
Abstract
Psychology has been a relatively disunified discipline since its inception, partly owing to the historical context in which it arose as a science. The interdisciplinary field of developmental science can potentially overcome this fragmentation. Specifically, the Developmental Systems (DS) approach recognizes that individuals, together with their contexts, constitute self-organizing, dynamic, complex systems best characterized by the processes that transpire when system components work together to build psychological attributes. This approach holds that these attributes emerge during development in ways that depend on spatial and temporal (i.e., historical) relations among system components. DNA works in collaboration with other factors in its local environment, so genetic determinism can be rejected; psychological traits cannot be predicted from genetic information alone. Research on the epigenetic control of gene expression has bolstered this insight, revealing how experiences influence genetic activity and, ultimately, behavior. Behavioral epigenetics likely bears on all psychological phenomena, putting it in a position to help unify the behavioral sciences. The DS perspective encourages skepticism of constructs that assign static traits to individuals and of tests that rely on genetic information alone to predict developmental outcomes. Because all characteristics develop—and because individuals are cohesive, highly integrated systems—psychology’s unification is an attainable goal that will be advanced by adopting this perspective.
Keywords
Introduction
On August 9, 2025, in Denver, Colorado, I had the honor of delivering the “Arthur W. Staats Lecture for Unifying Psychology” at the annual convention of the American Psychological Association. This paper is a formal presentation of the ideas conveyed in that talk. My contention is that a developmental perspective will be vital for theorists hoping to build a unified psychology in the 21st century. However, before making that argument, it will be useful to consider why psychology currently remains disunified.
A Brief History of Psychology’s Fragmented Nature
When psychology first emerged as a distinct science, it inherited ideas that were part of the zeitgeist at the time. One such idea was that a person’s characteristics can be independently influenced by two different kinds of factors, namely the experiences we have as we develop and the evolutionary history of our species. For example, it was in 1874 that Charles Darwin’s half-cousin, Francis Galton, gave his book “English Men of Science” the subtitle “Their Nature and Nurture,” thereby introducing the centuries-old Nature-vs-Nurture dichotomy into scientific discourse (Galton, 1874; Moore, 2002; Plomin, 1994). A mere 5 years later, Wilhelm Wundt established the very first experimental psychology research laboratory, formally establishing psychology as a science rather than a branch of philosophy (Boring, 1950). And by the 1890s, Sigmund Freud and William James were both addressing psychological questions in ways that proved foundational for our field (Breuer & Freud, 1893; Freud, 1894a, 1894b; James, 1890). Thus, it is perhaps unsurprising that Galton’s nature/nurture dichotomy became an organizing conceptual framework in psychology from the inception of the field. Soon, psychologists were thinking of behavioral characteristics as being caused primarily by “Nature” or “Nurture.”
Toward the end of the 19th century, Galton coined the word “eugenics” to refer to “the science of improving stock.” As he saw it, the goal of this science should be to increase the number of people “hereditarily endowed with noble qualities” (Galton, 1883, p. 17). In the wake of Galton’s work, the start of the 20th century found eugenic ideas taking hold in scientific circles (Kevles, 1995; Moore, 2013c); these ideas contributed to the belief that some people are, by their very nature, possessed of distinctive behavioral tendencies, such as being inclined toward violence (See Figure 1) or particular cognitive dispositions (such as “genius” (Galton, 1869) or “feeble-mindedness” (Goddard, 1912)). As is well-known, the idea that some people could be considered inherently “mentally defective” (e.g., Laird, 1935, p. 1607) or belonging to an “inferior race” (e.g., Viteles, 1928, p. 175) became central to Nazi ideology in the early 1930s, ultimately contributing to that regime’s industrialized murder of millions of innocent people (Kevles, 1995; Lerner, 1992; Müller-Hill, 1998; Proctor, 1988). Original photographs and composites used around 1885 by Francis Galton to display features he considered “common among men convicted of crimes of violence.”
By the mid-20th century, the pendulum had swung to the opposite extreme, at least in part as a reaction to what had transpired in Nazi Germany. Rather than attributing people’s psychological characteristics to “Nature,” academic psychologists became almost exclusively concerned with how experiences—“Nurture”—lead people to develop their behavioral characteristics. One root of this branch of psychology can be traced to John B. Watson’s book Behaviorism, which contained the following famous quotation: Give me a dozen healthy infants, well-formed, and my own specified world to bring them up in and I'll guarantee to take any one at random and train him to become any type of specialist I might select—doctor, lawyer, artist, merchant-chief, and yes, even beggar-man and thief, regardless of his talents, penchants, tendencies, abilities, vocations, and race of his ancestors (Watson, 1924, p. 82).
Clearly, Watson was confident that Nurture could always overcome any Nature-based predispositions. Likewise, well into the second half of the 20th century, B.F. Skinner was arguing strongly that man is “indeed controlled by his environment” (Skinner, 1973, p. 210).
Even as behaviorists rejected the philosophical nativism of an earlier generation of psychologists, the idea that some of our psychological characteristics are genetically or evolutionarily predetermined continued to motivate work in some branches of our field. For example, by 1960, Galton’s ideas about using twins to study genetic and environmental contributions to human traits had led to the emergence of behavior genetics, a distinct branch of psychology devoted to elucidating how genes influence behavioral and psychological differences between individuals (Loehlin, 2009). Then, in 1973, the first-ever Nobel Prize awarded for the study of behavior was given to Tinbergen, von Frisch, and Lorenz, recognizing their work on “instinctive” behaviors that were thought to be free of the influence of “Nurture.” These researchers established the subdiscipline of behavioral science known as ethology, which examines the behavior of non-human animals in natural settings. Shortly thereafter, Wilson (1975) published Sociobiology and Dawkins (1976) published The selfish gene, together heralding the soon-to-emerge subdiscipline now known as evolutionary psychology. Dawkins’ commitment to the primacy of “Nature” over “Nurture” was apparent in his well-known claim that people are “robot vehicles blindly programmed to preserve the selfish molecules known as genes” (p. vii). Contemporary scientists continue to work within the frameworks of behavior genetics, ethology, and evolutionary psychology (e.g., Costello & Thomas, 2024; Plomin & von Stumm, 2018).
