
Abstract
Ferroptosis plays a crucial role in regulating tumor growth and represents a promising therapeutic target for nonsmall cell lung cancer (NSCLC). RNA-binding protein with serine-rich domain 1 (RNPS1) has been closely associated with the development of various cancer types, but its role in NSCLC remains unclear. In this study, we used lentiviral vectors to silence or overexpress RNPS1 in NSCLC cells and then assessed cell proliferation along with ferroptosis markers, such as lipid reactive oxygen species (ROS). Bioinformatics analysis revealed that RNPS1 was upregulated in clinical NSCLC samples. Consistently, functional experiments showed that overexpression of RNPS1 promoted cell proliferation, while RNPS1 knockdown inhibited cell proliferation. Furthermore, RNPS1 overexpression attenuated erastin-triggered ferroptosis by suppressing the accumulation of lipid ROS and malondialdehyde, as well as by preventing the depletion of glutathione. Mechanistic investigations identified that RNPS1 stabilized ETS variant transcription factor 4 (ETV4) mRNA. Importantly, blocking ETV4 expression partially reversed RNPS1 overexpression-mediated suppression of ferroptosis. Collectively, these results support the notion that RNPS1 acts as a novel suppressor of ferroptosis in NSCLC progression.
Introduction
Lung cancer is one of the leading causes of cancer-related deaths worldwide, with over 2.2 million new cases and 1.79 million deaths annually, posing a threat to human health (Thai et al., 2021). In addition, the prognosis for patients with lung cancer is poor, with a 5-year survival rate ranging from only 4% to 17% (Rodriguez-Canales et al., 2016). Nonsmall-cell lung cancer represents the predominant histological type, accounting for ∼80% to 85% of all lung cancer cases (Liang et al., 2024). Despite therapeutic advances, such as surgery, chemotherapy, radiotherapy, and combined therapies for nonsmall cell lung cancer (NSCLC; Monzani et al., 2021), the prognosis for patients with advanced NSCLC remains unsatisfactory, with a low 5-year survival rate (DeSantis et al., 2014). The two most common NSCLC subtypes are lung adenocarcinoma and lung squamous cell carcinoma (Herbst et al., 2008). Consequently, recent research has focused on elucidating the molecular mechanisms driving NSCLC progression to inform clinical development. Ferroptosis, a form of cell death triggered by iron-dependent accumulation of lipid peroxides (Dixon et al., 2012), has recently emerged as a promising therapeutic target in NSCLC (Walters and Mousa, 2022; Zhang et al., 2022). Thus, investigating the regulatory mechanisms of ferroptosis in NSCLC is essential for delaying disease progression and improving patient outcomes.
RNA-binding protein with serine-rich domain 1 (RNPS1) is a key splicing regulator that promotes splicing fidelity by participating in spliceosome assembly and activating mRNA splicing (Sakashita et al., 2004; Zhong et al., 2022). Accumulating evidence indicates that RNPS1 is involved in cancer progression. For example, RNPS1 is highly expressed in cervical cancer samples and has been shown to promote cancer cell proliferation, migration, and invasion (Deka et al., 2023). Similarly, in endometrial cancer, RNPS1 knockdown inhibits tumor growth and induces apoptosis (Liu et al., 2021). Mechanically, in cervical cancer, RNPS1 contributes to tumorigenesis by ensuring the accurate splicing of pro-oncogenic genes and maintaining their mRNA levels (Deka et al., 2023). In addition, RNPS1 stabilizes NAT10 expression (Wang et al., 2024), while NAT10 suppresses ferroptosis in multiple cancers (Dalhat et al., 2023), suggesting that RNPS1 may participate in ferroptosis. Nevertheless, the role of RNPS1 in NSCLC remains unclear, and there is little research on ferroptosis-based therapy in NSCLC involving RNPS1. Through bioinformatics analysis, we found that RNPS1 is highly expressed in tumor tissues of NSCLC patients, suggesting its potential involvement in NSCLC progression.
