
Abstract
Based on a diverse Late-Holocene (1889–1743 cal yrs BP) small mammal assemblage (14 species, 204 individuals) resulting from the trophic activity of owls, we reconstruct the paleoenvironmental conditions of Las Chacritas area, Catamarca Province, northwestern Argentina. Twenty modern and one additional Late-Holocene samples from the region were used as comparative parameters in the paleoenvironmental reconstruction. Through direct comparison with a present-day assemblage from Las Chacritas, and using Nonmetric Multidimensional Scaling, spatial and temporal beta diversity (employing the Sorensen dissimilarity index), and the temporal beta diversity index (TBI) including all the small mammal samples, we quantitatively reconstruct the relationships among modern and Late-Holocene assemblages. Direct comparison showed significant differences, with relative abundances changing markedly over time, with several species of the Late-Holocene assemblage absent in the modern sample whereas many species frequent in the modern community absent in the Late-Holocene. NMDS analysis associated the Late-Holocene assemblage with modern samples of western drier environments. Beta diversity was low for presence-absence and moderate for abundance data, with higher contribution of turnover than nestedness, although for abundance data the proportion of nestedness was higher. TBI showed high variation among both Holocene sites and their present-day counterparts, whereas species gains dominated in modern samples. Our results suggest that the habitats surrounding Las Chacritas have modified markedly since the Late-Holocene. Presence of species today frequent in arid environments and absence of forest-dwelling species suggest that open, dry and relatively rocky habitats characterized this area in that interval.
Introduction
Small mammals have been widely used as a tool to infer changes in environmental conditions during Quaternary times due to its fossil record mostly represented by living species and that most of these species have narrow ecological requirements (Blois et al., 2010; Fernández and Pardiñas, 2018; López et al., 2016, 2020; López-García et al., 2014; Mignino et al., 2018; Tammone et al., 2014, 2020; Terry, 2010a; Terry et al., 2011). Taxonomic comparison between Quaternary small mammal assemblages and extant communities can be used to document temporal changes in diversity and relative abundance of the species as well as yield important insights about environmental changes. In northwestern Argentina (hereafter NWA) the study of several small mammal fossil assemblages has revealed noteworthy variations in the composition of these communities during Late Pleistocene and Holocene (e.g. Ortiz et al., 2011a, 2011b, 2012, 2019; Ortiz and Jayat, 2007a; Ortiz and Pardiñas, 2001). In mountainous areas of this region, Quaternary small mammal communities would have recurrently expanded or retracted their populations following the movements of the vegetation belts upward and downward the slopes derived from cold/dry and warm/humid alternating climatic pulses (Haselton et al., 2002; Ortiz et al., 2011b, 2019; Ortiz and Jayat, 2012). One of the most remarkable features shared by all these Late Pleistocene-Holocene mammal assemblages when they are compared with present samples from the same area is the loss of diversity in modern times. Indeed, with only one exception, in all these fossil assemblages the species richness is higher than in the living communities. However, a higher richness and diversity in Holocene samples compared to modern assemblages has often been attributed to time-averaging, the mixing of species that did not live contemporaneously but are found together within a single stratum (Kidwell, 2013; Olszewski, 1999; Terry, 2008, 2010a, 2010b).
In this work we studied a fossil small mammal assemblage recently recovered from Late-Holocene deposits in mountainous areas of central-eastern Catamarca, NWA, with the aim to offer a paleoenvironment hypothesis for the area. We identified the most relevant taphonomic features of the sample, characterized the taxonomic composition and the relative abundance of the species of the assemblage, and studied the pattern of temporal beta diversity change. Through comparisons of the taxonomic structure of the assemblage with other penecontemporaneous fossil sample and 20 current samples of this region, we quantify changes in the fauna of the area since the Late-Holocene. In addition, based on ecological requirements of the extant species, we use the direct comparison and the quantitative analyses to explore about the evolution of the small mammal communities and possible environmental changes over the past ~ 1800 years in this area, characterized by high geomorphological heterogeneity.
Study area and current environmental context
The fossil assemblage was recovered in Las Chacritas (Ambato Department, Catamarca Province, Argentina, 27°38′09″S, 65°57′55″W; 2100 meters above sea level – hereafter asl–), northernmost end of the Ambato range, central-eastern Catamarca, NWA. This mountain range presents a southwest-northeast orientation and extends along 150 km reaching a maximum altitude of 4500 m asl at its central portion. At its northern end the altitude of this range decreases significantly reaching 2000–2200 m asl. This zone of the Ambato range, in which the fossil site is located, is limited to the east by the Narváez range that reaches there 2500 m asl, and to the west by the Campo del Pucará, a relatively flat intermontane depression whose altitude ranges from 1650 to 1800 m asl (Figure 1). The average annual temperature in the area is 15°C, with −0.3°C the average minimum temperature for the coldest month and 28.1°C the average maximum for the warmest month. Annual rainfall varies between 383 and 482 mm (mean = 420 mm) and it is concentrated in the summer months (December to March) (d’Hiriart et al., 2015). Climatic and geomorphological features provide to this area an ecotonal nature between humid environments on the eastern slope and the arid areas located to the west of the main mountain ranges. The predominant vegetation consists of highland grasslands (Pastizales de Altura belt of the Yungas ecoregion, sensu Burkart et al., 1999), characterized by dense grasslands dominated by species of the genera Festuca, Deyeuxia, Lamphorothyrsus, Paspalum, Anthaenanthiopsis, and Stipa. Patches of Yungas montane forests (Bosque Montano belt of the Yungas ecoregion) dominated by alders (Alnus acuminata) also exist in the area at around 2.5 km from the fossil site (Figure 1). The topography of the area and the exposure of the slopes and ravines have determined the presence of these forest patches within a wider matrix of highland grasslands. Nowadays, moderate overgrazing produced by domestic cattle has partially modified the physiognomy of these grasslands, which are well developed only on the steepest slopes.