These new subdisciplines emerged in the 20th century specifically to study behaviors thought to be inherited through biological mechanisms. At the same time, traditional subdisciplines of psychology were also analyzing their subjects using a “Nature-or-Nurture” lens. For example, following work by McCrae and Costa in the 1980s and 1990s, psychologists converged on the five-factor model of personality, a model that remains today the dominant framework in the subdiscipline of personality psychology (Bainbridge et al., 2022). In 2000, McCrae and colleagues wrote that “both broad personality factors and the specific traits that define them are best understood not as characteristic adaptations, but rather as endogenous [i.e., “biologically based”] basic tendencies” (p. 174). Unconcerned about putting too fine a point on their contention, McCrae and colleagues (2000) chose to bluntly title their paper “Nature over nurture.”
Lest the preceding paragraphs make it seem like Psychology now consists only of subdisciplines that study putatively “unlearned,” “evolved,” “instinctive,” “biologically based,” or “genetically determined” behaviors, it should be noted that other branches of psychology have remained committed to the idea that experiences are the only important contributors to human behaviors. Proponents of the “Cognitive Revolution” that had swept most American university psychology departments by 1970 claimed that their work had “replaced” a “failed” behaviorism (Mandler, 2002, pp. 339−340), but behaviorist principles continue to animate the work of a very large number of psychologists. As of May 20, 2026, the Behavior Analyst Certification Board (BACB) recorded that more than 342,000 professionals held BACB certifications (Behavior Analyst Certification Board, 2025). Similarly, psychologists adopting the theoretical perspectives associated with subdisciplines like educational psychology and school psychology tend to concentrate exclusively on the effects of people’s experiences with their families, communities, schools, and workplaces (Burns et al., 2015).
Given this history, it should not be surprising that psychology is a splintered discipline, one in need of unification. There are, of course, several ways in which branches of this field diverge. On one hand, a subdiscipline like behavioral neuroscience approaches psychological phenomena from a relatively molecular standpoint, focusing on factors like neurotransmitters, cells, and neural circuits, whereas a subdiscipline like social psychology approaches these phenomena from a relatively molar standpoint, focusing on factors like political contexts, cultures, and group dynamics. On another hand, psychological subdisciplines concerned with affect focus on phenomena like emotions and feelings whereas subdisciplines concerned with cognition focus on phenomena like thoughts and memories. Unification of the field across all of these divisions will be a major challenge. However, a theoretical tool already exists that can facilitate unification, at least across subdisciplines committed to explanations that rely on “Nature” or “Nurture” alone: the developmental systems perspective (see Michel, 2013, for a concordant point of view).
The Emergence of Developmental Science
Even as psychologists took sides in “Nature-vs-Nurture” debates in the mid-20th century, there were already theorists arguing against the dichotomous view that grew out of Galton’s 19th century speculations. Among them were the comparative psychologists Daniel Lehrman (1953) and T. C. Schneirla (1957), the ethologist Frank Beach (1955), the experimental psychologist Zing-Yang Kuo (1967), the evolutionary biologist Richard Lewontin (1974), and the psychobiologist Gilbert Gottlieb (1970). What these scientists shared was an awareness of the importance of development; later writers also emphasized the importance of the spatial and temporal relations that give rise to our traits (Ford & Lerner, 1992; Lerner, 2015; Overton, 2015). By the 1990s, these theorists’ ideas had led to the advent of a new interdisciplinary endeavor called “developmental science.” Emerging at the interface of developmental psychology, cognitive science, philosophy of science, and evolutionary, developmental, and molecular biology, a unifying theme in the new field of developmental science was that behavioral scientists should study the processes that give rise to psychological and behavioral characteristics (i.e., behavioral “phenotypes” or traits). In addition to effectively undermining the Nature/Nurture dichotomy, the developmental approach highlights organisms’ potential for the kinds of change that enable improved adaptation. Appreciating the value of this new discipline, the U.S. National Science Foundation released its first “Developmental and Learning Sciences” program solicitation in 2006. Of course, there are still psychologists who vigorously defend nativist positions (e.g., Hamlin, 2013, 2023; Spelke, 2022; Wynn, 2008); these researchers appear not to be interested in the developmental processes that give rise to the psychological characteristics they study. Regardless of any value of their work, these theorists ought not be considered developmental scientists per se.
A focus on developmental processes differs markedly from the focus that characterizes most of psychology’s subdisciplines. Many researchers continue to write as if inherited DNA sequences influence characteristics like intelligence (e.g., Plomin & von Stumm, 2018), personality (e.g., McCrae et al., 2000), or psychopathology (e.g., Harden, 2026; Sullivan et al., 2003) in ways that manifest as “genetic predispositions” (see Furrer & Turkheimer, 2026, for a critical evaluation of this idea). Accordingly, these characteristics are conceptualized as predictable features of individuals’ psychologies. In contrast, while developmental scientists acknowledge that genetic factors influence all behaviors and psychological states, they reject the idea that any characteristics are predetermined in ways that permit accurate predictions on the basis of genetic factors alone. Instead, they understand that all psychological phenotypes develop when biological and non-biological factors act in concert to build those phenotypes. Therefore, phenotypic outcomes cannot be specified prior to the unfolding of development itself (Lickliter & Moore, 2023), a process that continues across the lifespan (Mousley et al., 2025).
These insights carry with them at least one notable advantage and at least two important implications. A significant advantage of the developmental perspective is the way it draws researchers’ attention to biopsychosocial mechanisms that afford experimental manipulation and the possibility of discovering interventions that can be deployed to individuals’ benefit. Further, one implication of this perspective is that psychological phenotypes in general can be recognized as inherently dynamic; living organisms are continuously adapting to their contexts. Finally, because all psychological phenotypes develop, the developmental perspective has the potential to unify the study of behavioral/psychological phenomena, at least to some extent.
The Developmental Systems Perspective
At the heart of much developmental science is the systems orientation that is a distinctive feature of Dynamic Systems Theory. Dynamic Systems Theory emerged from progress in mathematics and physics in the 1980s and helped describe how some natural physical phenomena—such as stars, tornadoes, snowflakes, and ecosystems—are able to self-organize (Nicolis & Prigogine, 1989; Stewart, 1989). These systems were observed to develop over time into complex, structured entities even in the absence of any pre-existing “design” specifying those entities. That is, organized entities with characteristic traits can emerge spontaneously from events that involve the components of these dynamic systems, even when none of those components hold “instructions” for creating the entities (Moore, 2002).
