
Abstract
Introduction
ADHD is the most common neurodevelopmental disorder, with a prevalence rate of 1.4% to 3.0% in children and adolescents worldwide (Thapar & Cooper, 2016). ADHD is highly heritable and multifactorial; multiple genes and non-inherited factors contribute to the disorder (Akutagava-Martins, Rohde, & Hutz, 2016; Thapar & Cooper, 2016; Thapar, Cooper, Eyre, & Langley, 2013). Prenatal and perinatal factors have been implicated as risks (Botting, Powls, Cooke, & Marlow, 1997; Braun, Kahn, Froehlich, Auinger, & Lanphear, 2006), but definitive causes remain unknown.
Evidence from pharmacological studies (Greenhill, Halperin, & Abikoff, 1999), genetic studies, particularly those on the dopamine transporter (DAT) genes (Cook et al., 1995), and imaging studies (Dougherty et al., 1999; Ernst, Zametkin, Matochik, Jons, & Cohen, 1998; Krause, Dresel, Krause, Kung, & Tatsch, 2000) strongly suggests that the dopaminergic system is involved in ADHD. Recent neuroimaging studies continue to contribute to emerging models of the pathophysiological mechanisms underpinning ADHD. Early structural magnetic resonance imaging (MRI) studies found a significant reduction in gray matter volume in the basal ganglia (putamen, globus pallidus, and caudate nucleus) in line with frontostriatal models of ADHD pathophysiology, and stimulants are associated with a reduction or even normalization of structural abnormalities in ADHD (Cortese & Castellanos, 2012). Recent studies also detected abnormalities in the frontostriatal areas, temporo-parietal lobes, basal ganglia, corpus callosum, cerebellum, amygdala, hippocampus, and thalamus (Cortese, 2012). Functional imaging has shown that ADHD is associated with deficits in the connectivity of frontostriatal, parietotemporal, and frontocerebellar networks (Konrad & Eickhoff, 2010).
In the imaging studies, various parameters of the dopaminergic system have been measured, including DAT (Spencer et al., 2005), dopamine D2 receptors (Lou et al., 2004), dopamine synthesis (Ernst et al., 1999), and extracellular dopamine levels (Ilgin, Senol, Gucuyener, Gokcora, & Sener, 2001). The most widely investigated parameter has been the DAT, especially variations in the density of presynaptic DATs. Studies have revealed either higher DAT availability (Cheon et al., 2003; Krause et al., 2000; Spencer et al., 2005), ranging from a 70% increase in the initial report (Dougherty et al., 1999) to a less pronounced increase most recently (Larisch et al., 2006; Silva et al., 2014), unaltered levels of DAT (van Dyck et al., 2002), or reduced in vivo DAT availability (Jucaite, Fernell, Halldin, Forssberg, & Farde, 2005; Volkow et al., 2007). The inconsistency in the results might be due to differences in imaging techniques, radiotracers and the methods used, differences in patients’ characteristics, including medication history, comorbid conditions, previous drug or cigarette exposure, ethnicity, gender and age, and differences in study group sizes. Currently, no unanimous conclusion had emerged from the various studies. Therefore, more extensive investigations are required to define the relationship between DAT density and the pathophysiology of ADHD.