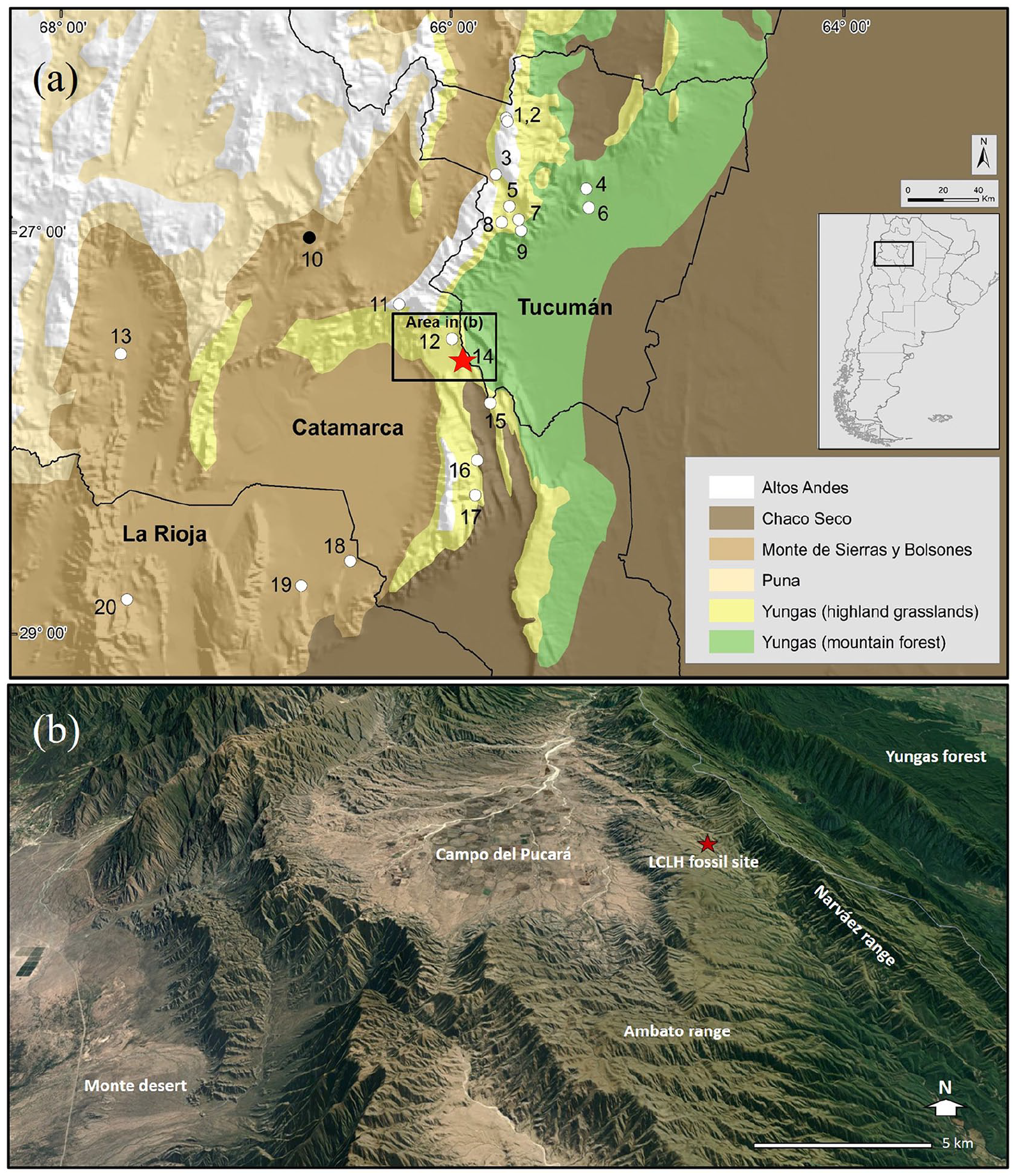
Maps of the study area in east-central Catamarca. In (a), a large portion of Catamarca and neighboring provinces of Tucumán and La Rioja is shown, with the location of Las Chacritas Late-Holocene site (star), the Late-Holocene Las Máscaras cave site (black dot) and the 20 locations at which modern owl pellets were collected (white dots) noted. In (b), a detailed map of the Las Chacritas and Campo del Pucará area is shown; the star indicates the Las Chacritas Late-Holocene site at which fossil material was collected. Numbers correspond to the following localities: 1, Lara 1; 2, Lara 2; 3, El Infiernillo; 4, Residencia Universitaria; 5, Tafí del Valle; 6, Ingenio San Pablo; 7, La Angostura; 8, La Mesada; 9, Quebrada Los Sosa; 10, Las Máscaras cave (Late-Holocene and present); 11, Las Golondrinas cave; 12, Alumbrera school; 13, Fiambalá; 14, Las Chacritas (present); 15, Cuesta de Singuil; 16, Las Juntas; 17, El Rodeo; 18, Villa Mazán; 19, La Calera; 20, Los Corrales.
The ecotonal character of the Las Chacritas area determines that many small mammal species are at the limits of their distributional range. The extant non-volant small mammal community of this area includes at least 18 species (d’Hiriart et al., 2015; Jayat et al., 2008b; Ortiz-Tempel et al., 2022) and is extensively dominated by rodents of the subfamily Sigmodontinae, represented by 14 species. There are also three species of caviomorph rodents and at least one small didelphid marsupial. Here, the montane forest is the habitat preferred by the sigmodontines Abrothrix illutea, Akodon caenosus, Akodon simulator, Oxymycterus wayku, Oligoryzomys brendae, and O. cf. O. occidentalis, while A. spegazzinii, Necromys lactens, Calomys musculinus, Phyllotis tucumanus, Reithrodon auritus, and Cavia tschudii show a marked preference for highland grasslands (d’Hiriart et al., 2015; Jayat et al., 2008a, 2008b, 2010, 2011; Ortiz et al., 2011b, 2019; Ortiz and Jayat, 2012). In grassland areas more affected by overgrazing Calomys musculinus is usually the most frequent species (Ortiz et al., 2011b, 2019; Ortiz and Jayat, 2007a).
Geochronological context
The fossil small mammal assemblage was recovered in Quaternary loessoid sediments exposed in a small area of the northern end of the Ambato range and restricted to gullies and ravines. These loessoid sediments characterize several hillside deposits and thick valley fillings and are the result of erosion/sedimentation of the preexisting loess itself accumulations as well as of thicker materials. Similar well-studied accumulations occupy a very important extension in other nearby basins located on the eastern humid slopes of mountainous systems of Tucumán and Catamarca provinces, such as Cumbres Calchaquíes, Aconquija, and Ambato ranges. According to a geomorphological evolutionary model developed by Sampietro-Vattuone and Peña-Monné (2016) for the nearby Tafí valley, 80 km further north of Las Chacritas in Tucumán Province, most of these surface formations comprises the final Pleistocene and all the Holocene, where the main outcrops are found in areas where loess was dismantled by subsequent erosion (see also Ortiz et al., 2019: 155–158). Similar outcrops dated to the final Pleistocene and the Holocene were described by Sayago et al. (2005) from Balcozna, about 40 km southeast of Las Chacritas, and by Ortiz et al. (2011a) for loessoid formations from Las Juntas, situated 50 km south of Las Chacritas.
The remains of small mammals studied here were collected from the same level which does not exceed 40 cm thick, exhumed from silt-clay sediments that were deposited during a period of slow and low energy flow conditions. The age of the Las Chacritas assemblage was established by a conventional radiocarbon dating on a unique sample of selected postcranial bones. The material provided a conventional radiocarbon age of 1930 ± 60 14C yr BP, which was calibrated to the 1889–1743 cal yrs BP interval, with a 68.5% probability range (cal AD 58 to cal AD 174).
Materials and methods
Specimens were collected by the authors in 2018 and 2019 and are housed in the Colección de Paleontología of the Dirección Provincial de Antropología (CAT-DPA-Pv), San Fernando del Valle de Catamarca, Catamarca Province, Argentina. The remains were part of a small mammal assemblage consisting of around 1500 cranial and postcranial specimens (sensu Grayson, 1984), corresponding to a minimum of 204 individuals. The material was recovered through wet sieving with a mesh of 0.5 mm according to the methodology described by McKenna et al. (1994). In order to establish the origin of the assemblage, taphonomic analysis were performed following Andrews (1990), Fernandez-Jalvo and Andrews (2016), and Fernández et al. (2017). We calculate the proportion of isolated teeth and tooth loss following Andrews (1990). Marks and surface alterations in bones and teeth caused by digestive corrosion were analyzed under microscope and categorized according to Andrews (1990), Fernández-Jalvo and Andrews (2016), Fernández et al. (2017), and Montalvo and Fernández (2019). In addition, the material was examined to detect possible post-depositional taphonomic traits such as weathering, manganese staining, carbonate crusts, root-marks, and cracking by humidity. Evidences of polishing and rounding associated to abrasion produced by water transport processes also were evaluated (García-Morato et al., 2019).
Radiocarbon dating on postcranial bones was carried out at the Laboratorio de Radiocarbono, Centro de Investigaciones Geológicas (CONICET- Universidad Nacional de La Plata), La Plata, Argentina (sample LP- 3683). The calibration was done with the program CALIB 8.1.0 on the base of the Southern Hemisphere SHCal20. 14c radiocarbon calibration curve (Hogg et al., 2020).
