea, wood, and charcoal, indicating a paludifying black spruce wooded fen with a hummocky ground layer. Charcoal is abundant in the samples from 543 to 429 cm, with wood abundant from initiation to 339 cm – dated at 6750 cal yr BP. Transitions in the early core were from Tomentypnum nitens, to Scorpidium scorpioides, to Sphagnum warnstorfii at 7978 cal yr BP, indicating at first treed conditions, then open conditions, and back to treed habitats. Larix is present from 446 cm to 354, 299–263, and 191–171 cm in low abundance. At about 400 cm depth, Sphagnum species abruptly decrease and species of open flark conditions increase, notably Meesia triquetra, followed by Hamatocaulis vernicosus and Pseudocalliergon trifarium at 339 cm and 6750 cal yr BP. Wood is abundant in the core until 339 cm and decreases upward, and after from 425 cm to the surface H. vernicosus and herbaceous macrofossils dominate the core (Figure 2). Organic matter accumulated at a rate of 0.31 mm yr−1 for the first 3332 years, then increased to 0.86 mm yr−1 during the period of Sphagnum presence and transition to wet conditions, followed a rate of 0.50 mm yr−1 for the last 6750 years. In summary, organic matter began accumulating in a paludifying forest at 11,320 cal yr BP and continued until approximately 7460 cal yr BP when trees decreased and rich fen bryophyte species became more abundant. From about 5800 cal yr BP until the present, species characteristic of very wet rich fens (including flarks) are variously abundant.
Core 6-Western edge of southern patterned fen (Figure 2)
Basal date 6863 cal yr BP, depth 316 cm, mineral contact at 296.4 m elevation, LTAR = 0.46 mm yr−1
The site of this core is 1.24 km west of core 5 with the mineral contact 2.9 m higher than the elevation of core 5. The site is located just west of the north/south patterned area, in an area of indistinct flark/string patterning (Figure 1). The surface elevation of 300 m is slightly higher or the same as areas to the west and east, suggesting a shallow drainage divide. Peatland initiation occurred at 6863 cal yr BP at a depth of 316 cm. At 318 cm depth charcoal macrofossils are present within a matrix of sand. Wood is present at 318 cm and Larix needles at 314–310 cm, and twigs are abundant from 318 to 314 cm, all suggesting an area of scattered shrubs and Larix trees. Bryophytes are present at 318 cm and again at 314 cm continuing upward to the present. Hamatocaulis vernicosus is abundant from 310 to 298 cm, replaced by Scorpidium scorpioides from 294 to 254 cm. At 246 through 190 cm depth a drier period is indicated by increases in shrub and root components. From 190 to 72 cm, H. vernicosus and S. scorpioides occur in alternating bands, and from 38 cm to the surface Larix was present on the site with drier conditions. The large portion of the core with no recovery makes detailed accumulation rates uncertain, but the rate of 0.46 mm yr−1 (based on the depth of 316 cm) is just below the overall fen average. This site initiated on an upland landscape with woody vegetation with an increase in Larix abundance, and quickly paludified to rich fen conditions. The site maintained wet, rich fen habitats varying in dryness and chemistry over the past 6000 cal yr BP, becoming drier with increases in Larix over the past 900 years.