In addition, ADHD has a strong genetic background and high familiarity, with an estimated heritability of about 80% (Faraone et al., 2005). The findings from electroencephalography (EEG) showed strong sibling similarity and parent–offspring correlations, molecular genetic, candidate gene, and genome-wide linkage/association studies have revealed that several different classes of genomic variants have been identified to be associated with ADHD risk (Loo et al., 2010). The decreased performance observed in some neuropsychological functions (Rommelse et al., 2008), and deficits in attentional control (Slaats-Willemse, Swaab-Barneveld, de Sonneville, & Buitelaar, 2007) and response inhibition (Slaats-Willemse, Swaab-Barneveld, de Sonneville, van der Meulen, & Buitelaar, 2003), in unaffected relatives of patients with ADHD have provided strong evidence to support the familial overlap between ADHD and executive dysfunction (Seidman, Biederman, Monuteaux, Weber, & Faraone, 2000). A recent study demonstrated executive dysfunctions in unaffected siblings of ADHD children in an ethnic Chinese population (Gau & Shang, 2010). It was also found that parents of ADHD children may suffer from impaired cool executive function (Nigg, Blaskey, Stawicki, & Sachek, 2004). The association between the dopaminergic system and executive function was confirmed by neuroimaging studies (Cropley, Fujita, Innis, & Nathan, 2006). The finger tapping test (FTT) is widely used to represent central dopaminergic function (Yang et al., 2003). In our previous study, we found that the FTT performance of healthy individuals with ADHD offspring was poorer than those without ADHD offspring (Hung et al., unpublished data). As families of individuals with ADHD are expected to share some of the genes involved in the disorder, we hypothesized that relatives of ADHD patients would present different DAT availabilities compared with healthy individuals. To the best of our knowledge, the role of DAT availability in healthy participants with a family history of ADHD has not yet been explored. We aimed to examine whether DAT availability differs between healthy individuals with and without ADHD offspring.
Method
Ethics
The institutional review board at the National Cheng Kung University Hospital approved the previous study proposal (BR-100-112). Before any procedure was performed, all participants signed an informed consent form.
Participants
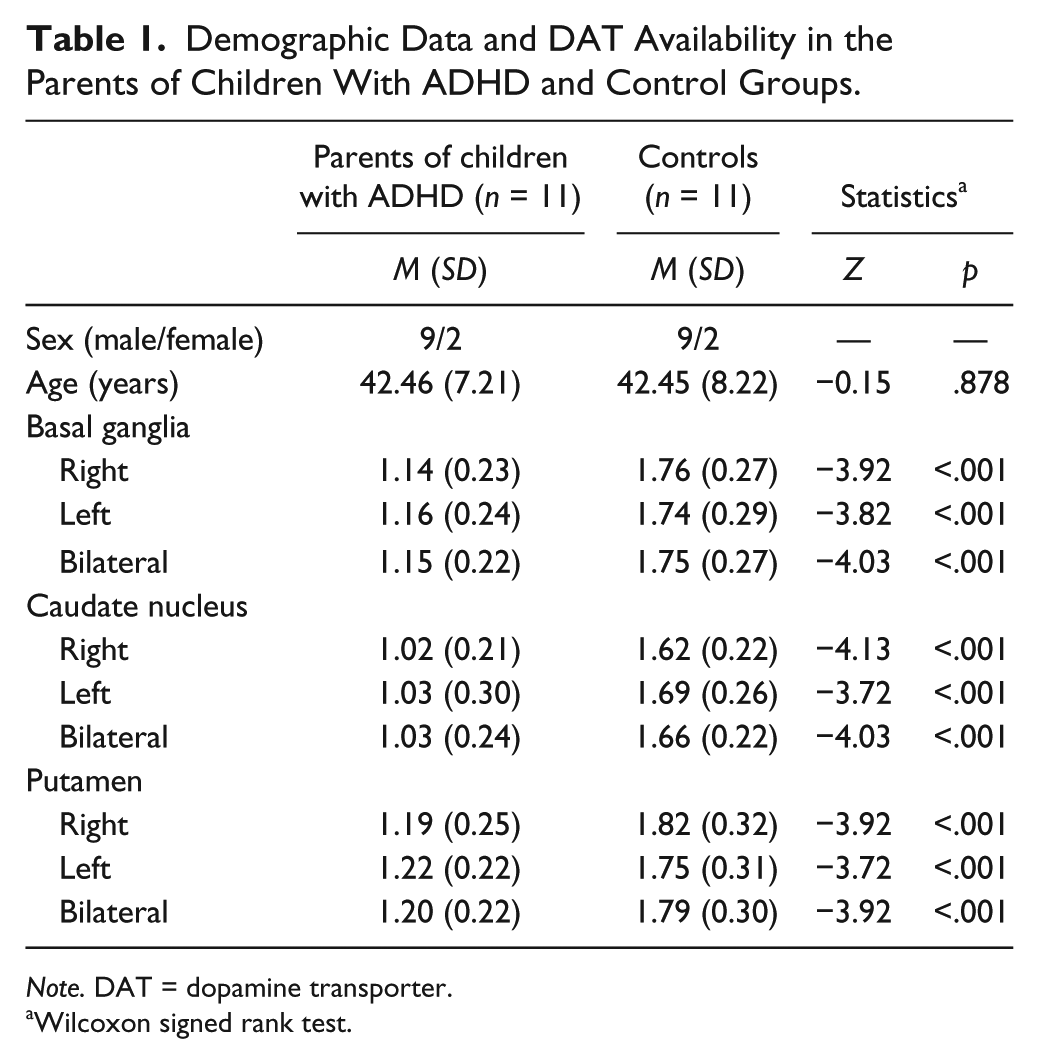
Two groups of participants were enrolled in the study. The first group comprised 11 healthy participants with ADHD offspring (n = 11). The second group comprised 11 healthy participants without ADHD offspring (n = 11). The two groups were matched for age and gender (Table 1). The participants underwent comprehensive medical and neurological examinations to ensure the absence of disease and were interviewed by senior psychiatrists who conducted the Mini International Neuropsychiatric Interview (Sheehan et al., 1998) to exclude individuals with other psychiatric disorders.
Demographic Data and DAT Availability in the Parents of Children With ADHD and Control Groups.
Note. DAT = dopamine transporter.
Wilcoxon signed rank test.
Inclusion criteria
Group 1 comprised healthy parents with offspring who fulfilled the Diagnostic and Statistical Manual of Mental Disorders (4th ed.; DSM-IV; American Psychiatric Association, 1994) criteria for children with ADHD. Group 2 comprised healthy parents without ADHD offspring. All participants were aged between 20 and 60 years.