ETS variant transcription factor 4 (ETV4), a key member of the ETS transcription factor family, plays critical roles in various physiological and pathological processes. In NSCLC, ETV4 overexpression has been shown to promote tumor progression by transcriptionally activating downstream targets such as PXN, MMP1, and MSI2 (Wang et al., 2020b). Furthermore, ETV4 knockdown induces ferroptosis in papillary thyroid carcinoma cells (Wang et al., 2021). Bioinformatics analysis predicted an interaction between RNPS1 and ETV4 transcripts, and a significant positive correlation was observed between RNPS1 and ETV4 expression levels in patients with lung cancer.
This study aimed to elucidate the role and underlying mechanisms of RNPS1 in NSCLC progression. We confirmed the upregulation of RNPS1 in NSCLC and its function in promoting tumor cell proliferation. Furthermore, we investigated its impact on ferroptosis, revealing a novel role for RNPS1 in suppressing this form of cell death. Mechanistically, we demonstrated that RNPS1 stabilizes ETV4 mRNA. Finally, rescue assays established that the RNPS1/ETV4 axis regulates NSCLC pathogenesis, highlighting its potential as a therapeutic target.
Materials and Methods
Cell culture
The human NSCLC cell lines A549 (iCell-h011) and SK-MES-1 cells (iCell-h192) were purchased from iCell (China). A549 cells were cultured in F-12K medium (iCell-0007, iCell, China) supplemented with 10% fetal bovine serum (FBS, TIANHANG, China) at 37°C with 5% carbon dioxide. SK-MES-1 cells were maintained in minimum essential medium medium (41500, Solaibao, China) containing 10% FBS under the same conditions. To induce ferroptosis, cells were treated with 5 μM erastin (Macklin, China) for 24 h, followed by subsequent analysis.
Cell infection
To generate lentiviral vectors for gene knockdown, shRNAs targeting RNPS1 or ETV4 were inserted into the BamHI/EcoRI sites of the pLVX-shRNA1 vector (BR004, Fenghui, China). The corresponding shRNA sequences are listed in Table 1. For gene overexpression, the coding sequence of RNPS1 was inserted into the XhoI/NotI sites of the pLVX-IRES-puro vector (BR025, Fenghui, China). Viral particles were produced by transfecting HEK293T cells (iCell-h237, iCell, China) with packaging vectors pSPAX2 (BR036, Fenghui, China) and pMD2.G (BR037, Fenghui, China) using Lipofectamine 3000. The viral supernatant was collected 48 h posttransfection and filtered through a 0.45-μm membrane. A549 and SK-MES-1 cells were seeded in culture plates at a density of 2 × 105 cells/well, infected with the prepared lentivirus, and cultured for 48 h before being harvested for analysis.
shRNA Sequences
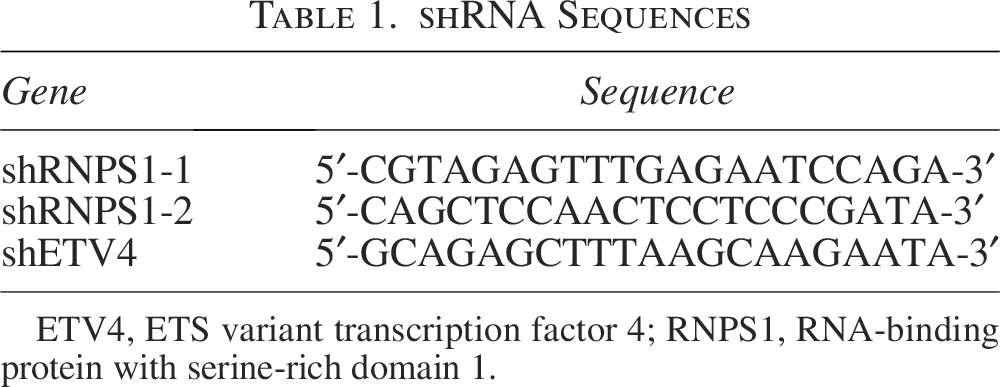
ETV4, ETS variant transcription factor 4; RNPS1, RNA-binding protein with serine-rich domain 1.