Core 7-Southern marginal treed fen (Figure 3)
Basal date 10,421 cal yr BP, depth 279 cm, mineral contact at 294.3 m elevation, LTAR = 0.27 mm yr−1
The core site is a wet area within forest dominated by Larix laricina, south of the main patterned area. The area surrounding the site appears to be a part of inflow through the upper peat column from a north-flowing stream and separated from the patterned fen by an area of wooded fen. Peat initiated about 1000 years after earliest initiation occurred in the central patterned area. From the mineral contact at 279 to 243 cm depth (dated at 7632 cal yr BP), the core contains sandy mineral material mixed with wood and Larix needles indicating a Larix-dominated swamp. During this 2789 year period, bryophytes were scarce and the rate of organic matter accumulation was very low (at 0.13 mm yr−1, accumulating only 36 cm of peat). From 243 cm to the surface, the core contains numerous Larix needles, abundant wood, and periodic bryophyte indicators of wooded conditions (Calliergon richardsonii, Tomentypnum nitens). Periodic wetter conditions (indicated by increases in Hamatocaulis vernicosus) occur at 203–112 cm and at 50–30 cm depth. The relatively high peat/mineral contact elevation – 3.2 m higher than core 4, and 4.5 m higher than core 3, both within the patterned area, coupled with the early basal date (10,421 cal yr BP), and very low rates of organic matter accumulation all indicate a marginal (ecotonal) site influenced by seepage water from inflowing streams, with wooded conditions continually present at the site for the entire Holocene.
Core 8 Southern marginal treed fen (Figure 3)
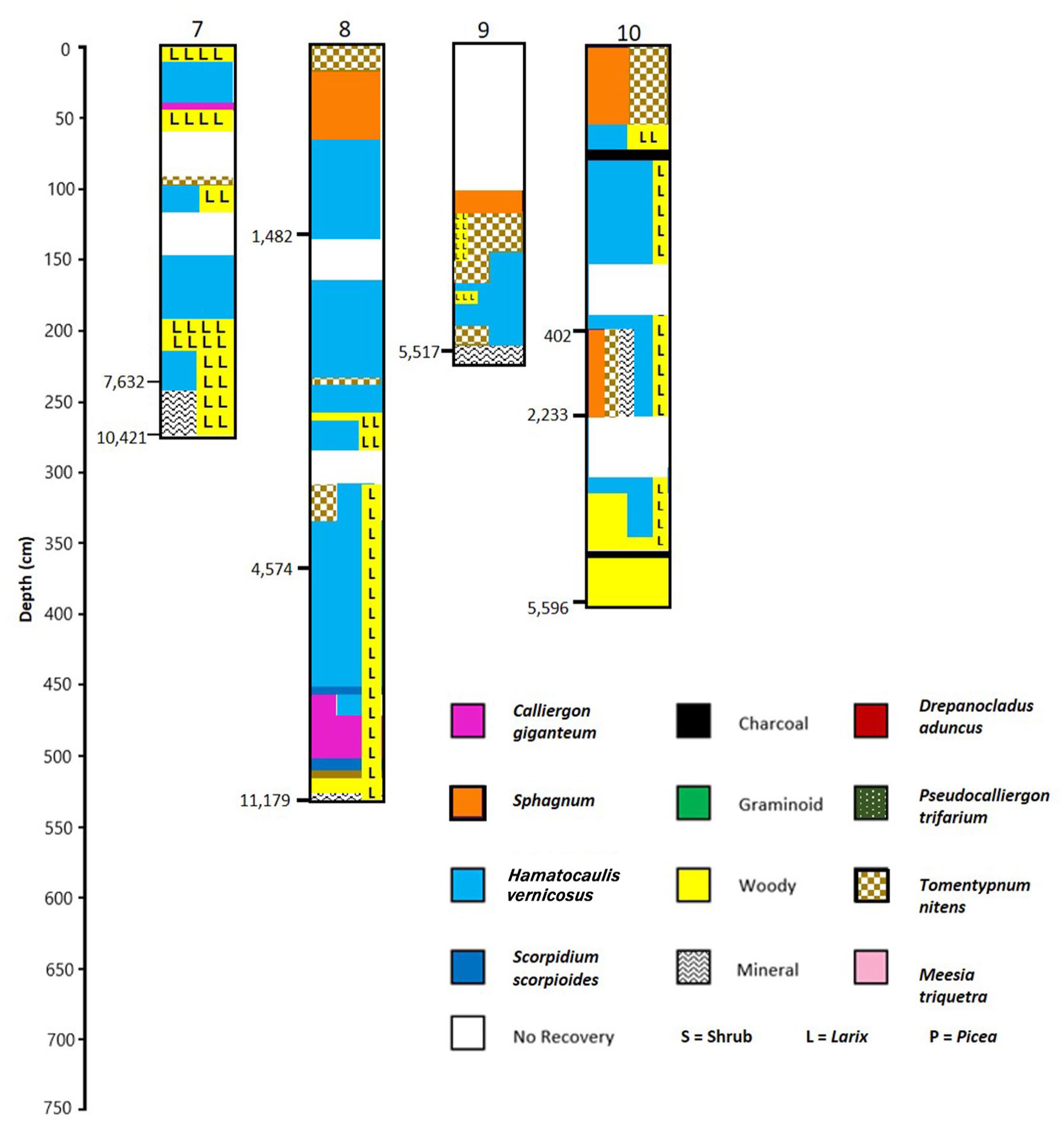
Summarized profiles of core lithologies for Cores 7–10. Dominant macrofossil components color coded. Radiocarbon dates shown on left of profiles (cal yr BP). Profiles based on data in Supplemental Figure 1, available online.