Exclusion criteria
The exclusion criteria were as follows: (a) individuals with a history of physical systemic illness or other comorbid psychiatric or neurological illness; (b) individuals who were mentally retarded, pregnant, at risk of suicide, or experiencing unstable clinical psychiatric symptoms; (c) individuals with a history of alcohol or other substance abuse or dependence; (d) female participants of child-bearing age had to take an acceptable form of contraceptive throughout the duration of the study to be included. All female participants underwent an instant urine pregnancy test prior to starting the experiment.
Imaging: [99mTc] TRODAT-1 Single-Photon Emission Computed Tomography (SPECT) and MRI
The imaging procedure was identical to that used in our previous study (Chen et al., 2011). For brain imaging, each participant was intravenously administered 740 MBq (20 mCi) [99mTc] TRODAT-1 (a radio-labeled form of a tropane derivative for the selective labeling of DAT) in a quiet environment about 10 min after the insertion of an intravenous line. SPECT data were obtained using an energy window of 15% centered on 140 keV for [99mTc]. Imaging of [99mTc] TRODAT-1 was initiated approximately 240 min after injection, and SPECT images were acquired over a circular 360° rotation in 120 steps, 50 s per step, in a 128 × 128 × 16 matrix. The images were then reconstructed using Butterworth and Ramp filters (cutoff frequency = 0.3 Nyquist; power factor = 7) with attenuations according to Chang’s method, and the reconstructed transverse images were realigned parallel to the canthomeatal line. The slice thickness of each transverse image was 2.89 mm. In addition, all participants underwent MRI (Signa CV-I, 1.5 Tesla, GE Medical Systems, Milwaukee, Wisconsin, United States). Using the commercial software PMOD (PMOD Technologies, Zurich, Switzerland), each participant’s SPECT image was automatically co-registered with the corresponding T2-weighted MRI image and was then manually finely adjusted by an experienced nuclear medicine physician. The MRI image was used as a reference, so the slice thickness of the co-registered images was the thickness of the T2-weighted MRI images (3.3 mm). For co-registration, rigid transformations were defined by 6 parameters: the rotation angles and translation distances in the three spatial directions. The interpolation method was trilinear. On the co-registered images, the two contiguous transverse slices that contained the most intense striatal radioactivity were further examined to ascertain whether the SPECT and MRI images were co-registered accurately, and whether the striatum was best seen on the two slices of the MRI images. If that was not the case, further adjustment of co-registration was performed manually until a satisfactory outcome was achieved. Regions of interest (ROIs) in the striatum (basal ganglia, caudate nucleus, and putamen) and occipital cortex were then drawn on the two contiguous MRI transverse slices, and these ROIs were projected onto the co-registered SPECT images. The ratio of the radioactivity [(St-Oc)/Oc ratio] was then derived by dividing the difference between the average activity in the striatum (St) and the average activity in the occipital cortex (Oc) by the average activity in the occipital cortex (Oc; Hwang, Yao, Wey, & Ting, 2004).