RNA extraction and quantitative real-time PCR
The total RNA extraction procedure was performed as follows: After cell collection, TRIpure lysis reagent (RP1001, BioTeke, China) was added to the cells. Following a 5-min incubation, chloroform was added, and the mixture was thoroughly inverted for homogenization. The aqueous phase was then transferred to a clean microcentrifuge tube, mixed with an equal volume of isopropyl alcohol, and stored overnight. After centrifugation for 3 min, the supernatant was carefully removed and discarded. The resulting RNA samples were dissolved with RNase-free ddH2O. cDNA was synthesized from the extracted RNA using All-in-One First-Strand SuperMix (MD80101, Medgene, China). DNA amplification was performed using primers, listed in Table 2. PCR reactions were carried out on a real-time PCR system with SYBR Green (SR4110, Solarbio, China). β-actin mRNA expression was used as an internal reference for normalization.
Primers Used for qRT-PCR

ETV4, ETS variant transcription factor 4; FL, full length; qRT-PCR, quantitative real-time PCR; RNPS1, RNA-binding protein with serine-rich domain 1.
Western blot
When cells reached ∼80% to 90% confluence, they were harvested. The cells were lysed using cell lysis buffer (P0013, Beyotime, China) containing 1-mM PMSF and incubated on ice for 5 min. The lysate was then centrifuged at 10,000 g for 5 min at 4°C, after which the supernatant was collected as the total protein extract. The protein concentration was determined using a BCA Protein Assay Kit (P0011, Beyotime, China). Proteins were separated by SDS-PAGE, transferred to a polyvinylidene fluoride membrane (ab133411, Abcam, UK), and then blocked. The membranes were then incubated at 4°C overnight with primary antibodies: anti-RNPS1 (FNab07371, FineTest, China), anti-ETV4 (220231, zenbio, China), and anti-SLC7A11 (R382036, zenbio, China). Following washes, the membranes were incubated with a goat antirabbit IgG secondary antibody (A0208, Beyotime, China). After the final wash, the membranes were incubated with ECL substrate (P0018, Beyotime, China) and visualized using a chemiluminescence imaging system (5200, Tanon, China).
Cell proliferation
Cell proliferation was assessed using the CCK-8 assay. Briefly, cells were seeded in 96-well plates at a density of 5,000 cells per well. After incubation for 0, 24, 48, or 72 h, CCK-8 solution (BS350A, Biosharp, China) was added to each well, followed by incubation for 2 h. The absorbance was then measured at 450 nm.
EdU assay analyzed by flow cytometry
Cell proliferation was further evaluated using an EdU assay analyzed by flow cytometry. Cells were seeded into six-well plates and incubated with EdU working solution (BL915A, Biosharp, China), which was prewarmed. After incubation, cells were collected, and the supernatant was carefully removed. The cell pellet was fixed by adding fixation solution for 15 min. Subsequently, cells were permeabilized by incubation with permeabilization solution for 15 min. After washing, the Click reaction was performed by incubating the cells with the Click reaction solution for 30 min. Finally, cells were resuspended in an appropriate buffer and subjected to flow cytometry analysis.
LDH release assay
The level of LDH in the cell supernatant was measured using an LDH detection kit (A020-2, Jiancheng, China). Briefly, cells were seeded into 96-well plates. Then, the LDH release reagent from the kit was added, followed by incubation according to the manufacturer’s instructions. Subsequently, absorbance was measured at 440 nm using a UV-vis spectrophotometer.
Lipid ROS analysis
For lipid reactive oxygen species (ROS) analysis, cells were seeded in plates and incubated with 2 μM BODIPY 581/592-C11 (Maokang, China) at 37°C for 20 min. After washing the cells, they were resuspended and analyzed immediately using flow cytometry.
Glutathione and malondialdehyde (MDA) analysis
Cellular glutathione (GSH) and MDA levels in cells were measured using a GSH assay kit (A006, Jiancheng, China) and an MDA detection kit (A003-1, Jiancheng, China), respectively, following the manufacturer’s protocols.