Basal date 11,179 cal yr BP, depth 538 cm, mineral contact at 297.7 m elevation, LTAR = 0.48 mm yr−1
The core was extracted from the edge of the western-most bog island on the southeastern side of the patterned area. No permafrost thaw is seen on Google images at the coring site. The mineral/peat contact is between 3.4 and 9.3 m higher than the two nearest coring sites. The current surface elevation is likewise between 6.1 and 7.8 m higher than the surrounding coring sites. Peat accumulation rates at the site have gradually increased, with an accumulation rate of 0.21 mm yr−1 during the first 6600 years, followed by a rate of 0.83 mm yr−1 for the next 3000 years and the past 1500 years had a rate of 0.96 mm yr−1. Earliest organic matter accumulation developed in a wooded habitat with both Picea and Larix needles present, along with the mosses Calliergon giganteum and Tomentypnum nitens. Larix macrofossils and wood were continually present throughout most of the core. Picea macrofossils appear abundantly in the core at 274 cm depth (about 3000 cal yr BP) and remain frequent in the core until 198 cm, and then become dominant in the core at 82 cm. About 750 cal yr BP, Sphagnum fuscum becomes dominant. This site began as a wet swampy Larix-dominated forest that accumulated peat at a low rate for the first 5000 years or so. With gradual peat accumulation the site developed into a wooded (Larix) rich fen. The presence of Picea macrofossils at about 3000 years indicates development of a wooded poor fen or bog. The recent occurrences of Sphagnum fuscum indicates ombrotrophy, either the result of permafrost development in the past ~800 years or dry raised peat surfaces. An ash layer at 26 cm and the change from S. fuscum to S. teres in the subsequent samples indicates a wildfire on site with increases in wetness and minerotrophy.
Core 9 Northern marginal treed fen (Figure 3)
Basal date 5517 cal yr BP, depth 223 cm, mineral contact at 293.4 m elevation, LTAR = 0.40 mm yr−1
This location of this core is just west of a bog island, with the peat/mineral contact at 293.4 m elevation, 5.6 m above the central portion of the northern patterned area (core 1) and near the source of the surface water flow into the northern fen area. The top 100 cm of the extracted core is missing. The site paludified at 5517 cal yr BP with abundant Larix. At 189 cm, Larix presence is reduced and Hamatocaulis vernicosus abundance increases suggesting wetter conditions. At 161 cm, the hummock-forming moss, Tomentypnum nitens, is co-dominant with H. vernicosus, and at 150–118 cm, Larix abundance increases and T. nitens becomes the dominant moss indicating drier conditions. At 114–106 cm, Sphagnum fuscum is dominant. Picea, although sporadically present throughout the core, is never abundant and probably never was present at the site, but occurred nearby. This site has remained a moderate-rich fen dominated by Larix for its duration; however, a pronounced wet period was present between 185–153 cm depth (approximately 1900 and 800 cal yr BP).