Twenty modern owl pellet samples generated by the strigiform Tyto furcata and one additional Late-Holocene sample, recovered from localities surrounding Las Chacritas, were used as comparative parameters in the paleoenvironmental reconstruction (Figure 1; Table 1) (e.g. d’Hiriart et al., 2015, 2017, 2021a; Gomez et al., 2012; Nanni et al., 2012; Ortiz et al., 2011a, 2011b, 2012, 2019; Ortiz and Pardiñas, 2001; Ortiz-Tempel et al., 2022). The modern samples represent small mammal assemblages of the two main ecoregions present in the study area (Yungas [Mountain forests, Highland grasslands] and Monte de Sierras y Bolsones; sensu Burkart et al., 1999) (see Supplemental File 1, available online). The additional Late-Holocene sample used in the comparison comes from Las Máscaras cave, Catamarca Province, located about 100 km northwest Las Chacritas and is contemporary to this last sample (Figure 1, Table 1; see Ortiz et al., 2012: 104). For each sample, the minimum number of individuals (MNI) was determined based on examination of the mandibles and maxillae recovered. For the paleoenvironmental reconstruction, we considered the habitat preferences of current populations of the species recorded in the Late-Holocene sample from Las Chacritas (hereafter LCLH) as well as the environmental characteristics of the areas where they are potentially found in sympatry (d’Hiriart et al., 2015, 2021a, 2021b; Jayat et al., 2008a, 2008b, 2010; Ortiz, 2003; Ortiz et al., 2011a, 2019; Ortiz and Jayat, 2007a, 2007b, 2012).
Minimum number of individuals (MNI) and relative abundance (MNI%) per species for all taxa identified in the Late-Holocene Las Chacritas and Las Máscaras cave assemblages and the 20 present-day small mammal samples.

To determine the number of genera and species in each sample, cranial and dental elements were identified through comparisons with recent and fossil reference specimens housed in the following Argentine collections: Colección Mamíferos Lillo (CML), Tucumán; Colección Paleontología de Vertebrados Lillo (PVL), Tucumán; Colección de Egagrópilas del Instituto Superior de Correlación Geológica (CEI), Tucumán; Colección Nacional de Mastozoología del Museo Argentino de Ciencias Naturales “Bernardino Rivadavia” (MACN), Ciudad Autónoma de Buenos Aires. The taxonomy of the species follows that proposed by Teta et al. (2018), with exceptions indicated in each particular case (e.g. Jayat et al., 2021). Species richness and diversity values for Late-Holocene and modern small mammal assemblages (Table 1) were expressed using the following index and estimators: number of species (S), the nonparametric estimator Chao 1, and the Shannon index (H). Diversity statistical calculations and analyses were conducted in the software PAST v.4.6b (Hammer et al., 2001).
Additionally, we quantitatively reconstruct the relationships between modern and Holocene samples through an ordination method. We developed a Nonmetric Multidimensional Scaling (NMDS) analysis using the sites as entities and the small mammal abundances as ordering variables. To reduce the effect of the dominant species on the results, the abundance value of the species was previously transformed to log 10 (to avoid the problem of the logarithmic transformations of 0 values we first added 1 to all cells of the matrix). The NMDS permitted us to order the modern sites in the high-dimensional space exploring the correlation of the NMDS scores with the small mammal abundances (correlation coefficients of the small mammal abundances were graphically presented as vectors from the origin in the multivariate space). With this analysis we search for similarities (as represented by the Bray-Curtis similarity index) between sites on the basis of the composition and abundances of the species, seeking to extrapolate modern to past climate under the assumption that more similar assemblages have more similar climates and environments. NMDS analyses were also carried out with PAST v.4.6b (Hammer et al., 2001) which implements the algorithm developed by Taguchi and Oono (2004). PAST conducts a sequence of 11 trials (one uses the results of a principal coordinate analysis as the initial condition and the others are random) from which the one with smallest stress is chosen.
Finally, to understand the patterns of species variation among sites and time, we analyzed beta diversity (changes in species diversity between different localities and changes between two different time periods for the same localities) employing a monotonic transformation of the Sorensen dissimilarity index (Koleff et al., 2003) and partitioning it into spatial turnover (ReplS) and nestedness (NesBS) components (Baselga, 2010). Each of these indices was also estimated for abundance data (Repl%diff and NesB%diff). Abundance turnover (Repl%diff) implies that the abundance of some species increases or declines from one site to another, while the abundance of other species does the same in the opposite direction. Meanwhile, for the abundance nestedness (NesB%diff) all species abundances increase or decrease from one site to another. We compared the LCLH with all other assemblages, in order to identify dissimilarities between them and to understand which mechanism determines variation in community composition. Also, we analyzed temporal beta diversity between the Holocene assemblages and their present-day counterparts from the same localities, using the TBI index (Legendre, 2019). This index measures the change in species composition between the same site at time 1 and 2, and allows to identify the sites that have changed in composition exceptionally. All analyses were carried out in R (R Core Team, 2017) using the beta.div.comp function (Legendre, 2014) and the TBI.R function available in the R package adespatial (Dray et al., 2018).
Results
Taphonomic features of the sample
All the materials of LCLH were deposited at the same level of the sedimentary matrix. The remains are well preserved and show high integrity, with very low levels of breakage. Most of the skulls were broken in some degree, but many (23%) preserved its entire anterior half including frontals and, in some specimens, parietals. Mandibles show a higher degree of completeness, most of them (44%) with all processes completely preserved. A great proportion of teeth remained in the alveoli. In maxillae, only 31% of molars were lost. Among the preserved premaxillae, 90% kept the incisors. In mandibles, molars are more frequently detached from their alveoli than incisors (molars in situ = 58% vs incisors in situ = 84%).
Examination of bones and teeth under microscope allowed to detect digestive corrosion on some incisors and molars affecting only the enamel; in the incisors the corrosion, concentrated on the extra-alveolar portion of the teeth, was slight and affected 12% of 325 incisors examined. For molars, digestion percentage does not exceed 5%.
Post-depositional taphonomic processes were null, with no evidence of weathering, root marks, manganese coatings, humidity cracking, or carbonate crusts. Polishing and rounding associated to water transport processes were absent.
Taxonomic composition
The taxonomy and abundances of small mammals recovered at LCLH assemblage are detailed in Table 1. At least 204 individuals corresponding to 14 species of small mammals were recovered, including both cricetid and caviomorph rodents (Figure 2; Table 1; see Supplemental Figure 1 and Supplemental File 2, available online). The assemblage shows a moderate codominance (based on MNI) of the sigmodontines Reithrodon auritus (27%), Akodon spegazzinii (17.6%) and Eligmodontia moreni (17.6%) with subordinate values of Calomys musculinus (12.7%), Phyllotis tucumanus (7.3%) and Phyllotis vaccarum (7.3%) (Figure 2; Table 1). The remaining species show representation percentages lower than 3%, such as Calomys fecundus (2.4%), Andinomys edax (2.4%), Necromys lactens (1.0%), and the caviomorph Ctenomys sp. (2.4%).
