
Abstract
The objective of this study is to characterize the taphonomic signature of Strix chacoensis in bone and tooth remains of microvertebrates (birds and micromammals). The chaco owl is a bird of prey whose diet and bone modification patterns are little known. In a sample of 41 pellets, an unusual proportion of bird remains over mammal remains was observed. We evaluate the ecological implications of this phenomenon, which could designate Strix chacoensis as a bioindicator of well-preserved forest and shrubland environments when combined with the specific ecological requirements of the prey species. Furthermore, taphonomic patterns vary across taxonomic groups: bird remains exhibited lower modifications due to breakage and digestion compared to micromammal remains. Therefore, this raptor can be characterized as having a minimal impact on bird remains and an intermediate impact on mammal remains.
Keywords
Introduction
Taphonomy has become an essential and unrestricted field for understanding the agents and processes involved in the formation of the archeological and paleontological record (Gutiérrez and Borrero, 2023). Taphonomic studies provide helpful inputs not only to evaluate the genesis of the assemblages but also to make paleoecological and paleobiological inferences from the archeofaunal record (Behrensmeyer et al., 2000).
Microvertebrate species are valuable indicators for evaluating shifts and consistencies in past environments and also for the understanding of the condition and human impact on present-day ecosystems, given their narrow ecological requirements at the microhabitat level (Andrews, 1990; Teta et al., 2014). This represents a promising contribution to what Lyman and Cannon (2004) defined as applied zooarcheology (Wolverton and Lyman, 2012).
The Chaco owl, Strix chacoensis, is a nocturnal, opportunistic, and medium-sized strigiform raptor endemic to southern South America. The species is relatively understudied and is known to inhabit dense and semi-open old or primary dry forests with arboreal and shrubby vegetation (López et al., 2018). Limited observations on the dietary habits of this raptor indicate a preference for small mammals (rodents and marsupials), insects, and occasionally, passerine birds and reptiles (López et al., 2018; Santander et al., 2012).
This raptor is usually associated with well-preserved dry forests, and considering the escalating and rapid degradation of these habitats due to intensive economic activities (Luque et al., 2010), it is likely that its historical distribution was more extensive than its current range. This assumption is supported by the considerable number of recent biological extinctions and local disappearances documented by various ecological, biological, paleontological, and paleoecological investigations (e.g. Barnosky et al., 2017; Teta et al., 2014).
Trees and forests in arid regions play a key ecological role by offering a habitat for biodiversity, preventing erosion and desertification, and regulating water, microclimate, and soil fertility (Cappa et al., 2022; FAO, 2019). However, dry forests are among the most threatened ecosystems worldwide due to agricultural expansion driven by increasing demands for food and energy in developed and emerging countries (Aguiar et al., 2022), often accompanied by intensive extractive activities lacking ecological oversight. The critical situation of South American dry forests, coupled with their limited chance of recovery, calls for prompt evaluations of the remaining biodiversity in these threatened environments. Effective conservation strategies demand a comprehensive grasp of the historical changes in the habitats targeted for preservation. Hence, ecologists have progressively turned to history and paleo-disciplines to elucidate, manage, and understand contemporary ecosystems and landscapes (Hayashida, 2005).
Given the precarious state of these habitats, our objective is to conduct a multi-taxa neotaphonomic study of pellet-derived vertebrate bone assemblages produced by the Chaco owl. Our aim is twofold: to present an actualistic taphonomic pattern that enables reliable inferences about the agent responsible for the formation of the fossil record, and to provide an ecosystemic perspective that allows to stablish a taxonomic “instantaneous” assessment (sensu Terry, 2010) of S. chacoensis prey assemblages in a threatened region of the South American Dry Chaco. Therefore, our intention is to establish a connection between this information and a multi-taxa neotaphonomic perspective by incorporating all skeletal remains found in Chaco owl accumulated pellets. Recognizing the deep potential of the data obtained from small fauna assemblages to reconstruct paleoenvironments, we anticipate that this perspective will make a substantial contribution to paleontological research through an exhaustive taphonomic analysis that can be compared with past microvertebrate accumulations in the region.
Study area
The sample comes from Traslasierra National Park (TNP) (30°59′40.84″S 65°37′36.12″W), with an elevation of 118 m a.s.l. (Figure 1). The area is located within the Arid Chaco subregion, on the border between the provinces of Córdoba and La Rioja. The landscape is characterized by a wide extension of preserved Chaco forest spanning approximately 200,000 hectares, dominated by white quebracho (Aspidosperma quebracho-blanco), mistol (Ziziphus mistol), chañar (Geoffroea decorticans), and brea (Parkinsonia praecox), among other species. Some endemic mammal species of this region include the giant armadillo (Priodontes maximus), different peccary species like the collared peccary (Tayassu tajacu), and the quimilero (Catagonus wagneri) (Torres and Tamburini, 2018). Other species present in the region are the Geoffroy’s cat (Leopardus geoffroyi), the Chacoan mara (Dolichotis salinicola), as well as some species with recent population declines, such as the guanaco (Lama guanicoe), and several micromammal species (Costa and Barri, 2018; Mignino et al., 2021, 2023; Torres and Tamburini, 2018).
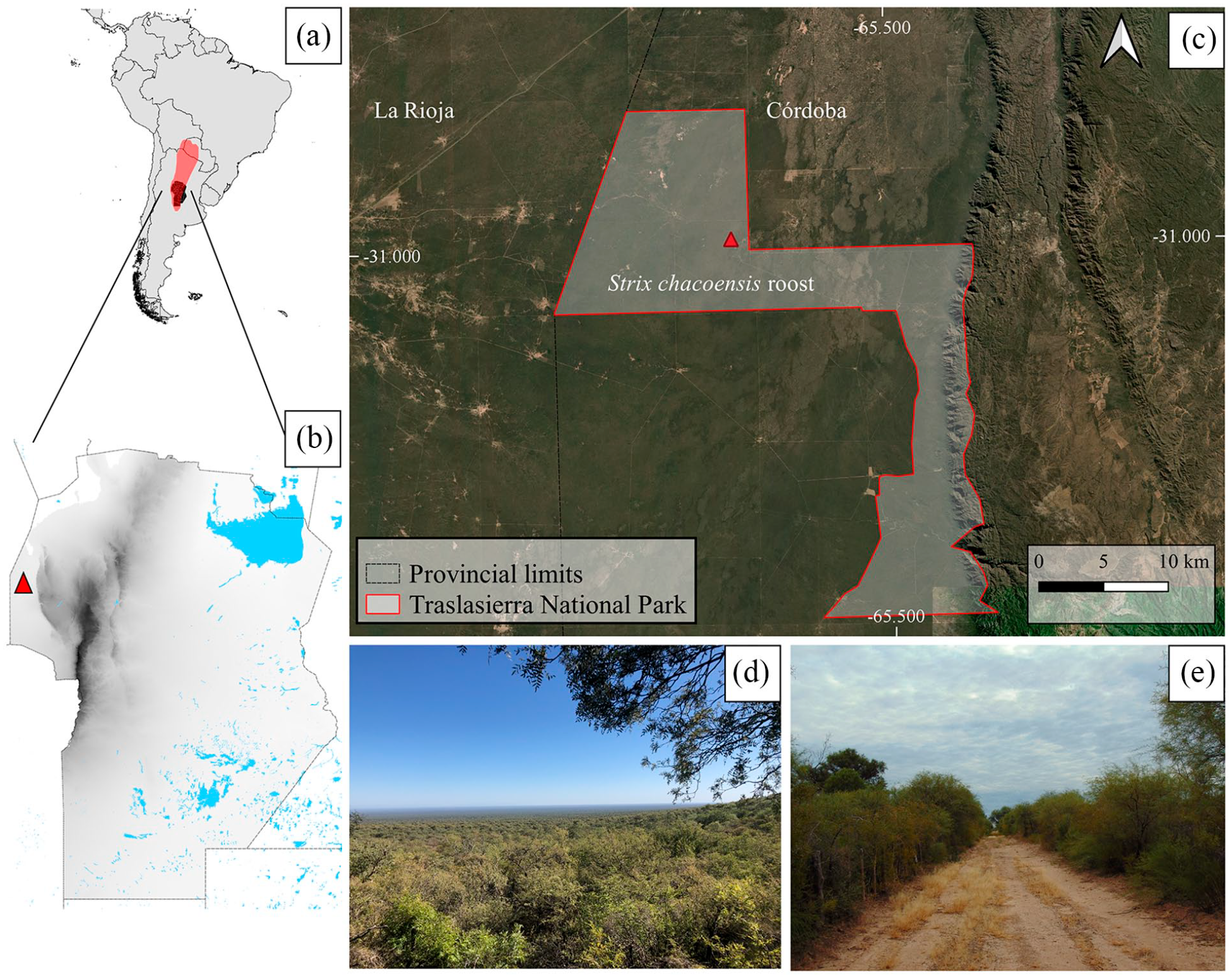
Location of the sample area. (a) General location of the area in South America and distribution (red color) of Strix chacoensis. (b) Location of the sampling area in the province of Córdoba. (c) Perimeter of Traslasierra National Park. (d) General view of Traslasierra National Park. (e) Characteristic environment of the area.
