
Abstract
In this paper, we explore and integrate different currencies that may underlie large-game hunting to guide a trans-Holocene analysis of variation in artiodactyl utilization using massive archaeofaunal data-sets from predominantly open-air sites from the Bonneville and Wyoming basins. The available empirical data continue to suggest that artiodactyls yield consistently higher return rates than lagomorphs allowing us to leverage predictions from both the prey choice and energetic risk-gain models that the relative importance of artiodactyl hunting should scale closely with climate-based change in their abundance on the landscape. We document with modeled climate data that seasonal variables correlated with the relative frequency of artiodactyl hunting, but that summer temperature had a significant overriding effect in both regions. Controlling for the negative relationship between summer temperature and artiodactyl abundances, we then document enhanced artiodactyl hunting in general and bison more specifically during the Fremont period that is consistent with a costly signaling hypothesis and the unique socio-ecological conditions of this context. Thus, climatic variation and its influence on artiodactyl abundances drives the overall trajectory of Holocene large game hunting variation but measurable and more subtle influences of costly signaling are also detected.
Keywords
Introduction
Large game hunting has long been viewed as a catalyst in human evolution, driving key changes in life history, social organization, locomotion, and other aspects of human behavior and biology (e.g. Hill, 1982; Isaac, 1984; Kaplan et al., 2000). Other evolutionary models, however, emphasize the pivotal role played by subsistence resources such as geophytes or small game that provide more reliable long-term returns accessible to a wider range of human consumers (e.g. Hawkes, 1991; Hawkes et al., 1993; O'Connell et al., 2002). Against this backdrop, considerable attention has focused on Holocene variation in the relative importance of large- and small-game hunting, gender differences in foraging, and the implications of hunting for other aspects of human behavior and biology in western North America.
Based on theoretical considerations—mostly from the prey model of foraging theory—and empirical evidence that larger animals often yield higher post-encounter return rates than smaller ones, archeologists have commonly used taxonomic indices comparing the relative abundance of large to small-sized prey (i.e. Abundance Index) to measure variation in overall return rates, diet breadth, and foraging efficiency from archaeofaunas. The latter variables have in turn been theoretically linked to variation in health, stature, and weaning age; workloads and osteoarthritis; the division of labor; settlement circumscription violence and warfare; hereditary inequalities; the emergence of agriculture; and technological changes associated with the adoption of lower return food resources, to name a few (Allen et al., 2016; Bartelink, 2006; Broughton et al., 2011; Broughton and O’Connell, 1999; Cannon, 2003; Cheverko and Bartelink, 2017; Greenwald et al., 2016; Hildebrandt and McGuire, 2002; Ugan et al., 2003).
Yet, despite the widely recognized utility of foraging theory models, there remains disagreement on how to best estimate certain model parameters that are required to apply them. Notably, debate continues on how to operationalize archeologically the key prey model components of (1) prey ranks and (2) the currencies and goals that motivated past hunting behavior. These issues have been raised in research on the potential climatic influences on Holocene artiodactyl populations, including pronghorn (Antilocapra americana), bighorn sheep (Ovis canadensis), deer (Odocoileus spp.), bison (Bison bison), and elk (Cervus canadensis), in both the Great Basin and Wyoming Basin of western North America (Broughton et al., 2008, 2011; Broughton and Bayham, 2003; Byers et al., 2005; Byers and Broughton, 2004; Byers and Smith, 2007). These analyses utilized a range of proxy estimates for artiodactyl population densities (i.e. artiodactyl fecal pellets, hunting tools), with the relative abundance of artiodactyls and lagomorphs (i.e. an Artiodactyl Index (AI): ∑artiodactyls/∑ ([artiodactyls + lagomorphs]) from archaeofaunas being key among them.
Following established convention, these studies argued that insofar as higher densities of artiodactyls elevated overall returns from hunting in past landscapes, lower-return animals especially lagomorphs, are predicted to less frequently enter the targeted set of prey. It follows that increasing abundances of artiodactyls in past environments should result in proportionate increases in their hunting, so that the relative frequencies of artiodactyl skeletal specimens in archeological faunas should, other things equal, track trends in their abundance on the landscape.
Based on data related to modern climatic factors that influence artiodactyl populations in interior western North America, it was then hypothesized that past artiodactyl populations should be controlled by measures related to climatic seasonality—namely intra-annual temperature range, and summer and winter precipitation intensity. Correlations between modeled reconstructions of these climatic variables and the artiodactyl index values revealed tight fits between the deductively derived predictions and the empirical records (Broughton et al., 2008, 2011). Most notable was a consistent pattern of increasing artiodactyl densities during the late-Holocene that correlates with reduced temperature seasonality, increased summer precipitation and decreased winter precipitation (Broughton et al., 2008).
These conclusions have been challenged by concerns that the studies focused on a limited number of sheltered archeological and paleontological deposits—caves and rockshelters—that may not reflect broader patterns of subsistence, or might be compromised by taphonomic issues concerning variation in the agents responsible for bone deposition (Grayson, 2011; Grayson and Fisher, 2009; Lupo et al., 2020; but see Broughton et al., 2011; Byers et al., 2005 for analyses of open-air deposits from the Wyoming Basin and Little Boulder Basin of eastern Nevada). In addition, recent syntheses of faunal datasets from the Bonneville Basin have found no meaningful Holocene trends in the relative abundance of artiodactyls and lagomorphs calling into question the typically employed relative ranking of these taxa (Lupo et al., 2020).
Additional concerns have arisen from considerations of the factors that motivate large game hunting. Among several hunter-gatherer groups (e.g. !Kung, Hadza, Aché), large game hunting appears to have been motivated more by social attention, signaling value, and mating opportunities than by foraging efficiency and nutritional provisioning (e.g. Hawkes, 1991; Hawkes et al., 1993, 2001). Following these observations, Hildebrandt and McGuire (Hildebrandt and McGuire, 2002; McGuire and Hildebrandt, 2005) proposed that major trans-Holocene trends in the relative importance of artiodactyl hunting in western North America were driven not by climate-based variation in large game abundance, but changes in past social contexts.
To address these methodological and theoretical issues, as well as provide an update on the current evidence for climatic controls on Holocene artiodactyl abundances, we examine the available archaeofaunal data from sheltered and open sites in two regions, the “Bonneville Basin” (BB) and “Wyoming Basin” (WY) study areas (Figure 1 and Supplemental Tables 1–4). We define the former to include the Bonneville Basin proper and several well-sampled directly adjacent areas in eastern Nevada, as well as the Uinta Basin and sites located on the north slopes of the Uinta Range. Our Wyoming Basin study area includes the Wyoming portions of the Green River and Washakie basins, as well as the Great Divide Basin in its entirety.
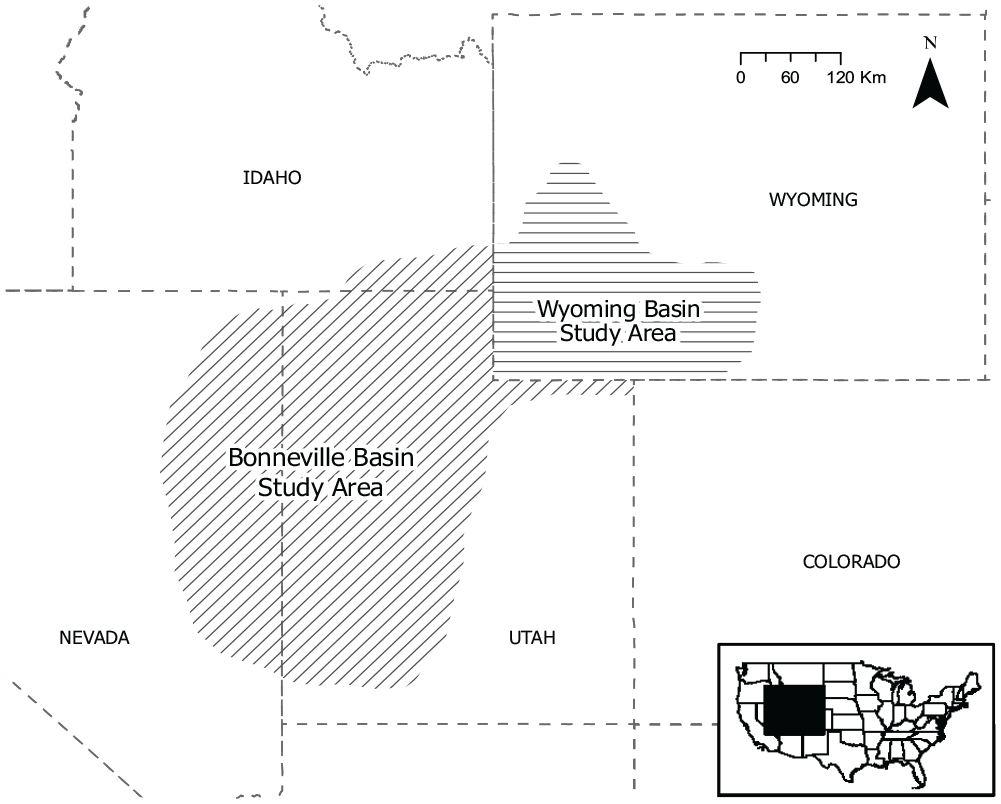
Map of the Bonneville and Wyoming basin study areas providing data for this study.