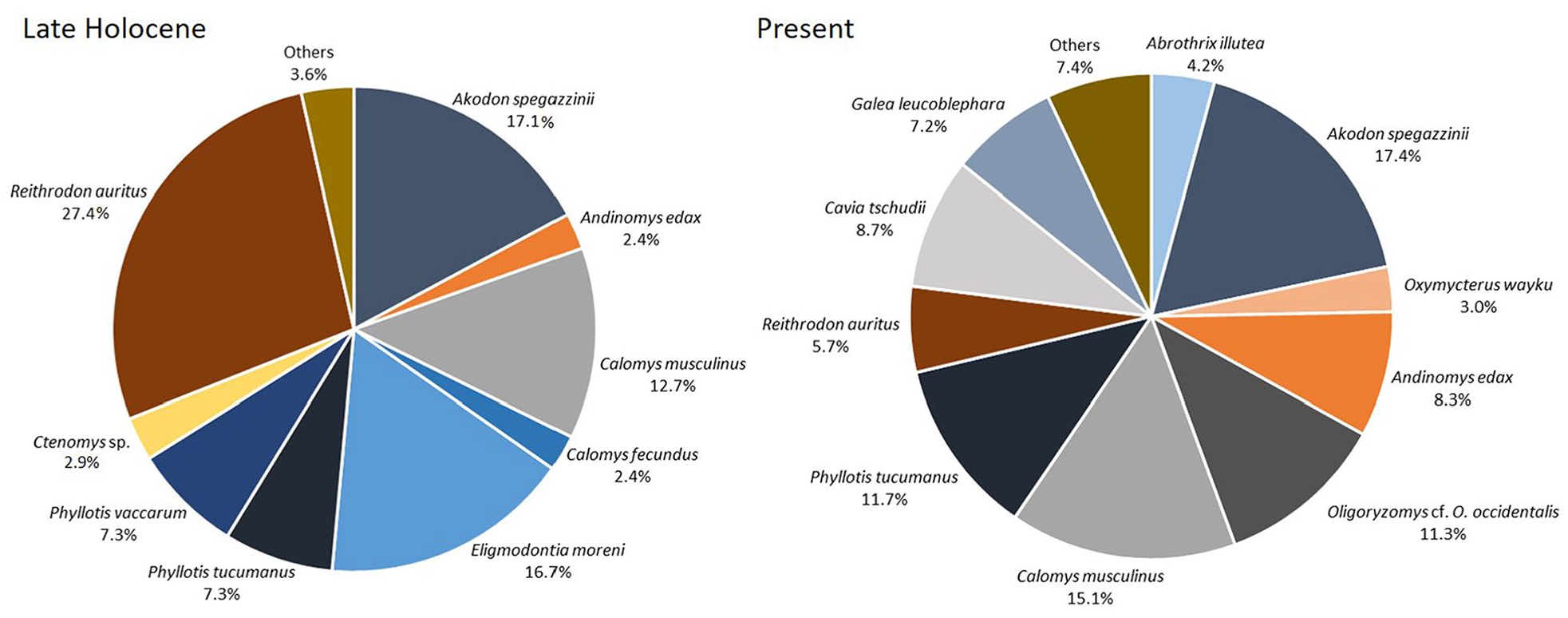
Taxonomic comparison (only for taxa with %MNI ⩾ 2.4) between the Late-Holocene and recent samples from Las Chacritas. Recent sample comes from Tyto furcata pellets.
The NMDS analysis (Stress = 0.09955; R2 Axis 1 = 0.6915, R2 Axis 2 = 0.2019) ordered modern localities mainly from east to west (Figure 3). Quebrada Los Sosa, Residencia Universitaria, Ingenio San Pablo, Tafí del Valle, La Mesada, La Angostura, Las Juntas, Cuesta de Singuil, El Rodeo, Las Chacritas, Las Golodrinas cave, Lara I and Lara II, all located on the eastern side of the pre-Andean mountain chains (from north to south, Cumbres Calchaquíes, Aconquija and Ambato ranges), were grouped on the negative values of the Axis I. Conversely, the sample representatives of localities coming from the western side of these mountain ranges (Las Máscaras cave, Fiambalá, Villa Mazán, La Calera, Los Corrales and El Infiernillo) were grouped on the positive values of this Axis. Between these two extremes is located the sample coming from Alumbrera school, which occupies an intermountain valley. The two Late-Holocene samples were located on the positive side of the Axis I, more associated with modern samples typical of the western side of these mountains, but with different relationships with respect to their present-day counterparts. While the Late-Holocene sample of Las Máscaras cave was located in the graphic very close to the current sample of this same locality, Late-Holocene and present-day samples from Las Chacritas are clearly separated (Figure 3).
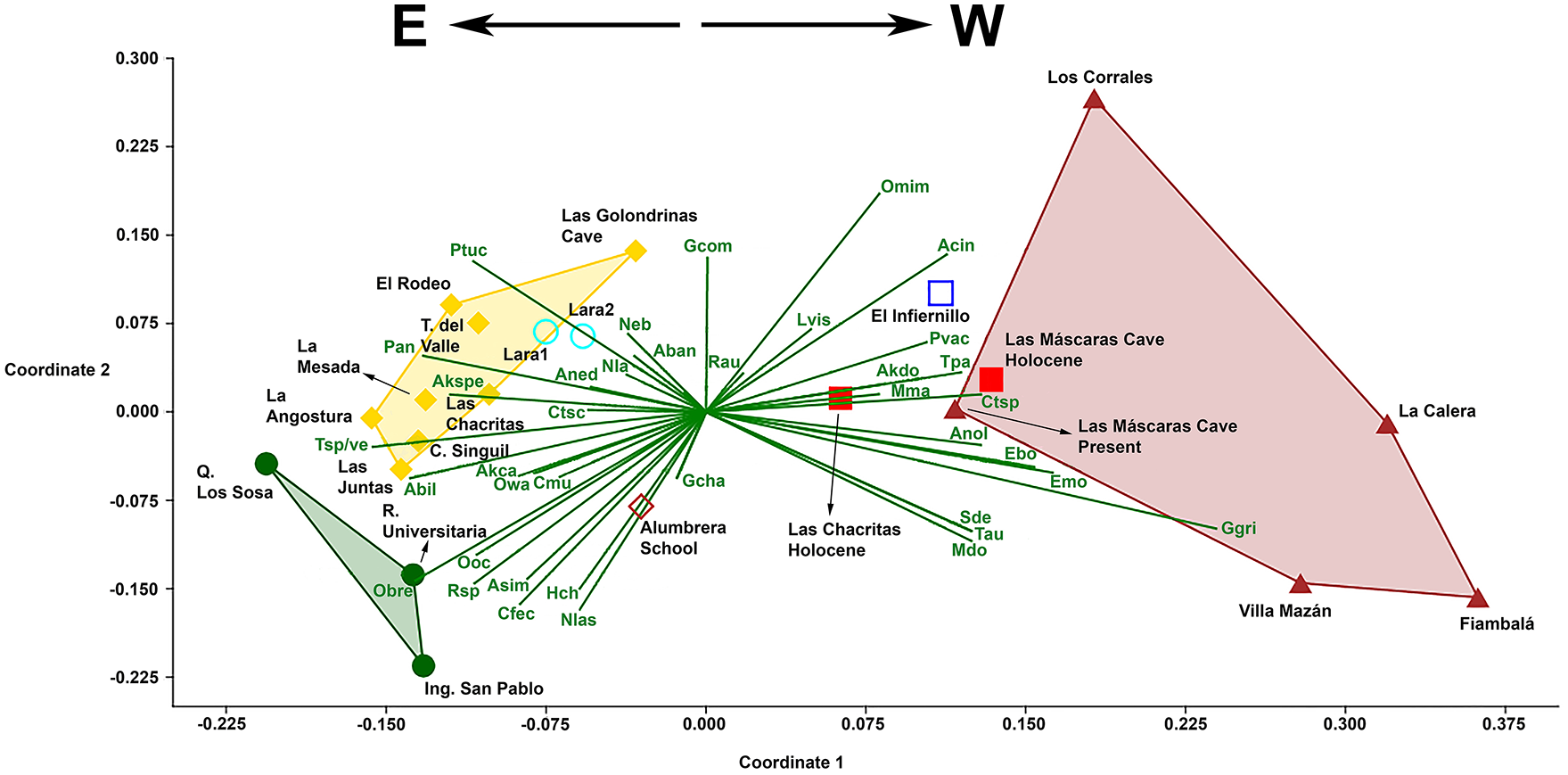
Result of the NMDS analysis. Two Late-Holocene assemblages and 20 modern owl pellet samples ordered by the small mammal abundances (correlation coefficients of the most influencial species presented as vectors from the origin in the multivariate space). Abbreviations: Aban (Abrothrix andina), Abil (Abrothrix illutea), Acin (Abrocoma cinerea), Akca (Akodon caenosus), Akdo (Akodon cf. A. dolores), Akspe (Akodon spegazzinii), Aned (Andinomys edax), Anol (Andalgalomys olrogi), Asim (Akodon simulator), Cfec (Calomys fecundus), Cmu (Calomys musculinus), Ctsc (Cavia tschudii), Ctsp (Ctenomys sp.), Ebo (Eligmodontia bolsonensis), Emo (Eligmodontia moreni), Gcha (Graomys chacoensis), Gcom (Galea comes), Ggri (Graomys griseoflavus), Hch (Holochilus chacarius), Lvis (Lagidium viscacia), Mdo (Mus domesticus), Mma (Microcavia maenas), Neb (Neotomys ebriosus), Nla (Necromys lactens), Nlas (Necromys lasiurus), Obre (Oligoryzomys brendae), Omim (Octomys mimax), Ooc (Oligoryzomys cf. O. occidentalis), Owa (Oxymycterus wayku), Pan (Phyllotis anitae), Ptuc (Phyllotis tucumanus), Pvac (Phyllotis vaccarum), Rau (Reithrodon auritus), Rsp (Rattus sp.), Sde (Salinomys delicatus), Tau (Timpanoctomys aureus), Tpa (Thylamys pallidior), Tsp/v (Thylamys cf. T. sponsorius/venustus). Green closed circles: Yungas forest localities; dark red closed triangles: arid Monte Desert localities; open blue square: semiarid high altitudinal grasslands- Monte Desert ecotone locality; red closed squares: Holocene localities; red open diamond: semi-humid transitional locality; turquoise open circles: transitional Yungas- high altitudinal humid grasslands localities; yellow closed diamonds: humid grasslands localities.
