
Abstract
Adaptive variation refers to the lock-and-key type relationship between an organism and its ecological niche. This includes a patterned relationship between human behavior and societal variation. As societies become more complex, for example, our capacity to process a great deal of information quickly increases (the Flynn effect). Such adaptive processes illuminate societal variation because there is a reliable pattern in comparative research whereby behavioral differences are predicted by ecological and demographic differences. This pattern cannot be explained in terms of genetic adaptation but reflects changes mostly occurring in the lifetime of the individual. It involves an evolved capacity of humans (and other species) to adapt their behavior developmentally to local challenges. The resulting phenotypes are ecologically predictable (or “adaptive”) but may not maximize reproduction (e.g., low fertility in modern urban cities). This innovative theoretical approach is scientifically valid and heuristically useful based on comparisons across species and between societies.
Adaptive variation (see Table 1) occurs whenever anatomy or behavior changes predictably in response to varied ecological pressures. For instance, birds adopt polygynous mating systems when living in a favorable habitat with plenty of food and cover from predators but are monogamous in poorer quality habitats (Emlen & Oring, 1977). Humans exhibit similar flexibility in their mating system (Barber, 2008a). Yet, in neither case do we truly understand how such different mating behavior arises. So the precise mechanism is not an issue in defining adaptive variation for birds or humans. In each case, we find that the observed mating system is suited to a specific environment, polygyny being understood as a response to favorable territory quality, to a scarcity of males, or to a heavy burden of infectious diseases and parasites. Note that the term adaptive variation is used in preference to adaptation, a narrower term that is often reserved for species-typical traits that have a long phylogenetic history (but see O’Steen, Cullum, & Bennett, 2002; Weiner, 1995, for rapid gene-based evolution of behavior and morphology) and are attributable to gene selection (Ruse, 1982; Weiner, 1995; Williams, 1966). In contrast, adaptive variation may be recent, vary across populations, and be mediated by associative or social learning as well as genes (Bateson, 2004; Newman, 2002). In either case, the phenotype is matched to the environment so as to solve an adaptive problem. A final distinction is that adaptations increase reproductive success, whereas adaptive variation may or may not as illustrated by rising fertility in agricultural societies and declining fertility in modern cities (Barber, 2010a; Kotkin, 2012).
The Distinction Between Adaptive Variation and Adaptation.

Many animal behaviorists have no trouble accepting that when birds switch from monogamous mating systems to polygyny, this is an adaptive response to environmental variation. Yet, they might have difficulty describing equivalent shifts in the human mating system as adaptive even though it evinces a similar pattern and changes in predictable ways to solve problems that are specific to the environment, whether it is a heavy pathogen load, a scarcity of males, or intense competition over resources (Barber, 2008a).
Scientists may assume a priori that birds have an evolved genetically mediated flexibility in their mating system such that monogamy is triggered by poor habitat quality. Yet, they have trouble accepting that the same could be true of humans, preferring alternative causal mechanisms, specifically associative and social learning. In making that distinction, they ignore the fact that (a) the same basic learning processes are found in other vertebrates and (b) that learning is the basis of much adaptive change for other species, for example, rodents learn which foods to eat by observing their mothers (Avital & Jablonka, 2000; Bateson, 2004; Richerson & Boyd, 2004; Newman, 2002). Rejecting an evolutionary approach to human societies based on presumed differences in the underlying mechanisms is a case of the nature-versus-nurture fallacy. In this article, I argue that adaptive social learning partly explains some human behavioral differences between local populations and between societies. I describe examples where this approach proves scientifically useful.
A formal evolutionary approach to societal differences (or evolutionary social science; Barber, 2007) makes four basic assumptions required for a determinist theory of societies: Modern societies owe their character to an interaction of genetic (“hunter-gatherer”) adaptations with divergent modern environments (interactionism), some changes in societies are due to changes in individuals (methodological individualism), historical changes and cross-societal differences may be due to similar adaptational mechanisms (counter cultural relativism), and different social contexts modify individual development in adaptive ways (adaptive development). For alternative evolutionary approaches to this problem, including behavioral ecology and evolutionary psychology, see Winterhalder and Smith (2000).
Social Learning in Adaptive Perspective
The view that individual associative learning is adaptive is a commonplace of the animal behavior literature (Bateson, 2004). Animals become more expert at predicting and controlling biologically significant events in their environment than they would be if they relied on rigid reflexes. A role for selection is implicated by the form that classical and instrumental learning take for a given species, with humans learning to fear snakes that pose a risk due to their venom more readily than plants, for instance (Ohman, Fredrickson, Hugdahl, & Rimmo, 1976; Seligman, 1993). Social learning, or learning by observation, has similar benefits.
Social learning is a feature of many vertebrate species, including lizards and fish as well as birds and mammals (Avital & Jablonka, 2000; Richerson & Boyd, 2004). It can be operationalized as behavioral differences between local populations that are not attributable to the physical environment or explainable in terms of genetic differences.
Rats (Rattus norvegicus) were observed diving to the bottom of the Po river to take shellfish, for instance, a practice that is not seen elsewhere for this animal (Galef, 1980; Gandolfi & Parisi, 1973; other scholars prefer to describe such phenomena as “animal culture,” but this term is not particularly meaningful—except that it points to an analogy with humans, and social learning is preferred in this article as being more precise and carrying less theoretical baggage; Barber, 2008b). Social learning is adaptively important among most or all social vertebrates (Avital & Jablonka, 2000; Richerson & Boyd, 2004) and may involve tool use.
Chimpanzees learn to “fish” for termites often using a specially prepared stick that is poked into entrances in the termite hill, for instance (Richerson & Boyd, 2004, p. 104). They usually pick this skill up from their mothers, rather than by individual learning, and this interpretation is supported by the existence of varied termiting traditions. In some local communities, chimpanzees strip the bark from the termiting stick, and in others they do not. Of those that strip the twig, some use the bark to catch termites, whereas most use the stick itself (Richerson & Boyd, 2004). Another example of complex social learning in chimpanzees is cracking large nuts using stone or wooden hammers (Luncz, Mundry, & Boesch, 2012). The fact that young adult female chimpanzees commonly out migrate to other groups implies that these behavioral variants are not explainable in terms of genetic differences both because groups are genetically diverse and because migrating females sometimes adopt the foraging practices of the recipient group. In each of these examples of socially learned foraging behavior, the learner acquires access to a significant food source that would not otherwise be available, so this is an example of adaptive variation. The behavior solves an adaptive problem and varies across populations in the absence of correlated genetic differences.
Social learning may well be the adventitious result of species-wide ancestral adaptations, however. Both rats and chimpanzees learn from conspecifics in part because they are highly social. They also have varied diets, so that the capacity to learn foraging techniques from other individuals would be especially valuable and subject to positive selection. Moreover, rats exhibit food neophobia and prefer foods that had been consumed by members of their colony (Avital & Jablonka, 2000, chap. 4). Each of these characteristics can be considered preadaptations for social learning of foraging activities, including novel practices, such as diving for shellfish.