We derive artiodactyl indices from these regions and array them against modeled paleoenvironmental data from PaleoView climate models (Fordham et al., 2017) and test the previously developed hypotheses that artiodactyl abundances should correlate with indices of climatic seasonality (Broughton et al., 2008). By examining two regions that differ in elevation, latitude and baseline temperature and precipitation regimes, our tests incorporate climatic influences on artiodactyls both through time and across space. After identifying the climatic controls on artiodactyl hunting, we are then able to test to what degree costly signaling may have influenced artiodactyl hunting during the unique socio-ecological context of the Fremont period in the Bonneville Basin. Our analysis has implications for the debates related to the motivations underlying men’s hunting in past human societies as noted above. Before providing those tests, we address the general issues raised above related to prey ranks and currencies used in archeological applications of foraging theory models.
Background
Prey ranks and zooarchaeological abundance indices
Prey ranks are typically defined in prey model formulations as post-encounter return rates, measured as e/h, where e represents the net energy provided by a prey type and h represents the handling costs associated with acquiring and processing it for consumption. Of these variables, e is more readily measurable since it generally scales directly with prey body size. Handling costs, by contrast, are highly variable and context dependent. Thus, early applications, notably Bayham (1979, 1982), adopted the modeling convention where variables recognized to be potentially important—but perceived to be intractable—are held constant in order to apply the prey model in archeological settings. Body size alone was thus used as proxy for prey ranks. This formulation was developed in the context of archeological studies in the American Southwest, where artiodactyls and lagomorphs dominate faunal collections and at a time when ethnographic or experimental information on return rates was lacking. After the first wave of empirical experimental and ethnographic data on return rates began to be published, the theoretical body size-return rate relationship received considerable support (e.g. Hill et al., 1987; Simms, 1987; Smith, 1991; Winterhalder, 1981; summarized in Broughton, 1994: Table 1), although exceptions to the pattern were also documented, in many cases related to mass capture, where variation in prey density becomes an important factor (e.g. Lindstrom, 1996; Madsen and Schmitt, 1998).
Correlations between AI and seasonality variables.

Ethnographic analyses have since provided additional insight in estimating h. For example, noting that prey mobility or velocity should be a positive measure of pursuit costs insofar as fleet prey yield more protracted chases, Codding et al. (2012); see also Bird et al., 2009) suggest e/h should be assessed qualitatively on a case-by-case basis considering both prey size and velocity. Others have suggested that prey ranks should be directly discounted for hunting failure rates by multiplying e/h values by ethnographically observed success rates (Lupo et al., 2020; Ugan and Simms, 2012).
A recent synthesis of a worldwide sample of empirical data on return rates for terrestrial vertebrates highlights the importance of handling-cost variation (Morin et al., 2022). After adjusting for inter-study differences in the way foraging returns have been calculated and correcting them using a standardized formula, Morin and colleagues examine several factors, such as escape strategy, hunting technology, and prey body size, that influence return rates. Their analysis underscores earlier work documenting high variability in body fat, tissue composition, and especially, pursuit costs, which results in a significant, weak, relationship between body size and return rate at the global level (n = 131 prey types; r = 0.39, p < 0.0001).
Most critical to archeological applications, however, is less to what degree a global body-size e/h relationship may exist across prey types, but whether differences exist in the returns for the prey taxa under analysis in a specific context. Here, we focus on the artiodactyls and lagomorphs available to the Holocene foragers in western North America. To this end, we summarize the return rate data in Morin et al. (2022) for North American artiodactyls and lagomorphs (Figure 2). These data indicate considerable variation across taxa and ethnographic contexts, although the mean return rates for artiodactyls are significantly higher than those for lagomorphs (Mann-Whitney U = 71.0, p < 0.001; Figure 2a; from Morin et al., 2022: Table 1 [Published e/h]). Similar results are indicated with recalculated values using a standardized formula and omitting cases where data were obtained from recreational hunters, were incompletely documented, or include search time (Mann-Whitney U = 10.0, p = 0.035; Figure 2b; from Morin et al., 2022: Table 3 [Rederived e/h]). We emphasize that the ethnographic data incorporate time costs associated with failed pursuits so that those costs are charged and incorporated into these return rate estimates. However, it is the case that the summed mean approach from which these calculations derive does not allow an assessment of variance in failed pursuits across foraging bouts that is most appropriate in analyses focused on risk-sensitive foraging that we discuss in more detail below (Bird et al., 2009; Morin et al., 2022).
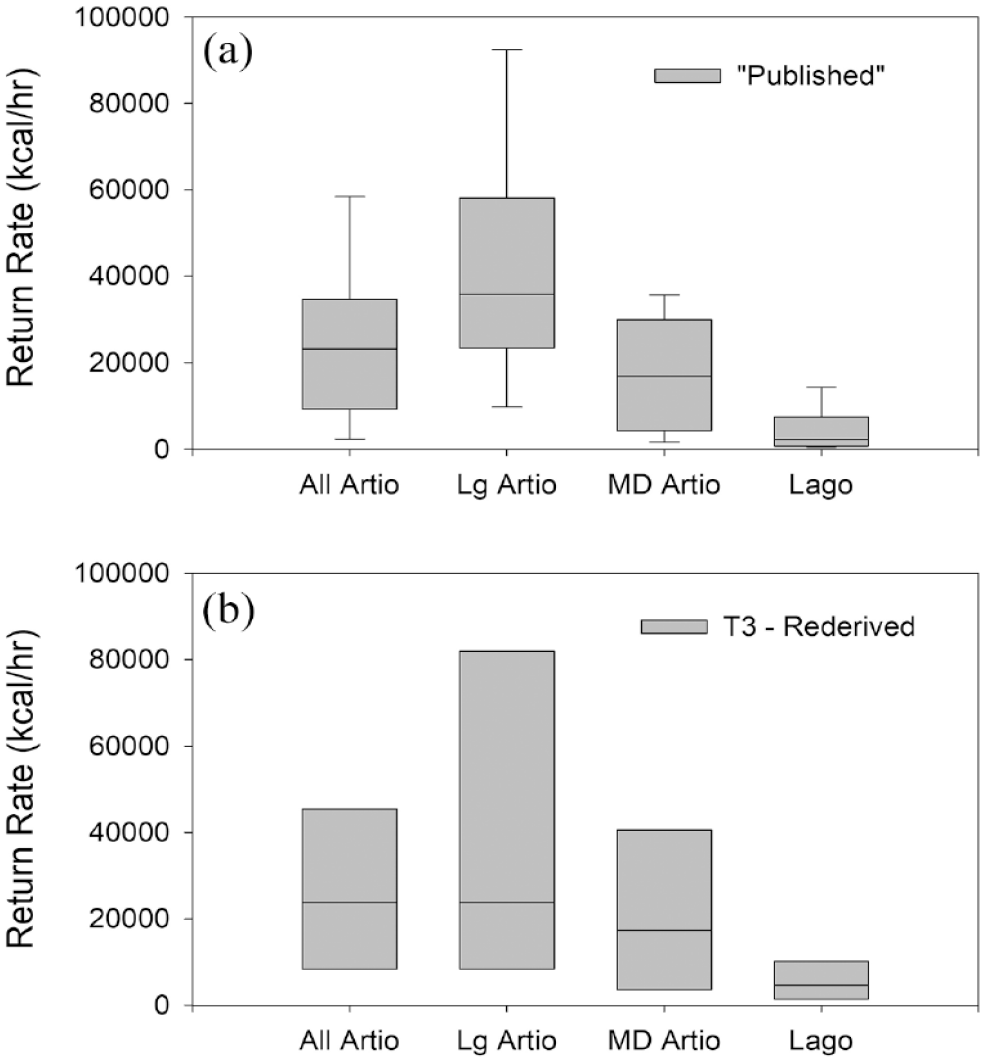
Ranges of North American artiodactyl and lagomorph return rates as provided in Morin et al. (2022). (a) Values presented in the literature (Morin et al., 2022, Table 1 [column headed “Published e/h”]). Artiodactyl taxa include moose (Alces alces), caribou (Rangifer tarandus), bighorn sheep (Ovis canadensis), mule deer (Odocoileus hemionus), white-tailed deer (O. virginianus), pronghorn (Antilocapra americana), bison (Bison bison), and elk (Cervus canadensis). Lagomorphs include snowshoe hare (Lepus americanus), black-tailed jackrabbit (L. californicus), cottontail (Sylvilagus sp.), swamp rabbit (S. aquaticus), and marsh rabbit (S. palustris). (b) Recalculated values using a standardized formula and omitting data from recreational hunters, and those that were incompletely documented, or include search time (Morin et al., 2022: Table 3 [column headed “Rederived e/h”). Artiodactyl taxa include moose (Alces alces), caribou (Rangifer tarandus), and pronghorn (Antilocapra americana). Lagomorphs include snowshoe hare (Lepus americanus), black-tailed jackrabbit (L. californicus), and cottontail (Sylvilagus sp.).