Spatial and temporal beta diversity
Beta diversity including the two Late-Holocene samples was rather low for presence-absence (D = 0.27) and moderate for abundance data (D = 0.40). For both data sets the contribution of turnover was higher than nestedness, although for abundance data the proportion of nestedness was higher (Figure 4). We compared LCLH species composition with all the other sites of the region (Figure 5). Reading the graph from left to right, as a resemblance of regional moisture increasing gradient, there is an increase of turnover among sites, with exception of Alumbrera school where turnover contribution is small. Thereafter, the values increase markedly toward sites where moisture is higher. Nestedness values were generally low (mean value = 0.15), with no clear pattern along the region, although sites La Calera, Fiambalá and Alumbrera school vary from LCLH with higher contribution of nestedness than other sites.

Overall beta diversity partitioning among presence/absence and abundance data.
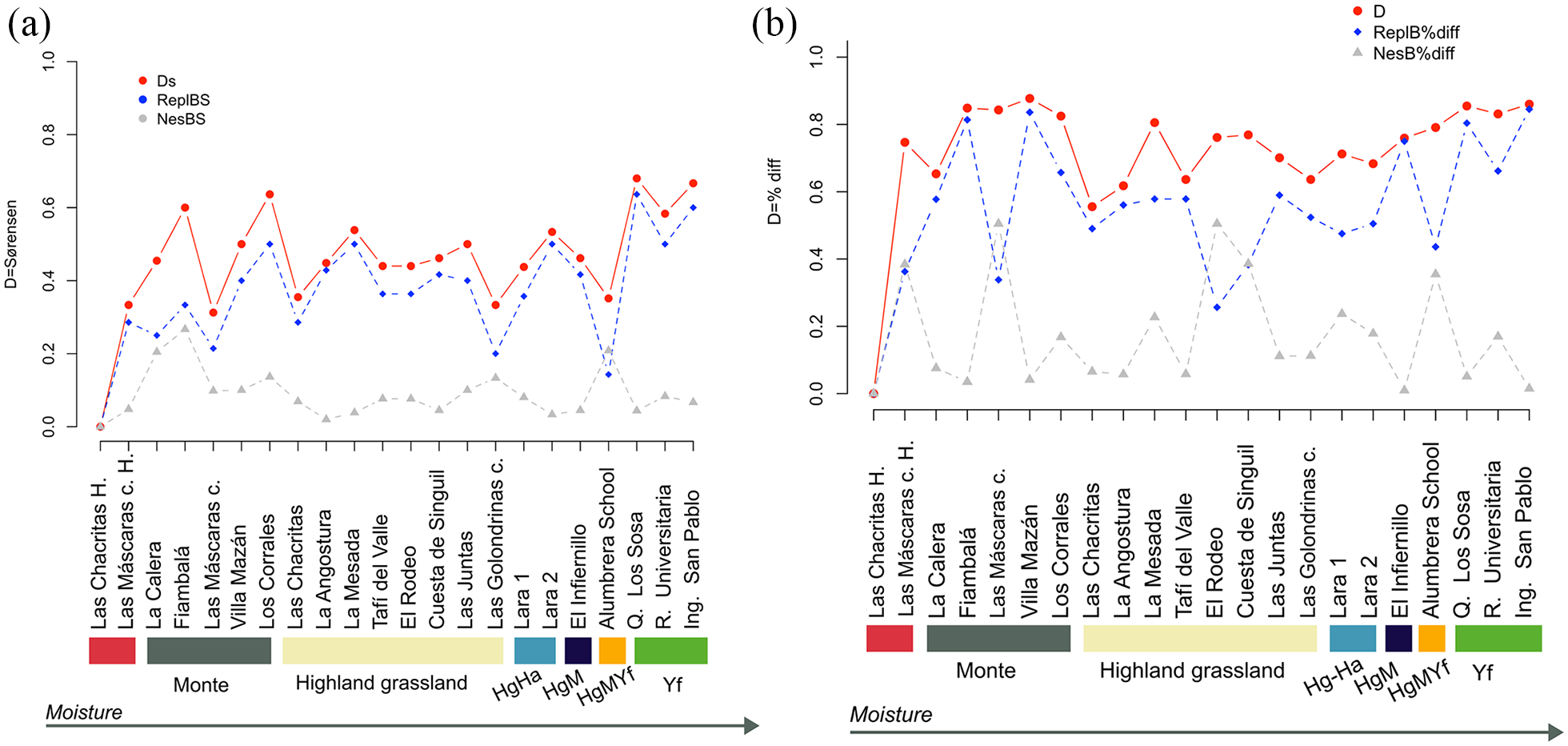
Changes in Beta diversity (Ds) of northwestern Argentina small mammal assemblages. (a) presence/absence data; (b) abundance data. ReplBs indicates species replacements while NesBs indicates nestedness for presence absence data. ReplB%diff and NesB%diff represent the same for abundance-based data.
Results obtained for abundance-based data do not show a clear variation pattern along the region (Figure 5B). In general, variation is explained mainly by turnover, except for both Las Máscaras cave assemblages (present-day and Holocene samples), El Rodeo, Alumbrera school and Cuesta de Singuil in which nestedness values are higher or quite similar than turnover (Figure 5). Temporal beta diversity index (TBI) shows high variation between both Late-Holocene sites and their present-day counterparts (Table 2). However, variation between time 1 and time 2 was significant only for Las Chacritas according to presence-absence and abundance data, but not for Las Máscaras cave (Table 2). For both sites species gains (D(gain)) was higher than species loss (D(loss)).
Temporal beta diversity (TBI) between Late-Holocene and present-day assemblages of the sites Las Chacritas and Las Máscaras cave.
