
Abstract
Background:
Anemia and micronutrient deficiencies remain major public health concerns among children in Ghana. Nutrient-rich food fortification offers a promising strategy to improve iron status and reduce inflammation. This study evaluated the effect of a 24-week egg powder–fortified Tom Brown meal on hemoglobin (Hb), serum ferritin, and C-reactive protein (CRP) levels among school-aged children in the Wa Municipality.
Methods:
A randomized controlled trial was conducted with 136 children aged 5 to 12 years. Children in the intervention group received egg powder–fortified Tom Brown 3 days/week for 24 weeks, while the control group consumed unfortified Tom Brown. The Shapiro–Wilk test (P > .05) validated the normal distribution of all biochemical variables, thereby endorsing the application of parametric analyses. Baseline comparisons between groups showed no significant differences in Hb, ferritin, or CRP levels.
Results:
After the 24-week intervention, hemoglobin levels increased from 11.81 ± 0.83 to 14.67 ± 0.93 g/dl (P < .001; d = 2.602) in the intervention group, while in the control group, hemoglobin levels increased from 11.43 ± 0.85 to 12.53 ± 0.54 g/dl (P > .051; d = 0.519). Ferritin levels increased from 14.32 ± 2.07 to 18.87 ± 3.82 ng/ml (P < .001; d = 2.640) in the intervention group, compared to an increase from 12.24 ± 2.72 to 15.10 ± 2.11 ng/ml (P > .052; d = 0.699) in the control group. In contrast, CRP levels decreased from 5.87 ± 1.67 to 3.93 ± 0.99 mg/l (P < .001; d = −1.542) in the intervention group, while in the control group, CRP levels decreased from 4.97 ± 1.148 to 3.911 ± 0.93 mg/l (P > .052; d = −0.719).
Conclusion:
The 24-week egg powder–fortified Tom Brown intervention resulted in substantial improvements in hemoglobin and iron stores and a marked reduction in inflammation in school-aged children. The findings revealed that fortifying and integrating affordable, locally acceptable foods into school-based feeding programs can be a high-impact, scalable public health intervention for reducing anemia and micronutrient deficiencies in resource-limited settings.
Background
Anemia and micronutrient deficiencies are still major public health issues globally, disproportionately affecting children and women in low- and middle-income countries. Approximately 40% of children worldwide are anemic, with iron deficiency accounting for nearly half of these cases. 1 Globally, iron deficiency anemia (IDA) affects over 1.6 billion people and is associated with impaired cognitive development, reduced physical capacity, and increased morbidity and mortality. 2 Despite decades of interventions, progress in reducing anemia has been slow due to its multifactorial etiology, including inadequate dietary intake, infections, genetic predispositions, and socio-economic restrictions.3,4 Iron supplementation strategies, which have become the convention in combating anemia, are further constrained by side effects and reduced effectiveness in populations with high infectious disease burdens. 5
Micronutrient deficiencies, especially iron, vitamin A, and zinc, often frequently coexist and are exacerbated by low dietary diversity and reliance on nutrient-poor staple foods, contributing to widespread “hidden hunger” and increased susceptibility to infections.6,7 These conditions are particularly severe in low- and middle-income environments, where undernutrition contributes significantly to preventable child mortality.8,9
In Sub-Saharan Africa, anemia prevalence among children often exceeds 50%, driven by complex interconnected health factors such as inadequate dietary intake, malaria, parasitic infections, and chronic inflammation.10,11 The region endures a disproportionate share of the global burden, with persistently high prevalence across population groups despite ongoing nutrition interventions, which highlights the inherent limitations in current intervention strategies. 12
Similarly, anemia remains a critical public health concern in Ghana, particularly among children and adolescents, with consistently high prevalence being reported across national and sub-national studies.13-16 A combination of factors such as inadequate intake of iron-rich foods, food insecurity, infections such as malaria, and dietary inhibitors of iron absorption serve as drivers of anemia burden in Ghana, demonstrating the complex interaction between nutritional, environmental, and socio-economic factors.10,17
Although interventions such as food fortification and micronutrient supplementation have demonstrated potential in improving iron status, their effectiveness in many African settings remains inconsistent.18,19 Enhancers such as NaFeEDTA, ascorbic acid and microbial phytase have been shown to improve iron absorption, while food-based approaches, including fortified maize-meal porridge and animal-source ingredients such as bovine blood powder, have yielded promising results. 20 However, these approaches often fail to account for physiological constraints that limit iron utilization in infection-prone individuals and populations.