Bioinformatics analysis
The expression of RNPS1 in various cancers was predicted using the GSCA database (https://guolab.wchscu.cn/GSCA/#/). The mRNA and protein expression of RNPS1 were predicted using TNMplot (https://tnmplot.com/analysis/) and the HPA database (https://www.proteinatlas.org/), respectively. The binding of RNPS1 to the ETV4 transcript was predicted using the CatRAPID database (http://service.tartaglialab.com/page/catrapid_group).
Statistical analysis
Data analysis and graphical presentation were performed using GraphPad. All data are presented as the mean ± SD. Cell experiments were performed in three biological replicates. Comparisons between the two groups were analyzed using the t-test. Comparisons among three or more groups were analyzed using one-way Analysis of Variance. A p-value <0.05 was considered statistically significant.
Results
RNPS1 is highly expressed in NSCLC
RNPS1 was found to be upregulated in multiple cancers, including NSCLC (Fig. 1A). The HPA database showed that RNPS1 was moderately expressed in NSCLC tissues (Fig. 1B). Furthermore, by analyzing TNMPlot database, we found that RNPS1 was upregulated in NSCLC tissues compared with normal tissues (Fig. 1C). The survival analysis using the Kaplan–Meier plotter also confirmed that patients with a high expression of RNPS1 had a poor prognosis (Fig. 1D). These findings suggest that RNPS1 is upregulated in NSCLC, and its high expression correlates with poor prognosis.

RNPS1 is highly expressed in nonsmall cell lung cancer (NSCLC).
RNPS1 promotes NSCLC cell proliferation
To determine whether RNPS1 drives malignant phenotypes, we established RNPS1-overexpressing and knockdown cell lines. As shown in Figure 2A, B, RNPS1 was successfully expressed in A549 and SK-MES-1 cells. Subsequent CCK-8 assays revealed that RNPS1 overexpression promoted cell viability (Fig. 2C, D), while RNPS1 knockdown inhibited cell viability. Furthermore, EdU staining confirmed that RNPS1 enhanced cell proliferation (Fig. 2E, F). These findings establish RNPS1 as a critical driver of NSCLC cell proliferation.

RNPS1 promotes NSCLC cell proliferation. RNPS1 overexpressing and knockdown cell lines were established by lentivirus infection; then, the expression of RNPS1 was detected by qRT-PCR and western blot in
Overexpression of RNPS1 suppresses erastin-induced ferroptosis
We next investigated the role of RNPS1 in ferroptosis. Erastin is a ferroptosis inducer that acts by inhibiting SLC7A11 (Wang et al., 2020a). Ferroptosis was confirmed through the LDH release assay. The results showed that erastin increased LDH release, while RNPS1 overexpression suppressed this trend (Fig. 3A). Subsequently, lipid peroxidation was measured using the C11-BODIPY581/591 fluorescent probe (a hallmark of ferroptosis). Erastin treatment elevated lipid ROS accumulation, whereas RNPS1 overexpression reduced erastin-induced lipid ROS production (Fig. 3B, C). Given that GSH and MDA are well-established markers of ferroptosis, we further evaluated ferroptosis by detecting their levels. RNPS1 overexpression inhibited the erastin-triggered increase of MDA levels and restored GSH levels (Fig. 3D, E). Moreover, SLC7A11 is a component of the key ferroptosis regulatory system, System Xc. The inhibition of SLC7A11 enhances cellular sensitivity to ferroptosis (Wang et al., 2020a). We validated this critical ferroptosis marker using western blot. The results showed that erastin treatment suppressed SLC7A11 expression, and this suppression was reversed upon RNPS1 overexpression (Fig. 3F).