In this area, rainfall is limited due to the natural barrier effect of the surrounding mountains, and therefore highly saline soils are prevalent. Based on rainfall data collected by local farmers over the last six years, there is a substantial distinction between the wet season (November–April) and the dry season (May–October). Between the years 2013 and 2018, the average accumulated rainfall stood at 74.2 mm for the coldest and driest months, while the average accumulated rainfall during the wet season was 424.5 mm (Mignino et al., 2021).
Strix chacoensis
The Chaco owl (Strix chacoensis) is a medium-sized strigiform raptor, with a total length ranging from 35 to 38 cm and weight between 350 and 500 g. This species is endemic to southern South America and inhabits an extensive range covering approximately 760,000 km2, spanning from the southern Chaco region of Bolivia and Paraguay to the Argentine provinces of Buenos Aires and Córdoba (Figure 1a). Its presence is documented in semi-open, arid environments with the presence of thorny shrubs and cacti of varying sizes, which serve as daytime perches (Kratter et al., 1993; Santander et al., 2012). Furthermore, it also occupies ancient and primary forests characterized by dense vegetation cover and intricate plant structures (Pereyra-Lobos and Jara, 2013). The Chaco owl is primarily active during nighttime, using the cover of darkness to prey on small vertebrates, including mammals, birds, amphibians, reptiles, and invertebrates such as scorpions (Santander et al., 2012).
Conservation studies have highlighted the vulnerable status of S. chacoensis (López-Lanús et al., 2008). BirdLife categorized this raptor as Near Threatened (NT) due to ongoing forest fragmentation, degradation, and loss across its geographical range. Remarkably, the presence of the Chaco owl has been established as a bio-indicator of well-preserved native forests in various South American biomes (López et al., 2018 and references therein).
Materials and methods
During August 2021, which corresponds to the cold and dry season, a total of 41 pellets produced by the Chaco owl were collected. These pellets were retrieved from an abandoned building as part of a biocultural conservation initiative focused on evaluating different predators of the Dry Chaco region as agents of accumulation of small vertebrate remains at local archeological and paleontological sites.
The collected pellets were meticulously processed with surgical instruments, enabling the separation of bone remains and teeth. A thorough taxonomic identification was carried out, aiming to achieve the highest possible level of accuracy. Taxonomic assignments for bird remains relied on morphological characteristics and diagnostic features of specific long bones (humerus, ulna, carpometacarpus, tibiotarsus, and tarsometatarsus) (De la Peña, 2020; Olsen, 1979). Craniomandibular remains were identified from general features. For micromammal identification, teeth morphology and diagnostic traits of mandibles and skulls were used (Patton et al., 2015; Udrizar-Sauthier et al., 2020). When required, comparative zoological collections from IDEVEA-CONICET and IDACOR-CONICET were consulted.
Anatomical and taxonomic determinations were made while considering bone specimens’ completeness, lateral position, and fusion. Minimum number of individuals (MNI) and minimum number of elements (MNE) calculations were carried out following Lyman’s (2008) approach to assess the abundance of identified specimens per taxon.
For the purpose of diet analysis, an estimation of prey biomass was conducted. This calculation is represented as (Bi) = [(Spi Ni)/Σ(SpiNi) ] × 100, where Spi represents the weight of species i, and Ni stands for the number of individuals of species i consumed. This calculation allowed to establish the proportion in which each taxon contributed to the total biomass consumed (Marti et al., 2007). Reference data for prey biomass were obtained from De la Peña (2020) for birds and from the Argentinian mammal categorization (https://cma.sarem.org.ar).
The taphonomic approach followed the methodological guidelines provided by Andrews (1990), Bocheński et al. (1993), Bocheński and Tomek (1997), Fernández et al. (2017), Bocheński et al. (2022), and Montalvo et al. (2020). To evaluate skeletal part representation, the relative abundance index of skeletal parts was computed for both bird and micromammal remains. This index, expressed as Ri, was calculated as: Ri = MNEi/(Ei × MNI) × 100. This formula considers the representation of each element in the sample (MNEi) relative to the number of times that element is present in the skeleton (Ei), and the minimum number of individuals (MNI). For bird remains, the wing/leg ratio was determined by assessing the representation of humerus, ulna, and carpometacarpus in comparison to femur, tibiotarsus, and tarsometatarsus (Ericson, 1987). In micromammals, the ratio of cranial to postcranial elements was evaluated using the PCRAP/CR and PCRT/CR indices (Andrews, 1990; Fernandez-Jalvo and Andrews, 1992). The PCRAP index is calculated as: PCRAP = (femur (f) + tibia (t) + humerus (h) + radius (r) + ulna (u)) × 8/(hemimandible (mb) + hemimaxillary (mx) + molars (m)) × 5] × 100. The PCRT index is expressed as: PCRT = [(h + f)/(mb + mx)] × 100. Additionally, the relationship between distal (D) and proximal (P) elements was assessed through the D/P index, calculated as: D/P = (t + r)/(f + h)] × 100.
Regarding breakages, evaluation included bird remains, specifically skulls, mandibles, and postcranial elements (scapula, coracoid, humerus, radius, ulna, carpometacarpus, femur, tibiotarsus, and tarsometatarsus) (Bocheński et al., 1993). For micromammals, breakage evaluation adopted the methodologies outlined by Andrews (1990) and Montalvo et al. (2020), focusing on skulls, mandibles, and commonly used postcranial elements (i.e. humerus, ulna, radius, femur, and tibia).
Potential modifications resulting from predation and consumption, such as perforations, furrows, and chewing marks, were analyzed across all anatomical units (Bocheński et al., 1998; Laroulandie, 2002).