By the start of the 21st century, some scientists were looking at biological and behavioral phenomena in this way, too (Kelso, 2000, p. 25). For example, the Nobel Prize-winning biologist Gerald Edelman wrote that “The brain is … a self-organizing system” (1992, p. 25). Similarly, Thelen and Smith (1994) argued persuasively that motor development in infants unfolds in an orderly fashion without the need for the kinds of biologically determined “motor programs” that earlier theories required. Their perspective, which was shared by theorists such as Ford and Lerner (1992) and Johnston and Lickliter (2009), came to be known as the Developmental Systems (DS) perspective (Moore, 2016). The DS perspective was dramatically different than traditional approaches that had imagined “instructions” for development being somehow encoded within the genome, instructions able to produce supposedly “innate” characteristics such as locomotor patterns, sexual orientations, Universal Grammar, and instincts. These older approaches had typically viewed behaviors as originating within individuals.
In contrast, proponents of the DS perspective conceptualize an organism and its environment as a single complex system. In this way, behaviors are understood not to originate within an individual, but rather to emerge from processes that come about when individuals interact with their environments. 1 Thelen and Smith’s (1994) position built on the work of Russian neurophysiologist Nikolai Bernstein (e.g., Bernstein, 1967), who discerned that when a person moves, their central nervous system is being presented with a continuously changing situation. We are perhaps most aware of this when we walk on uneven, unpredictable terrain. As a result of this state of affairs, human functioning is necessarily dynamic; we need to be able to adapt in a continuous manner to ever-changing contexts. Altering any one of the many components that contribute to a movement pattern—components that include joints, tendons, various types of muscle fibers, proprioceptive and vestibular input, the nervous system’s integration of visual information and motor feedback, features of the environment, the individual’s motivations, the presence of competitors or observers, etc.—will affect the pattern as a whole. And to compensate for altering any of these components, behaving organisms always adjust the functioning of other components involved in the movement.
From this perspective, movement patterns are processes that involve assembling many system components into an available state within particular contexts, and always in real-time. The importance of environmental features as a component of the dynamic system is easy to see when we consider human locomotion in various contexts: our gaits are quite different on concrete, grass, ice, sand, steep mountain trails, in a “bouncy house,” or on the moon. This is why the context of a behavior must always be considered a component of the system being studied, and why systems theorists reject that idea that behaviors arise strictly from within individuals. Instead, behaviors are recognized as emergent phenomena that reflect organisms-behaving-within-environments. Paraphrasing Bernstein in English, our actions are never repeated (as they would be if they were the product of a biologically determined “motor program”); instead, they are always newly constructed in each real-world context.
Focusing on bodily movement naturally draws attention to how neural and mental activity is embodied—integrally linked to the physical body’s interactions with its environment. However, for over 3 decades, cognitive scientists (Varela et al., 1991) philosophers (Hutto & Myin, 2017, 2018; Lakoff & Johnson, 1999) and psychologists (Marshall, 2014; Witherington & Heying, 2013) have been taking seriously the idea that psychological characteristics are similarly embodied, emerging from individual organisms’ interactions with the world. Fittingly, theorists such as Overton (2006), Witherington (Witherington et al., 2018; Witherington & Heying, 2013), and van Geert (van Geert & Steenbeek, 2005) have argued cogently that the systems perspective should be generalized beyond motor development and applied to cognition, affect, desires, beliefs and every other psychological feature. This perspective has the potential to help us understand the development of all psychological/behavioral phenotypes in the same, unified way. Once we accept that context is no more relevant for motor activity than it is for cognition, affect, and other psychological phenomena, it becomes easier to see development in general as a process that is never prescribed or inevitable. Although there are still contemporary psychologists who have not yet adopted this perspective (e.g., Spelke, 2022; see Moore & Lewkowicz, 2024), many behavioral scientists now recognize development to be a flexible, context specific, and adaptable phenomenon.
Importantly, “context” here refers both to an individual’s current environment and to the “developmental environment” of the behavior. This latter environment is a temporal context that is historical in nature, and as such, it is characterized by what preceded the behavior in question at multiple timescales. For example, behaviors are typically influenced by events on a millisecond (or shorter) timescale (e.g., what was the chemical state of the person’s brain just before the behavior?). Likewise, behaviors can be influenced by events on longer timescales, such as minutes (e.g., was the person given stressful news 20 min ago?), hours or days (e.g., did the person see a film the night before that has changed how they are interpreting their situation?), or even years or decades (e.g., what sorts of childhood experiences led the person to expect certain kinds of behaviors to yield desirable outcomes?). When we behave, we do so both in a current environment as well as in an historical, developmental context.
An important consequence of the DS approach is that it does away with efforts to identify the simple cause of any behavioral effect, because all behavior is understood to be dependent on the individual’s concurrent environmental context, their developmental history, and biological factors such as brain structure and chemistry, genetic and epigenetic factors, hormonal states, and the histories of all these factors. Thus, our behavioral characteristics—just like our anatomical and physiological characteristics—are influenced by an enormous number of biological and non-biological factors, all of which co-act to yield our psychological traits (Lerner, 2015; Overton, 2015; Witherington & Lickliter, 2016). And these phenotypes all emerge as a consequence of developmental processes. Unfortunately, although it is possible to study these amazingly complex systems, it is undeniably difficult to do so (after making this same point, Lickliter and Herrington (2024) offer examples of successful uses of the DS framework). Nevertheless, by adopting the DS perspective, we can dispense with single-cause, static, deterministic explanations for our psychological phenotypes; such explanations are, in the end, indefensible. In this way, we can bring the various phenomena of interest to psychologists under the same theoretical umbrella.