Core 10 Northern marginal treed fen (Figure 3)
Basal date 5596 cal yr BP, depth 404 cm, mineral contact at 291.1 m elevation, LTAR = 0.72 mm yr−1
This site is 0.76 km NNE of the edge of the northern patterned area in a Larix-dominated moderate-rich fen. The peat/mineral boundary is 3.3 m elevation above that of core 1 in the patterned area and 2.1 m lower than core 9 at the western edge of the northern patterned area. Two broad areas of no recovery in this core prohibit determining accumulation rates within the core, but the mean accumulation rate is among the highest recorded in the fen (0.72 mm yr−1). Organic matter accumulation began at 5596 cal yr BP (at 404 cm depth) with abundant wood and Larix needles. Bryophytes become abundant at 376 cm (mostly Hamatocaulis vernicosus). Larix remained abundant throughout the core, with sporadic occurrences of Picea. Charcoal occurs in most samples up to 60 cm depth, with large pieces especially noticeable at 384, 336, 104, 76, and 68 cm depth, indicative of local site wildfires. Tomentypnum nitens and Sphagnum are present in some abundance beginning at 270 cm, continuing to 210 cm, associated with sand grains, then with H. vernicosus until 68 cm, and at 60–12 cm, both species dominate the bryophyte fraction. From 270 to 210 mineral materials are present (1–20%). This site paludified in Mid-Holocene and remained a wet, wooded, rich fen until 60 cm depth when drier, more oligotrophic conditions prevailed.
Long term apparent organic matter accumulation rates (LTAR)
When 30 radiocarbon dates obtained from 10 cores are regressed against peat depth, there is an overall significant positive correlation between age and depth of organic matter (linear model R2 = 0.793 and polynomial model R2 = 0.806 (Figure 4). The linear model provides a