Concerning modifications due to digestion in avian remains, assessments were conducted both on the cortical surface of long bones and on broken edges. The categorization scheme proposed by Bocheński and Tomek (1997) was followed, which includes three categories of bone surface alterations (unmodified (A), rounded (B), and sharp (C)), and two categories of modifications on broken edges (sharp (A) and rounded (B)). For micromammal remains, digestion assessments followed the parameters defined by Andrews (1990) and Fernández et al. (2017). This involved considering incisor and molar teeth (both in situ and isolated), as well as distal humeri and proximal femurs. Digestion degrees were categorized as absent, light, moderate, heavy, and extreme, depending on the type of modification. Bone surface modifications were examined using a Motic SMZ-140-N2GG binocular microscope, and photographic documentation was carried out using a Motic 171-TLED stereomicroscope (8x–50x).
Results
A total of 1025 specimens (NISP) were retrieved from the collection of 41 pellets. Within this assemblage, bird remains accounted for 65.85% (NISP = 675) of the total, while the remaining 34.15% (NISP = 350) were attributed to micromammals. Bird remains were identified in 20 owl pellets, representing a frequency of 48.78%. In contrast, micromammals were present in 43.90% of cases (18 pellets).
Taxonomic composition
Table 1 shows the data derived from taxonomic identifications. In these taxonomic groups, a combined total of 49 individuals were successfully recognized, comprising 69.38% birds and 30.62% micromammals.
Taxonomic representation of birds and micromammalian remains contained in pellets of Strix chacoensis from Traslasierra National Park.
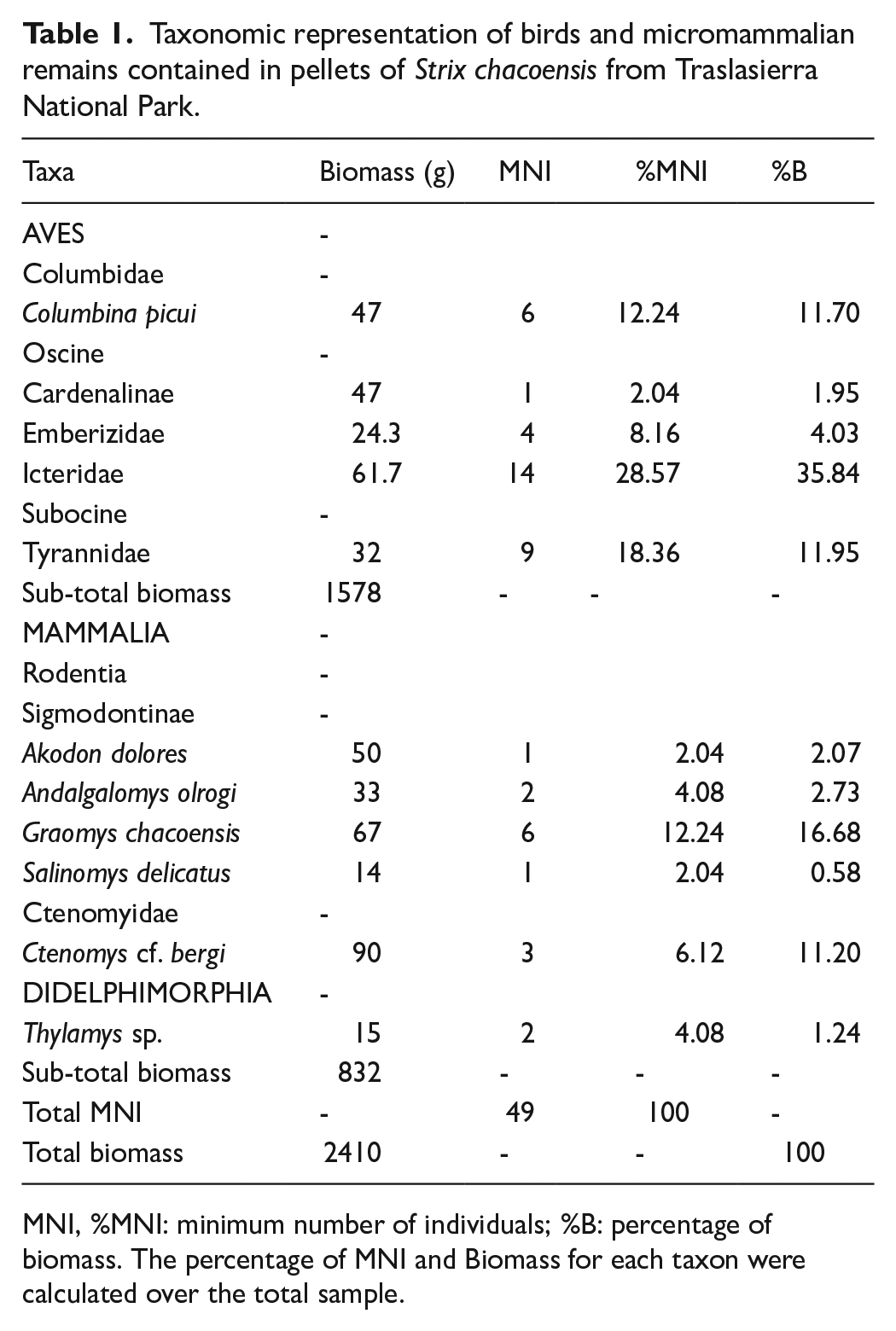
MNI, %MNI: minimum number of individuals; %B: percentage of biomass. The percentage of MNI and Biomass for each taxon were calculated over the total sample.
With an MNI of 34, the avian component was characterized by the presence of three families, one sub-family and one species (Table 1). In this sense, Icteridae birds played a prominent role in the sample followed by tyranid birds.
Micromammals comprised six taxa (one genus and five distinct species). In a total of 15 micromammals, the most abundant species was Graomys chacoensis (Table 1).
Concerning the estimation of dietary biomass, birds comprised a significant proportion, accounting for 65.47% of the total biomass consumed by S. chacoensis and micromammals constituted the remaining 34.53%. The Icteridae birds held a predominant position in biomass contribution (Table 1).
Anatomical representation
A minimal number of elements (MNE) was determined as 580 for bird remains and 312 for the micromammal assemblage (Table 2). The calculation of the relative abundance index (Ri) considered a minimal number of individuals (MNI) of 34 for birds and 15 for micromammals. Within this context, birds exhibited an average Ri of 47.94%, while micromammals displayed a value of 38.24%. In the case of birds, mandible, pelvis, and tarsometatarsus were the most prominently represented anatomical units. For the micromammal group, mandible, humerus, and femur were the most widely found (Figure 2).
Relative abundance of skeletal parts (Ri) of birds and micromammal prey contained in pellets of Strix chacoensis from Traslasierra National Park.