An early example of a developmental systems theory was Bronfenbrenner’s Ecological Systems Theory, established in the 1970s (Bronfenbrenner, 1977, 1979). However, it was not until 1986 that Bronfenbrenner formally introduced an element of developmental time into his theory, a key aspect of all later DS theories (Bronfenbrenner, 1986). And although he eventually incorporated a time dimension into his theory, Bronfenbrenner was primarily concerned with identifying the various kinds of “external environments” (Bronfenbrenner, 1986, p. 723) that influence individual development, sometimes via their effects on families. In contrast, Gottlieb’s research on the development of prenatal behavior had led him, by 1976, to explicitly acknowledge the importance of time and timing in developmental events (Gottlieb, 1976; see also Turkewitz & Devenny, 1993). In addition, rather than attending almost exclusively to external environments, Gottlieb was also interested in the components of developmental systems that are internal to either pregnant women or to the embryos/fetuses developing inside them. Following Gottlieb, I will consider the importance of both internal and external components of developing systems in later sections of this paper, consistent with the tenets of the interdisciplinary field of developmental psychobiology (Michel, 2013; Michel & Moore, 1995). For now, it is worth noting that Gottlieb was also interested in evolutionary processes (Gottlieb, 1992). Although a broad consideration of the role of evolution in the development of psychological phenotypes is beyond the scope of this review, there can be no doubt that natural selection’s effects on populations have influenced the developmental processes observable in individuals today (Witherington et al., 2023; see also Lala, 2025). Hence, a quick comment about evolutionary explanations for human characteristics is warranted.
A Brief Note on the Role of Evolution in the Development of Psychological Phenotypes
A particular type of Evolutionary Psychology—one associated with theorists such as Cosmides and Tooby (1992), Pinker (2002), and Buss (2015)—has been touted as a “revolutionary” approach that can serve as “a cogent meta-theory for [all of] psychological science” (Buss, 2020, pp. 316−317). However, this narrowly defined Evolutionary Psychology has been criticized by numerous theorists for several reasons (e.g., Barrett et al., 2014; Buller, 2005; Lickliter & Honeycutt, 2013; Moore, 2026; Narvaez et al., 2022). In the context of the current paper, I will restrict my criticism of so-called Narrow Evolutionary Psychology (Scher & Rauscher, 2003) to just one point.
There is a sense in which all behaviors reflect evolutionary processes, of course; behaviors depend on myriad lower-level biological factors including muscle fibers, neurons, neurotransmitters, organized neural circuits, etc., all of which have long evolutionary histories (Moore & Lewkowicz, 2024). However, there are many behaviors that do not demand evolutionary explanations at the behavioral level of analysis (Moore, 2026). For example, spending an afternoon reading a paranormal mystery novel does not require an evolutionary explanation, because this behavior is unlikely to be adaptive or maladaptive—it is unlikely to bear on survival or reproduction. Furthermore, even if a behavior does affect survival or reproduction, it will not necessarily require an evolutionary explanation. Individuals who send text messages containing sexual content are more likely to engage in sexual activity (Hicks et al., 2021; Rice et al., 2012); nonetheless, although sexual activity increases the likelihood of reproduction, sexting per se cannot be attributed to evolution, because the technology required for text messaging was not available to our ancestors. As Hauptman and colleagues (2024) have noted, many of our behavioral capacities have not evolved via natural selection.
In contrast to evolutionary explanations, developmental explanations are required to explain all phenotypes, since anatomical, physiological, and psychological phenotypes do not exist in undeveloped, fertilized eggs. All phenotypes—behavioral characteristics included—must develop (Moore & Lickliter, 2023). Moreover, developmental explanations can be sufficient explanations for behavioral characteristics that do not merit evolutionary explanations. So, whereas developmental explanations for a behavior are always necessary and are sometimes sufficient, evolutionary explanations are never sufficient and are only sometimes necessary to make sense of the behavior (Moore, 2013d; Moore & Lewkowicz, 2024).
Components of Developmental Systems & The Failure of Genetic Determinism