mean long term accumulation rate of 0.557 mm yr−1), varying from 0.27 to 0.72 mm yr−1. This rate agrees well with that reported by Bauer et al. (2003) for a fen complex near Calling Lake AB, wherein they reported a range of accumulation rates from 0.29 to 0.94 mm yr−1 and a mean of 0.529 mm yr−1 (from 16 cores dated between 1560 to 7440 cal yr BP).
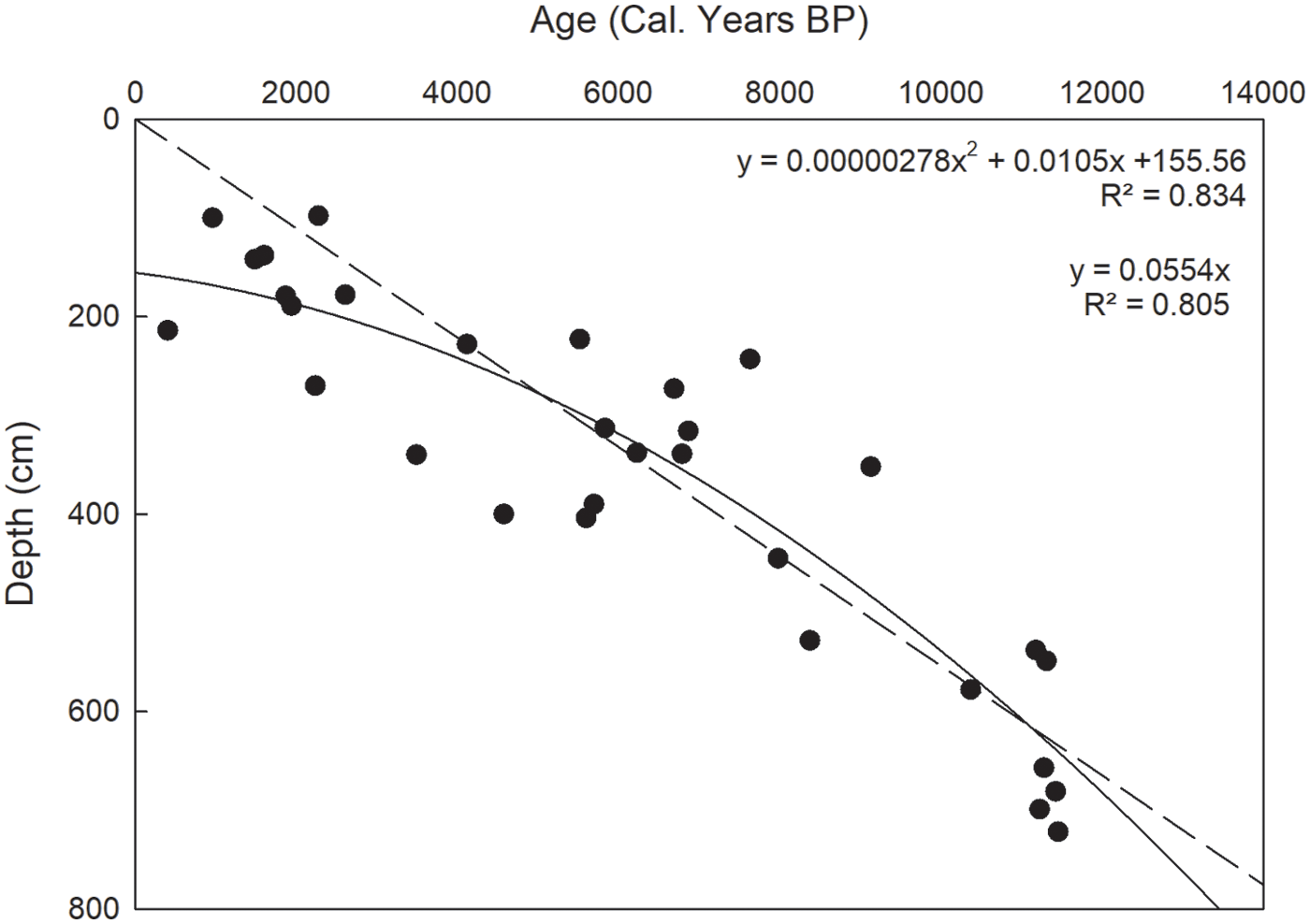
Peat depth (cm) as a function of age (cal yr BP) from 30 radiocarbon dates from 10 cores from McClelland Lake wetland Linear model R2 = 0.793, y = 0.0557x; polynomial model R2 = 0.806, y = 130.06e0.0001x.
Cores 1, 3, and 4, all located in the patterned fen areas with high moss abundance and early basal dates, have high accumulation rates. Cores 2, 6, 7, and 9, all with abundant woody materials indicating paludified, wooded sites, have lower accumulation rates. Except core 2, located on a mineral rise in the patterned fen, all of the other cores with woody materials are in located in present day wooded fens. Core 7, with the lowest accumulation rate (0.27 mm yr−1), stands out and is unique in its marginal landscape position with almost the entire core length containing abundant macrofossils of Larix. In contrast, core 10 had the highest accumulation rate (0.72 mm yr−1). This is true even if a zone with a mineral intrusion is subtracted from the total core length. Comparing mean accumulation rates for cores 1–6 and 13 along the central patterned axis (Figure 5) reveals higher accumulation rates eastward at sites at lower elevations (except core 2 located on a mineral rise). As a result, the 10.1 m initial mineral substrate gradient between core 13 and core one (over the 7.75 km) has been reduced by half, to a gradient of 5.0 m, over the 6800–11,000-plus years of organic matter accumulation.