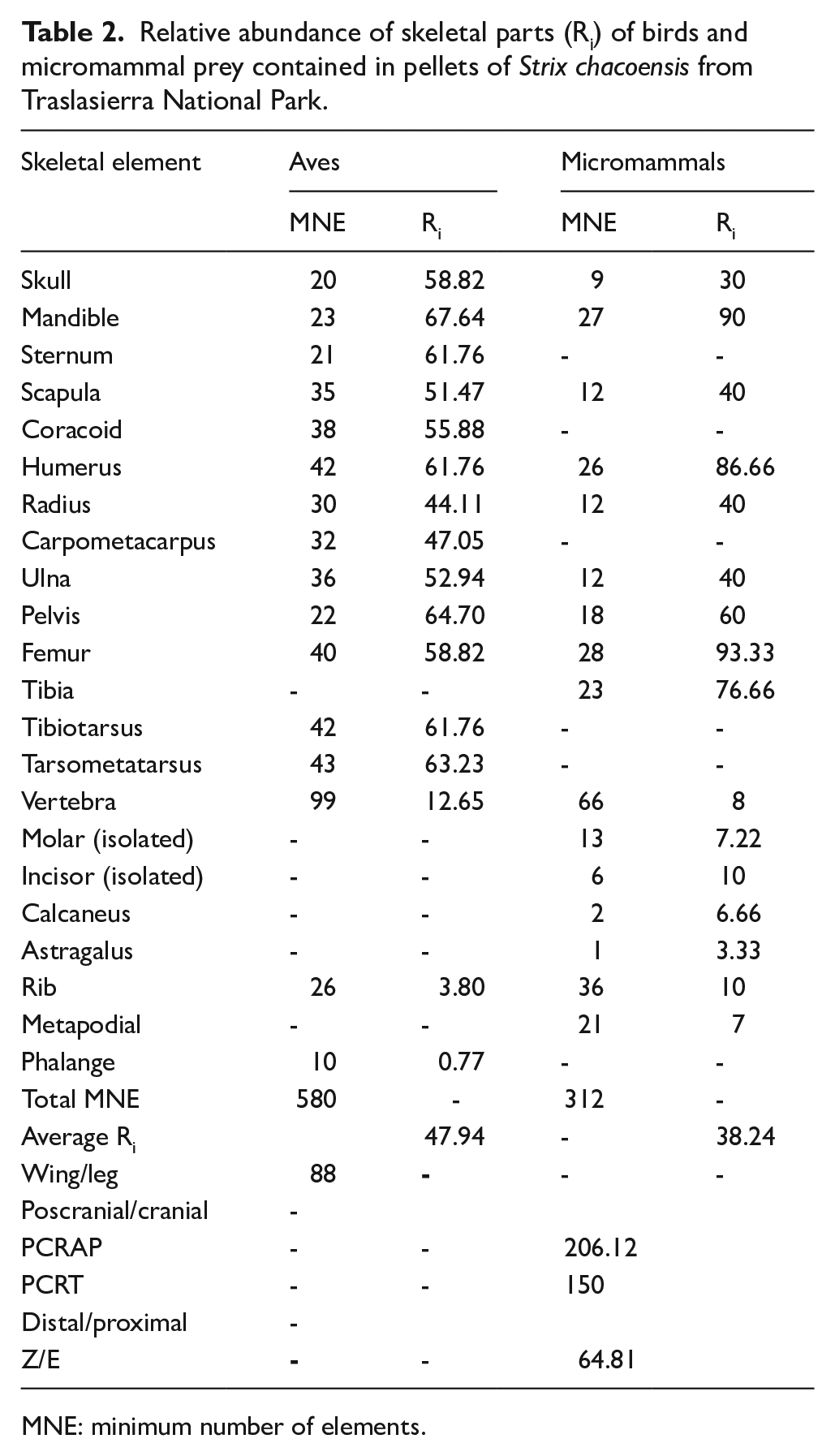
MNE: minimum number of elements.

Percentage of relative abundance of skeletal parts (Ri) of birds and micromammalian prey contained in pellets of Strix chacoensis from Traslasierra National Park. The percentages correspond only to the elements examined.
In terms of indices assessing the ratio between skeletal parts, the avian group demonstrated a marginally enhanced representation of leg components, namely femur, tibiotarsus, and tarsometatarsus. In contrast, among micromammals, the proportion between cranial and postcranial elements exhibited a more pronounced representation of the former group. Additionally, the ratio between distal and proximal units showed a notable trend toward the latter group (Table 2).
Breakage
In terms of cranio-mandibular remains within the avian assemblage, breakages were observed at a frequency of 44.23% (Table 3). A significant percentage of intact skulls were recovered, with smaller proportions consisting of skull fragments featuring premaxillary portions and calotte fragments with basioccipital attachments. Mandibles exhibited a nearly 60% frequency of completeness, and those with breakages primarily displayed damage to their articular processes. Postcranial remains displayed a completeness frequency approaching 80% (Table 3). Among these remains, scapula and femurs were the most affected by breakages. In contrast, humeri and carpometacarpals were the most well-preserved bones.
Breakage categories and frequencies in bird bones contained in pellets of Strix chacoensis from Traslasierra National Park.
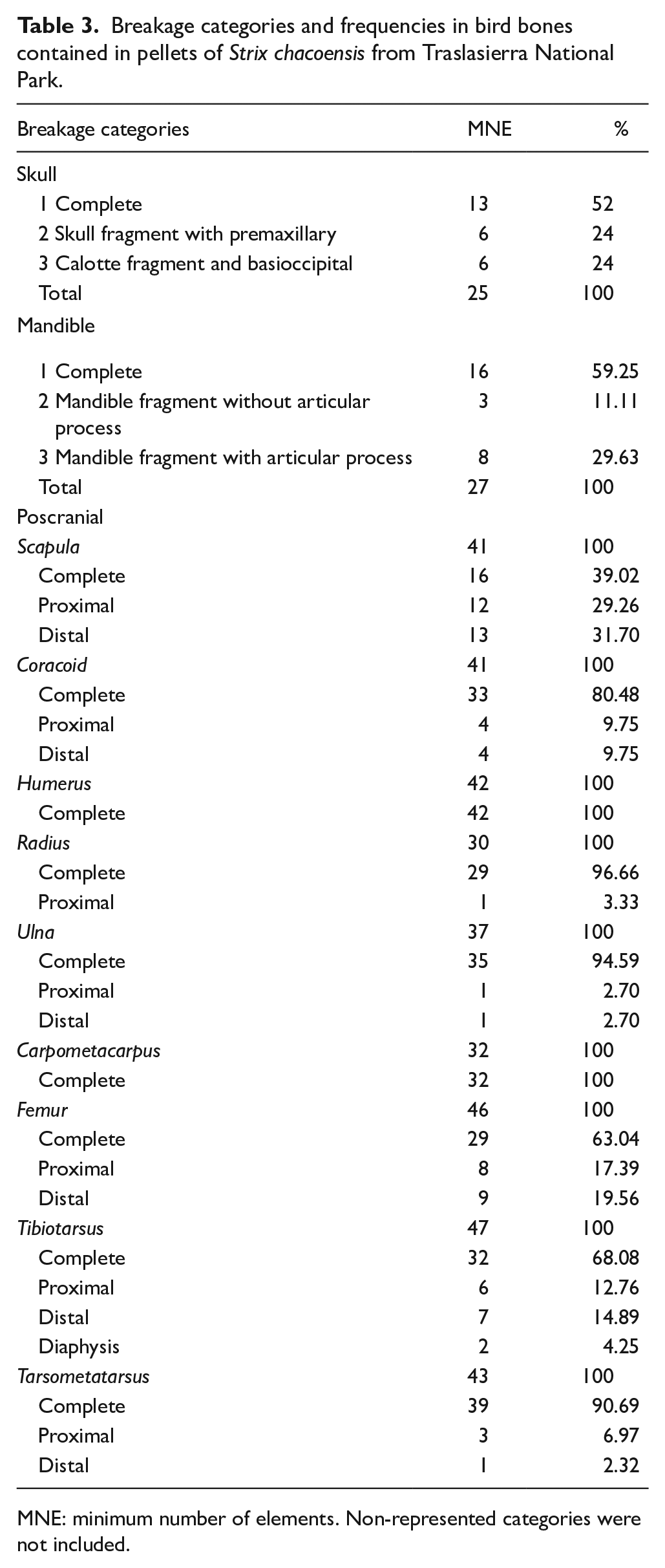
MNE: minimum number of elements. Non-represented categories were not included.