The diving feats of rats are not the only case of complex social learning for these animals, of course. In Israel, black rats (Rattus rattus) have recently branched out in a new direction by specializing on seeds from the pine cones of Jerusalem-pines. Generally speaking, animals avoid this food because it takes more energy to get at the seeds than they yield in nourishment. When the feeding animal strips seeds systematically along the conical whorl structure in which they grow, this energy equation is transformed, however. This elaborate cone-stripping technique is socially learned from the mother, and it has permitted a novel dietary specialization that completely alters the lifestyle of these animals (Avital & Jablonka, 2000). Cross-fostering experiments showed that pine cone stripping is not dependent on genetic differences and that it is transmitted socially. Experiments show that young rats (but not old rats) can acquire the complex pine-stripping sequence individually if they are given partly stripped cones, suggesting that social learning may be combined with individual acquisition in this case (Aisner & Terkel, 1992).
Social learning permits much more variation between communities than would be produced by other forms of learning. It is thus useful as a jumping-off point for an evolutionary understanding of adaptive variation in human societies with all their behavioral diversity.
Social Learning as a Different Kind of Adaptive Learning
Whereas natural selection can be thought of as a negative-feedback or error-correcting system, social learning is positive-feedback, promotes rapid behavioral change, and tends to extremes. Lacking genes as intermediaries in the selection process, social learning is selected directly based on the consequences of the behavior, and these often have profound fitness effects whether beneficial or detrimental (Avital & Jablonka, 2000). People who learn to ingest poisons by observing others doing the same may lead shorter lives, suffer from chronic illnesses, and have fewer healthy offspring, for instance. As far as we know, there are no genes that specifically promote the smoking of tobacco, although there may be numerous genes affecting vulnerability to tobacco addiction, including those that affect variation in brain receptors that respond to nicotine. Smoking is a damaging habit that derives from social learning and its persistence is all the more remarkable given that first exposures to smoking can be extremely unpleasant as the body reacts to the threatening toxins in tobacco smoke (Haberstick, Ehringer, Lessem, Hopfer, & Hewitt, 2011).
The mere fact that large numbers of people smoke tobacco was advanced as an indication of the absurdity of applying adaptationism to modern humans (Symons, 1992), but it may be too early to judge. Up to this point, much of the social learning about tobacco involved mastering its use as a pleasurable stimulant drug. In recent decades, however, we have learned to connect tobacco addiction with numerous diseases that destroy health and happiness. As soon as populations reach a high level of tobacco addiction, they are liable to associate smoking with many undesirable outcomes. Just as surely as smoking is promoted by social learning, avoidance and prevention of tobacco addiction may also be acquired by the same type of learning, even if this process is delayed. If so, smoking rates would follow an inverted U-shaped function if they are observed over a sufficient number of generations for the health consequences to be fully revealed. (Making the tobacco–disease connection requires scientific knowledge that may be unavailable to some contemporary populations where smoking rates continue to rise.)
These ideas are illustrated by changes in cigarette consumption in the United States during the last century (Figure 1) that follow an inverted U-shaped function. Cigarette consumption began at a very low level at the beginning of the 20th century, increased at a rapid pace until the 1960s, and subsequently entered a rapid decline (U.S. Department of Agriculture, 2007). The increase in cigarette consumption from 1900 on is partly attributable to increased supply as the product was machine produced rather than handmade, making it plentiful and cheap. The rise in consumption also likely reflects a social learning process in which smokers associated cigarettes with positive outcomes whether physiological (i.e., pleasure or satisfaction) or social (being successful or admired). The health risks of smoking were highlighted by research linking smoking and lung cancer. However, tobacco companies deliberately misled the public about such risks until the late 1950s when they conceded that smoking may cause cancer (Cummings, Brown, & O’Connor, 2007). Cigarette consumption peaked soon afterward and dropped by more than half over the next 40 years. This decline is consistent with social learning of the adverse health consequences of smoking.
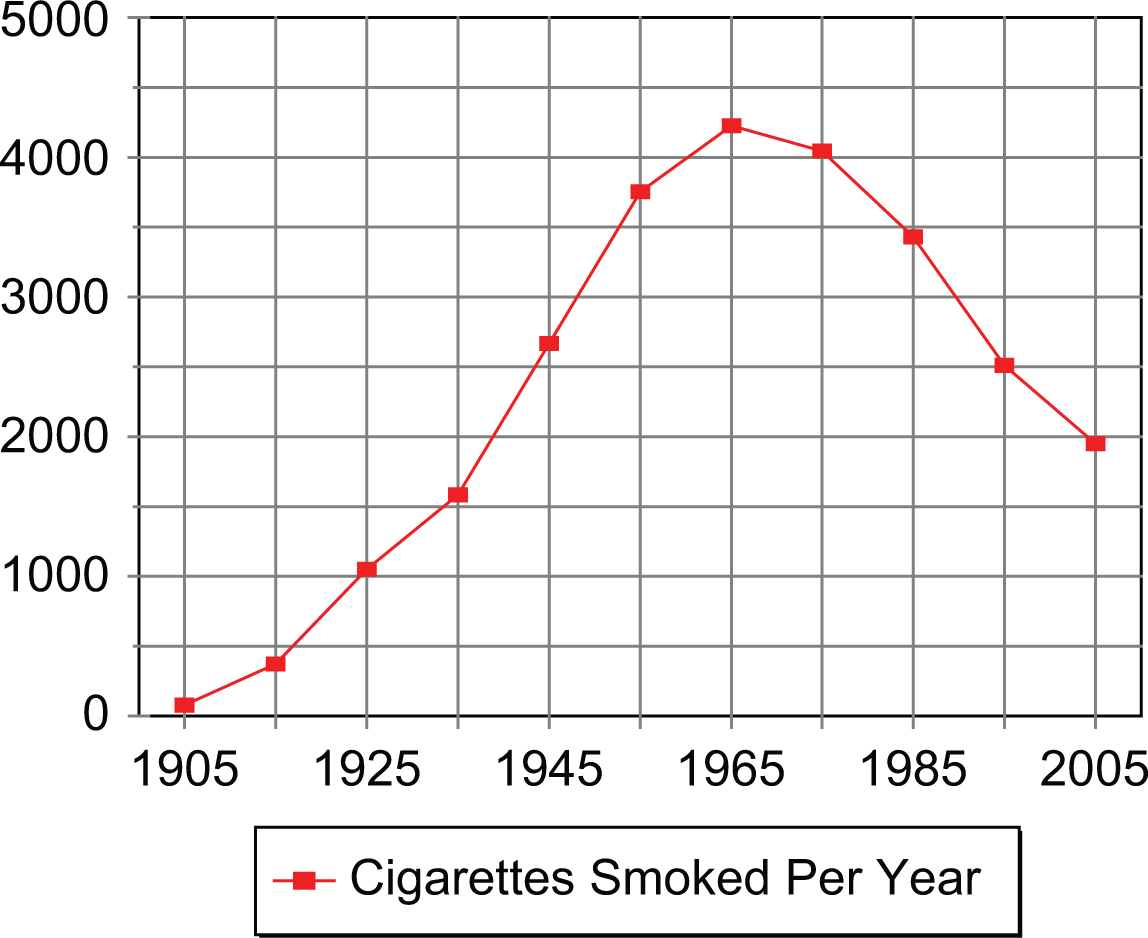
The average number of cigarettes smoked annually by Americans over18 years during the past century.