Longitudinal section for McClelland patterned fen through Cores 1–6, and 13 along the elevational gradient from west (on the left) to east (on the right). Total distance is 7.75 km. Age, Depth, and LTAR (long term apparent rate of organic matter accumulation) shown at bottom. Approximate organic matter accumulated by depth shown by colors for three interpolated ages (4000; 7300; and 11,300 cal yr BP). McClelland Lake is located to the right (east) of the profile.
Discussion
Developmental history of McClelland Wetland
The first evidence of organic matter accumulation at McClelland wetland is present at 11,200–11,400 cal yr BP, nearly synchronous with deglaciation, the opening of the Athabasca River northward drainage, and the Lake Agassiz Clearwater River flood. Soon after deglaciation, the lower portions of the McClelland fen basin (at elevations from 287.8 to 293.5 m) appear to have been a wet sandy landscape with shrubs and graminoids – seemingly a wet meadow with organic matter accumulation commencing from primary peat formation. At somewhat higher elevations, larch (Larix laricina) and black spruce (Picea mariana) were present. At a minimum, 4.5 km of the fen westward from the lake began organic matter accumulation before 11,000 cal yr BP, with all sites except that at Core 1 developing from and having woody vegetation on site, most sites with abundant larch. By 10,000 cal yr BP, these wooded sites had developed to moss-dominated moderate-rich fens. Paludification of the surrounding landscape continued until at around 6000 cal yr BP and sites to the west, north, and south of the peatland were accumulating peat at low rates from swampy larch woodlands. After 6000 years, these marginal sites maintained tree cover and today are either black spruce or more commonly larch-dominated rich fens. By 10,000 cal yr BP the dominant bryophyte species present today were present on site, with sites to the north dominated by Hamatocaulis vernicosus and those influenced by the southwestern water inflows dominated by Scorpidium scorpioides. These two species have continued to dominate to the present time.
Regional development of peatlands in the Early Holocene
In general, deglaciation of the northeastern part of Alberta took place around 11,000 cal yr BP (Dyke et al., 2003); however, large scale peatland initiation from paludified landscapes did not occur until after 7000–7500 cal yr BP (Campbell et al., 1998). Fisher et al. (2009) studied the stratigraphy of lakes associated with moraines in the Fort McMurray area. The Firebag moraine lies on the Fort Hills (adjacent to McClelland Wetland) continuing westward across the present-day Athabasca River channel. North flowing waters were blocked by ice until deglaciation of the moraine and opening of the drainage northward to the Arctic Ocean via the Mackenzie River that Fisher et al. (2009) determined took place 11,250 cal yr BP. Glacial Lake Agassiz discharged meltwater eastward (into the St Lawrence River) until 11,450 cal yr BP when this outlet closed. Subsequently, at 11,275 cal yr BP an earthen drainage divide was overtopped and incised, lowering the lake by 52 m (Smith and Fisher, 1993), creating a massive discharge event with maximum volume of 22,000 km3 that was discharged over a 1.5–3.0 year period and followed by continued flow until 10,750 cal yr BP.
First recognized as early as 1896 in Sweden by Gunnar Andersson, and well-documented cold climatic anomaly, the Younger Dryas, occurred from 12,900 to 11,600 cal yr BP, with abruptly warmer temperatures afterward (Carlson, 2013). Following the Younger Dryas, at 11,300 cal yr BP a brief (150–350 years) cooling event (the Preboreal Oscillation [PBO]) has been widely recognized. Fisher et al. (2002) argued that the close association of Lake Agassiz flood dates and opening of the northward drainage to the Arctic Ocean with the beginning of the PBO provide compelling arguments for the cause of this short cooling event. This cooling event is synchronous with the initiation of organic matter accumulation and the presence of peatland vegetation at McClelland Wetland.
The Early Holocene climate in northern and central Alberta was warmer with summer solar radiation reaching a maximum about 11,000 cal yr BP (Pisaric et al., 2003) and drier, with lowered lake levels suggested by diatom assemblages from 9200 to 8100 cal yr BP recorded at Otasan Lake (66 km WNW of McClelland Wetland – Prather and Hickman, 2000).