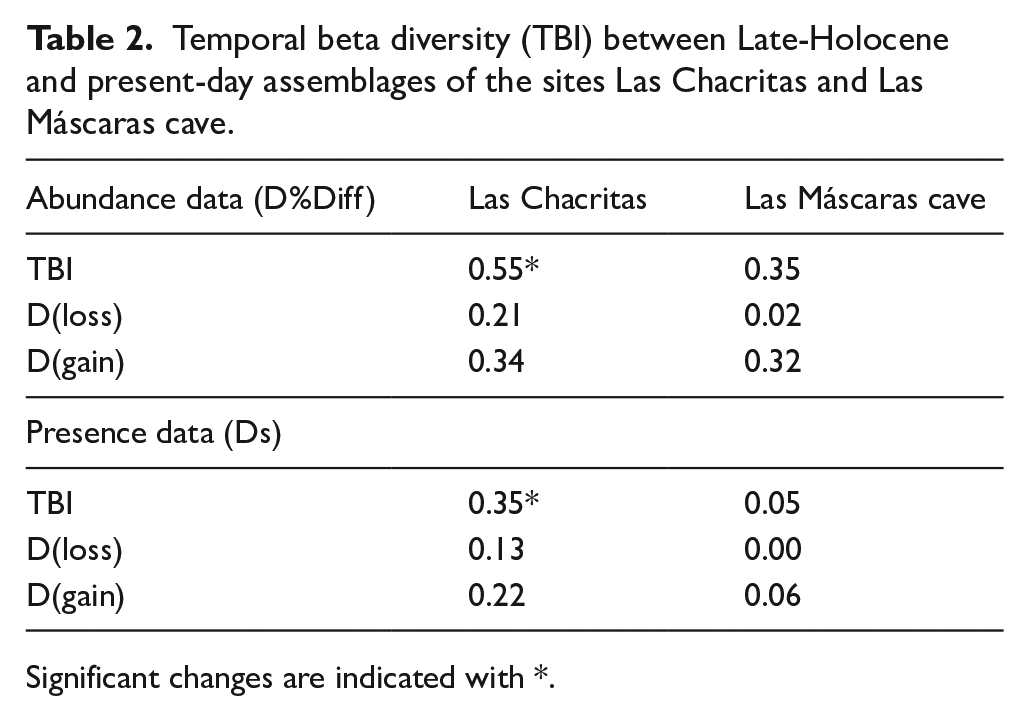
Significant changes are indicated with *.
Discussion and conclusions
The absence of weathering, abrasion or polishing signals in the remains and its high completeness and low levels of breakage suggests that the LCLH assemblage was quickly buried and that water transport processes were absent. The lack of other post-depositional taphonomic processes, such as root-marks, manganese coatings, carbonate crusts, and humidity cracking, indicates that there were no wet or damp conditions during the formation of the site. The presence of digestive corrosion on some teeth and post-cranial bones suggests the involvement of predators as the main source of production of the LCLH assemblage. The low percentages of elements affected and the slight degree of digestion of the small mammal remains indicate the involvement of a category 1–2 predator, which indicates that this assemblage was generated by some species of strigiform bird through the accumulation and fossilization of the content of their pellets (Andrews, 1990; see also Fernández et al., 2017: 41–43, and Montalvo and Fernández, 2019: 37). Since the LCLH sample has been formed by an opportunistic predator (and no transport processes have been observed), paleoecological information obtained from this assemblage should be accurate in indicating variations in the small mammal communities in response to naturally driven changes.
In NWA region, the most common owls included in category 1–2 are Tyto furcata and Bubo magellanicus. The average size of some rodent species in LCLH (e.g. Andinomys edax, Graomys griseoflavus, Reithrodon auritus, Galea leucoblephara, Ctenomys sp.) also indicates a medium to large size owl species, such as T. furcata or B. magellanicus, as the probable source of the assemblage. The involvement of this type of predator implies that the species recorded come from an area of maximum 4 or 5 km radius (Andrade et al., 2016; Andrews, 1990; Massa et al., 2015; Ortiz et al., 2019) and, in a zone with altitudinal gradients, the presence of different environments at that time is expected in the Holocene sample (García-Morato et al., 2024; López et al., 2016; Ortiz and Pardiñas, 2001).
The study of several Late Pleistocene-Holocene small mammal assemblages in NWA has revealed significant temporal variations in the diversity and composition of these communities (e.g. Ortiz et al., 2011a, 2011b, 2012, 2019; Ortiz and Jayat, 2007a; Ortiz and Pardiñas, 2001). In these areas of mountainous relief, small mammal species have recurrently modified their distributional range by expanding or isolating their populations to different altitudes following downward and upward displacements of the vegetation belts (Ortiz et al., 2019; Ortiz and Jayat, 2012). The fossil record has revealed that Reithrodon auritus and Akodon spegazzinii were dominant species in the small mammal communities along the eastern mountain slopes of the southeastern NWA during the Late Pleistocene and Holocene (Ortiz et al., 2011a, 2011b, 2012, 2019; Ortiz and Jayat, 2007a, 2012), areas that nowadays are situated close to the Yungas forest-highland grasslands ecotone.
Paleoenvironmental interpretations
Small mammals have long been widely used as paleoclimatic and palaeoenvironmental proxies (e.g. Barnosky et al., 2003; Grayson, 1984, 2000). Fossil and subfossil small mammal assemblages from Late Pleistocene-Holocene are mostly represented by living species which we know to have small home ranges and often relatively narrow habitat requirements (Blois et al., 2010; Fernández et al., 2012; Fernández and Pardiñas, 2018; Matthews et al., 2005; Ortiz et al., 2011b, 2019; Tammone et al., 2014, 2020; Terry, 2010a, 2010b), thus providing detailed information about the paleoenvironmental conditions at specific localities. Our analyses of richness and abundance revealed significant changes in the composition of the non-volant small mammal fauna from Las Chacritas area during the last ~1800 years, with both species richness and evenness higher in the living community than in the Late-Holocene. Total richness is high in the LCLH assemblage (14 species) although lower than in the modern sample from the same site (17 species) (Figure 2; Table 1), despite having been affected by time-averaging processes. Comparisons revealed that many species of the Holocene sample are absent in the current assemblage and, reciprocally, many species in the modern sample have not been recorded in the LCLH assemblage. Other striking difference between the Late-Holocene and present samples from Las Chacritas is linked to the relative abundances of several species, which have changed markedly during this interval. Exclusive species of the Late-Holocene are the cricetids P. vaccarum, C. fecundus and two species of Eligmodontia, while seven species are present only in the present-day sample: Thylamys cf. T. sponsorius/T. venustus, Abrothrix illutea, Akodon caenosus, Oxymycterus wayku, Oligoryzomys brendae, Oligoryzomys cf. O. occidentalis, and Cavia tschudii (Table 1). Among the species shared by the two samples, R. auritus shows the most conspicuous difference in relative abundance, from the highest frequency among all Holocene species to a remarkable decrease in modern times (5.68%). In marked contrast to the Holocene assemblage, dominated by R. auritus, A. spegazzinii, and E. moreni, in the current sample the prevalent species were A. spegazzinii (17.42%), C. musculinus (15.15%), P. tucumanus (15.15%), and Oligoryzomys cf. O. occidentalis (11.36%) (Figure 2; Table 1).
If we assume that the ecological requirements of small mammal species in NWA have not modified substantially over the last two millennia, these documented changes suggest that the environment surrounding Las Chacritas has modified significantly. Based on qualitative and quantitative taxonomic composition, several inferences about the habitats present at this locality around 1800 years ago can be drawn. First, the presence in LCLH sample of species today frequent in arid environments, such as Phyllotis vaccarum, Graomys griseoflavus, Eligmodontia moreni and Eligmodontia bolsonensis, suggests that open, dry and relatively rocky habitats characterized this area at that time. Second, the absence in LCLH assemblage of forest-dwelling taxa documented currently in the same area (e.g. Thylamys sponsorius/venustus, Abrothrix illutea, Akodon caenosus, Oxymycterus wayku, Oligoryzomys cf. O. occidentalis, and Oligoryzomys brendae) suggests the absence of humid Yungas forest patches which at present are developed in the area. Third, the dominance of Reithrodon auritus in the Holocene assemblage together with the relatively high representation of Akodon spegazzinii and Calomys musculinus is indicating that extensive grasslands dominated the environment. Finally, the occurrence of Calomys fecundus suggests the presence of some patches of more mesic habitats, perhaps more xeric Chacoan woodlands or shrubs in protected ravines associated with watercourses. All these evidences suggest that around 1800 cal yrs BP the area of Las Chacritas was characterized by the presence of open and arid to semiarid habitats, with persistent scattered grasslands, probably punctuated by somewhat mesic woodland or shrubland patches associated with water courses (Figure 6).