As Bronfenbrenner (1977) observed, development is influenced by several layers of environmental factors, including those surrounding a child in the microsystem (e.g., family, neighbors, peers), exosystem (e.g., government and educational systems, media), and macrosystem (e.g., beliefs, values, norms, and laws of the culture in which development is taking place). Likewise, as Gottlieb (1991) observed, development is also influenced by several layers of biological factors, including organs, tissues, hormones, cells, organelles, and nucleic acids such as DNA and RNA. To represent how these many environmental and biological factors mutually influence one another bidirectionally across multiple levels of analysis, Gottlieb drafted a simplified diagram showing just four system components interacting with one another across developmental time (see Figure 2). This sketch is a visual representation of the previously stated assertion, namely that behaviors, thoughts, and feelings should be recognized as emerging in development from the concerted actions of a multitude of both biological and non-biological factors at numerous levels of analysis. These levels include our cultural contexts, our current social situations, our immediate physical environments, our nervous systems, our internal organs, the compositions of our cells, our DNA, and other factors operating at these different levels. An adaptation of Gottlieb’s diagram capturing the essence of the developmental systems perspective. The figure shows just four (of many possible) levels of analysis; components at these levels co-act and influence one another bidirectionally.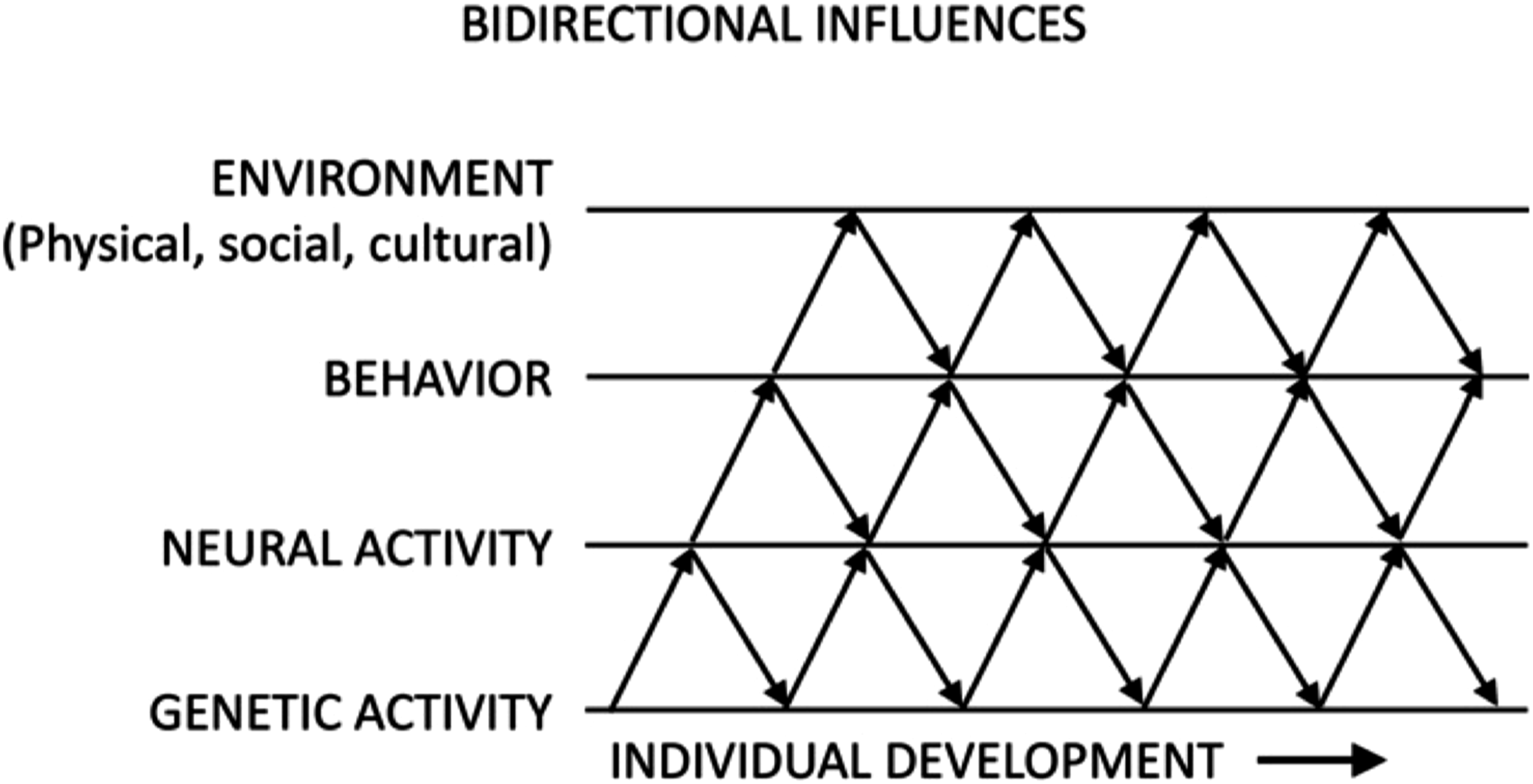
The DS perspective holds not only that these multifarious system components all contribute to developmental outcomes, but also that DNA and the other factors share a fundamental causal parity (Griffiths & Tabery, 2013; Oyama, 1985), meaning that DNA cannot be considered any more important in phenotype determination than can any other factor. Actually, one of the consequential conclusions arising from the DS perspective is that our genes cannot be understood to single-handedly cause any of our phenotypes, whether they be anatomical, physiological, or behavioral (Griffiths & Stotz, 2006; Kampourakis, 2021; Lewontin, 2000; Moore, 2002; Moore et al., 2025; Noble & Noble, 2023). In fact, the failure of genetic determinism has been widely acknowledged by biologists (Barresi & Gilbert, 2023; Gilbert & Epel, 2015; Keller, 2010; Lewontin, 2000).
It can be difficult to accept this conclusion because recent advances in molecular biology, genetics, and genomics have generated a different impression. Indeed, media and textbook portrayals regularly suggest that there are certain genes that cause certain traits (Carver et al., 2017; Gericke et al., 2014). Nonetheless, this is a misunderstanding. Confusion about genetic causation of traits can arise for several reasons (Moore, 2008), but I will consider just one of these reasons here, namely the way in which normal experimental controls contribute to the misperception that genetic factors can determine phenotypic outcomes.
Conducting experiments properly involves controlling variables in ways that allow for valid causal inferences. For example, a researcher might do a so-called genetic “knockout” study in which all of the individuals in a control group of test animals share the same genome—they are clones of one another—and another group (the experimental group) shares that same genome with one exception: it lacks a single gene that is still present in the control group’s genome. This sort of genomic editing can be accomplished using molecular biology techniques such as CRISPR-Cas9. Genetic knockout studies can give the impression that any novel outcome ultimately observed in a phenotype must have been caused by the gene that was experimentally manipulated. Even though genetic knockout studies are interesting and potentially valuable, there are two important things to keep in mind when interpreting them.
First, if animals with a specific gene develop a normal phenotype and animals without that gene develop an abnormal phenotype, it can seem as if the gene in question is single-handedly responsible for the normal phenotype. However, this is not correct. A simple analogy can be helpful here. If we drain all of the gasoline from a gasoline-powered automobile, the car will not move. But this does not mean that gasoline is single-handedly responsible for a car’s normal functioning! Instead, gasoline-powered cars move only when they have normally functioning wheels, motor oil, spark plugs, and a variety of other components, too. Just as gasoline-draining shows that gasoline is necessary—but not sufficient!—for normal functioning of the car, the results of a genetic knockout study can suggest that a gene might be necessary for normal development of a phenotype. Still, that does not mean that the gene single-handedly causes that phenotype. Despite this logic, if the absence of (or a mutation in) a particular gene is associated with a particular disease, it seems natural for us to jump to the incorrect conclusion that we have identified “the gene for” that disease, as if the genetic abnormality causes the disease without any contributions from other system components (Kampourakis, 2017; Moore, 2002).
Second, although a particular gene might be shown in a knockout study to play a role in the development of a normal phenotype, it is surprisingly easy to misinterpret that gene’s role. This is perhaps best understood via an analogy that Rose and colleagues proposed in 1990. Although these theorists offered their analogy in the context of a discussion about psychosurgery—which entails “knocking out” a particular area in a brain—their logic applies just as well to genetic knockout studies: “If you remove a transistor from a radio and the result is that the radio thereafter emits nothing but howls, you are not entitled to assume that the function of the transistor that was removed was that of a howl suppressor. Rather, what we see in the radio with the transistor removed is the workings of the rest of the system in the absence of the transistor. But the most likely effect of removing a transistor or disconnecting parts of the brain is indeed some sort of howl” (Rose et al., 1990, p. 191).