While these data are clearly imperfect, and undoubtedly variation in e/h exists among the artiodactyl and lagomorph species included in this sample, they are also consistent with the body size and velocity approach noted above—North American artiodactyls and lagomorphs, such as hares, are both exceptionally fast, yet the former are substantially larger. These considerations suggest that artiodactyls, on average, should yield higher e/h relative to lagomorphs so that their relative abundance in archeological faunas should gauge their abundance on past landscapes based on the logic of the prey model, other things equal.
Alternative models and currencies
The currency and strategic goal that anchors the prey model is the maximization of net energetic returns. However, other currencies and goals can underpin foraging behavior in general and large game hunting in particular, most notably the minimization of risk in acquiring sufficient calories and the value large prey may hold in costly signaling, prestige rivalry, and display (e.g. Bishop et al., 2023; Codding et al., 2011; Elston et al., 2014; Hawkes, 1991; Hildebrandt and McGuire, 2002; Lupo et al., 2020; Pinson, 1999; Speth et al., 2013).
Codding et al. (2011) provide a useful framework for considering how different ecological contexts may structure the prevalence of these different goals that focuses on a trade-off between “offspring provisioning” and “social provisioning.” Offspring provisioning refers to foraging geared toward the nutritional needs of dependent young and other kin. By contrast, social provisioning refers to foraging motivated by delayed benefits in kind or other indirect fitness benefits such as those derived from signaling. In the latter, fitness benefits may accrue to hunters strictly from the information conveyed about them through successful kills—“from the different ways that others treat them in light of the quality they reveal” (Hawkes and Bliege Bird, 2002). Insofar as offspring and social provisioning represent a trade-off, and men and women vary in how they calculate the costs and benefits attendant to it, gender differences in foraging may emerge (e.g. Codding et al., 2011; Elston et al., 2014; Hawkes, 1991; Smith and Bliege Bird, 2005).
Whether foragers aim to minimize the risk of daily caloric shortfalls or maximize energetic returns will often play into such calculations and Codding et al. (2011) emphasize that such decisions should vary by ecological context—most notably, the relative abundance of differently ranked resources on the landscape (Codding et al., 2011; Elston et al., 2014). Specifically, they propose that when foragers frequently encounter high-return, but high failure-rate resources, such as large game, maximizing energetic returns can satisfy both provisioning offspring and prestige-signaling goals. If so, then the sexual division of labor should converge on the pursuit of such prey, with the foraging efforts of women shifting to logistical support of men’s hunting. Such an adaptation may underlie the convergence of effort by both sexes toward hunting-related work that characterize the Aché of Amazonia and many groups in the circumpolar north (see also Waguespack, 2005). However, when encounter rates with high-variance large game decline, the division of labor should diverge, with men still pursuing risky, high-return resources, but women shifting to more reliably acquired resources with lower energetic returns such as small game or plant resources. With the latter, collective diet breadths should widen as large game abundance declines and women contribute more reliably acquired, small game. This “energetic risk-gain” model thus yields expectations that closely align with standard expectations of the prey model—the relative contribution of harvested large, high-ranked prey should scale with their natural abundance on the landscape.
Additional socio-ecological factors may, however, further alter the payoffs for offspring and social provisioning especially those that can enhance costly signaling motivations for male large game hunting. These relate to conditions that either reduce the fitness costs of forgoing the provisioning of offspring or increase the fitness value of social display.
As originally suggested by Hildebrandt and McGuire (Hildebrandt and McGuire, 2002; McGuire and Hildebrandt, 2005), increases in the productivity of female resource acquisition may increase their level of offspring provisioning and thereby reduce the opportunity costs for men to withhold it. Hildebrandt and McGuire suggested this may have occurred in late Archaic (4000–1000 BP) contexts in California insofar as thresholds of productivity were eclipsed through intensive female-generated acorn harvests. In extreme cases, this may even have allowed women’s subsistence efforts to subsidize the risk-prone foraging of men, as has been suggested to be the case at times for Martu and Hadza foragers (Codding et al., 2011).
Different socio-ecological contexts may also alter the fitness value of large game hunting as a medium for social display and costly signaling. Depending on a range of other factors such as life history stage or marital status, larger, more entwined social networks and human population aggregations should be associated with larger pools of potential recipients to receive costly displays leading to higher payoffs for such activity (Codding et al., 2011; Martin, 2019; Simms et al., 2014). Higher payoffs for display may also stem from factors that strengthen the signaling value of large game acquisition, such as when they become locally uncommon through resource depression that is also more likely to occur adjacent to higher density human settlements (Martin, 2019). Signaling strength in these contexts may also be amplified through the acquisition of especially large, risky, and/or dangerous prey types (e.g. Oommen and Shanker, 2021; Speth, 2010).
Insofar as strong climatic drivers can be documented for Holocene artiodactyl populations in our study area, signaling motivations for large game hunting should be detectable by large game hunting levels that exceed baseline trends established between artiodactyl abundance records and the climatic variables that control them. In other words, if ancient hunters acted in ways consistent with optimal foraging predictions, the abundance of large game in the archeological record should reflect their abundances on the local landscape. During times with environmental conditions favorable to large-game reproduction, game populations will increase and, subsequently, so too should hunter encounter rates and the relative representation of artiodactyls in archeological sites. Teasing out signaling from provisioning motivations is, however, difficult and the dearth of empirical tests of costly signaling speaks to this issue.
Here, we suggest that costly signaling might be identified empirically as AI values exceeding expectations based on climatic conditions. In such circumstances, the archaeofaunal record would reflect hunters who were not randomly encountering large game, but instead seeking out such resources as costly signals. In other words, optimal foraging (and risk minimizing) should reflect calorie-maximizing decisions during on-encounter hunting, whereas signaling hunting (and effectively risk maximization to maintain honest signals) should reflect targeted hunting.