Social learning can produce dangerous and maladaptive extremes (Edgerton, 1992), but these tend to be eliminated over time as illustrated by the South African Xhosa cattle cult where religious sacrifice of the cattle herds produced a disastrous famine, a consequence that weakened the cult itself. Either the adverse consequences become evident, as in the smoking example, or the group itself is eliminated. For example, religious groups such as the Quakers that prohibit sexual intercourse dwindle in numbers until their practices disappear via natural selection. So the existence of “sick” societies does not invalidate the notion of adaptive variation across societies or within the same society over time. Indeed, these societies furnish some compelling examples of natural selection at work culling out “sick” variants that reduce Darwinian fitness.
In dealing with the problem of “sick” or maladaptive human behavior, some theorists suggest that there is an independent selection process that applies to information acquired via social learning that effectively liberates it from Darwinian selection. This meme theory of cultural selection encounters many serious problems (Avital & Jablonka, 2000; Barber, 2008b).
Even those who are sympathetic to the meme perspective doubt that there is any fundamental unit of social learning analogous to the triplet code of genetics, doubt that memes (Dawkins, 1976) can replicate themselves faithfully as genes do, and question whether memes can be observed in any way except through the behaviors they are assumed to produce, thereby raising the problem of tautology (Barber, 2008b; Richerson & Boyd, 2004). Even if cultural variants are not particulate, or separable into small independent pieces, Richerson and Boyd (2004) argue that they can nevertheless evolve through natural selection, but this view is controversial (Alvard, 2003; Flinn, 1997).
Unlike genes, memes cannot be replicated unless the behavior they represent is expressed and copied by another individual. As two critics of the meme concept (Avital & Jablonka, 2000) write,
Transmissibility is often sensitive to local conditions and variations are induced during development . . .. In non-human animals, social learning, including learning through imitation, and even instruction, does not involve the transmission of encoded or symbolic information. That is why it is impossible to decouple the transmission of information and its developmental function. Most transmission is not function-insensitive “copying.” It is reconstruction—a function-sensitive developmental process. With this type of information transmission, there is no unit of heritable variation that is not also at the same time a unit of function that is constructed during development. (p. 359)
The solitary exception here is human symbolic communication through writing and other media where ideas are transmitted without behavioral expression. With the exception of literate humans, discussing memes adds little that cannot be gleaned directly from behavioral observation. The concept of meme selection thus seems unworkable and has not led to important new discoveries in comparative research. Moreover, one does not require either meme selection or another intermediate selective system (such as genes) to produce an adaptive outcome: It may be produced by natural selection acting directly on the phenotype rather than via heritable variation.
Gene selection is a powerful error-correcting system for ensuring that an organism is adapted to its habitat, and some theorists might prefer to imagine that the capacity to respond differently in different environments is genetically encoded. It certainly may be as is likely true for the temperature-dependent sex determination of some reptiles, for example. Yet, there are many other examples where adaptive change is entirely behavioral, and this is illustrated by the Mauritius kestrel adopting the novel practice of nesting on cliffs.
The Mauritius kestrel found itself driven to the edge of extinction because its nests built in tree cavities proved vulnerable to monkeys that had been introduced to the island (Colias & Colias, 1984). Then, in 1974, one pair of kestrels nested on a cliff face that was safe from monkeys. The young prospered and bred at the same site possibly because they had developed a strong nesting preference for the place where they were raised (i.e., imprinting). The resulting tradition of cliff-nesting saved this endangered species.
Two important points can be made. First, adaptive behavioral change (or “adaptive variation”) can occur without genetic change or any other selective intermediary. Second, the Mauritius Kestrel possesses an evolved flexibility in its nesting habits. Similar flexibility is documented for other species that allows them to adapt to varied ecological conditions and thereby expand their range. The same general point applies to humans who prosper in many different habitats including extremes such as tundra, deserts, and high mountains.
Like the Mauritius kestrel, humans manifest an evolved propensity to respond to environmental factors in a fashion that promotes our chances of survival and reproduction. Admittedly, there is much controversy about whether people in developed countries strive for reproductive success (as our subsistence ancestors did; Turke, 1990) or for economic success in the modern world where fertility is often below replacement levels even after correcting for very low infant and child mortality (Barber, 2010a; Kaplan, Lancaster, Tucker, & Anderson, 2002; Kotkin, 2012; Turke, 1990). Although the demographic shift remains incompletely understood, it is nevertheless a fairly predictable pattern. This pattern smacks of adaptive variation in the sense that the modern urban environment reduces fertility due to lack of affordable living space (Kotkin, 2012) among other factors, whereas agriculture boosted fertility due to increased food production (Bentley, Goldberg, & Jasienska, 1993).
Such complex responses to environmental variation illustrate adaptive variation where the varied fertility outcomes are a predictable (i.e., patterned) consequence of environmental factors. A similar point can be made about health where increased nutrition generally improved health and reproductive success throughout human history (Turke, 1990) but undermines health due to over nutrition and obesity-related illnesses in developed countries where the population is mostly sedentary (Fontaine, Redden, Wang, Westfall, & Allison, 2003).
On balance, it seems reasonable to predict adaptive variation both across human societies and in the same society over time. The remainder of this article focuses on instances of each type of adaptive variation, and their underlying psychological and developmental mechanisms.
Adaptive Variation Across Human Societies
One of the simplest approaches to finding adaptive patterns across human societies is to look at adaptive variation across animal populations and to investigate whether similar patterning applies to our species. This method can be useful for developing testable hypotheses in comparative research although the number of topics where this approach is relevant is limited, and it can be faulted as overly simplistic. Vocal communication is one topic where cross-species comparisons seem useful.
Vocal communication of different species encounters similar technical problems related to sound transmission through space despite obstacles and interference, as well as the problems of learning and imitation. There are many striking parallels between the imitative processes of learning complex songs in birds and language acquisition for humans as indicated by differences among bird populations including local dialect variation and the decay of complex song traditions with diminished population size (Laiolo & Tella, 2007). Research also finds that the sonority of human language varies adaptively with the amount of time spent outdoors where ambient noise impedes the vocal signal in much the same way that bird song is adapted to the density of vegetation (C. R. Ember & Ember, 2007). So human language encounters many of the adaptive problems of vocal communication for other species.
Animal behavior also inspires research on human mating systems and associated variation in sexual behavior. The mating system of birds is affected by three key factors: resource availability, pathogen load, and the ratio of males to females in the population (Barber, 2008a). Emlen and Oring (1977) concluded that birds adopt polygynous breeding systems when the territory defended by a single male is of sufficiently good quality to support two or more females. When the territory quality exceeds that theoretical value (the polygyny threshold), a female sharing the territory with another enjoys greater reproductive success than a monogamous female on a territory that is below the polygyny threshold.
The productive capacity of the local ecology is related to human marriage systems also, and most polygamous societies are clustered close to the equator, where food plants produce throughout the year, whereas more strongly seasonal ecologies of higher latitudes are associated with monogamy according to cross-cultural research (Marlowe, 2007). The logic is that where there is less gatherable food, women rely more on male hunting and fishing, so that monogamy is ecologically enforced at higher latitudes.
A highly productive ecology is just one factor in animal polygyny. Females may choose to share a mate if he is of exceptionally high genetic quality. Good genes are of particular importance in mate selection for species, or habitats, where there is a high incidence of parasites and endemic diseases and where resistance to them is partially heritable (Hamilton & Zuk, 1982). Among birds, polygynous breeding systems are more likely to occur if a particular species has a high burden of parasitic illnesses. The offspring may cope with parasites better if sired by a vigorous male who passes on his resistance to them.