Therefore, even a seemingly straightforward genetic knockout study cannot prove that a particular gene causes a particular phenotype, even if it might seem like it can. Discovering that the presence of (or a mutation in) a gene makes a difference in a phenotypic outcome is not the same thing as understanding what normally brings that phenotype into being. The fact is, a gene that has a bold effect on a phenotype should not be considered the cause of that phenotype.
Psychologists may have not yet uniformly accepted this fact, but many geneticists have understood it from the earliest days of their discipline. For example, in 1915, the pioneering geneticist Alfred Sturtevant wrote: “Although there is little that we can say as to the nature of Mendelian genes, we do know that they are not ‘determinants’ … The difference between normal red eyes and colorless (white) ones in Drosophila [fruit flies] is due to a difference in a single gene. Yet red is a very complex color, requiring the interaction of at least five (and probably of very many more) different genes for its production … We can then, in no sense identify a given gene with the red color of the eye even though there is a single gene differentiating it from the colorless eye … All that we mean when we speak of a gene for pink eyes is, a gene which differentiates a pink eyed fly from a normal one—not a gene which produces pink eyes per se …” (Sturtevant, 1915, cited in Carlson, 1966, p. 69, emphasis added).
This distinction between a difference-making factor and a causal factor is one reason biologists know that there is no such thing as a gene “for” a trait, in the sense that the gene causes the trait (Kampourakis, 2017, 2021; Moore et al., 2025; Tabery, 2014).
Although our schools and media often make it seem like some phenotypes—eye colors, for example (Moore, 2013b)—are single-handedly caused by specific DNA sequences (Gericke et al., 2014), contemporary molecular biologists know that genes do not build phenotypes on their own. Genes cannot be thought of as agentic controllers of events that take place in bodies, because DNA is an inert molecule; a bowl of naked DNA on a table will do nothing at all (Noble, 2015). Instead, DNA only does what it does in collaboration with a variety of factors in its local environment, working with these other factors to build other biological molecules such as cell membrane receptors, antibodies, neuromodulators, chaperone proteins, etc. Importantly, some of the factors in DNA’s local environment reflect the broader environment in which an individual is developing—for example, the individual’s diet or the presence of hostile competitors. And it is because phenotypes emerge from developmental processes involving co-acting genetic and non-genetic factors that an individual’s phenotypes cannot be accurately predicted from information about the individual’s genome alone (Gottlieb, 1991, 1992, 1998, 2007; Gilbert & Sarkar, 2000; Keller, 2005; Lickliter & Honeycutt, 2003; Lickliter & Witherington, 2017; Moore, 2016, 2024).
Epigenetics
Although it was apparent to some early geneticists that phenotype construction is a complex process that is never determined strictly by genetic factors, this basic fact has been thrown into high relief in the past 25 years, as we have learned more about the epigenetic control of gene expression. The prefix “epi” refers to something on or over something else, so the word epigenetic in the current context refers to factors “above” DNA that influence the functioning of the genome, however directly or indirectly (for more information on the several available definitions of “epigenetics,” see Moore & Flom, 2020). In some cases (e.g., DNA methylation), these factors are literally on DNA. In other cases, they are “higher” than DNA in a less literal sense; such factors operate at higher levels of analysis and include molecules like hormones that are involved in stress responses, cytokines that are involved in inflammatory responses, and dietary factors like vitamins. In every case, these epigenetic factors contribute to the regulation of genetic activity. Specifically, they increase or decrease the speed at which gene products—like proteins or functional RNAs—are produced. They can even modify the expression of genes in more striking ways, effectively turning those genes “off” or “on” (Moore, 2017a). Discoveries about the epigenetic regulation of genetic activity have had a remarkable effect on thinking about genetic contributions to phenotypes. If a gene can be “turned off” in an epigenetic process, then the gene’s presence in an individual’s genome can be inconsequential. As a result, whereas it once seemed like what really matters in phenotype development is what genes you have, what matters now is what those genes are doing. Merely having a gene need not be predictive of any particular developmental outcome. (The precisely specified mechanisms by which epigenetic factors affect genetic activity are beyond the scope of this article; interested readers can consult Moore (2015) for more details.)
In some cases, the factors that influence gene expression are the products of other genes (e.g., transcription factors and long non-coding RNAs). But in other cases, the influences on gene expression can be traced to non-genetic factors, such as light (e.g., Naruse et al., 2004), nutrients (e.g., Waterland & Jirtle, 2003), or pollutants (e.g., Crews et al., 2007), all of which reflect the individual’s broader environmental or experiential contexts (Moore, 2013a). Some of the other factors of this type that are capable of altering genetic activity are exercise, exposure to drugs, and some early-life experiences (Moore, 2015). This is why research on the epigenetic control of gene expression has proven to be of particular interest to psychologists: our experiences have considerable power over how our genomes function (Lester et al., 2011; Weaver, 2007). Epigenetic processes play important roles in development starting at conception (Gilbert, 1992) and continuing throughout our lifetimes. In addition, these processes occur in every part of the body, including the brain, so they are able to influence our thoughts, feelings, and behaviors (Lester et al., 2011; Moore, 2015, 2017b, 2024).
Behavioral epigenetics has sparked interest among psychologists for several reasons, one of which is the possibility that this work could help explain the unforeseen lack of concordance often found in monozygotic (i.e., MZ, or “identical”) twins. Because MZ twins share nearly identical genomes whereas dizygotic (i.e., DZ, or fraternal) twins do not, many theorists long assumed that MZ twins would develop very similar traits more often than DZ twins do. While MZ twins are typically more similar to one another—unsurprisingly, given that genetic factors do contribute to our phenotypes, and MZ twins have exceedingly similar genomes—empirical studies have often found that they are less similar than expected (Caspi et al., 2004; von Stumm & Plomin, 2018). Research on epigenetics has revealed one potential explanation for such discordance.