Furthermore, a total of 18.79% (MNE = 109) of bird remains were recovered articulated. Within this subset, the presence of tendon and skin remnants was identified (Figure 3a). A total of 96% of these articulated remains belonged to the anterior limbs.
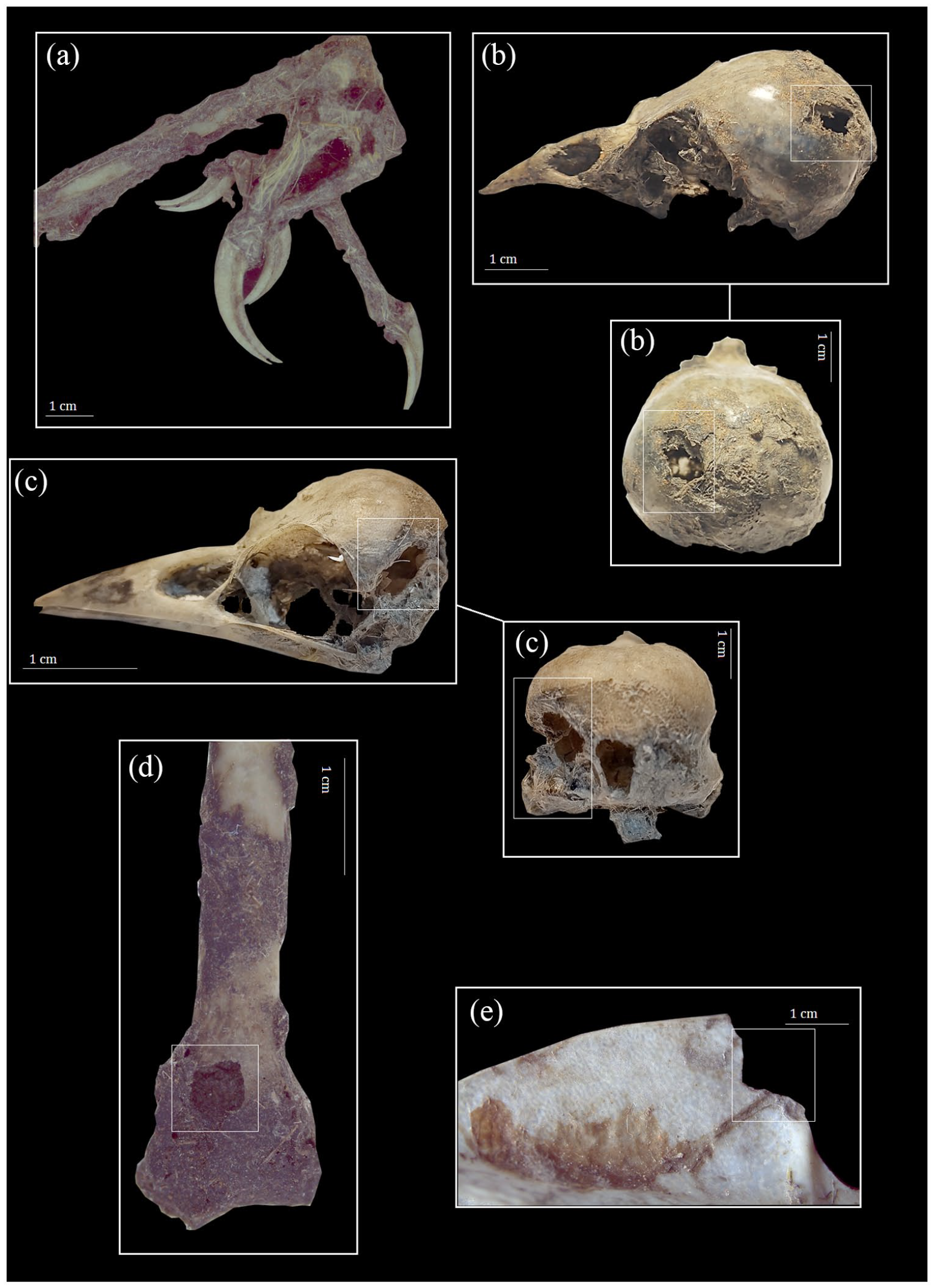
Remains of birds with punctures and chewing marks. (a) Articulated forelimbs with the presence of tendons, skin, and feathers. (b and c) Skull with punctures on occipital region caused by raptor beaks. (d) Humerus with a puncture on distal epiphyses caused by raptor beaks. (e) Sternum with chewing marks on the lateral border.
Fragmentation in micromammal remains displayed proportions similar to those observed in avian craniomandibular remains, with higher frequencies recorded in postcranial anatomical units (Table 4). Maxillae and mandibles exhibited breakages at a proportion near 45%. Only one complete skull was recovered; the others were identified primarily through the frontal bones and zygomatic region. Conversely, mandibles demonstrated a relatively high completeness frequency of around 70%. Incomplete mandibles typically displayed partial breakage of the ascending branch (Table 4). Postcranial remains showed a fragmentation frequency of approximately 35% and ulna and tibia were the elements most significantly affected. In contrast, humerus, radius, and femur were the best-preserved remains, each exhibiting completeness rates exceeding 75%. Finally, no articulated micromammal remains were identified, and the presence of tendons or skin remnants was not recorded.
Breakage categories (sensu Montalvo et al., 2020) and frequencies in micromammals bones contained in pellets of Strix chacoensis from Traslasierra National Park.

MNE: minimum number of elements. Categories 4, 5, 6 and 8 were not included because they were not represented in the sample.
Perforations and chewing marks
A frequency of 2.75% of the bird specimens showed perforations marks (Figure 3). In this sense, 87.50% of specimens present this type of alteration within the skull remains. The perforations were primarily concentrated in the frontal and parietal bones as well as the temporal fossae, with a high prevalence (78.57%) in these regions (Figure 3b and c). Additionally, perfortations were identified in only two cases, affecting the distal humerus and sacrum region of the pelvis (Figure 3d).
Conversely, 7.41% of the remains exhibited modifications indicative of chewing. In 60% of these cases, the alterations were found on the lateral edges of the sternum (Figure 3e). In a smaller proportion, chewing-related damage was detected in skull, coracoid, ulna, pelvis, and tibiotarsus bones.
Modification by digestion
All degrees of digestion were identified within the avian assemblage (Table 5 and Figure 4). A 20.80% of the total remains showed signs of digestion. Category B alterations (rounded) predominated significantly among the affected elements, while three elements displayed modifications characteristic of category C (sharp). In cases falling under category B, pits and depressions with rounded edges were discernible. Conversely, elements categorized as category C showed pits and depressions with angular, rough, and sunken edges.
Degrees and frequencies of digestion in bird remains contained in pellets of Strix chacoensis from Traslasierra National Park. Modification categories: unmodified (A); rounded (B); sharp (C).

MNE: minimum number of elements.