Low (1990) found that human polygamy is correlated with pathogen prevalence also. In societies with a high risk of illnesses such as malaria, women benefit from mating with healthy men who provide enhanced heritable parasite resistance to offspring. Men also benefit genetically from polygamous marriage because their offspring are more genetically variable and thus less likely to be wiped out by any particular parasite infestation or disease epidemic. For a wide variety of illnesses (schistosomiasis, malaria, leprosy, leishmanias, trypanosomes, filariae, spirochetes), disease risk correlates with the practice of polygamy in societies described in the Human Relations Area Files (M. Ember, Ember, & Low, 2007).
The third major reason for polygyny is the arithmetic of the sex ratio. If there is a scarcity of males in the population, monogamy does not work well for many females because they would be excluded from reproducing. M. Ember and Ember (1979) found that for species of birds and mammals with a scarcity of males, polygynous reproductive systems were more common. A scarcity of males in human populations is also associated with reduced paternal residence with, and investment in, children (Barber, 2002, chap. 9).
So each of the three main explanations for polygynous reproductive systems in other species is also supported in human comparative research. Such ecological explanations for polygamous marriage receive comparatively little attention in the social sciences where cultural-determinist views prevail, usually without rigorous empirical tests. Polygamy is attributed to economic underdevelopment or “backwardness,” to patriarchal repression of women, or to a lack of education.
Each of these explanations for polygamy was evaluated by analyzing data on 32 polygynous countries that met a criterion of at least 5% of women being in polygamous marriages (Barber, 2008a). The proportion of married women having co-wives is recorded in Demographic and Health Surveys (DHSs) that are administered to urban and rural residents in nationally representative samples. The outcome was clear. Humans engage in polygynous breeding systems under similar ecological circumstances as other vertebrates: because of male resource competition, to fight infectious diseases, and to make the most of a limited supply of males. However, cultural-determinist predictions received little or no support.
Exposure to modern media did not reduce polygamy. In fact, where women were exposed to more mass media, polygamy was higher rather than lower, challenging the economic underdevelopment interpretation (Barber, 2008a). While women in polygynous societies were less educated due to an earlier age of marriage, power inequality between the sexes did not matter either. Thus, female acceptance of wife-beating was unrelated to polygamy, implying that polygamous wives do not accept physical domination and abuse by their husbands. Perhaps the most surprising failure of a cultural-determinist prediction was that adherence to a religion sympathetic to polygamy had no influence on the prevalence of this form of marriage once the other influences were considered.
One limitation of comparative research of this kind is that it cannot specify the causal mechanisms through which the marriage system comes to match local conditions. Nevertheless, a more complete picture can be built up through analysis of related psychological variables, such as sexual motivation. If polygynous marriage is more common in societies where infectious diseases are more of a threat, then the animal behavior literature suggests correlated differences in sexual psychology.
Given that female birds are attracted to bright plumage displays in populations where there is a heavy parasite load (Hamilton & Zuk, 1982), one might predict that women would be more swayed by physical attractiveness in their choice of a mate in environments where heritable disease resistance is more important due to heavier pathogen loads. This prediction was tested, and supported, in cross-cultural research (Gangestad & Buss, 1993).
If women benefit greatly from mating with attractive men in places having a heavy pathogen load, then one would predict that they might be more interested in extramarital sex, so as to pass on disease resistance to their children. This is what Barber (2008c) found in an analysis of women’s interest in “casual” sex (sociosexuality) in 48 countries. Barber found that women—but not men—were more interested in casual sex in countries having a heavier pathogen load and that the gender difference in sociosexuality (normally higher in men) declined in such countries. This finding suggested that women might be shopping for good genes for their offspring via temporary sexual partners. Women are also more attracted to stereotypically masculine faces in places with a heavy disease load. The rationale is that facial masculinity is an index of heritable disease resistance analogous to the bright plumage of birds (Penton-Voak, Jacobson, & Trivers, 2004; Thornhill & Gangestad, 2008).
In conclusion, both the form of human marriage and correlated variation in sexual psychology are consistent with the assumption that human behavior responds to some ecological factors much as other species do. That conclusion raises further questions, however. In particular, one might ask, what are the underlying developmental, psychological, and neurological, mechanisms? Such questions do not invalidate the evolutionary interpretation, of course, because the same (largely unacknowledged, much less answered) problem haunts the relevant animal behavior research.
If ecological considerations help us understand population differences in the form of marriage, perhaps they can do the same for geographic variation in the underlying sexual psychology and behavior. The same approach might be useful for investigating the factors underlying population differences in intelligence, criminality, and other dependent variables that are correlated with sexual and reproductive behavior. Each of these topics is functionally related to mating competition and the marriage market (Barber, 2007). Thus, a marriage market adverse to women due to a scarcity of marriageable men promotes many of the conditions that social scientists label as social problems: teenage reproduction, single parenthood, crime, and social inequality, as well as challenges to somatic and psychological health (Barber, 2002). This wide range of topic areas in psychology is introduced to demonstrate the potential usefulness of the concept of adaptive variation in comparative research.
The Marriage Market and Adaptive Variation in Premarital Sexuality
The form of marriage varies adaptively across societies as discussed above. Even within the same form of marriage, there are signs of adaptive variation. Sexual behavior varies from one predominantly monogamous society to another, for instance. In some societies, women almost never have sex before marriage as was true of European Americans in 1900, just 8% of whom had sexual intercourse by age 19 years (Caplow, Hicks, & Wattenberg, 2001) or women in Beijing in 1989, just 15% of whom had intercourse before marriage (Larson, 2012). In others, they may seek, or accept, sexual intimacy on a first meeting as illustrated by the contemporary phenomenon of “hooking up” on U.S. college campuses (Bogle, 2008).
Why is there so much historical and societal variation in female sexual behavior, as well as that of men? Evidently, young people adapt their dating/mating strategies to their social milieu and, specifically, to variation in local marriage systems or dating dynamics. This point can be illustrated by variation among college campuses. Cashdan (1993) found that college women’s sexual behavior varied depending upon what they expected from men in their dating pool. The shallower that men were seen to be, and the less women expected out of them in terms of emotional commitment, the more short term their own perspective became. In a dating environment perceived to be full of “cads” (i.e., sexual opportunists), women dressed provocatively and initiated many brief sexual relationships. If they encountered more “dads” (men offering emotional commitment and marriage) than cads, they behaved differently, emphasizing their own chastity and sexual fidelity.
Cashdan (1993) found that college men were similarly flexible in their dating behavior. Cads attracted women by emphasizing their physical appearance and sexuality. Dads promoted themselves by emphasizing their capacity to do well in school and their desire for a permanent relationship, that is, advertisements of their capacity for paternal investment in children. Such evidence implies that men and women adjust their dating behavior to that of the other gender. This phenomenon suggests a mechanism through which sexual behavior varies from one society, or community, to another, accommodating varied levels of paternal investment. If paternal investment prospects in a country are high, for example, premarital sexuality declines, and single parenthood is reduced (Barber, 2003).