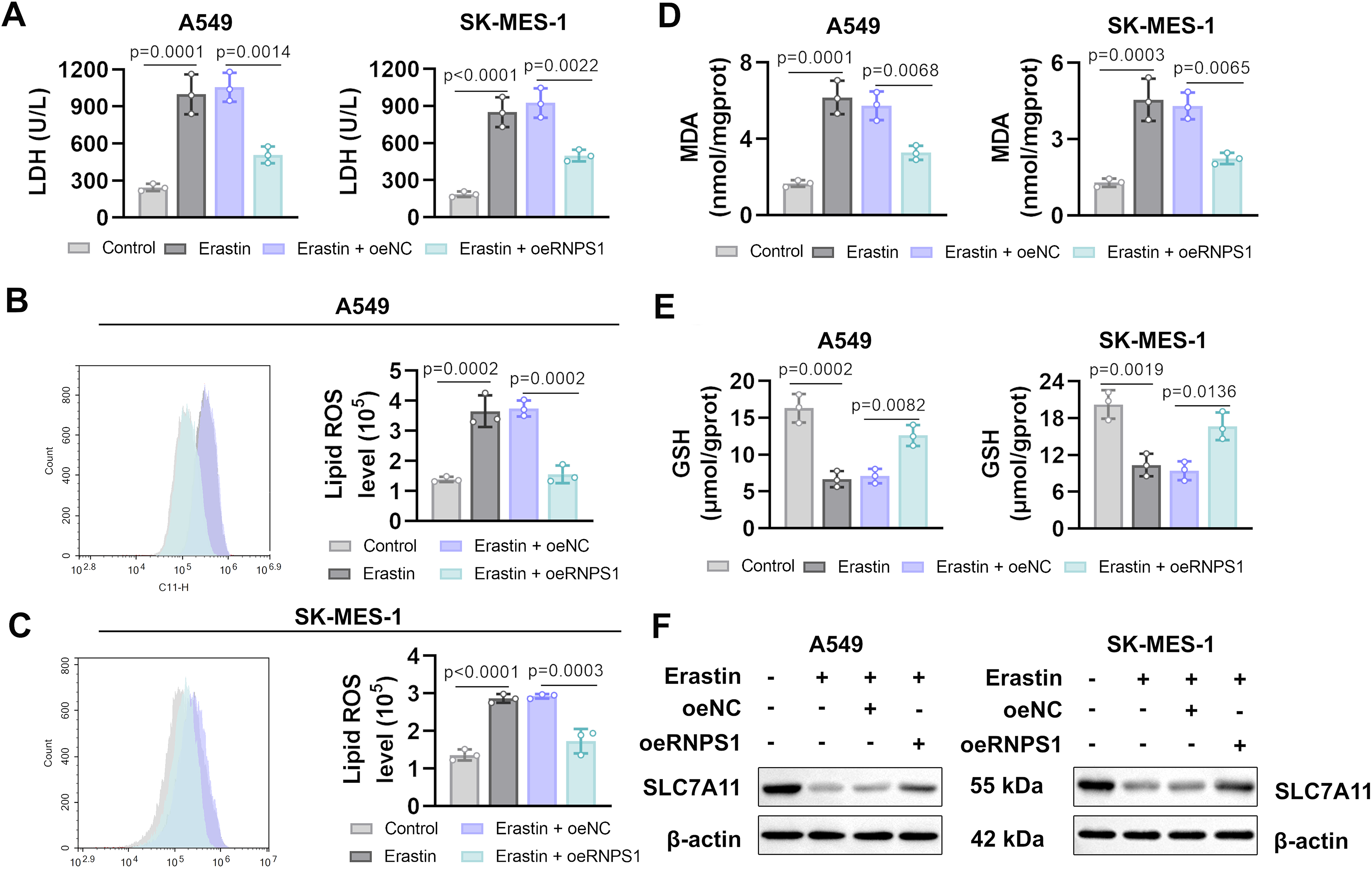
Overexpression of RNPS1 suppresses elastin-induced ferroptosis. After infection, cells were treated with 5-μM erastin for 24 h.