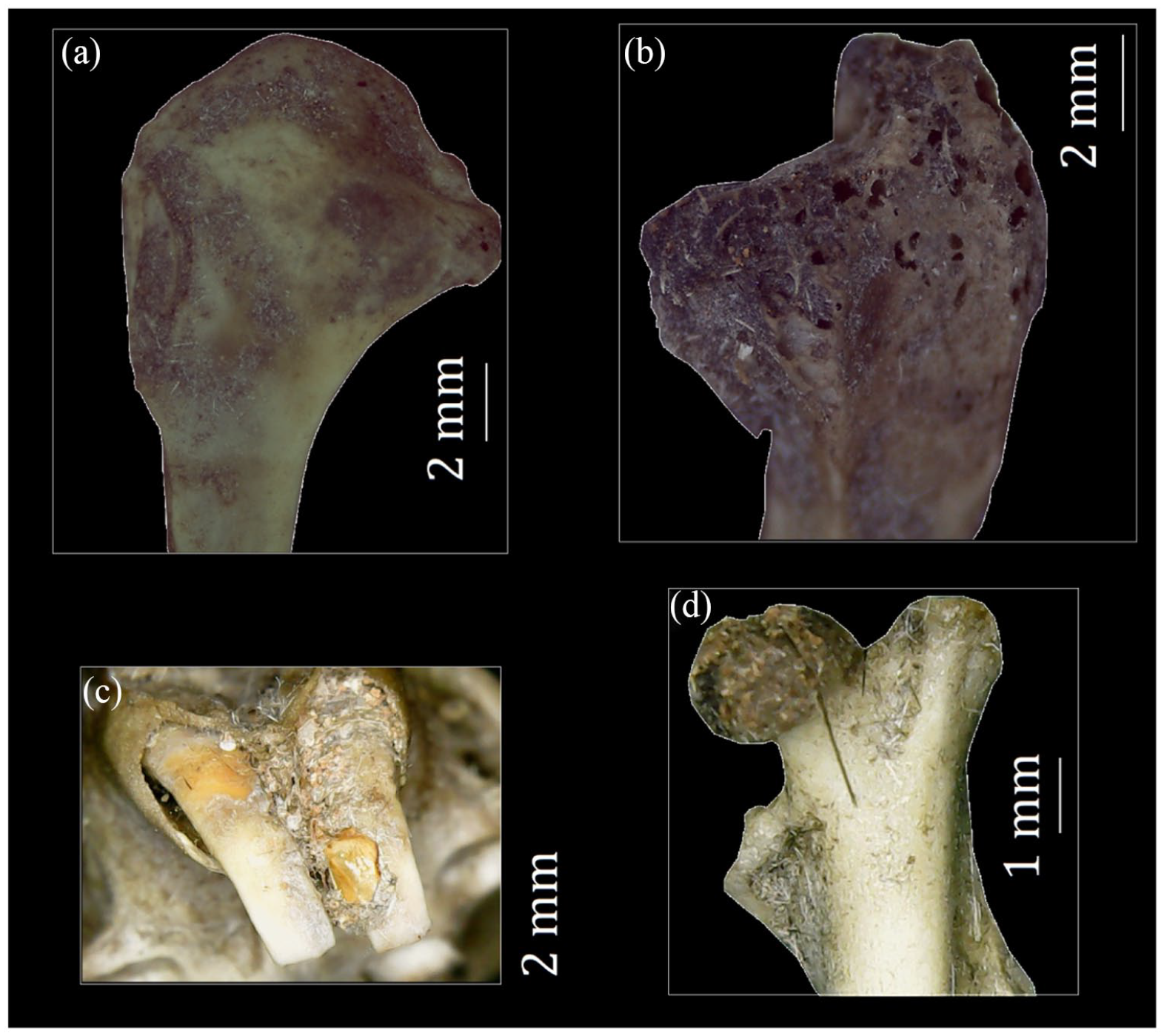
Examples of bird and micromammal remains with digestion modifications generated by Strix chacoensis. (a) Light digestion on humerus. (b) Light digestion on tarsometatarsus. (c) Moderate digestion on upper incisors of Ctenomys cf. bergi. (d) Moderate digestion on the proximal femur of the sigmodontine rodent.
Craniomandibular remains showed a digestion frequency of 17.54%, while postcranial remains demonstrated a proportion of 21.15%. The first set only displayed light modifications (category B). In contrast, the second group exhibited a mix of alterations covering categories B and C. Tibiotarsus, femur, and radius were the most clearly affected units, whereas no modifications were observed in furcula, fibulae, vertebrae, ribs, and phalanges (Table 5).
The micromammal remains showed moderate digestion frequencies, comprising a wide spectrum of alteration degrees (Table 6 and Figure 4). A total of 45.45% of the analyzed remains underwent modifications ranging from light to heavy alteration categories, with no instances assigned/attributed to the extreme category. Postcranial remains showed an alteration frequency due to digestion of 35.18%, mostly exhibiting light to moderate levels of digestion (Figure 4). Tooth remains, including incisors and molars, evidenced a digestion proportion of 44.66%. Among incisors, 58.13% showed primarily light damage, although two cases displayed moderate and heavy digestion. On the other hand, the 39.25% of molars (both in situ and isolated) exhibited signs of digestion. The light digestion category prevailed, with only one instance showing a moderate degree of alteration (Figure 4).
Degrees and frequencies of digestion in micromammal remains contained in pellets of Strix chacoensis from Traslasierra National Park.

MNE: minimum number of elements.
Discussion
Ecological inferences and implications for archeological and paleontological contexts
This study revealed an atypical abundance of bird remains in the pellets of S. chacoensis, representing about 70% (NISP) of the total remains, in contrast to micromammals which accounted for around 30%. Among the avian prey, Icterid birds showed the highest prevalence, followed by representatives from the Tyrannidae family and C. picui. Among the micromammal prey, G. chacoensis was the most represented species. Ctenomys cf. bergi, Salinomys delicatus, Akodon dolores, Andalgalomys olrogi, and the marsupial Thylamys sp. were present to a lesser extent.
Significantly, the identification of A. olrogi marks the first report of this species in Chacoan environments in the province of Córdoba. In particular, the conservation status of both A. olrogi and S. delicatus, two co-occurring species in this assemblage, has been classified as Least Concern (LC) and Vulnerable (VU) respectively, according to assessments conducted by the Argentine Society for the Study of Mammals (Rodríguez et al., 2019). These species, characterized by their adaptations to desert conditions, remain relatively elusive and have been infrequently detected through conventional sampling methods like live trapping (Mignino et al., 2023).
In a comprehensive examination of the seasonal diet variation of S. chacoensis in the Monte Desert of Central Western Argentina, Santander et al. (2012) reported a distinct dietary composition that the present study. Their study indicated that the most abundant prey in the Chaco owl pellet-derived assemblages were micromammals, insects, and scorpions. Remarkably, birds were represented with an exceptionally low frequency. Across the entire sample, micromammals reached proportions close to 40%, whereas birds exhibited a frequency of 2.5%. Notably, the highest occurrence of bird prey was observed during summer, reaching a percentage of 8.7% (Santander et al., 2012).
Kratter et al. (1993) also identified notable trends studying samples from two stomachs of S. chacoensis in the Chaco environments of southern Bolivia. Their analysis identified scorpions, cockroaches, large spiders, centipedes, and blind snakes as the predominant dietary components. However, no mention was made of the discovery of micromammals or birds in this case study.