Subsequently, this hypothesis provides a path to identifying signaling behaviors empirically. This means that in statistical analyses, signaling should be reflected in residuals that occur above regression lines that describe relationships between climatic drivers (e.g. summer temperature or precipitation) and Artiodactyl Index values. Additionally, such trends should be pronounced for the largest-sized artiodactyls available to human consumers (e.g. bison). As others have suggested (e.g. McGuire and Hildebrandt, 2005; Simms et al., 2014), features of the Fremont Complex may have provided a fruitful context for costly display and a useful case study to explore these issues. Given the context specific nature of prestige hunting, we only make predictions about the Fremont period. We do note, however, that the method deployed here might be used to explore other anthropogenic trends, such as resource depression, in the archeological record. Exploring every deviation from the best fit line, is, however, beyond the scope of this paper and we limit our focus to identifying signaling behaviors in the Fremont record.
Regional settlement histories, the Fremont Complex, and costly signaling
The two study areas highlighted in this research each display very different settlement histories and we leverage these differences to explore the influence of centralized villages on the motivations behind large-game hunting. Prior to ~2000 cal BP, hunting and gathering characterizes the lifeways of both regions. After this time, however, the adaptive strategies in the two regions diverge, with village sites appearing in the Bonneville Basin, while hunting and gathering remains the primary lifeway in the Wyoming Basin, to the north. In fact, beyond a poorly understood, middle Holocene housepit phenomenon characterized by seemingly short-term occupations in low investment structures, adaptations such as sedentism, villages, or any kind of permanent structures are absent from Wyoming Basin during the Holocene (Thompson and Pastor, 1995).
In contrast, the Fremont complex is represented by a mixed farming and foraging adaptation that occurred across much of the eastern Great Basin and Colorado Plateau between ~1950 and 650 cal BP (Madsen and Simms, 1998). Importantly, the Fremont are known for villages associated with maize agriculture. Our Bonneville Basin faunal sample falls entirely within the established Fremont geographic range with 65 sites dating to this period. In addition to unique aspects of material cultural—including distinctive basketry, rock art, ceramic vessels, metates, and unfired clay figurines— the Fremont is distinguished from the preceding Archaic and subsequent Late Prehistoric groups by several features relevant to the expression of costly signaling.
First, a dramatic surge in human population densities occurs within the period to judge from radiocarbon date frequencies and other archeological indicators, with a peak in dates near 950 cal BP (Louderback et al., 2011; Madsen and Simms, 1998; Massmino and Metcalfe, 1999). And although adaptive diversity has been emphasized and a range of settlement-subsistence patterns appear to have operated within the period, the Fremont is associated with many large sedentary agricultural village sites with multiple, complex structures such as large semi-subterranean timber and adobe houses adjacent to public spaces, and large above ground rock-walled granaries (Madsen and Simms, 1998). There is also evidence of status differentiation and inequality in some Fremont sites based on spatially variable architectural features and associated artifact contents, the differential distribution of grave goods (Hockett, 1998; Janetski and Talbot, 2014; Madsen and Simms, 1998), and, in some cases, variation in the consumption of maize (Coltrain and Leavitt, 2002).
On the latter, ethnographic, historical, and experimental analyses of Fremont maize agriculture suggest that increasing investment would have produced higher productivity per unit of land, but at the expense of declining return rates—establishing the two key features of resource intensification (Barlow, 2002). Stable isotope analyses have confirmed the importance of maize for many, but not all, Fremont peoples, where it appears to have often approached a staple comparable to the role it played in the diets of Ancestral Pueblo groups to the south (Coltrain and Leavitt, 2002).
The convergence of large population aggregations with complex social contexts, and expanded productivity associated with maize agriculture, would seem to have promoted a context favorable to the expression of signaling-based hunting. But as noted above, identifying such behavior archeologically first requires a control on climate-based variation in artiodactyl abundances.
Artiodactyls and climatic seasonality
The seasonality of temperature and precipitation both condition artiodactyl abundances with the interplay between seasonal temperature and moisture influencing forage quality and free water availability, locomotion energetics, and metabolism. New plant growth provides more digestible, higher energy forage that typically emerges in wetter, cooler conditions in the spring and early summer growing season. Subsequently, the duration and quality of the growing season directly effects artiodactyl foraging success, which in turn, governs maternal condition, gestation success, newborn survival, recruitment rates and, ultimately, herd size (Brown et al., 2006; Byers and Hogg, 1995; Craine et al., 2009; Douglas, 2001; Fox et al., 2000; Gedir et al., 2015; Hamlin et al., 1982; Klein, 1965; Leslie and Douglas, 1979; Mackie et al., 1998; Peek et al., 2002; Purdue, 1989; Stephenson et al., 1985).
While artiodactyl taxa of the arid west generally prefer cool, moist conditions, high moisture levels in association with extreme cold temperatures can negatively impact foraging efficiency, locomotion, and metabolism (e.g. Axelrod, 1967; DeLgiudice et al., 2002; Mech et al., 1987; Severinghaus, 1947). Severe winter conditions regulate ungulate herds occupying temperate regions through the proximate effect of snow cover. Deep snowpacks restrict mobility and reduce food availability. Winter severity can thus determine annual weight loss and the ability of animals to recover during the rest of the year. However, in warmer settings at lower elevations and latitudes, winter precipitation and subsequent spring run-off can provide more favorable foraging conditions (Brown et al., 2003; Byers and Smith, 2007; Douglas, 2001).
Highly seasonal and unpredictable climates can also impact artiodactyls (see also Axelrod, 1967; Broughton et al., 2008; Slaughter, 1967). These factors stem from the observation that larger mammals have longer, more variable gestation periods than small ones, increasing the likelihood that favorable conditions for birthing will be “missed.” Because gestation time is typically longer than the favorable period for reproduction, large mammals must rely on correlated cues to initiate breeding (e.g. photoperiod). Correlated cues will increasingly fail, however, as the length and predictability of the favorable period decreases—features that characterize highly seasonal environments—and depressed reproductive rates follow.
While we anticipate broadly similar climatic responses to the different artiodactyl species represented in our study sample (bison, deer, pronghorn, bighorn sheep, elk), these taxa exhibit varying thresholds of toleration to specific climatic parameters (Broughton et al., 2008; Byers and Smith, 2007). Bison are most notable in this context and earlier work has documented in the Wyoming Basin that they were more sensitive to lower amplitude Holocene fluctuations in aridity compared to pronghorn (Byers and Smith, 2007)—trends consistent with modern ecological studies of these taxa (Gedir et al., 2015; Van Vuren and Bray, 1986). And in our Bonneville Basin study area, the Holocene abundance—and even the presence—of bison appears to have fluctuated dramatically in relation to effective precipitation and the extent of local grassland communities with a notable pulse that aligns roughly with the Fremont period (Grayson, 2006; Lupo and Schmitt, 1997). Over twice the size of the medium artiodactyls (pronghorn, deer and bighorn sheep), bison abundance would have potentially influenced not only overall hunting return rates but the availability of targets garnering the highest signaling values. To explore these issues, we examine separately the climatic response of bison below.
Records of paleoclimatic seasonal variation
A small body of paleoclimatic data documents season-specific variation in Holocene temperature and precipitation across western North America, with most analyses focused on summer climate. These data provide evidence for a summer thermal-maxima in the early to middle Holocene, with cooling during the Late-Holocene. Despite the general pattern, climate records demonstrate geographic variability, including regional variation in the timing of the Holocene thermal maxima. Much of the evidence for Holocene variation in summer temperature derives from pollen- and macrofossil-based vegetation records, often coupled with high-resolution charcoal sequences. These records suggest expansion of xerophytic plant communities and maximum fire frequencies during the early to middle Holocene. These patterns are consistent with high summer temperatures, intensified summer drought, low fuel moisture, and dry thunderstorms (e.g. Anderson, 1990; Barron et al., 2003; Brunelle and Anderson, 2003; Carré et al., 2021; Carré and Cheddadi, 2017; Davis et al., 1985; Davis and Moratto, 1988; Doerner and Carrara, 2001; Gavin et al., 2003; Hallett et al., 2003; Heusser et al., 1985; Long et al., 1998; Louderback et al., 2015; Louderback and Rhode, 2009; Lundeen et al., 2013; Lundeen and Brunelle, 2016; Minckley et al., 2007; Mohr et al., 2000; Smith and Anderson, 1992; Strong and Hills, 2003; Thompson et al., 2016; West, 2004; Whitlock et al., 2012; Whitlock and Bartlein, 1993).