RNPS1 promotes ETV4 mRNA stability
Studies have shown that ETV4 promotes NSCLC progression (Wang et al., 2020b). Analysis of the GEPIA database revealed a positive correlation between the expression of ETV4 and RNPS1 in NSCLC tissues (Fig. 4A). To validate this, we overexpressed RNPS1 in cells and observed elevated ETV4 mRNA levels (Fig. 4B). In addition, previous studies have reported that SLC7A11 is a downstream target of ETV4 (Wang et al., 2021). RNPS1 overexpression increased SLC7A11 mRNA levels, indicating that RNPS1 promoted ETV4-mediated SLC7A11 expression (Fig. 4C). Using the CatRAPID database, we predicted potential binding between RNPS1 and the ETV4 transcript (Fig. 4D), leading us to hypothesize that RNPS1 might regulate ETV4 stability. To further explore the mechanism, ETV4 mRNA stability was assessed with the actinomycin D (ActD) treatment. The results showed that RNPS1 overexpression prolonged the half-life of ETV4 mRNA (Fig. 4E), indicating that RNPS1 enhanced ETV4 expression by promoting ETV4 mRNA stability. To further investigate the impact of RNPS1 as a splicing regulator on ETV4, we analyzed the expression of the ETV4 full-length (FL) transcript and known splice variants. Based on previously reported ETV4 splice variants (ETV4 Δ4, Δ7, and Δ6–8) (Cosi et al., 2023), we designed specific primers to detect the FL ETV4 transcript and these variants separately by specific RT-PCR. Our results revealed that in RNPS1-knockdown cells, the transcript level of FL ETV4 was decreased. Concurrently, the expression of ETV4 Δ4, Δ7, and Δ6–8 splice variants, which arise from exon skipping, was increased (Fig. 4F). This finding suggests that RNPS1 also ensures the splicing of ETV4 pre-mRNA, thereby promoting the production of the FL ETV4 isoform. Furthermore, we generated ETV4-overexpressing cells (Fig. 4G). The results revealed that erastin increased LDH release and lipid-ROS accumulation, which were suppressed by ETV4 overexpression (Fig. 4H, I). In addition, ETV4 overexpression reduced MDA accumulation and restored GSH levels (Fig. 4J). Moreover, erastin-induced downregulation of SLC7A11 expression was restored by ETV4 overexpression (Fig. 4K). These findings indicate that ETV4 also inhibits ferroptosis in NSCLC cells.

RNPS1 promotes ETV4 mRNA stability.
RNPS1 inhibits ferroptosis in NCSLS cells by regulating ETV4
To establish a causal link between the RNPS1-ETV4 axis and the regulation of ferroptosis, we conducted rescue experiments by knocking down ETV4 in the context of RNPS1 overexpression. Figure 5A confirmed successful ETV4 knockdown, as both mRNA and protein levels were reduced. Subsequently, cells were infected with RNPS1 overexpression lentivirus and shETV4 lentivirus, and LDH levels in the cell supernatant were measured. RNPS1 overexpression inhibited LDH levels, but the levels increased by ETV4 knockdown (Fig. 5B). Similar trends were observed in lipid-ROS levels, where ETV4 knockdown reversed the RNPS1-induced decrease (Fig. 5C). In addition, RNPS1 overexpression inhibited MDA accumulation and restored GSH levels, whereas the opposite effects were observed upon ETV4 silencing (Fig. 5D). Furthermore, ETV4 knockdown inhibited SLC7A11 protein levels in RNPS1-overexpressing cells (Fig. 5E). These results demonstrate that the inhibitory effect of RNPS1 on ferroptosis in NSCLC cells is mediated by ETV4.

RNPS1 inhibits ferroptosis in NSCLC cells by regulating ETV4.
Discussion
The incidence rate of lung cancer and its associated mortality rate has been increasing annually (Rankin et al., 2020). Despite significant advances in lung cancer therapy, the overall treatment outcomes remain suboptimal due to the disease’s complex clinical manifestations and heterogeneity (Epsi et al., 2019). NSCLC is the most common pathological type of lung cancer. Thus, it is essential to conduct in-depth research on the mechanisms underlying NSCLC progression.
RNPS1 is involved in the progression of various cancers. Functionally, RNPS1 promotes the proliferation of cervical cancer cells (Deka et al., 2023), while inhibition of RNPS1 suppresses tumor growth in endometrial cancer (Liu et al., 2021). Consistent with previous studies, we demonstrated that RNPS1 enhances the proliferation of NSCLC cells, and the knockdown of RNPS1 significantly inhibits cell proliferation.