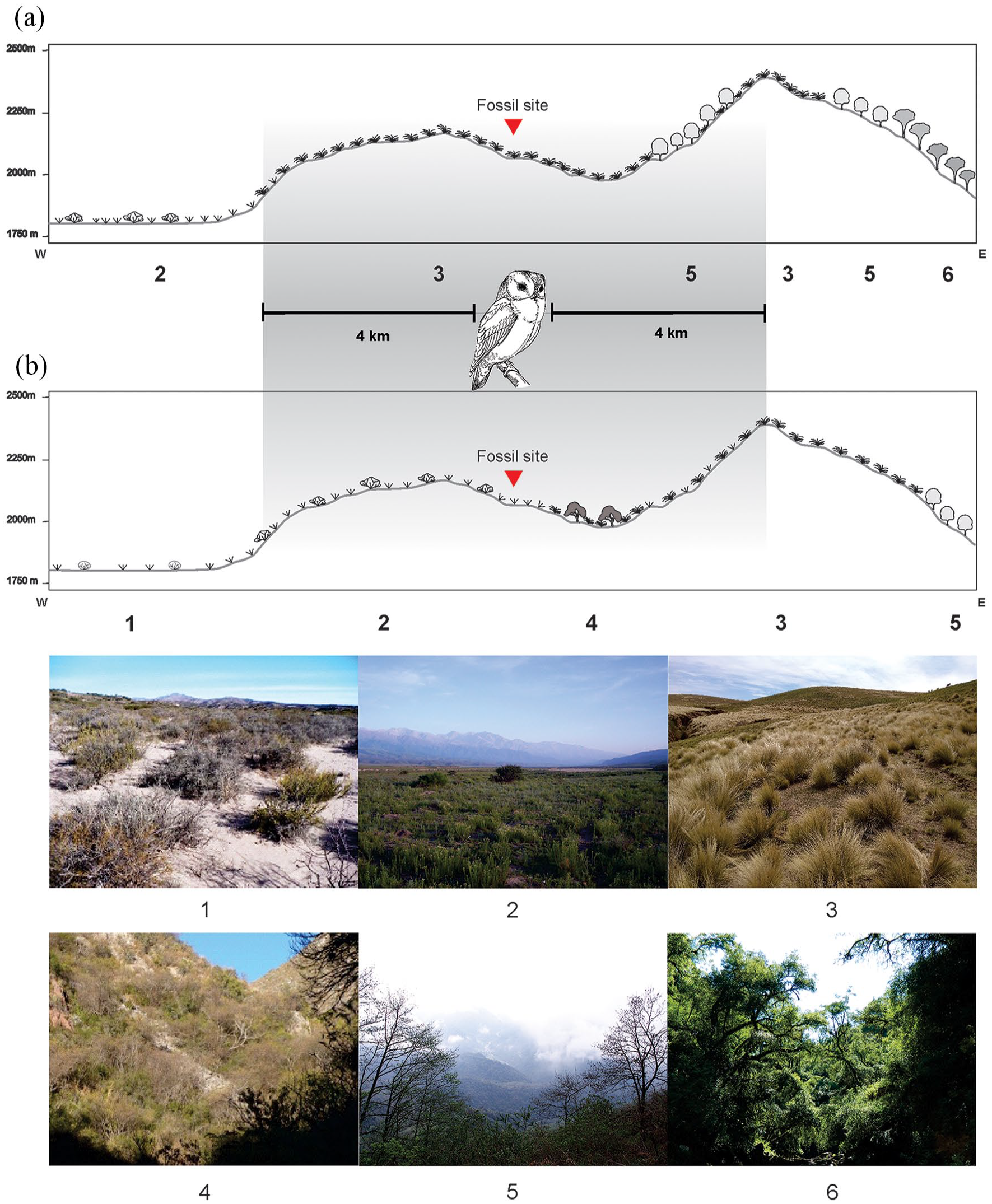
East to west schematic profile from the area of Las Chacritas (Catamarca) depicting vegetation communities and the area where the owl would have predated (indicated by the gray shading). (a) Modern vegetation communities represented by: 1, Monte desert (Monte de Sierras y Bolsones ecoregion); 2, Monte – Yungas highland grasslands ecotone; 3, Yungas highland grasslands (Yungas ecoregion); 4, Dry Chaco woodlands (Chaco Seco ecoregion); 5, Yungas mountain forests (Yungas ecoregion); 6, Yungas forests (Yungas ecoregion). (b) Environmental reconstruction for the same area around 1800 yBP inferred from the small mammals assemblage predated by the owl.
A similar conclusion can be reached on the basis of the NMDS analysis, which takes into account both the presence of the species and the distribution of their abundances. This analysis ordered modern localities mainly on a humidity and vegetation physiognomy, east to west, gradient. Localities representatives of forested areas (Quebrada Los Sosa, Residencia Universitaria, and Ingenio San Pablo), humid grasslands (Tafí del Valle, La Mesada, La Angostura, Las Juntas, Cuesta de Singuil, El Rodeo, Las Chacritas, Las Golodrinas cave), and transitional mesic areas (Lara I, Lara II), characteristically developed on the eastern side of the pre-andean mountain ranges, were grouped on the left side of the Axis I. On the opposite side of this Axis, samples representatives of localities coming from arid (Las Máscaras cave, Fiambalá, Villa Mazán, Los Corrales, and La Calera, all referable to the Monte desert [Monte de Sierras y Bolsones ecoregion, sensu Burkart et al., 1999]) and semiarid (El Infiernillo, located in a transition zone) areas located on the western side of these mountain ranges, were grouped. Alumbrera school, situated in a semi-humid transitional area, was located between these two climatic and environmental extremes. The two Late-Holocene samples clearly grouped with those nowadays assemblages coming from localities characterized by arid environments. However, while the Holocene sample of Las Máscaras cave closely matches those coming from present-day Monte desert, the LCLH sample appears more typical of the transition between highland grasslands and Monte desert areas as those observed nowadays at El Infiernillo, 100 km to the north in Tucumán Province. In this locality the environment corresponds to a wide ecotone between Yungas highland grasslands and more arid habitats typical of Monte. In this general context, the composition and abundance distribution of the registered taxa in LCLH suggests that in this area around 1800 years BP the Monte environments moved eastward, invading intermontane valleys and surrounding areas.
In the diversity analysis, overall beta diversity is mainly explained by turnover which implies the replacement of some species by others and that the abundance of some species increases from site to site in the same magnitude as the abundance of other species decreases. This replacement dominated pattern may be a result of niche partitioning, limitations to dispersal, and species sorting (Legendre, 2014; Petsch et al., 2017; Victorero et al., 2018), which could be promoted by the high heterogeneity and complexity of environments and the pronounced variation along altitudinal vegetation belts in the region (d’Hiriart et al., 2021b; Ferro, 2013; Ferro and Barquez, 2009, 2014; Sandoval et al., 2015).
Small mammal species composition of LCLH differs from other nearby sites mainly due to turnover, highlighting the temporal changes and species distributional shifts observed in previous studies for the region (e.g. Ortiz et al., 2011a, 2012, 2019; Ortiz and Jayat, 2007a, 2012). On the other hand, comparatively high nestedness values indicate an intermediate position of LCLH between hyper arid Monte desert localities with few species and an environmentally transitional site encompassing Monte desert and Yungas highland grasslands and forests environments (see Ortiz-Tempel et al., 2022 :4–5). Localities with a higher or similar contribution of nestedness than turnover have been mentioned as affected by selective extinctions, possibly due to harsh environments and reduced connectivity (Dobrovolski et al., 2012; Qian, 2009).