Fewer Holocene records for the region address winter climate (but see Lundeen et al., 2013; Minckley et al., 2007; Walker and Pellatt, 2003: 541; Whitlock et al., 2012), but several documenting conditions of the early Holocene for the Great Basin are especially noteworthy. Pollen analysis from Mono Lake, in the western Great Basin, for instance, shows evidence for synchronous expansion of xerophytic vegetation and higher lake levels during the early Holocene (Davis, 1999). This apparent contradiction is interpreted to reflect insolation-driven seasonality. During the early Holocene, higher summer solar radiation increased the likelihood of droughts, while lower winter insolation produced deeper snowpacks, greater spring runoff, and higher lake levels (Davis, 1999; Whitlock et al., 2012).
While these data suggest a general pattern of enhanced seasonality and/or aridity during the early and middle Holocene they also indicate considerable geographic and temporal variability in these trends. Understanding fine-grained climatic influences on artiodactyl populations should thus be based on localized paleoclimatic information from the Bonneville and Wyoming basins in the form of trans-Holocene, time-series data. To generate this information, we use the widely-applied PaleoView climate model (Fordham et al., 2017).
Materials and methods
PaleoView
We use the PaleoView software (Fordham et al., 2017) to generate climate data for our analyses. PaleoView generates global coupled atmosphere–ocean–sea ice–land general circulation models based on daily simulations provided by the Community Climate System Model ver. 3 (CCSM3). Ground truthing the PaleoView model has demonstrated that it successfully retrodicts long-term trends in climate and associated events, such as the termination of the Pleistocene, as well as successfully simulating present-day climate patterns (Fordham et al., 2017).
PaleoView output provides simulations of temperature and precipitation at user defined temporal and spatial scales. We prompted PaleoView to provide data for 100-year periods beginning at 9500 cal. BP for latitude 37.5°–40.0°N and longitude 112.5°–115°W (west central Utah) and latitude 40°–42.5°N and longitude 110°–112.5°W (southwest Wyoming). We note that while the vast majority of sites from each study area lie within these spaces, not all do. This results from the limitations of the PaleoView software which partitions space based on 2.5° × 2.5° blocks. Increasing the climate simulation block size causes overlap of the two study areas, resulting in the homogenization of the simulated climate datasets. Annual and seasonal precipitation (mm/day) and temperature data were extracted for each region (see Supplemental Information). We define winter as December, January, February and summer as June, July, August.
Faunal data sets
The Wyoming Basin (Supplemental Material 1 and 2; see also Byers and Smith, 2007) and Bonneville Basin (Supplemental Material 3 and 4) archaeofaunas document the relative abundances of artiodactyls and lagomorphs recovered from mostly non-sheltered archeological sites. The Wyoming dataset, collated in Byers and Smith (2007), includes sites with NISP counts of 25 or more from 172 components representing over 182,000 identified specimens that span the period from 9501 to 181 years cal. BP (Figure 1). The Wyoming dataset offers an important contrast to the one from Utah—agriculture and the associated social complexity, as in the Fremont, did not emerge in the region (Kornfeld et al., 2016; Thompson and Pastor, 1995). The Bonneville Basin dataset includes sites with NISP counts of 25 or more from 129 components representing over 195,000 identified specimens that span the period 11,125–100 years cal. BP. Both datasets were calibrated using the Calib 8.1.0 software (Stuiver and Reimer, 1993) and the Intercal20 calibration curve (Reimer et al., 2020).
To make the faunal data consistent with the PaleoView output, we created 700-year running averages for AI values incremented every 100 years and beginning at 9500 cal. BP from each dataset. We chose the 700-year interval for two reasons. First, this bin size serves as a compromise between displaying as much short-term variability as possible, while at the same time averaging samples large enough to clearly demonstrate overall patterns. Second, we have used this bin size in previous studies (e.g. Broughton et al., 2008; Byers et al., 2005; Byers and Smith, 2007) and wish our results to remain internally consistent. We also observe that patterns in the spatial distribution of prey can alter encounter rates (e.g. Broughton, 2002), our regionally expansive data sets nest within the recognition that our method estimates the on-average encounter rates across each region within a given time frame.
Regardless of the methodological rational, the patterns in our data appear robust. For example, we reapportioned the Wyoming Basin data into more-narrow 300-year bins and compared it to the 700-year bin dataset. In this case, the two datasets correlate significantly (rs = 0.839, r < 0.001). Re-binning the Bonneville Basin data into 300-year bins resulted in 10 bins containing no data, yet the remaining data correlate significantly with the 700-year bin dataset (rs = 0.862, r < 0.001). These comparisons suggest that in both cases, the patterning in the data appears robust, and does not change significantly with modifications to our analytical methodology.
Finally, we evaluate relationships between climate variables and artiodactyl abundance by using the R statistics package (R Core Team, 2021) to calculate tests for difference, correlation analyses, and a generalized linear model (GLM). Because AI ranges from 0 to 1, we rely on logistic regression models with a binomial distribution, logit link, and quasi-likelihood estimation. This quasibinomial GLM accounts for the overdispersion present in our data (Wood, 2017).
Artiodactyl abundance indices
The Artiodactyl Index we use here maximizes the available faunal data in our study areas. It is important to understand that in this study, we are not attempting to measure diet breadth per se, but instead use prey model logic to track the abundance of artiodactyls on ancient landscapes. Regardless, the AI lends itself best to large datasets of often fragmentary materials generated by a range of analysts and that often identify specimens simply to ordinal-level taxa (e.g. “artiodactyl,” “small mammal” or “lagomorph”) too general for commonly-used ecological diversity measures. Many other specimens are identified to the medium artiodactyl category that includes pronghorn, deer, and bighorn sheep or a large artiodactyl category that includes bison. Our AI thus incorporates specimens identified as “artiodactyl,” “medium artiodactyl”, and “large artiodactyl,” as well as specimens identified to specific artiodactyl species or genera. To explore the influence of climatic variation on bison and control for the influence of this taxon in the Fremont context, we examine a Bison Index (∑bison + large artiodactyls/[bison + large artiodactyls + medium artiodactyls]), and a Medium Artiodactyl Index (∑medium artiodactyls/∑ ([medium artiodactyls + lagomorphs]), in addition to the Artiodactyl Index defined in the introduction.
Results
Holocene artiodactyl abundance and climate change
The PaleoView simulation retrodicts several climate trends in our study areas. First, these results suggest that during the Holocene, annual precipitation was greater in the Wyoming Basin and temperature was higher in the Bonneville Basin (Figure 3a and b). While these annual scale trends are important (Byers and Broughton, 2004; Byers and Smith, 2007), here we focus on how seasonal variation in climate conditioned artiodactyl populations. While Wyoming had moister winters than Utah and summer precipitation differed little between the two regions, precipitation for either season lacks any obvious, long-term trends across the Holocene (Figure 3c and d). Temperature variation, however, decreases across the Holocene, with colder winters and hotter summers giving way to more equable temperatures during the later Holocene (Figure 3e and f).