Dating on college campuses has declined in the years since Cashdan’s (1993) study, possibly replaced by hooking up, or casual physical intimacy of varying intensity that lasts for a short period, perhaps a single evening (Bogle, 2008). Such casual sex would appear to have analogs in the anthropological literature, involving societies as diverse as the Muria of India (Symons, 1979), the Trobriand Islanders of New Guinea (Malinowski, 1929), and the Quechua of South America (Morales, 1995), although the emphasis is mostly on adolescent sexuality where females are quite unlikely to conceive.
Hooking up may reflect the comparatively safe environment of college campuses and other settings where most students are linked by similarity of background and by extensive friendship networks (Bogle, 2008). This would explain why hooking up is practiced more on college campuses than in other settings. College campuses may facilitate hooking up, but this alone cannot explain why it is so common today but occurred so rarely in the past. One novel factor behind this change could be a declining proportion of males to females, or the sex ratio, on college campuses (Bogle, 2008). On college campuses with relatively more females, women are more likely to be sexually active but less likely to have steady boyfriends (Uecker & Regnerus, 2010). Whether on campus or off, a scarcity of men liberates sexual behavior, specifically among women as they adjust their sexual behavior to the proclivities of men who are in greater demand. Human sexual behavior—and gender differences therein—is evidently sensitive to contextual factors, including the availability of mates or spouses. Such variation seems adaptive in the sense that it facilitates successful competition over desirable partners in different milieus (Cashdan, 1993).
Important as the sex ratio may be to marriage markets, it is just one of various interacting factors that contribute to the likelihood of marriage. For instance, the impact of the sex ratio is mediated by economic conditions. Men whose wages are too low are generally disqualified from marriage as illustrated by declining marriage rates among African Americans with declining wages for unskilled labor (Wilson, 1997). This helps explain why single parenthood is more heavily concentrated among low-income women (Abrahamson, 1998; Barber, 2005a, 2007). Similarly, in predominantly agricultural societies, ownership of sufficient land is a key qualification for marriage (Voland & Engel, 1990). The number of men who are economically qualified for marriage (i.e., land owners) may thus be substantially lower than the number of single men of appropriate age for marrying.
So there is abundant evidence of adaptive variation in sexual behavior across societies, across time, and across socioeconomic groups, among other potential influences. Further evidence of adaptive variation can be found by looking at the developmental mechanisms underlying such societal variations. This research strategy is illustrated by developmental influences on violent crime, and intelligence test scores, as well as sexual behavior—three topics that are generally considered separately by psychologists but turn out to be functionally linked to reproductive competition (Barber, 2007).
Developmental Mechanisms
Intelligence
Adaptive variation is defined in terms of a match between phenotype and environment such that changes in behavior from one society to another tend to increase reproductive success, or to enhance some outcome that is (or was) correlated with reproductive success or inclusive fitness, such as accumulation of resources or acquisition of higher social status (Turke, 1990). Identifying such patterns in comparative research implies that the relevant phenotypes vary in adaptive ways. Strictly speaking, we do not need to understand the developmental processes whereby the phenotype gets matched up with the social environment. Yet, grasping the relevant developmental mechanisms strengthens the case for adaptive patterning (or “adaptive design”). It is also a fruitful approach to hypothesis testing in comparative research as recognized by cross-cultural researchers for many decades (Barry, Child, & Bacon, 1959; Berry, 1967; Whiting & Whiting, 1975).
Human behavior is not always adaptive, of course (Edgerton, 1992), but fitness-decreasing actions are not challenging to the concept of adaptive variation: They may represent natural selection at work weeding out maladaptive variants. Indeed, many of Edgerton’s “sick societies” were already in a steep population decline, implying that their maladaptive actions were being selected against. For example, the slaughtering of their cattle by the South African Xhosa tribe inspired by a child’s religious vision brought on a devastating famine.
Other examples of societal sickness such as the loss of boat building technology by Tasmanian Islanders are not nearly so devastating to an adaptationist approach as commonly assumed (Henrich, 2004). One explanation is that when their community was cut off from the mainland, the population size was too small to sustain transmission of complex skills (Barber, 2008b). A closely similar phenomenon is found for song birds where song complexity declines in small isolated communities compared with their parent populations (Laiolo & Tella, 2007). So what are often presented as text book examples of the failure of adaptation specific to human societies can actually illustrate natural selection at work in ways that apply to other species.
Once adaptive patterns are established in cross-societal comparative research, it is worth asking whether these societal differences can be explained in terms of adaptive variation in child rearing and other developmental phenomena, such as early nutrition or exposure to modern electronic technologies (in the case of intelligence). An evolutionary approach to societal variation in intelligence and other outcomes is bolstered by a variety of well-established developmental and causal mechanisms. In addition to individual learning, and social learning, these include adaptive variation in brain development.
The central evolutionary mechanism of gene selection is highly controversial when applied to human group differences (including variation in intelligence). Yet gene-based natural selection produces adaptive local biological differences in skin color, body build, digestive enzymes, disease resistance, and so forth (Cavalli-Sforza, Menozzi, & Piazza, 1996), and there are also geographic differences in behavior (as indexed by personality tests; Allik & McCrae, 2004) that might be products of genetic selection (Fincher, Thornhill, Murray, & Schaller, 2008; Nettle, 2006).
In addition to any genetic variation, there is abundant evidence of gene–environment interactions affecting intelligence (as measured by IQ tests). Adaptive variation in brain development may reflect ancient mammalian mechanisms that track the level of danger in the environment, or food availability, or level of maternal emotional closeness, and so on. Stress hormones play a key role in mediating such adaptive flexibility in development (Barber, 2007; Kalinichev, Easterling, Plotsky, & Holtzman, 2002; Teicher, Andersen, Polcari, Anderson, & Navalta, 2002). This helps explain why stressful rearing conditions associated with low social status are correlated with lower intelligence, although verbal stimulation and positive interactions are also reduced, suggesting an environmental impoverishment effect (Hart & Risley, 1995).
IQ scores reflect variation in the capacity of the human brain to process complex information quickly and accurately. Variation in IQ scores in different social groups provides a quasi-experiment assessing the impact of varied environmental inputs on cognitive ability. The possibility for adaptive variation in IQ scores is consistent with copious animal research on environmental enrichment boosting cognitive ability (Rosenzweig, 1996). Such research demonstrates that the information-processing capacity of the mammalian brain is enhanced as more demands are placed on it for dealing with complex novel information (Barber, 2005b, 2006a; Rosenzweig, 1996).
This animal behavior background points to predictions about societal differences in information-processing ability or intelligence. Intelligence would increase in a society as the cognitive demands of that society increased. Consistent with this prediction, intelligence rises with increased importance of literacy as societies move away from agricultural production and become increasingly urbanized (Barber, 2005b, 2010b). The economic driver here may be the great value of literacy for keeping records in a mercantile economy (Justman & Gradstein, 1999). Conversely, in societies where agriculture remains important, parents invest relatively little in the education of their children, often preferring to keep them at home from school where they can provide needed labor for the household economy (Admassie, 2003). One thus finds that national IQ scores increase as agriculture declines in importance (Barber, 2005b). There is good evidence that this is partly due to increased formal education (Ceci, 1991).
Copious evidence shows that IQ increases with years of education whether one compares individuals or nations (Barber, 2005b; Ceci, 1991). Education may thus provide an advantage in economically developed urban societies where the ability to process a lot of printed information quickly and accurately is a key marketable skill. (Interestingly, generalized problem-solving increases more in developed countries than does crystallized intelligence, suggesting that education is not the only factor here; Dickens & Flynn, 2001.)