In a study of 80 Spanish MZ twins, Fraga and colleagues (2005) found that young individuals within a twin-pair were “epigenetically indistinguishable” from one another (p. 10604). However, as they aged, twins in a pair diverged epigenetically in noteworthy ways. Even more telling was the fact that twins in a pair “who were older, had different lifestyles, and had spent less of their lives together” (p. 10608) were more epigenetically different than were twins who had shared similar lifestyles. Thus, life circumstances are able to alter epigenetic states, which can be expected to lead to altered gene expression profiles and different observable characteristics. Other studies have reported that different experiences early in life (e.g., exposure to higher versus lower socioeconomic status, exposure to caring versus abusive adults, etc.) can also lead to longstanding phenotypic differences via epigenetic alterations, strengthening the impression that our experiences can effectively “get under our skin” and influence our traits, in some cases producing effects that are detectable decades later (Borghol et al., 2012; Heijmans et al., 2009; Suderman et al., 2014).
Epigenetic effects occur on relatively short timescales as well. When travelers cross several time zones in a short period, they typically experience jet lag; their circadian rhythms initially remain entrained to the time zone they departed from, and not until some days later do their bodies adjust to the new time zone, allowing them to operate on the same schedule as the locals. Research with rodents revealed that mammals begin to adjust to new schedules when we are exposed to light at times when our circadian rhythms anticipate darkness (Reppert & Weaver, 2002). Specifically, this unexpected exposure to light leads to epigenetic changes that drive increased expression of “clock genes” in neurons, genes that are involved in the control of circadian rhythms (Naruse et al., 2004; Rusak et al., 1990). In this way, exposure to a simple visual stimulus brings about the rapid biological change required to adjust to a local time zone. Together with other research results, these findings suggest that our epigenetic states may be perpetually changeable, and that stimulation of our neurons can alter the epigenetic state of those neurons’ DNA, altering how those neurons function. Accordingly, this jet lag example reveals a mechanical way in which our experiences can alter how our brains operate, physically incorporating into our brains information about the world we inhabit.
Another line of research on behavioral epigenetics revealed that when newborn rat pups are groomed by their mothers, this experience alters their epigenetic states in ways that make them less reactive to stress later in life (Meaney, 2017). Rat pups that experience less maternal grooming typically grow up to have increased stress reactivity in adulthood (Weaver et al., 2004). But it is not just social interactions early in life that can have these kinds of significant effects. Cole (2009) documented how a person’s sense that they are lonely influences their epigenetic state in ways that help explain why they are more susceptible to viral infections. By the 1990s, researchers knew that social isolation is associated with increased replication of viruses that cause the common cold, AIDS, and other disease states, leading to poorer health. Cole and colleagues (2007) found that people who reported feeling lonely had reduced expression of genes involved in immune responses, offering an explanation for these people’s increased vulnerability to infection. Notably, this effect was a function of the individuals’ perception of their social isolation, not of the number of people with whom they were actually socially connected. That is, what mattered more than objective reality was a person’s psychological state. Capturing the essential insight of behavioral epigenetics, Cole (2009) wrote “Because the molecular composition of our cells constitutes the physical machinery by which we perceive and respond to the world around us, and because that molecular composition is itself subject to remodeling by socioenvironmental influences, gene expression constitutes both a cause and a consequence of behavior” (p. 135).
As the subdiscipline of behavioral epigenetics grew in the first 25 years of this century, it became apparent that there are significant correlations between epigenetic modifications to the genome and a wide variety of human psychological phenomena. For example, empirical or theoretical work has considered the epigenetic components of all of the following disorders: schizophrenia and bipolar disorders (Chen et al., 2021; Dempster et al., 2011; Labrie et al., 2012), depression (Mill & Petronis, 2007; Mitsis et al., 2023), addiction (Hamilton & Nestler, 2019; Wong et al., 2011), eating disorders (Campbell et al., 2011; Steiger et al., 2023), suicidal behavior (Dwivedi et al., 2025; Keller et al., 2010; Poulter et al., 2008) and some age-dependent or neurodegenerative memory impairments, such as dementia (MacBean et al., 2020) or Alzheimers disease (Villa & Combi, 2024). Epigenetic effects have also been associated with experiencing abuse as a child (e.g., Lawn et al., 2018) or intimate partner violence as an adult (González-Martínez et al., 2024). Crucially, epigenetic effects are also central in normal psychological processes, including memory formation (Day & Sweatt, 2010, 2011), emotional functioning (Assis et al., 2025), regulation of circadian rhythms (Naruse et al., 2004), and pair-bonding (Wang et al., 2013). At this point, it is increasingly recognized that epigenetic factors are integrally involved in the emergence of all psychological phenotypes, whether they are psychopathological or not.
It is evident that gene expression can be epigenetically regulated by factors originating in individuals’ environments, including sensory stimuli (e.g., light, sound, pressure on the skin), diet (Choi & Friso, 2010), drugs (Hamilton & Nestler, 2019; Maze & Nestler, 2011), pheromones (Cardoso-Junior et al., 2020), or environmental pollutants (Anway et al., 2005). Furthermore, the functioning of an individual’s genome can also be influenced by the individual’s own behavior, such as through exercise (Fernandes et al., 2017) or exploring an enriched environment (Fischer et al., 2007). The realization that DNA is an inert molecule that has the effects it does only because of its interactions with other molecules—some of which reflect the body’s broader context—has gone a long way toward eliminating the perception that DNA can dictate phenotypic outcomes in context-independent ways. In the wake of this failure of genetic determinism, researchers have increasingly recognized that understanding psychological phenotypes demands enquiries into how genetic and non-genetic factors co-act to bring those phenotypes into being.
At the same time, because all of our thoughts, feelings, and behaviors are dependent in part on the structure and functioning of our brains—a biological entity—strict environmentalism has failed as well. One consequence of the failure of both genetic and environmental determinism is that psychologists should acknowledge that our predictions about behavior will, at best, always be probabilistic. Gottlieb suggested as much decades ago, when he noted that development is best described by the words “probabilistic epigenesis” (Gottlieb, 1998, 2007). Thus, phenotypic outcomes are not predetermined but instead emerge in developmental time through epigenetic processes involving bidirectional influences across multiple levels of analysis, as captured in Gottlieb’s 1991 sketch (see Figure 2).