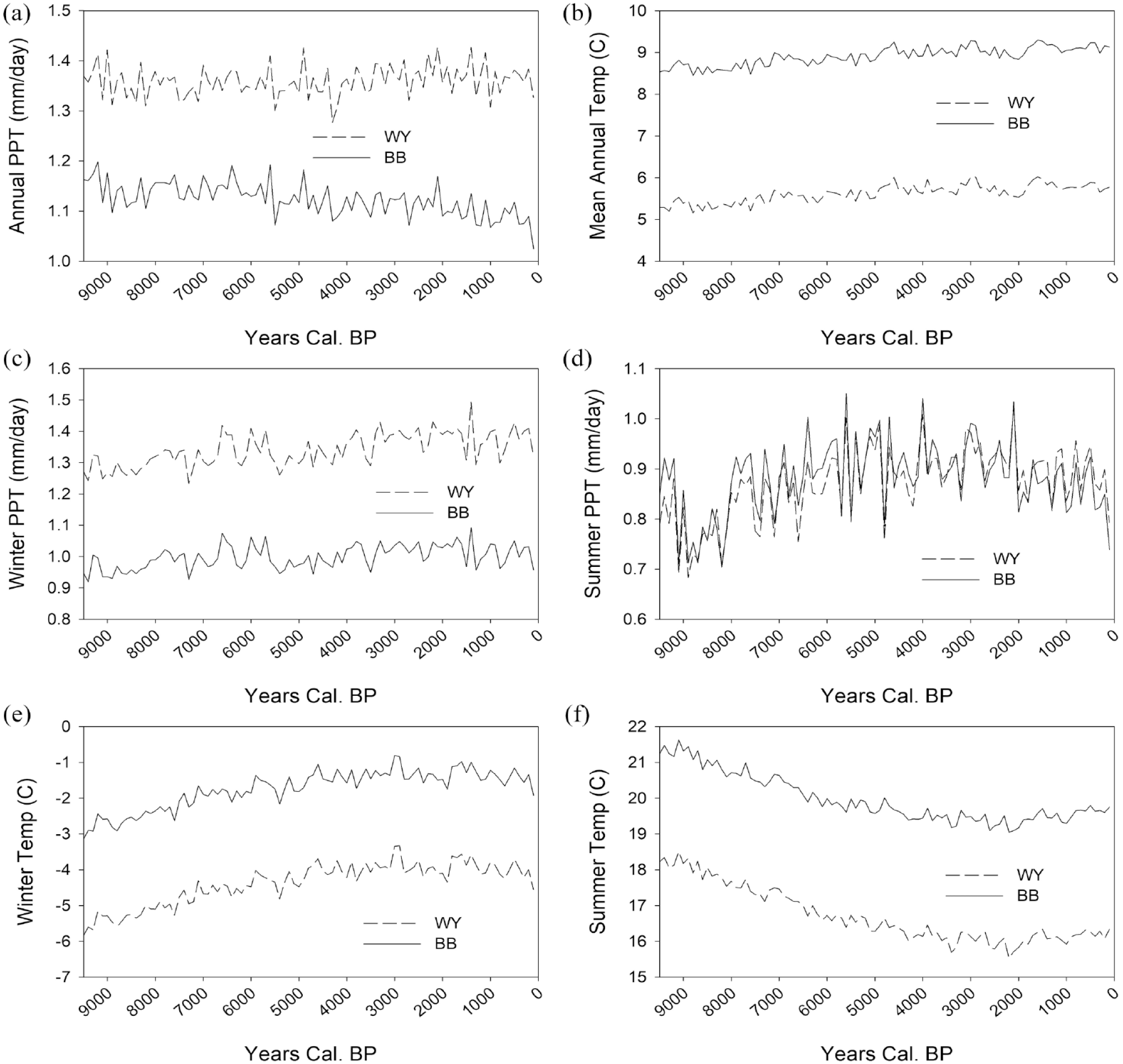
PaleoView climate simulations for the Bonneville and Wyoming basins: (a) mean annual precipitation, (b) mean annual temperature, (c) winter precipitation, (d) summer precipitation, (e) winter temperature, and (f) summer temperature.
These paleoenvironmental data, when linked with the regional archeological datasets and identified relationships between artiodactyl reproductive ecology and climate patterns, provide a basis for predicting relationships between climate and artiodactyl population histories in the Wyoming and Bonneville basins. Overall, these data suggest that the higher seasonality and warmer, dryer conditions of the early/middle Holocene would have limited artiodactyl populations. Conversely, the more mesic and less seasonally variable climate of the late-Holocene would have provided an environment favorable to artiodactyl herd growth. The result would have been a higher density of artiodactyls on the late-Holocene landscapes of Utah and Wyoming—but always higher in the latter at any given point in time. Each of these general trends are reflected in the data (Figures 4 and 5).

Holocene trends in artiodactyl indices (AI) for the Bonneville and Wyoming basins.
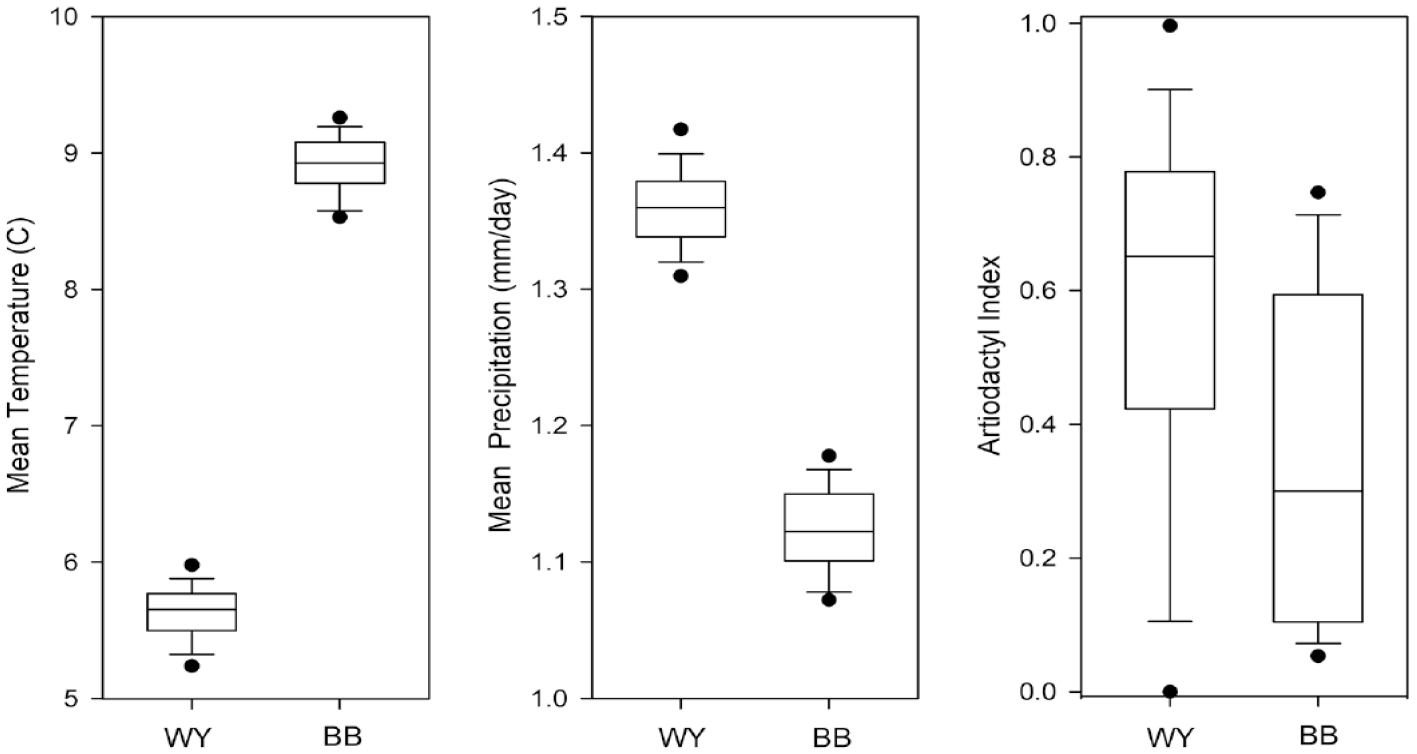
Comparison of temperature, precipitation and artiodactyl abundances for the Bonneville and Wyoming basins. Filled circles represent the 5th and 95th percentile.
Figure 6a–h displays the AI values for both areas arrayed against the PaleoView datasets for seasonal precipitation and temperature. Both datasets exhibit rank-order correlations between each of the seasonal climate variables and the artiodactyl records (Table 1). These relationships show negative correlations between AI and summer temperature, and positive correlations between winter temperature and both winter and summer precipitation. As expected, the highest AI values associate with times of lowest temperature seasonality (Figure 7).