On the other hand, Figueroa et al. (2006) studied the diet of S. rufipes and documented the prevalence of insects, constituting over 70% of the diet. Nevertheless, micromammals played a significant role in biomass contribution (89.2%), accounting for 24% of the diet in a sample obtained from the Andean Nothofagus-Araucaria forest in southern Chile. In line with these findings, birds showed a particularly low frequency (1%). Similarly, in forested environments with high occurrence of Aextoxicon punctatum and Podocarpus saligna along the Valdivia coast of southern Chile, Martínez (1993) highlighted the predominance of insects (55.31%) and micromammals (42.55%) in the diet of S. rufipes. Notably, micromammals represent a substantial 94.7% of the consumed biomass. Conversely, birds comprise only 1.32% of the total and contributed a mere 1.5% to the overall biomass.
These comparative studies underscore the remarkable variability in the dietary preference and composition of S. chacoensis and S. rufipes across different geographical regions and ecological contexts.
In summary, existing studies on the ecology and diet of raptors within the Strix genus have consistently highlighted the dominance of mammals, often followed by insects, in the dietary preference of these owls. As a result, it has been generally assumed that birds constitute occasional items in their diet. However, the unexpectedly high proportion of bird remains, both in terms of individual frequency and biomass contribution in the S. chacoensis samples from Traslasierra National Park (TNP) presents an intriguing finding.
Several interrelated hypotheses could potentially shed light on this phenomenon. One primary factor is the extensive preservation of large areas of primary native Chaco forests in TNP, covering approximately 200,000 hectares. This preservation becomes especially significant given the widespread trend of vegetation transformation and composition change in central Argentina due to the expansion of agroecosystems associated with soybean cultivation, real estate development, and infrastructure projects. These activities have led to the replacement of open grasslands and native forests, resulting in higher rates of deforestation and alterations in vegetation structure and composition.
The critical role of vegetation as habitat, providing shelter, feeding grounds, and nesting sites for birds, is crucial to understanding this phenomenon. Alterations in the composition and structure of vegetation due to disturbances can directly impact the composition, structure, and density of bird assemblages of any area. Silvetti et al. (2023) recently examined the impact of human disturbances on the Chaco forest in Córdoba province and highlighted the correlation between these disturbances and changes in the taxonomic structure of the bird community. They argued that significant changes in forest ecosystems have affected not only specialist bird species but also those adapted to shrubland environments, which often use mid-stratum shrubs for nesting and feeding. This suggests that habitat preservation could play a vital role in influencing the dietary preferences of S. chacoensis.
Furthermore, the specific habits of both predator and prey might contribute to the high occurrence of bird remains. Small birds, particularly passerines, are primarily diurnal, while S. chacoensis is a raptor with strict crepuscular and nocturnal habits. The owl’s opportunistic hunting behavior during times when its avian prey is resting in nests or roosting may lead to an increased representation of birds in its diet.
Seasonal variations in rodent abundance could also play a role. Many sigmodontine rodent species, which constitute a significant portion of S. chacoensis’ diet, reproduce in spring and summer, leading to higher abundance during that time. Toward the end of summer and beginning of autumn, rodent populations peak, but by the end of autumn and beginning of winter, their numbers decrease significantly (O’Connell, 1982).
Considering that the TNP sample corresponds to the end of the cold season (winter), it is plausible that the decrease in rodent abundance during this time could have led S. chacoensis to rely on birds as a food source. Overall, the complex interplay of habitat preservation, predator-prey interactions, and seasonal variations in prey availability could collectively explain the unusual dietary composition observed in the Chaco owl pellet-derived vertebrate bone assemblage from TNP.
The diverse micromammal prey species found in the pellets of S. chacoensis suggest that this raptor forages in a range of environments, with a notable prevalence of scansorial and arboreal species. Each of these prey species is adapted to specific ecological niches: G. chacoensis is associated with complex dry forest environments; S. delicatus and A. olrogi thrive in semi-arid and arid settings with shrubland areas and proximity to salt flats; A. dolores occupies grassland and shrubland environments; and C. cf. bergi is exclusively found in grassland areas (Patton et al., 2015). This prey selection showcases the versatility and adaptability of S. chacoensis in exploiting various ecological niches for hunting.
The combination of the high proportion of bird remains and the ecological requirements of the micromammal prey species provides valuable insights into paleoenvironmental reconstructions based on microvertebrate bone remains found in archeological and paleontological contexts. The presence of S. chacoensis in such contexts could potentially serve as an indicator of past dry forest environments. As an accumulating agent, the owl’s dietary preference and its sensitivity to disturbance in dry forest habitats can contribute to understanding the historical distribution and composition of these environments.
Furthermore, given its negative response to disturbance in native ecosystems, the presence or absence of this raptor could potentially provide an insight into the historical impacts of human activities or other ecological changes on these environments. The presence of S. chacoensis could suggest well-preserved and less disturbed areas, while its absence might indicate areas that have undergone significant environmental changes.
The Chaco owl as the accumulator of vertebrate material: Taphonomic considerations
By analyzing the modifications and damage inflicted on different taxonomic groups, such as birds and micromammals, it is evident that the same raptor, S. chacoensis, produced distinctive damage on different preys. The recovery of various anatomical units from both bird and mammal skeletons suggests that owls like S. chacoensis tend to ingest a wide range of skeletal elements. The relative abundance percentages observed in this study indicate that the predator ingests the whole prey.
Moreover, the findings of this study provide an opportunity for comparative analysis with previous research on S. chacoensis and its interactions with both bird and micromammal assemblages. By examining previous works such as Andrews (1990) and López et al. (2018), it is possible to identify distinct patterns of damage and modification caused by S. chacoensis on different prey. This comparison underscores the predator’s ability to adapt its feeding behavior and bone modification to different taxonomic groups, shedding light on the complexity of its interactions with various prey species.
The average index revealed slightly better preservation than that reported by López et al. (2018) for a sample of S. chacoensis from the Monte Desert of central western Argentina. Likewise, Andrews (1990) obtained a similar average abundance value for samples produced by S. nebulosa and results that were below those achieved in our work for S. aluco.
Moderate and equivalent frequencies of fragmentation were observed in cranio-mandibular remains between birds and micromammals (ca. 45%). Considering only the remains of the skull, micromammals had only one instance where the element was completely recovered. Contrarily, birds exhibited completeness levels exceeding 50%. Micromammals showed better completeness levels than birds in their mandibles, with proportions of more than 70% and 59%, respectively. The difference in skull fragmentation may be due to the stronger cranial vault of birds who were preyed upon, which was likely a result of early cranial suture fusion compared to the weaker posterior region of micromammal skulls (e.g. sigmodontine rodents and marsupials). In contrast, postcranial bones showed better preservation in the anatomical units of birds than in those of micromammals. As a result, bird remains displayed a breakage rate of 20% while micromammal remains had a frequency of about 35%. The long bones of birds have a thin cortical structure, and contain trabeculae that increase their hardness, probably due to their adaptation to flight (King and McLelland, 1984). Nevertheless, these bones are not shown to be more durable and flexible than those of micromammals. We believe that the most logical explanation comes from the occurrence of articulated remains and the presence of soft tissue.
Just over 18% of the bird remains were recovered in an articulated condition. Nevertheless, a considerable percentage was identified in the postcranial units. In contrast, none of the micromammal anatomical units showed any connection or presence of soft tissues in any of the instances. This characteristic may be crucial in understanding variations in the frequency and severity of the breakage of this type of remains.