Variation among assemblages based on abundance is also dominated by turnover; however, in some cases, nestedness is higher or almost equal than replacement. For example, several species abundances increase from LCLH to Las Máscaras cave and Alumbrera school (A. spegazzinii, A. edax, C. musculinus, E. bolsonensis, E. moreni and G. griseoflavus), revealing higher contribution of nestedness to beta diversity among sites. This abundance change shows an increase of species which are typical of Monte desert (E. bolsonensis, E. moreni, G. griseoflavus) and highland grasslands (A. spegazzinii, A. edax, C. musculinus). On the other hand, the opposite trend is registered when comparing LCLH with El Rodeo and Cuesta de Singuil which evidence a clear decrease in species abundances (e.g. A. spegazzinii and C. musculinus) that posits both localities as a subset of LCLH.
Regarding temporal analysis, temporal beta diversity between Holocene and present-day counterparts evidenced high variation in species presence and abundance, excepting the species richness for Las Máscaras cave, where TBI values were markedly low. Space-time interaction was significant only for Las Chacritas locality. The replacement of some species (loss of species with affinity for Monte desert environments and gain of species more related to Yungas montane forest and high-altitude grasslands) indicates a conversion from an open and more arid habitat, with sparse grasslands and some scattered scrubs, to a more homogeneous landscape dominated by dense highland grassland with some patches of montane forests. This conclusion reinforces the vision of changes in environmental conditions obtained by observing variations in richness and abundance and in the analysis of NMDS.
There is very limited or null information on paleoenvironmental conditions for the study area involving the temporal interval represented by LCLH small mammals sample. Most of this limited paleoenvironmental evidence for this period in NWA comes primarily from archeological sources, sparsely distributed in the region, and indicates that around 1800 years BP more humid conditions developed at a regional level that would have lasted until around 900 years BP (e.g. Garralla, 2003; Gómez Augier and Caria, 2012; Morales et al., 2009; Olivera et al., 2004; Oxman et al., 2020; Peña-Monné and Sampietro-Vattuone, 2016, 2018; Sampietro-Vattuone, 2010). Thus, although our result of a more arid paleoenvironment based on the small mammal assemblage contradicts previous evidences, it should be considered that those proposals are quite lax, involving long temporal intervals, with low resolution, and have not considered the wide geomorphological and environmental variability of the region. It is significant to note that the Late-Holocene small mammal sample from Las Máscaras cave, dated to 1800 ± 35 yBP (layer 7 in Ortiz et al., 2012), indicated arid conditions similar to those currently developing in that area, somewhat more arid than those found later in more modern samples (layers 5, 4 and 3) of the same sequence (see Ortiz et al., 2012: 106–107). In this regard, when comparing the LCLH and the Holocene sample of Las Máscaras cave respect to their modern counterparts, it is interesting to note that while there is a notable change between the Holocene and present day samples from Las Chacritas, in Las Máscaras cave the differences between the Holocene and modern samples are negligible, which clearly indicates the inconvenience of extrapolating paleoenvironmental information from different areas in regions with extreme orographic and environmental variability.
Our results represent the only evidence that supports a more arid interval for that time of the Late-Holocene in the region. The inferred paleoenvironment for LCLH seems to be the result of climatic conditions, with minimal or no influence of human activities. Despite human settlements and associated agropastoral activities were established in this area of the NWA at least several centuries before the deposition of the fossil assemblage (Grill et al., 2013; Roldán, 2014; Sampietro-Vattuone, 2010), in the vicinity of the LCLH site there are no suitable areas for the development of extensive agriculture, allowing us to interpret our results minimizing the effects of human influence on the assemblage composition.
Our conclusions would not have been possible without the noteworthy increase in knowledge on taxonomy, distribution, ecological requirements and fossil record of small mammals of the region in recent years, which begins to show general trends on the evolution of these communities in the Quaternary and their close connection with the environmental changes (e.g. d’Hiriart et al., 2015, 2017, 2021a, 2021b; Gomez et al., 2012; Guilardi et al., 2020; Jayat et al., 2007, 2008a, 2008b, 2010, 2011, 2016, 2017, 2018, 2020, 2021; Nanni et al., 2012; Ortiz et al., 2000, 2011a, 2011b, 2012, 2019; Ortiz and Jayat, 2007a, 2012; Ortiz and Pardiñas, 2001; Ortiz-Tempel et al., 2022; Teta et al., 2011, 2013, 2022, 2023). Nevertheless, although several Quaternary small mammal assemblages were described for NWA from different periods, our knowledge about short-, medium-, and long-term changes in these communities is still incipient. Additional studies of small mammals recovered from Quaternary deposits and the use of more quantitative methodological approaches will allow us to improve our understanding on the evolution of these communities and the environments in which they developed.
Supplemental Material
sj-docx-1-hol-10.1177_09596836241236349 – Supplemental material for Late-Holocene environmental conditions revealed by a small mammal assemblage in mountain areas of Northwestern Argentina
Supplemental material, sj-docx-1-hol-10.1177_09596836241236349 for Late-Holocene environmental conditions revealed by a small mammal assemblage in mountain areas of Northwestern Argentina by Pablo E Ortiz, J Pablo Jayat, Agustina Novillo, Vanessa Torres-Carro and Franck Barbière in The Holocene
Supplemental Material
sj-docx-2-hol-10.1177_09596836241236349 – Supplemental material for Late-Holocene environmental conditions revealed by a small mammal assemblage in mountain areas of Northwestern Argentina
Supplemental material, sj-docx-2-hol-10.1177_09596836241236349 for Late-Holocene environmental conditions revealed by a small mammal assemblage in mountain areas of Northwestern Argentina by Pablo E Ortiz, J Pablo Jayat, Agustina Novillo, Vanessa Torres-Carro and Franck Barbière in The Holocene
Supplemental Material
sj-docx-3-hol-10.1177_09596836241236349 – Supplemental material for Late-Holocene environmental conditions revealed by a small mammal assemblage in mountain areas of Northwestern Argentina
Supplemental material, sj-docx-3-hol-10.1177_09596836241236349 for Late-Holocene environmental conditions revealed by a small mammal assemblage in mountain areas of Northwestern Argentina by Pablo E Ortiz, J Pablo Jayat, Agustina Novillo, Vanessa Torres-Carro and Franck Barbière in The Holocene
Supplemental Material
sj-tif-4-hol-10.1177_09596836241236349 – Supplemental material for Late-Holocene environmental conditions revealed by a small mammal assemblage in mountain areas of Northwestern Argentina
Supplemental material, sj-tif-4-hol-10.1177_09596836241236349 for Late-Holocene environmental conditions revealed by a small mammal assemblage in mountain areas of Northwestern Argentina by Pablo E Ortiz, J Pablo Jayat, Agustina Novillo, Vanessa Torres-Carro and Franck Barbière in The Holocene
Footnotes
Acknowledgements
We thank G. Ortiz-Tempel and E. Bustamante for helping us during fieldwork. S. García-Morato and two anonymous reviewers provided valuable comments on an earlier version of this paper. This work was made possible thanks to the institutional support of the CONICET, the Instituto Superior de Correlación Geológica (Universidad Nacional de Tucumán-CONICET), the Cátedra de Paleontología (Universidad Nacional de Tucumán), the Unidad Ejecutora Lillo (CONICET- Fundación M. Lillo), the Universidad Nacional de Chilecito (UNdeC), and the Instituto de Biodiversidad Neotropical (Universidad Nacional de Tucumán-CONICET).
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was partially funded by the Consejo de Investigaciones de la Universidad Nacional de Tucumán granted to PEO (PIUNT 26/G626), and Agencia Nacional de Promoción Científica y Tecnológica (PICT 2012-0050) granted to JPJ.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.