The probabilistic nature of development means that we need to proceed with extraordinary care (if at all) when using measures of children’s psychological characteristics to predict their prospects. So far, these measures cannot forecast individuals’ futures in accurate ways. Even the latest statistical tool designed to extract genomic information with the goal of making such predictions—polygenic scores (PGSs)—cannot accurately predict developmental outcomes for individuals (Harden & Koellinger, 2020; Morris et al., 2020). Moreover, these scores cannot be understood as revealing even just a “genetic predisposition” in a person (Furrer & Turkheimer, 2026). This should not surprise us, as PGSs do not take epigenetic phenomena or developmental contexts into account at all (Moore, 2023). Recognizing the essential role of developmental processes in phenotype construction, Burt (2023) provided a powerful critique of PGSs, writing that “the potential scientific rewards of adding PGSs to social science are few and greatly overstated and the scientific costs, which include obscuring structural disadvantages and cultural influences, outweigh these meager benefits for most social science applications” (Burt, 2023, p. 1).
In addition to being wary of attempts to predict developmental outcomes starting in infancy or early childhood, it would probably be wise to remain skeptical of constructs that assign static traits to individuals. Developmental changes continue to accrue across the lifespan (Mousley et al., 2025), and because decades of research have revealed the astonishing plasticity of neural structures and functions (Eagleman, 2021; Kandel et al., 2021), our behavioral characteristics remain open to change. Of course, there is a long history in psychology of characterizing individuals as possessing static traits. Yet the developmental perspective, which can be valuably applied across the field’s subdisciplines, suggests that we might more profitably view individuals’ psychological phenotypes as changeable, albeit sometimes in states of stable equilibrium (or “attractor states,” to use the terminology favored by dynamic systems theorists like Thelen & Smith, 1994), states that simply appear static.
Conclusion
All psychological characteristics reflect the history and current presence of (and relations between) an immense number of factors, including biological molecules like hormones, neurotransmitters, and DNA, cells like neurons, myocytes, and pancreatic beta cells, the presence in our environments of distinctive foods, social partners, and meteorological climates, the rewards and punishments meted out by societies, the traditions of cultures, and more. Because these same factors contribute to most—if not every one—of our psychological traits, the field of psychology should be unifiable. After all, the overarching message of DS theorizing is that each human being (along with their contexts) is a single cohesive, integrated system with inextricably linked component parts (Lickliter, 2009). Unification of psychology should be considered an attainable goal (Michel, 2013, 2026).
Nonetheless, the exceptional complexity of these systems means that a common understanding of all psychological phenomena will be difficult to achieve in practice. The sheer number of relevant influences on behaviors, thoughts, and feelings, coupled with the awe-inspiring complexity of the processes that give rise to these phenomena means that researchers, clinicians, and other individuals who are hoping to operate within a single psychological theory are in a difficult spot. As it happens, this state of affairs explains why scientists still understand the developmental origins of such a relatively small number of traits. Even so, the fact that all psychological phenotypes develop at least opens the possibility of unifying psychology with a developmental perspective. At present, this perspective can leave researchers, clinicians, and other individuals with at least a few helpful hints.
First, the DS perspective encourages us to recognize that because we are inherently dynamic entities in continual interaction with our environments, what we do is extremely important. It is easy to lose sight of this fact when we regularly encounter claims that our unalterable genetic constitutions determine aspects of our futures. Although we obviously cannot gain complete control of our futures, the fatalism of genetic determinism is demonstrably unwarranted. We should always remain aware that changing one’s context—including one’s diet, exercise, social partners, dwelling place, sleep schedule, drug use, job, country of residence, etc.—can have remarkable effects on our behaviors and psychological states. And even if changes to these states sometimes occur at timescales that make the changes difficult to detect, we should remember that people are constantly changing.
Second, no one should credulously accept a claim that a gene can be the sole cause of a phenotype, because genes always operate as they do in part because of their spatial and temporal contexts. Believing that a gene is (or genes are) responsible for a phenotype in a context-independent way renders clinicians relatively helpless in the face of a behavioral disturbance, and it renders researchers less likely to search for the actual developmental cause of the phenotype. As Lehrman wrote over 70 years ago, any theory that conceives of behaviors as “immanent, preformed, [or] inherited … is bound to divert the investigation of behavior development from fundamental analysis … [Such a theory] inevitably tends to short-circuit the scientist’s investigation of intraorganic and organism-environment developmental relationships which underlie the development” of such behaviors (Lehrman, 1953, p. 359). If a researcher wants to discover a good treatment option for a particular psychopathology, that researcher should study how the pathology develops rather than merely searching for DNA segments correlated with that pathology. Asking questions about the development of phenotypes will invariably reveal a multi-factor (and typically biopsychosocial) process that actually causes the phenotype. Consequently, the results of experimental (rather than correlational) studies of development will increase the likelihood that interventions will be discovered that can ease suffering.
Third, because phenotype development results from events occurring at more than one level of analysis, understanding these processes will typically require interdisciplinary teams of researchers. Therefore, research psychologists should endeavor to constitute (or join) teams of researchers that include biologists as well as social scientists with expertise beyond individuals’ behaviors. If a team of developmentally oriented researchers studies how a psychopathological phenotype emerges in real-time from the co-actions of myriad factors inside and outside of the body, they will always discover multiple influential factors each of which will suggest a possible way to intervene helpfully. Multidisciplinary approaches to scientific work have become increasingly common in recent decades and seem likely to characterize successful research endeavors in the future. If we pull researchers with diverse intellectual backgrounds under the same, increasingly large umbrella, unification of our field will become increasingly likely.
Even though elucidating the developmental processes that build phenotypes will be arduous, it is important to rise to the challenge. Because if we fail to focus on the processes that build our traits in development, we are at risk of examining someone’s genome on the day they are born—or, for that matter, the socioeconomic context into which they are born—and telling them that they are doomed to success or failure, when in fact they might not be. And this would often lead to an unhappy ending in either case. For the moment, at least, we should refrain from projecting confidence about our predictions for individuals, as we currently understand the developmental origins of so few of our psychological characteristics.
Footnotes
Funding
The author received no financial support for the research, authorship, and/or publication of this article.
Declaration of Conflicting Interests
The author declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.