Statistical Analysis
As the sample size was small, the Mann–Whitney U test was used to examine the differences in age and DAT availability between the groups. Supplemental ANCOVA and independent t tests were used to test the robustness of the findings before and after controlling for the effect of age. The level of significance was set at p < .05 (two-tailed). All analyses were performed using SPSS software (Version 17.0, SPSS Inc., Chicago, Illinois, United States).
Results
No significant differences were found in terms of age and sex between groups, as shown in Table 1. DAT availability was significantly lower in the basal ganglia (p < .001), caudate nucleus (p < .001), and putamen (p < .001) of healthy participants with ADHD offspring compared with controls (Table 1). The results of the independent t test and ANCOVA also showed that DAT availability was lower in healthy participants with ADHD offspring compared with controls.
Discussion
The results of the present study showed that healthy parents with ADHD offspring had lower DAT availability in the bilateral basal ganglia, caudate nucleus, and putamen than healthy controls without ADHD offspring.
Previous studies have demonstrated disruption in DAT levels in participants with ADHD, although the findings are inconsistent. The most recent meta-analysis included nine studies addressing striatal DAT density in ADHD, suggesting that striatal DAT density was 14% higher on average in the ADHD group than in the healthy comparison group. However, there were methodological limitations of the study, including small sample size, limited statistical power for meta-regression analysis, artificial comparison across several radiotracers, and the causality of the regression findings could not be tested in cross-sectional designs (Fusar-Poli, Rubia, Rossi, Sartori, & Balottin, 2012). There was another recent study reported by Silva et al., revealing that 11 adolescents with ADHD had higher striatal DAT compared with 19 healthy controls (Silva et al., 2014). Among the above studies related to DAT alterations in ADHD, the first report was 70% increases in DAT in participants with ADHD (six participants with ADHD compared with norms; Dougherty et al., 1999). The study by van Dyck et.al (2002) found unchanged striatum DAT binding in nine adults with ADHD, and that by Jucaite et al. (2005) found lower DAT availability in midbrain but not in striatum. However, previous exposure to stimulants or other drugs and smoking status contribute to the contradictory study results. Otherwise, age and gender may play a role on the divergent results. Cheon et al. (2003) investigated nine children and found an increased striatal DAT binding, and Jucaite et al. (2005) compared 12 adolescent ADHD participants with 10 young adults. Silva et al. (2014) investigated only male adolescents, and the findings may not be generalized to females or other age groups. Due to difficulty in recruitment of participants with ADHD without any neurological, significant clinical disease or comorbid psychiatric disorder, most studies reported small sample size as limitation, while Volkow et al. (2009) proposed relatively large sample size (lower DAT availability in 53 ADHD participants compared with 44 healthy controls).