Following deglaciation at Mariana Lake (178 km south of McClelland Wetland at 55°57′N), Hutton et al. (1994) described early vegetation as dominated by Artemisia and Gramineae, with limited abundance of arboreal species, and recorded abundant Sphagnum spores in their lake core, with these spores suggesting the presence of peat-forming wetlands in local area between ca. 11,200 and 10,200 cal yr BP.
Between ca. 11,400 and 8500 cal. yr BP, spruce forest, including both Picea mariana and P. glauca, began to dominate the MacKenzie Basin (MacDonald, 1987) and Larix pollen first appears at Mariana Lake between ca. 8400 and 10,200 cal yr BP.
At about 10,000 cal yr BP, Sphagnum disappears, reappearing in the core at ca. 6900 cal yr BP, increasing in abundance to the present. At 8300–6200 cal yr BP, Populus reached its maximum Holocene occurrence. Beginning around 7300–6800 cal yr BP, peatlands increased, with extensive paludification after 5700 cal yr BP (Hutton et al., 1994). In concordance, at an associated large complex peatland at Mariana Lake, early peatland formation from lake infilling was present at 9100 cal yr BP, and continued until 8000 yr BP, with portions of the landscape paludifying by 7300 cal yr BP. By 4800 cal yr BP, organic terrain had extended to one-third of the present-day peatland. Extensive paludification began around 5700 cal yr BP (Nicholson and Vitt, 1990). Differentiation of bog islands interspersed with fen water tracks were evident at about 5000–5700 cal yr BP. Basal dates from a large poor fen near to the Mariana Lakes site studied by Nicholson and Vitt (1990) have basal dates ranging from 5910 to 7260 cal yr BP (5150–6310 RC yr BP – Yu et al. (2014)). These vegetation changes occurred simultaneously with increased aridity between ca. 10,000 and 5700 cal yr BP (Ritchie, 1976; Viau and Gajewski, 2009).
Peatland initiation and development in boreal western Canada
A global postglacial ~1450-year climatic periodicity has been recognized (Bond et al., 1997). In continental western Canada, the ~1450-year periodicity has been identified as wet and dry cycles in Late-Holocene sediments (Campbell et al., 1998). Additionally, Campbell et al. (1998, 2000) proposed for southern Alberta a paleoclimatic model with 19 wet/warm periods at ~1450 year intervals over the past 15,000 years, including wet periods at about approximately 8500, 10,000, and 11,500 years (cal yr BP). Periodicity in peat accumulation rates and ash-free bulk densities were found to be associated with wet periods of 1400–1600 year duration at 6900, 5500, and 4000 cal yr BP at a rich fen in western Alberta (Yu et al., 2003), and other fens in other parts of the province (Yu et al., 2014). The wet events were contemporary with warm periods in the North Atlantic (Bond et al., 1997), probably in response to solar activity (Yu et al., 2003). Associated with these functional changes, basal dates (71) from paludified peatlands across continental western Canada show the same regularity at a millennial time scale (Campbell et al., 2000). Most peatlands initiated during wet periods, especially at 7000, 5200, and 3800 cal yr BP with a 400–500 year time lag from the beginning of wet events at 7500, 5600, and 4200 cal yr BP. This lag may be due to sampling bias by incorporating younger peat into compacted basal samples for dating. Somewhat fewer initiation events are also present at 8500 cal yr BP.
Summary – McClelland Wetland: A unique early peatland
In summary, it appears that at around 11,000 cal yr BP peatlands, including Sphagnum-dominated ones, were present, but uncommon across the northern boreal areas of western Canada (Halsey et al., 2000). These decreased or were eliminated between 8300 and 6200 cal yr BP, with peatlands restricted to small areas surrounding bodies of water, and only after 7000 cal yr BP did extensive paludification occur. Differentiation of peatland landforms (bog islands, water tracks) were secondary features developing only after 5000–5700 cal yr BP.