The fact that parents invest in children’s education in urbanized societies more than they do in agricultural ones (Admassie, 2003; Justman & Gradstein, 1999) probably reflects an effort to train children in ways that promote social success in their own society (Low, 1989). In addition to such specific targeting of desirable skills through education, exposure to an information-rich environment likely enhances information-processing capability for humans as it does for other mammals (or vertebrates more generally, Greenfield, 1998). Conversely, exposure to childhood stressors may reduce academic capacity and performance and impair development of social skills (Barber, 2000a; Delaney-Black et al., 2002). This phenomenon likely reflects an adaptive flexibility in brain development, given that dangerous conditions inhibit exploration and reduce opportunities for development of cognitive skills (Barber, 2007).
Neuroscientists now recognize that the brain is quite flexible, even in adult life, so that well-educated people, whose brains are therefore more active, resist degenerative diseases of the brain associated with aging and actually live longer (Molla, Madans, & Wagener, 2004). This lifelong modifiability of brain function is adaptive in the formal (or gene-centered) sense that it derives from an ancient mammalian capacity to respond to varied environmental inputs in ways that promote survival and reproduction. For instance, a stressful early life makes mammals more responsive to stressors as adults (Kalinichev et al., 2002), and this likely helps the individual to survive and reproduce in dangerous environments where there are many predators, or where intraspecific competition is severe, or vital resources are scarce. Mammals also enhance their information-processing capacity in response to the demands of the physical and social environment in ways that are clearly relevant to human variation in intelligence. For instance, laboratory animals raised in more stimulus-rich environments have improved learning ability (Rosenzweig, 1996).
If the environments of nonhuman mammals vary significantly in the amount of cognitive challenge, the same is even more obviously true of our species, particularly in modern contexts. Arguably, we live in a more stimulus-rich environment than that experienced by our ancestors, although partially isolated from the sensory inputs of the natural environment due to modern housing, the use of dogs for hunting, and other factors that shrink sensory processing areas of the brain (Groves, 1999; Stephan, Frahm, & Baron, 1981). Brain stimulation increases rapidly with major changes in communication technology, such as the emergence of printing, radio, cinema, television, and the Internet (Barber, 2006a).
Generalizing from the environmental enrichment effects for other species, one would expect intelligence to increase with the development of modern electronic media. Consistent with this view, very rapid increases in IQ scores occurred in developed nations over the past century (known as the Flynn effect; Dickens & Flynn, 2001; Flynn, 1987). In some cases, these amount to a rise of more than 30 IQ points (or two standard deviations). It is likely that some of this change is due to improved health and nutrition that accompanied economic development (Martorell, 1998). In addition, intelligence test scores likely increased due to greater information-processing demands of negotiating a modern urban way of life (Greenfield, 1998), increased years of formal schooling (Ceci, 1991), and increased exposure to modern electronic technologies (Barber, 2006a; Carter, 2001; Mielke, 1994; Okagapi & Frensch, 1994).
Barber (2006a) found that reading and math achievement in a country increased with exposure to mass media including television and computers as well as newspapers—school achievement being strongly correlated with IQ scores (Lynn & Vanhanen, 2002). Moreover, children’s daily use of mass media statistically explained the fact that reading scores were higher in economically developed countries. Students use of information technology, particularly television, and computers, evidently sharpens their intellectual skills allowing them to perform better in school.
In conclusion, one finds that the Flynn effect resembles the environmental enrichment effects established for other species. This looks like adaptive variation in information-processing capacity from one society to another and with varied stimulation levels within a given society. By acquiring cognitive skills that are useful in a technologically advanced society, we may grow rusty in other capacities, however, including sensory acuity and sensitivity to events in the natural environment, such as the ability to track wild animals and such changes are accompanied by a reduction in specialized sensory regions of the brain (Groves, 1999; Stephan et al., 1981).
Clearly, there are many developmental mechanisms underlying adaptive variation in IQ scores, or information-processing capacity more generally, and these can be investigated in comparative research. Given that societies vary widely in the propensity to express damaging violence, one might imagine that societies exhibiting a lot of violence—specifically violent crime—might expose their children to different developmental experiences compared with those growing up in less criminally violent places.
Violent Crime
Measurement of violent crime is plagued by technical problems having to do with biased reporting and inaccurate record keeping (Neapolitan, 1997). This problem is exacerbated by somewhat different definitions of crimes around the world. Despite these problems, much of the variance in serious crimes such as murders, rapes, and assaults is systematic, implying that the error is small enough to permit meaningful comparative research (Barber, 2000a).
Although a great deal remains unknown about the development of violent criminal behavior, psychologists have now amassed considerable evidence about developmental correlates of crime (Barber, 2007). These factors are conveniently organized at two levels of complexity. At the lower level, a pattern of coercive parenting that involves low empathy, scolding, corporal punishment, withdrawal of affection, and so forth promotes the development of antisocial tendencies of delinquency, precocious sexuality, drug use, and academic under-performance (Barber, 2000a; Lykken, 1995; Rowe, Rogers, Meseck-Bushey, & John, 1989). At the higher level of complexity (i.e., communities, socioeconomic groupings, ethnic groups, countries), this pattern of parental behavior is elicited by factors associated with a weak marriage market from a women’s perspective, including low sex ratios (i.e., scarcity of males; Barber, 2000b, 2004 a, b, 2006b; Walsh, 2003), poverty, high male unemployment, high single parenthood ratio, and so forth (Barber, 2009). Of course, the marriage market is not the only factor in interpersonal aggression. Warlike societies socialize children for aggression through physical privation, for instance (C. R. Ember & Ember, 1994; Field, 2001). Whichever level of complexity is analyzed, violent crime arises indirectly from the antisocial behavior that the relevant conditions produce.
Antisocial tendencies may be an adaptive response to difficult or uncertain social environments (Barber, 2000a; Rowe et al., 1989). Under such conditions, a more cooperative disposition is a disadvantage because agreeable individuals get taken advantage of by peers having a more selfish, or exploitative disposition. Uncertain conditions also favor earlier reproduction because later reproduction is selected against by high mortality (Barber, 2000b; Draper, 1989; Ellis et al., 2003; Quinlan, 2003).
Violent crime may be interpreted as one manifestation of adaptive variation in antisocial tendencies in response to family background and insensitive parental behavior. This perspective diverges from the more mainstream view that coercive parenting is inherently dysfunctional. This mainstream perspective seems reasonable in view of the harm caused to children and to their communities after they mature. Yet, the dysfunctional perspective is not supported, given that insensitive parenting resists most efforts to change it, implying that it is predictably elicited by particular social environments.
Even after parents in disadvantaged communities are alerted to the adverse consequences of coercive parenting and are trained in more empathic behavior, their use of corporal punishment and other insensitive practices does not change (Nightingale, 1993). This suggests that different parenting practices are elicited by divergent social environments with the more stressful conditions of low-income neighborhoods eliciting more corporal punishment and less empathy compared with a middle-income neighborhood, for instance (Barber, 2009; Hart & Risley, 1995). To the extent that the outcomes of these varied parental practices shape adults who are better suited to their specific social environment, they constitute adaptive variation.