In fact, the role of ADHD subtype and severity of symptoms of the DAT availability remained unclear now. The lifelong plastic change of brain and dopaminergic system, the ability to adapt, and the intrinsic compensation all contribute to the heterogeneity of ADHD. Thus, the developmental, neurophysiological, and neuropathophysiological phenomena may also be responsible for the obvious differences detected (Ludolph et al., 2008).
To our knowledge, this is the first DAT availability study focusing on parents with ADHD offspring to investigate the pathophysiology and heritability of ADHD. Previous DAT imaging results were mainly based on ADHD participants. Therefore, comparing the similarity between previous results and this result could be not easy. However, our results of lower DAT availability in healthy adults with ADHD offspring revealed that ADHD could be a heritable condition via altered DAT levels. Furthermore, the participants in our study were healthy adults who had never received psychostimulant treatment; therefore, a secondary brain response to psychostimulants could be excluded, and also less possibility of intrinsic compensation to dopaminergic system. Currently, it is unclear whether low DAT availability is the primary presentation of the ADHD genotype or a secondary effect due to down-regulation under intrinsic DAT dysfunction or loss of dopaminergic nerve cells. Our results could provide evidence that lower DAT availability might be the preliminary presentation in the dopaminergic system.
In these results, the affected regions included bilateral basal ganglia, caudate nucleus, and putamen, which are related to the mesolimbic, mesocortical, and nigrostriatal tracts of the dopaminergic system. The dopaminergic system has been reported to be positively related to a wide spectrum of cognitive functions (Hsieh et al., 2010). This result is in line with previous functional MRI findings that parents of ADHD children also suffer from impaired cool executive function (Nigg et al., 2004), as they showed altered frontostriatal connectivity during a cool cognition task (i.e., go/no go task; Casey et al., 2007).
Our result is consistent with those of Volkow et al. (2009) and Hesse et al. who also reported decreased DAT availability in ADHD patients. Hesse et al. reported lower DAT availability in striatum, head of the caudate, and the putamen, while Volkow reported lower DAT availability in left ventral striatum (including accumbens and caudate), left midbrain, and left hypothalamus in ADHD patients but not in putamen (Volkow et al., 2009). Our result demonstrated lower DAT availability in healthy parents with ADHD offspring over bilateral basal ganglion, caudate, and putamen without left-to-right asymmetry. The reason for the discrepancy in affected regions might be the different imaging techniques, radiotracers, methods, small sample size, and characteristics of the sample in our study. The definite reason remained unclear and needs further investigation.
There are several limitations to the present study. First, our sample size was small. Second, both hyperactive pattern and inattention are associated with altered striatal dopamine concentrations and DAT levels (Volkow et al., 2007). However, due to the small sample size in the current study, it was hard to analyze the subtype specific role of the DAT in the striatum. Besides, we did not measure the ADHD subtype nor rating the hyperactivity/impulsivity/inattention scores of ADHD offspring. Further study may consider correlating the results with the subtype of the ADHD offspring.
Footnotes
Acknowledgements
The authors also wish to thank Professor Yuan-Hwa Chou from Taipei Veterans General Hospital, Ms. Tsai Hua Chang, Mr. Chien Ting Lin, and all of the research participants.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the National Science Council of Taiwan (NSC 100-2314-B-006-041-MY3, NSC 101-2314-B-006-065, and NSC 102-2420-H-006-007-MY2), the Ministry of Science and Technology, Republic of China (MOST 104-2314-B-006-053-MY2), and National Cheng Kung University Hospital (NCKUH-10104019 and NCKUH-10405033). This research also received funding (D102-35001 and D103-35A09) from the Headquarters of University Advancement at the National Cheng Kung University, which is sponsored by the Ministry of Education, Taiwan, Republic of China.