Based on known references on fragmentation in bird remains, Bocheński et al. (1993) and Bocheński and Tomek (1994) found greater modifications by this variable in samples produced by S. aluco and Asio otus. Consequently, for the former, 7% of the skulls were complete, and 39% of the mandibles exhibited no damage. Meanwhile, the samples produced by A. otus did not show completeness in skulls, and mandibles reached a frequency of 16%.
As mentioned, breakage of postcranial bird remains reached a frequency of 20%. In contrast, a higher and considerable frequency of postcranial remains (more than 50%) was broken in samples produced by S. aluco (Bocheński et al., 1993).
In our study, the elements with the highest frequency of completeness were humeri, carpometacarpus, radii, and ulna. Coincidentally, these units correspond to the set of hind limbs. This may be particularly connected to the type of locomotion of the preyed birds. Given that the taxa that make up the assemblage are fliers; their forelimbs are more robust than their hind limbs. This may generate greater resistance to breakage processes (Cruz, 2003). In turn, the highest values of bone mineral density are located in the bones of the forelimbs and shoulder girdle (i.e. humerus and coracoid) and the lowest in the hind limbs (i.e. femur and tarsometatarsus).
Regarding breakage in micromammal bones, close to 90% of the skulls exhibited some type of breakage. The mandibles evidenced higher completeness, with a frequency of 70%. This agrees with that reported by López et al. (2018) for samples produced by S. chacoensis in the Monte Desert of central western Argentina. These values showed greater breakage than those reported by Andrews (1990) for S. aluco and were close to those reported for S. nebulosa.
We observed higher frequency and intensity of postcranial breakage than observed by López et al. (2018) for S. chacoensis and by Andrews (1990) for samples of S. nebulosa and S. aluco.
The record of modifications caused by beaks and claws exclusively in bird remains is an interesting aspect for the characterization of the taphonomic “signature” of this bird of prey. Generally, this damage can occur at times when the raptor hunts and transports its prey (usually by biting the head) or because of the impact of the beak to finish killing its prey and ingesting it. This could be a probable explanation for most records of these marks in the remains of bird skulls. The configuration and structure of micromammal skulls may explain the difficulty in finding this kind of modifications, although the high breakage may be linked to this process.
In this study, modifications by digestive corrosion showed a notable difference between birds and micromammals. Consequently, we detected 20% of modifications by digestion in bird remains, and 45% in micromammal remains. Within these assemblages, postcranial bird units showed a digestion frequency slightly higher than 20%, whereas in micromammal remains, modifications by digestion were close to 35%. We consider a number of aspects that could be acting together in this case. On the one hand, the sample could be the result of an accumulation generated by individuals of different ages, causing dissimilar records of digestive corrosion signals. Young individuals have been found to produce a more significant number of modifications in ingested remains than adults (Taylor, 1994). This seems to represent a plausible scenario, as young individuals exert greater prey use to optimize ingestion during the initial stages of development (Andrews, 1990; Taylor, 1994; Williams, 2001). On the other hand, on some occasions, remains of birds showing articulated bones of tendons and feathers were recorded. In these cases, the remains were not affected by digestive corrosion. Indeed, given the high abundance of birds in the environment, the remains may have been ingested and expelled in a short period. This may have resulted in the existence of tissues and feathers and, therefore, less digestion damage.
The remains mostly affected by digestion were tibiotarsus, femur, radius, and ulna. This was consistent with that demonstrated for strigiform raptors, which exhibited greater involvement of long bones than that of the rest of the anatomical units.
Regarding alteration by digestive corrosion in micromammal remains, the frequency and intensity of affectation in incisors were considerably higher than that reported by Andrews (1990). At the same time, López et al. (2018) demonstrated a very similar proportion of modification by digestion for assemblages produced by S. chacoensis to that reported in the present study. Indeed, both the present work and López et al. (2018) found that frequencies above 50% of modified remains were detected. In both cases, light degree prevailed, and moderate modifications were detected at low frequencies. A contradictory trend reported by López et al. (2018) was found in the proportion of digestion on molars. These authors revealed a percentage of occurrence close to 10%, and in this work, molars were affected by almost 40%. In the samples from TNP, light and moderate modifications were detected. In comparison, López et al. (2018) indicated light modifications exclusively.
According to what was observed in the postcranial elements (distal humeri and proximal femora), the percentage of digestion for TNP samples was close to 35%, and light and moderate alterations were detected. This was higher than what was observed in the sample of S. chacoensis from the central Monte Desert (López et al., 2018). Here, the authors indicated a frequency of 20% with light, moderate, and heavy damage.
Conclusions
In this work, we report a multi-taxa neo-taphonomic analysis of prey remains digested by S. chacoensis. This is an owl associated with well-preserved forest environments, whose knowledge based on its diet and modification is scarce. The high occurrence of birds over micromammals could be a correlate of an interdigitation of elements such as habitat preferences of S. chacoensis, greater abundance of birds in areas of preserved forest, and seasonality to which the analyzed samples correspond to.
The complex interplay between dietary preferences, prey ecology, habitat selection, and sensitivity to disturbance makes S. chacoensis a promising candidate for informing paleoenvironmental reconstructions, acting as a potential bioindicator of ecological impact.
Also, given that it is a stenoic owl, this species can be used as a predictor of ecosystem impact and ecological preservation, by finding its own bone remains from archeological sites and/or its action over the preyed faunal remains.
The results of taphonomic patterns in bird remains showed scarce bone loss, low to moderate breakage, and a low proportion of remains with alteration by digestion. In contrast, micromammal remains evidenced moderate bone loss, with a preponderance of postcranial elements and zygopodial units, moderate breakage of skeletal remains, and a high proportion of diagnostic elements modified by digestion. We attribute these differences primarily to the presence of articulated remains, tendons, muscles, and feathers that may be protecting the remains against loss, breakage, and digestion damage.
Following the categorization of Bocheński et al. (1998) by breakage and digestion damage on bird remains, S. chacoensis could be considered as a category 1 (light) modifier. In contrast, taphonomic patterns on micromammal anatomical units would confirm S. chacoensis in modification category 2 (intermediate) (Andrews, 1990; López et al., 2018).
Considering the importance of knowing the agents causing the accumulation of microvertebrates in archeological and paleontological contexts, this work contributes to enhancing the development of modern analogs of greater precision at a regional scale. The detection of the Chaco owl as an accumulating agent of fossil remains is a valuable input not only to reinforce and strengthen paleoenvironmental inferences but also to provide knowledge and tools capable of being applied to ecosystem conservation policies.
Finally, it is necessary to expand the studies, including samples from different seasons of the year and from other predators in the same environment. This should allow us to clarify what has been discussed in this work.
Footnotes
Acknowledgements
We are grateful to the staff of Traslasierra National Park, specifically to the park ranger Nelson Salvi. Special thanks to Jessica Manzano-García, Paul Quintero, Nicky Quinteros, and Lucho Garay, whose contributions were essential in the fieldwork. The quality of this paper was enhanced by the valuable feedback provided by two anonymous reviewers. We would like to acknowledge the valuable comments and bibliographic contributions made by Luna Silvetti, Heimo Mikkola, Guillermo Sferco, Pablo Teta, Tracy Fleming, and Celeste Samec. Mercedes Izeta reviewed the English version of this manuscript. This work was carried out thanks to the support of National Parks and CONICET. Two institutions that are pillars of Argentine scientific development.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.