Sexual Behavior
Both IQ scores and violent crime rates manifest adaptive patterns in comparative research, and these patterns reflect differing developmental responses to varied rearing environments. There are many aspects of human sexuality that also suggest adaptive patterning and for which the relevant developmental mechanisms may be investigated. These patterns cut across different measures of sexual behavior, including the single parenthood ratio, premarital sexuality, teenage childbearing, divorce rates, number of lifetime sexual partners, and more subjective measures such as interest in casual sex (or sociosexuality; Barber, 2007; Guttentag & Secord, 1983).
One key dimension of sexuality is the extent to which different societies, or different social groups, experience extramarital, or premarital, sexual intercourse. Most evolutionary scholars begin from the assumption of Symons (1979) that sexuality is a female service provided to men—although this assumption may need to be revised in the light of female sexual behavior being much freer, following the widespread use of contraception in developed countries as well as the entry of more women into paid employment and careers. Such secular changes alter the cost–benefit analysis for low-commitment sexuality given that risks of unwanted pregnancy are reduced and that women rely less upon economic support from husbands and are therefore less vulnerable to reputational damage that reduces their desirability as brides (Barber, 2008c; Bogle, 2008; Klinenberg, 2012).
In sexually restrictive communities, women may withhold their sexual services until after marriage (Dickemann, 1979). By doing so, they make themselves more desirable as brides on the assumption that men seek partners who provide a high confidence of paternity (Alexander, 1979; Symons, 1979).
Perhaps the best evidence that free sexual expression weakens marriage is to be found in the extraordinary efforts made to control sexuality in some societies. Free sexual expression is treated not just as a threat to a specific marriage, but to all marriages, and is resisted not just by husbands or wives, and their families, but by the entire community (Dickemann, 1979; Strassman, 1996). Sexually restrictive societies (or societies of honor) impose stiff penalties for single parenthood and adultery. In such societies, there is a relative scarcity of brides, as indicated by a negative correlation between the sex ratio and approval of casual sex in cross-national studies (Barber, 2008c; Schmitt, 2005).
At the opposite end of the sexual behavior continuum are less restrictive societies where premarital sexuality is common, single parenthood ratios are high, and divorce rates are elevated, and so on. Such relatively unrestricted societies are characterized by an oversupply of brides relative to grooms and this correlation is explainable in terms of women competing for the romantic attention of men by offering greater sexual intimacy (M. Ember, 1974; Guttentag & Secord, 1983). The rationale is that in such an environment, men are in greater demand than women and therefore set the rules of dating relationships to suit their own interest in advancing the time when sexual intimacy occurs in the relationship. In other words, men pursue sex partners rather than brides (a strategy that would be almost impossible in sexually restrictive societies).
Whereas Guttentag and Secord (1983) emphasized the supply of prospective brides relative to grooms as the key determinant of sexual restrictiveness in a society, the level of premarital sexuality (and weakness of marriage) is affected by several other factors. These include reduced familial supervision (Klinenberg, 2012); economic conditions (with recessions working against new household formation and marriage; Abrahamson, 2000); female labor participation with more participation delaying, and hence weakening, marriage (Barber, 2005a); contraceptive use with effective contraception increasing premarital sexuality and delaying marriage (Stevenson & Wolfers, 2007); and the high cost of raising children in cities reducing the attractiveness of marriage and childbearing, thereby reducing marriage rates and increasing non-marital sexuality (Kotkin, 2012).
How does sexual behavior develop so differently in different societies? The key theoretical question to ask about such variability is whether it reflects situational pressures like the impact of claustration and chaperoning that may be interpreted as efforts by kin to control and manipulate women’s fertility (Dickemann, 1979) or developmental variation partially attributable to hormonal effects early in life (Ellis et al., 2003).
In addition to efforts by authority figures to prevent sexual expression, female sexuality is affected by psychological and hormonal influences on psychological development. Although a detailed account of such influences is beyond the scope of this article, at least three deserve mention. These are as follows: the probable effects of father’s absence during childhood (Ellis et al., 2003), the impact of abusive parenting (Barber, 2000a), and what may be termed the female challenge effect (i.e., hormonal effects of female–female competition; Archer, 2006).
If female children do not have a co-resident father, or father figure, they are predisposed to precocious sexuality, consistent with the pattern of environmental insecurity advancing the age of sexual activity and increasing mating effort for both genders (Barber, 2000a; Ellis et al., 2003; Quinlan, 2003). This pattern of social behavior is often discussed in pathological terms because it is associated with high rates of violent crime (particularly for males) and other social problems. Yet, the pattern can be interpreted as an instance of adaptive variation, as already mentioned.
Research on children of abusive parents reveals a similar pattern of short-term or low-investing sexuality that is partly attributable to the impact of stress hormones on the developing brain (Barber, 2007; Teicher et al., 2002). Psychosocial stressors, such as high population density, produce similar effects on aggression and reproduction for other mammals (Kalinichev et al., 2002).
The male challenge effect refers to an activational effect of hormones rather than a developmental effect, strictly speaking. According to the male challenge hypothesis, when a territorial male is challenged by a rival and succeeds in repelling the rival, there is increased production of testosterone and other androgens. This has the effect of increasing both the aggressiveness and the sex drive of the victorious male (Archer, 2006). Although psychologists are cautious about accepting that hormones have much of an impact on human social behavior, Archer (2006) presents a great deal of evidence that high testosterone production is correlated with victory, high social status, and sex drive for humans, and it also correlates with marital problems (Booth & Dabbs, 1993; Mazur & Michalek, 1998).
Cashdan (2008) reviewed evidence that competition among women boosts sex hormones as well (in addition to stress hormones). She also showed that increased mating competition (operationalized as a scarcity of men of reproductive age relative to women) increases hormone production as inferred from widening of the waist due to increased abdominal fat.
Women’s interest in casual sex in a country increases as the marriage market becomes more competitive (Barber, 2010b; Schmitt, 2005). This implies that women respond to reproductive competition (and also economic competition; Cashdan, 2008) with an increase in sex drive and, possibly, an earlier onset of sexual activity (Barber, 2003, 2005a). Conversely, if there is a relative excess of single men, women delay sexual activity until after marriage, thereby enabling them to marry on favorable terms (Guttentag & Secord, 1983). It should be clear that the challenge hypothesis, when applied to women, has a developmental component—as reflected in altered body shape and age of beginning sexual behavior—as well as an activational one. The same argument can be made for males and for other species, where the development and intensity of bodily displays are hormone dependent.
Controversial though these conclusions may be, they suggest numerous lines of inquiry for comparative researchers. Human sexuality is expected to vary considerably around the world in response to varied ecological conditions, and these differences may be large enough to swamp gender differences (Barber, 2008c) as noted previously. So human sexuality varies adaptively in response to social and ecological influences, and such variation can reflect developmental influences, such as the impact of psychological stress on brain development and sexual motivation.
Conclusion
The concept of adaptation can be applied to phenomena such as the change in size of equine leg bones over tens of millions of years (Ruse, 1982), but it is less useful when applied to human behavior. Indeed, it is doubtful whether one can point to even a single example of a human behavioral adaptation that evolved over millions of years via gene selection (Symons, 1992) and that project is complicated by the various hominid species arising and going extinct on such large time scales.