Ferroptosis, distinct from apoptosis and necrosis, is a novel form of cell death triggered by the accumulation of iron-dependent lipid peroxides (Dixon et al., 2012). Ferroptosis holds promise for cancer treatment and may inform novel therapeutic strategies against NSCLC (Huang et al., 2021). Ferroptosis is characterized by the accumulation of lipid ROS, the lipid peroxidation product MDA, as well as GSH depletion (Stockwell et al., 2017). Previous studies have reported that RNPS1 stabilizes the expression of NAT10, and NAT10 promotes cancer progression by inhibiting ferroptosis (Dalhat et al., 2023; Liu et al., 2024). Besides, RNPS1 knockdown inhibits MAPK8 expression (Deka et al., 2023), whereas knockdown of MAPK8 promotes ferroptosis (Luo et al., 2022). These findings suggest that RNPS1 may be associated with ferroptosis. Our study found that RNPS1 reversed the ferroptotic effects of erastin. Erastin is a specific small molecule that blocks SLC7A11-mediated cystine transport (Stockwell et al., 2017). Our results confirmed that RNPS1 overexpression largely reversed SLC7A11 downregulation induced by erastin. These findings demonstrated that RNPS1 promotes NSCLC progression by suppressing ferroptosis.
Furthermore, RNPS1 possesses RNA-binding activity and regulates the expression of downstream genes by binding to mRNA, such as AURKB (Luo et al., 2022). Among the downstream factors predicted by the CatRAPID database to be potentially regulated by RNPS1, ETV4 has drawn our attention, as it has been reported to promote NSCLC progression (Wang et al., 2020b). Moreover, overexpression of RNPS1 also upregulates ETV4 expression. Subsequent half-life assessment by ActD treatment confirmed that RNPS1 enhances the stability of ETV4 mRNA. Beyond stabilizing ETV4 mRNA, our experiments demonstrate that RNPS1 acts as a splicing fidelity factor, suppressing splicing of ETV4 pre-mRNA to ensure the efficient expression of its functional FL isoform, which aligns with the established role of RNPS1 as a splicing regulator (Deka et al., 2023). It is noteworthy that the ETV4 splice variants (Δ4, Δ7, Δ6–8) upregulated upon RNPS1 knockdown. Consistent with this, in prostate cancer, the FL ETV4 isoform demonstrates functional superiority over the Δ4, Δ7, and Δ6–8 splice variants, as evidenced by its stronger transcriptional activity on oncogenic targets and enhanced ability to promote cell proliferation, underscoring their higher oncogenic potential (Cosi et al., 2023). Therefore, we speculate that the function of RNPS1 is to promote cancer by facilitating the expression of the FL ETV4. In addition, ETV4 has been found to alleviate erastin-induced ferroptosis by upregulating SLC7A11 (Wang et al., 2021). Our study evealed that RNPS1 overexpression promotes SLC7A11 expression, while knockdown of ETV4 in RNPS1-overexpressing cells reversed SLC7A11 upregulation, suggesting that RNPS1 inhibits ferroptosis through the ETV4/SLC7A11 axis (Fig. 5F).
Conclusion
In conclusion, our study identified RNPS1 as a tumor-promoting target, which regulates the ETV4/SLC7A11 axis to inhibit ferroptosis in NSCLC. These findings provide insights into the underlying mechanisms of NSCLC progression and facilitate the development of effective strategies against NSCLC.
Authors’ Contributions
L.W. contributed to conceptualization, methodology, supervision, funding acquisition, resources, and writing—review and editing. S.M. contributed to methodology, investigation, formal analysis, visualization, and writing—original draft. Y.S. and N.Z. contributed to investigation, methodology, and formal analysis. X.L. and Y.Q. contributed to investigation and methodology. H.D. and P.X. contributed to investigation.
Footnotes
Funding Information
This research was funded by the Doctoral Startup Project of Liaoning Province (2022-BS-351), Shenyang Science and Technology Program (22-321-33-60), and the Funding for Key Specialties of Liaoning Province.
Ethics Approval
This study does not involve any studies involving human or animal participants.
Data Availability
Data used in this study are available from the corresponding author upon request.
Disclosure Statement
The authors declare no conflicts of interest.