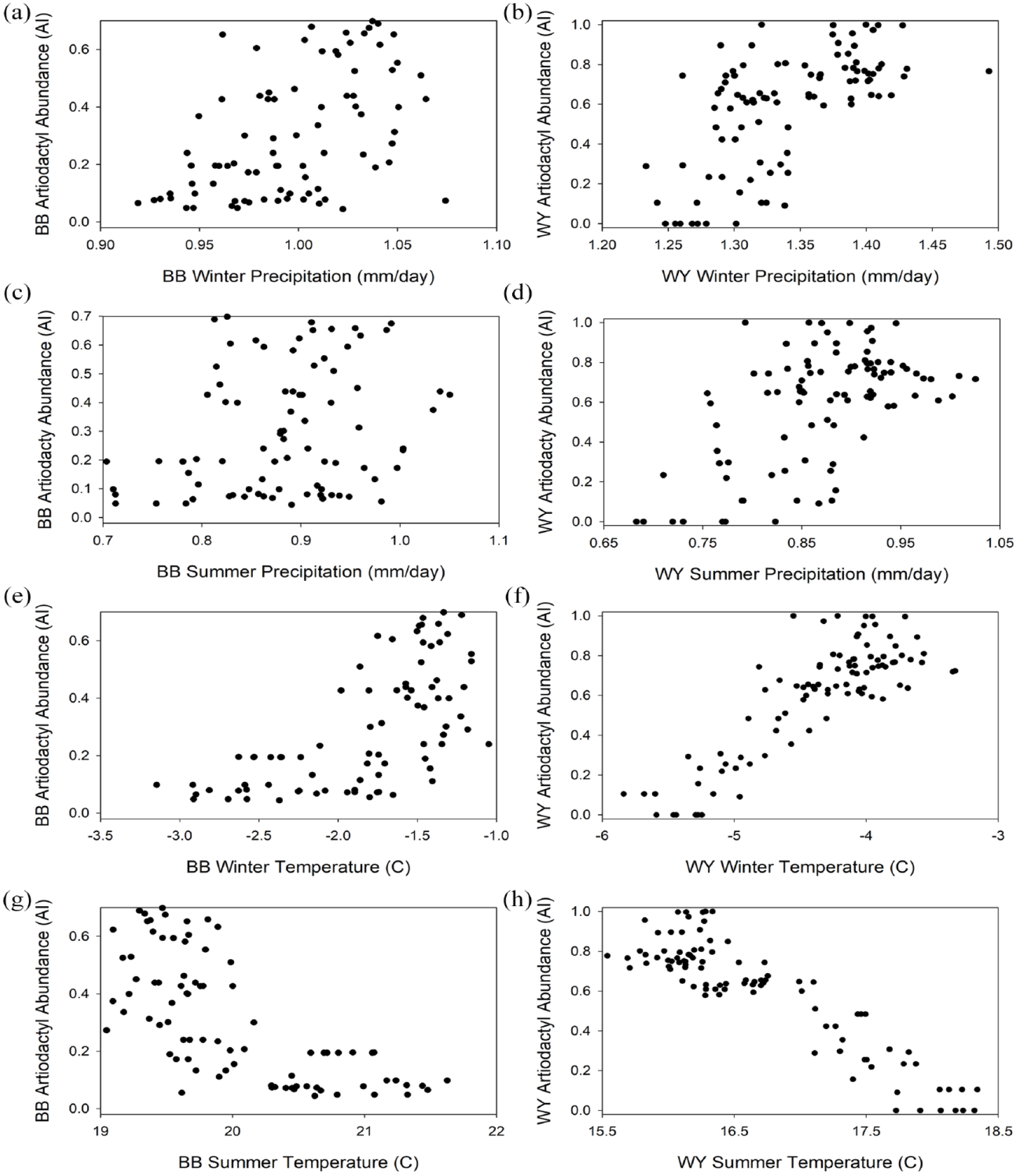
Artiodactyl indices (AI) arrayed against, winter precipitation (a, b), summer precipitation (c, d), winter temperature (e, f), and summer temperature (g, h).

AI values as a function of seasonal relationships with temperature for our study areas. AI values above and below the median value were plotted with different symbols to identify under which climate regimes artiodactyls were most abundant.
To determine which climate variables best explain patterning in the artiodactyl records, we fit a multivariate logistic regression model using a generalized linear model. Regressing the Bonneville Basin AI values on the Bonneville Basin PaleoView data reveals that summer temperature is the only significant predictor variable (β = −1.27; p < 0.001; Supplemental Table S1). The model performs well (pseudo-r2 = 0.58), with residuals slightly skewed and the ACF plot indicating some temporal patterning out to four lags (400 years). Similar relationships exist between the Wyoming Basin datasets. Here again, only summer temperature is included in the model as a significant predictor variable (β = −1.41; p < 0.001). The Wyoming Basin model also performs well (pseudo-r2 = 0.76), displaying evenly distributed residuals and the ACF plot with some temporal patterning out to three lags (300 years). Finally, the combined model which incorporates state as an interaction term again indicates that summer temperature best explains AI, although the model fits differ by state. The rate of decline for AI as summer temperature increases is faster in Wyoming than it is for the Bonneville Basin study area (β = −1.52; p < 0.0216). A combined model demonstrates the greatest predictive power of the three (pseudo-r2 = 0.80). Here too, model residuals are evenly distributed and the ACF plot indicates some temporal patterning in the residuals out to four lags (400 years). In all cases, as summer temperature increases, AI declines, suggesting that summer temperature, by itself, best explains artiodactyl abundance in both study areas.
Signaling-based large game hunting in the Fremont?
As noted above, the Fremont period in the Bonneville Basin would have provided contexts conducive to prestige rivalry and costly display in which the motivation for large game hunting could be expected to shift to optimize those goals. The signaling hypothesis thus predicts that Fremont faunas will display levels of artiodactyl hunting greater than predicted by the relationship between Artiodactyl Index values and climatic variation.
Given that summer temperature is the only significant climatic driver, we examine the relationship between this variable and the AI for the Bonneville Basin again with a generalized linear regression model, this time with special attention to the Fremont period (Figure 8a). The data document that Fremont period AI values fall consistently above the regression line, indicating that during this time, more artiodactyls are present than would be predicted by the relationship with summer temperature. In fact, the mean standardized residual for the 14, 100-year bins we associate with Fremont lies 1.34 standard deviations away from the best fit line. Fremont faunas are thus consistent with the expectation that some portion of large game hunting was motivated by costly signaling.