Yet, there is abundant evidence for adaptive variation in human behavior. Such adaptive variation is revealed in cross-societal comparisons, including adaptive developmental differences. Such adaptive variation is mostly, or entirely, independent of changes in gene frequency over time and reveals itself in predictable associations between ecological and social environments and behavioral phenotypes as humans adjust to ecological pressures in predictable ways, just as other species do. For example, residents of urban environments have much lower fertility than residents of agricultural communities, thanks to the varied costs and benefits of raising children in these different ecologies (Barber, 2010a; Kotkin, 2012).
Such adaptive patterning was illustrated in comparative variation for phenomena as varied as IQ scores, the form of marriage, violent crimes, and patterns of premarital sexuality. Adaptive variation is thus a useful approach to organizing comparative data and generating testable hypotheses in the field. By adaptive patterning, I mean the predictable match between ecological influences (broadly construed) and phenotypes that is not to be confused with the narrower concept of adaptation where such patterning is due to gene selection.
One of the simplest approaches to devising testable hypotheses from the perspective of adaptive variation is to generalize from animal behavior and to ask whether the same ecological factors predict variation across human societies as is established for ecological research on other species. One example of this approach developed above was the impact of ecological factors on the degree of polygyny in the marriage system. Another involved analysis of brain responses to the complexity of environmental stimulation and the Flynn effect in respect to IQ variation resembles environmental enrichment experiments on animals. Note that this research strategy can be heuristically useful even if the underlying developmental mechanisms are different or unknown. Once such patterns are established, they provide a rationale for comparative research on developmental variation from one society to another, as illustrated by environmental enrichment effects, the form of marriage, and sexual behavior.
Although comparative researchers have long tested functional evolutionary hypotheses, work in this and related fields lacked a clear theoretical or empirical picture of adaptive variation, both of which this article set out to provide. Research was cramped by a view of adaptation drawn from comparative anatomy that was too restrictive to deal with human behavioral variation (both by requiring an extended evolutionary time frame and by over emphasis on gene selection).
Human behavior does not always maximize Darwinian fitness, of course. One of the most intriguing examples of this is the low fertility characteristic of developed societies. Yet, one finds that modern behavior is quite predictable in terms of adaptive variation. Whereas agricultural societies convert increased food production into enhanced fertility, and have increased family size compared with foragers, the opposite occurs in urban societies where food is even more plentiful but it is much more costly to raise children in monetary terms and monetary constraints evidently win out (Kotkin, 2012). In effect, family size is responding adaptively to resource availability and the costs and benefits of reproduction in these varied ecologies, although differing resources are limiting in each case.
Modern societies are unusual in generating a fertility level that is well below replacement. Such low fertility is nevertheless predictable as a patterned response to urbanization and economic development (Barber, 2010a). This pattern is adaptive in the sense of being a predictable response to ecological variation, but it is not an adaptation because it reduces reproductive success rather than increasing it. That is why one is much better off analyzing these phenomena in terms of adaptive variation (or comparative patterns) rather than in terms of fitness maximization and Darwinian adaptation that cannot account for sub-replacement fertility.
Modern fertility is constrained by various unique modern factors, and this phenomenon is analogous to birds reducing clutch size in response to diminished food availability in some years (Lack, 1968). The difference, of course, is that reducing clutch size allows parent birds to successfully raise small clutches rather than wasting their parental investment on larger clutches if most of the offspring would perish in a year of diminished food availability. This strategy contributes to lifetime reproductive success. It is an adaptation. Low fertility for urban humans is not an adaptation. As a predictable response to ecological variation, it is identifiable as adaptive variation, however.
The approach to comparative research outlined in this article does not reject Darwinian adaptation as a viable approach to human societies but rather insists upon appropriate theoretical modifications to preserve the usefulness of adaptive explanations, particularly to modern societies where these had been most problematic (Turke, 1990). My approach differs from evolutionary psychology by playing down genetic determinism and differs from behavioral ecology in its focus upon modern societies rather than hunter gatherers (Smith & Winterhalder, 2000). It differs from mainstream social science in rejecting cultural determinism (Barber, 2008b). The main theoretical changes it requires are as follows:
Recognizing that gene selection is just one of many mechanisms through which human behavior becomes adaptively matched to different ecological or social conditions. Others include individual learning of different kinds and social learning. In some cases, adaptive change occurs without any change in gene frequency for other species as well as for humans. This principle is widely accepted by animal behaviorists, of course (Avital & Jablonka, 2000), but it is under appreciated in many Darwinian accounts of human behavior.
Accepting methodological individualism whereby changes to individual psychology and behavior can be observed as differences among nuclear families, local communities, or entire societies. For example, young people are sexually active before marriage in societies where there is a scarcity of marriageable males and are sexually restricted in their psychology and behavior where there is a scarcity of young females. Such variation may be at least partly mediated by psychological adjustment of the individual to the social environment of families, communities, and societies.
Abandoning narrow cultural relativism so as to permit investigation of phenomena that generalize across societies, or even across species. Instances of the latter were discussed in this article such as polygyny intensity and the Flynn effect construed as a brain enrichment phenomenon. Such counter cultural relativism goes against the grain of much social science, but it is empirically successful and therefore a fruitful avenue for further inquiry.
Appreciating that maladaptive behavior does not invalidate a research agenda based on adaptive variation. Perhaps the biggest theoretical difficulty faced by comparative researchers is the fact that human behavior in modern societies does not maximize reproductive success, leading some scholars to question the value of Darwinian adaptationism in modern societies (Edgerton, 1992; Symons, 1992). Yet, Edgerton’s “sick societies” can be thought of as maladaptive societies that are in the process either of changing to more adaptive alternatives, or sinking into reproductive decline and being removed by natural selection. The precipitous decline in fertility in advanced urban societies—some of which are at less than half replacement levels (Kotkin, 2012)—can be viewed in the same terms and it may be that developed societies are so hostile to childbearing that they are destined to fail. This problem is often falsely interpreted as an issue that is unique to our own species but that is a matter of perception. The fact that human societies change so rapidly and generate so much variation means that maladaptive variants are far more salient. Conversely, animal societies seem to be much better adapted to their ecological niche. Yet, over the grand sweep of evolutionary time, far more species failed to adapt and went extinct than persisted to this day.
Some scholars might lament the loss of theoretical tools, such as mathematical modeling, that are predicated on gene selectionism and its social learning analogs (Richerson & Boyd, 2004). What this approach gains in theoretical rigor, it frequently loses in empirical applicability, and there are few, if any, instances of complex human behavior in modern societies that are satisfactorily illuminated in these terms (Barber, 2008b; Buss, Haselton, Shackelford, Bleske, & Wakefield, 1998; Symons, 1992).
Finally, it may be objected that whereas gene selectionism is of limited heuristic usefulness, referring to adaptive variation suffers from the opposite problem, of being over-inclusive, thereby raising the specter of tautology. There are two valid responses to this objection. The first is that the adaptive variation approach does produce genuinely falsifiable hypotheses, several of which were discussed above (e.g., the possibility that electronic technologies would render people intellectually lazy and reduce their intelligence). The second response is that the approach developed in this article benefits from its heuristic usefulness for identifying adaptive patterns that generalize across species as well as across societies and is thereby capable of stimulating a great deal of research.
Footnotes
Acknowledgements
I am grateful to the reviewers for exceptionally constructive and detailed comments on an earlier draft of this article.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Author Biography
![]()