Few peatland initiation events have been recorded before 8500 cal yr BP, with basal dates (71) from paludified peatlands across continental western Canada exhibiting a regularity at a millennial time scale (Campbell et al., 2000). Most peatlands initiated during wet periods, especially at 7000, 5200, and 3800 cal yr BP apparently corresponding to warm events in the North Atlantic (Yu et al., 2003).
A few initiation events prior to 10,000 cal. yr BP are known from northern Alberta, including at Rainbow Lake in northwestern Alberta (10,230 cal yr BP) where peat accumulated in depressions (Bauer and Vitt, 2011); however, widespread peatland expansion in the area occurred only after 8000 cal yr BP (Bauer and Vitt, 2011). This early peatland initiation is in contrast to sites farther south where Bauer et al. (2003) reported the earliest dates for peat accumulation in the Athabasca area of the province were at around 7000 cal yr BP.
McClelland wetland – A wetland resistant to allogenic change
Over the 10,000 year history of McClelland Wetland, the central portions of the wetland have been remarkably resistant to change, with little alteration in dominant species. The resistance to change is set against a background of fluctuations in regional Holocene climate and local varying water balance (reconstructed from 18O preserved in moss cellulose - Gibson et al., 2022). These dominant bryophyte species continued to play a foundational role on site for the duration of the Holocene responding to persistent long-term ground water sources. Accumulation rates have remained steady (or decreased somewhat using a polynomial model) once a dominant bryophyte layer was established, but with accumulation rates at paludifying marginal sites lower than those of the central moss-graminoid-dominated areas. The greater organic matter accumulation present in the central portion of the wetland (nearest the lake) compared to areas farther westward has decreased the elevational gradient by half, increasing the occurrence of ponding in the largest flarks. The resistance to changing Holocene events, together with stabile ecosystem functions, provide a case study for how species in rich fens may resist change against future climate-related and anthopogenic disturbances to the surrounding boreal forest regime. This resistance to vegetation change for rich fens across the region appears to be associated with persistent long-term groundwater inflows, and suggests that surrounding watersheds are an important part of long-term survival of patterned rich fens.
Supplemental Material
sj-docx-1-hol-10.1177_09596836231169997 – Supplemental material for An 11,000 year record of plant community stability and paludification in a patterned rich fen in northeastern Alberta, Canada
Supplemental material, sj-docx-1-hol-10.1177_09596836231169997 for An 11,000 year record of plant community stability and paludification in a patterned rich fen in northeastern Alberta, Canada by Dale H Vitt and Melissa House in The Holocene
Supplemental Material
sj-pdf-2-hol-10.1177_09596836231169997 – Supplemental material for An 11,000 year record of plant community stability and paludification in a patterned rich fen in northeastern Alberta, Canada
Supplemental material, sj-pdf-2-hol-10.1177_09596836231169997 for An 11,000 year record of plant community stability and paludification in a patterned rich fen in northeastern Alberta, Canada by Dale H Vitt and Melissa House in The Holocene
Footnotes
Acknowledgements
We especially want to thank Dr Xiaoying Fan (Suncor Canada) for her encouragement and thoughtful discussions. Also, thanks to Robert McLaughlin (Suncor Canada) for providing the sonic cores. Drs Jean Birks and John Gibson (Innotech Alberta) were instrumental in coordinating our research and providing significant logical support, especially in shipping materials and help in the field. Lilyan Glaeser (SIU) and Alexandra Tsalickis (SIU) provided technical assistance in the laboratory and Neal Tanna (Innotech) and Don Jones (Innotech) provided much necessary assistance in the field. Comments from an anonymous reviewer improved the paper for which we are grateful.
Correction (October 2023):
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This project was funded by Innotech Alberta contract number C2017000679 from funds supplied by Fort Hills Energy L.P, for which we are grateful.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.