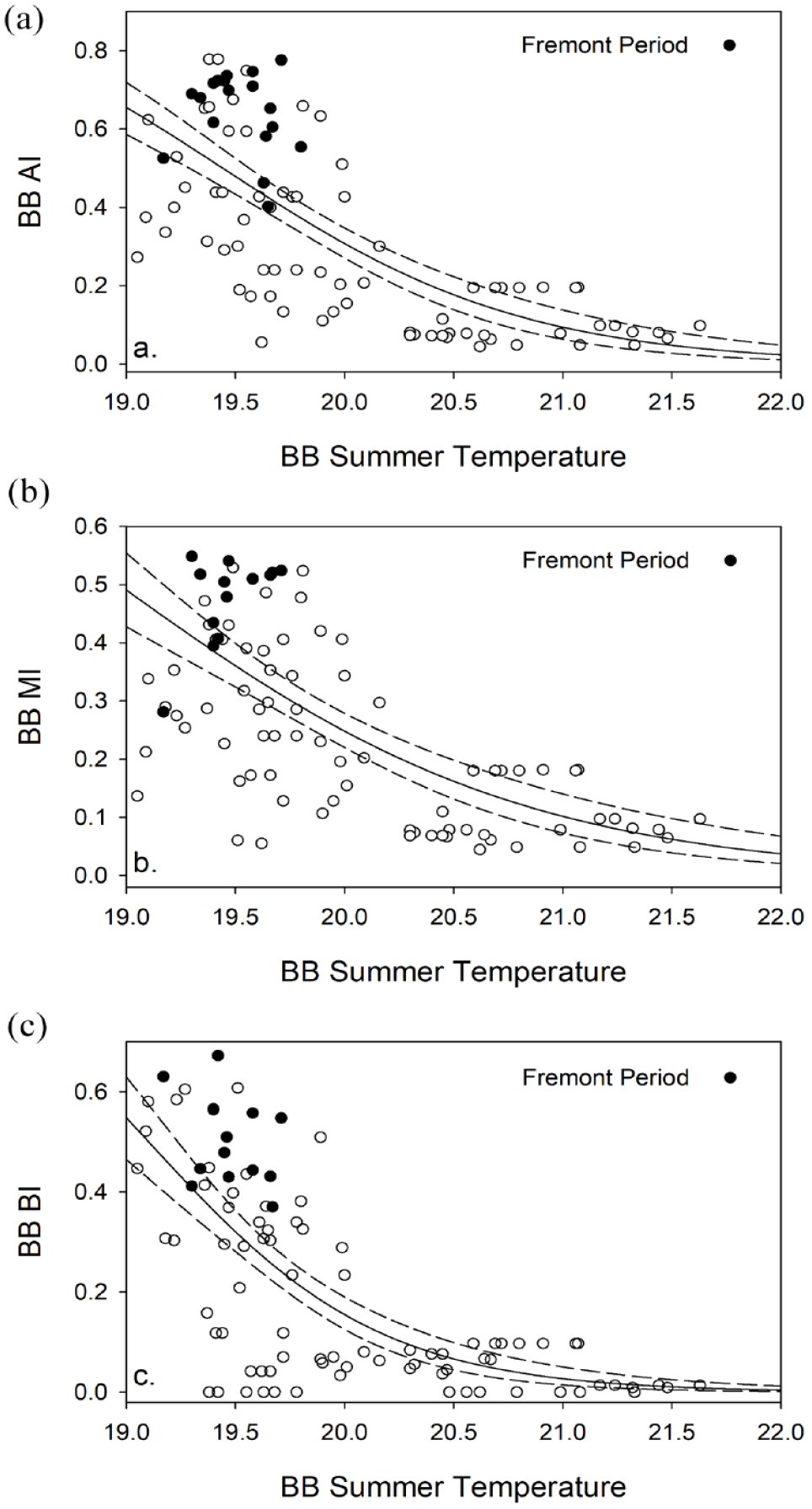
The generalized linear model regression relationships for summer temperature and artiodactyl indices for the Bonneville Basin; circles represent the Fremont period: (a) AI = Artiodactyl Index (artiodactyls/(artiodactyls + lagomorphs), (b) MI = Medium Artiodactyl Index ((∑medium artiodactyls/∑ ([medium artiodactyls + lagomorphs]), and (c) BI = Bison Index (∑bison + large artiodactyls/[bison + large artiodactyls + medium artiodactyls]).
Variation in the abundance of bison does not exert a detectable influence on this pattern as a similar result is indicated when bison specimens are excluded from the equation (mean standardized residual = 1.16; Figure 8b). Moreover, controlling for summer temperature, bison exhibit higher abundances in Fremont sites (mean standardized residual = 0.98)—relative to the medium artiodactyl taxa—than would be expected given the prevailing summer temperature conditions of the period (Figure 8c). Thus, both artiodactyls as a group and bison more specifically are better represented in Fremont sites with the driving variable of summer temperature controlled.
Conclusions
Our analysis documents strong relationships between Holocene seasonal climatic variation and artiodactyl hunting in the Bonneville and Wyoming basins. These trends follow from the prey model and the energetic risk-gain model, as they both predict that proportional increases in artiodactyl hunting will track climate-driven changes in their abundance. Unlike earlier work, this analysis employs a vast sample of open-air assemblages, limiting the incorporation of materials from non-human agents and the ambiguities of analyses focused on sheltered contexts. This analysis also represents the first bi-regional comparison of the impact of climate on large-game populations and demonstrates that the two adjacent regions exhibited similar variability in artiodactyl hunting through time, each showing substantial increases across the Holocene. Notably, the hunting of artiodactyls was consistently higher in the Wyoming Basin at any point in time owing to the cooler summer temperatures that characterized the region.
Estimating resource rankings continues to be a significant challenge to applying foraging models in archeological settings. However, we emphasize that such estimates are testable in empirical applications and can be evaluated through comparisons between deductively derived predictions and the empirical record—in this case correlations between climatic variation and the proportional hunting of artiodactyls and lagomorphs. The close fits between the predictions stemming from two separate foraging models in which artiodactyls are positioned as higher ranked than lagomorphs, suggests this placement approximates the returns experienced by past foragers of the Bonneville and Wyoming basins.
While our analyses show that each climatic variable exhibits significant rank-order correlations with artiodactyl abundances, linear modeling that controls for variable interaction effects and collinearity indicates that summer temperature, by itself, best explains artiodactyl abundance. This result suggests that while factors such as summer precipitation may show significant positive correlations with artiodactyl abundances when the two variables are examined in isolation, the effect is eclipsed by summer temperature in multivariate analyses that include the influence of the latter. This may be the case if summer temperature controlled the effect of summer moisture availability through its direct influence on effective precipitation and plant evapotranspiration.
Although we identify summer temperature as the principal climatic driver, our conclusions remain consistent with earlier work suggesting artiodactyls reached their lowest densities during the Pleistocene-Holocene transition and the early Holocene (Broughton et al., 2008, 2011; Byers et al., 2005). However, as documented by comparisons between the Bonneville and Wyoming basins that are characterized by different temperature regimes, we certainly anticipate spatial variation in this overall trend. We also expect variation at an intra-regional level and it is likely that localized mesic refugia supported higher artiodactyl densities. Sites adjacent to primary migration corridors may have, for instance, contained higher densities of artiodactyls and several records from the adjacent Colorado Plateau in Utah suggest this may have been the case (Louderback, 2022; Rampton, 1999). It is possible that wetlands may have also promoted higher densities of artiodactyls in some settings during the Pleistocene-Holocene transition and the early Holocene (Elston et al., 2014; Elston and Zeanah, 2002).
We also present an empirical test of the costly signaling hypothesis for large game hunting. In the Bonneville Basin, our results are consistent with the hypothesis that proportional increases in large game hunting during the Fremont period was associated with larger audiences and platforms for social display and driven by signaling-based hunting. That period exhibits not only higher artiodactyl abundances than expected given the prevailing summer temperature conditions, but higher bison frequencies—a taxon likely to have garnered the highest signaling value due to its great size.
At the same time, our analysis continues to underscore the overriding importance of climatic controls on the relative contribution of large and small game to Holocene diets in western North America. Indeed, in order to approach any analysis of costly signaling in relation to past large game hunting, climatically-driven variation in artiodactyl abundance must be controlled as we have done here. Further analyses within the Fremont comparing artiodactyl abundances from large sedentary villages relative to more short-term, smaller residential localities, or temporal divisions that capture variation in human population densities and levels of intensification, may be productive.
Finally, the trans-Holocene relationships between climatic variation and artiodactyl abundances may play an important role in forecasting future change in the population dynamics of these taxa. Although the relationships between modern climatic variables and western artiodactyl species are known to some degree—and we have drawn on them to generate expectations for the past—the modern data are limited by the narrow range of variation represented by the recent time series that extend no more than several decades. Our results indicate that summer temperature has been the single critical factor controlling artiodactyl populations in both the Bonneville and Wyoming basins across the past 10,000 years and they also suggest critical summer temperature thresholds in both regions, beyond which artiodactyl populations are invariably depressed. Specifically, artiodactyls occur at consistently low Holocene abundances when mean summer temperature exceeds 17° in the Wyoming Basin and 20° in the nearby, but lower elevation, Bonneville Basin (Figure 6). Similar values would likely apply to many other areas of western North America, which are expected to experience a warming trend as part of the ongoing climate change process. These results are especially concerning given our increasing recognition of the impact of large mammal biomass reduction on ecosystem physical and trophic structures, changes in vegetation community composition, modification of biogeochemical cycling, and further regional and global climate change (e.g. Barnosky et al., 2015; Gill et al., 2009; Malhi et al., 2016).
Supplemental Material
sj-pdf-1-hol-10.1177_09596836231211809 – Supplemental material for Climatic controls and costly signaling: An integrated analysis of Holocene hunting in the Bonneville and Wyoming Basins, USA
Supplemental material, sj-pdf-1-hol-10.1177_09596836231211809 for Climatic controls and costly signaling: An integrated analysis of Holocene hunting in the Bonneville and Wyoming Basins, USA by David A Byers, Ryan McGrath, Peter Yaworsky, Theresa Popp, Gideon Maughan and Jack M Broughton in The Holocene
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.