Research has shown that iron metabolism in such contexts is strongly influenced by inflammatory pathways and iron homeostasis is regulated by the hepcidin–ferroportin axis, and infection-induced increases in hepcidin reduce intestinal iron absorption and iron availability for erythropoiesis. 21 In malaria-endemic settings such as northern Ghana, chronic low-grade inflammation may therefore attenuate the effectiveness of conventional iron interventions. 22 Also, from a nutritional and biochemical perspective, the dietary matrix affects both iron bioavailability and utilization. The body absorbs heme and protein-bound iron sources more efficiently and is less affected by inhibitors such as phytates and polyphenols than it is by non-heme iron. 23 Furthermore, high-quality dietary proteins and essential amino acids enhance erythropoiesis by improving the synthesis of globin and overall hematopoietic function. Eggs provide highly digestible protein and key micronutrients, including vitamin B12, riboflavin and choline, which are essential for red blood cell production. 24 Additionally, eggs may also exhibit anti-inflammatory and antioxidant properties that could enhance iron utilization in inflammation-mediated anemia. 25
Notwithstanding the progress made, nutrition interventions in Ghana have largely focused on pharmacologic iron delivery or generic micronutrient fortification, with limited emphasis on integrated food-based strategies that simultaneously address iron bioavailability, protein quality, and inflammation. 26 This gap is particularly evident among school-aged children, a group that remains neglected but vulnerable due to rapid growth, increased cognitive demands and continued exposure to infectious diseases. 26
The traditional, conventional Tom Brown, a widely consumed cereal-based food in northern Ghana, though it offers a platform for food-based intervention, is often limited by low iron bioavailability due to its plant-based composition and high levels of phytate. Fortification with egg powder, a nutrient-dense animal-source ingredient rich in highly bioavailable protein, iron and erythropoietic micronutrients, presents a biologically plausible and culturally acceptable strategy to improve iron utilization and hematologic consequences. 27
However, empirical evidence on the hematologic and biochemical effects of egg powder-fortified Tom Brown among school-aged children remains limited, particularly within the Wa Municipality of Ghana. This study therefore addresses a critical gap by evaluating the effectiveness of this food-to-food fortification approach in improving anemia outcomes and supporting iron utilization among school-aged children in a high-burden, infection-prone background. In doing so, the study sought to provide evidence to address the following critical questions:
Whether daily consumption of egg powder–fortified Tom Brown significantly improves hemoglobin levels among school-aged children in the Wa Municipality; and
How the intervention affects serum ferritin and other biomarkers of iron status.
Materials and Methods
Study Design
The study was a randomized controlled trial conducted in the Upper West Region, Ghana. Using a computer-generated random sequence, eligible children who met the inclusion criteria were assigned to the intervention group, which received egg powder-fortified Tom Brown, or the control group, which received unfortified Tom Brown. To guarantee allocation concealment and reduce bias, the allocation was carried out by an independent research assistant who was not involved in participant recruitment, intervention administration, or outcome assessment. Group allocations were only communicated to participants and their caregivers upon enrollment. This process minimized selection bias and ensured that the groups were comparable at baseline for a valid assessment of the effects of the intervention.
Study Area
The study was conducted in the Wa Municipality of the Upper West region of Ghana. The municipality is predominantly peri-urban, with a significant proportion of the population engaged in subsistence farming and petty trading. The area is characterized by a high burden of infectious diseases and nutritional deficiencies, making it an ideal setting for studying anemia.
Study Population
The target population included school-aged children (
Sample Size Determination
The sample size for this study was calculated to detect a clinically meaningful difference in mean hemoglobin (Hb) levels between the intervention and control groups. Previous studies among school-aged children reported a baseline mean Hb of 12.65 g/dL. For this study, the standard deviation (SD) of Hb was assumed to be 0.96 g/dl. 28 A clinically relevant absolute difference of 0.5 g/dl in mean Hb between groups was considered appropriate, reflecting the expected improvement from the egg-powder–fortified Tom Brown intervention.
The minimum sample size per group was calculated using the formula for comparing 2 independent means:

Zα/2 = 1.96 (5% significance level)
Zβ = 0.84 (80% power)
SD = 0.96 g/dl
Δ = 0.5 g/dl (absolute difference)
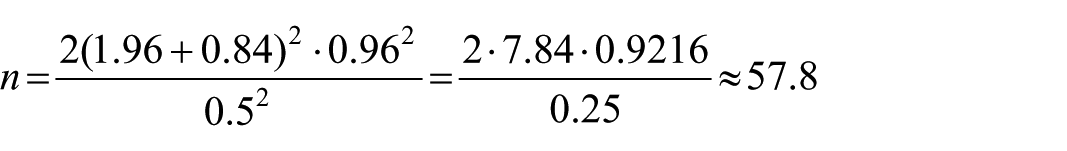
Thus, approximately 58 participants per group were required.
To account for a 15% attrition rate, the adjusted sample size was calculated as:


Rounding it up, the final sample size was set at 136 participants, which was considered sufficient to detect meaningful changes in Hb and iron status biomarkers while accounting for possible lost-to-follow-up participants. Thus, a total of 136 school-aged children were recruited, with their parents or caregivers serving as respondents for relevant questionnaires; 15 were lost to follow-up, and 121 children completed the study, and their data were included in the final analysis.
Inclusion and Exclusion Criteria
All school children aged 5 to 12 years old in public government primary schools in the Wa Municipality were eligible to participate in the study. Children of the same age group in private primary schools were not allowed to take part in the study. Furthermore, children with special conditions, those with known allergies to egg and egg products, those on special medications, and children with special disabilities that may interfere with their physical activity levels or whose parents/guardians declined to consent to their participation were exempted from the study.
Sample Recruitment
Control of Confounding Factors
Baseline nutritional status was assessed using anthropometric measurements (weight, height, and BMI-for-age Z-scores). Inflammation was evaluated using C-reactive protein (CRP), and ferritin values were adjusted using the BRINDA regression correction approach. Participants' screening was employed to gather information on recent illness, medication use, and dietary supplement intake. Children taking iron supplements or medications that could impact iron metabolism were excluded from the study. Individual randomization was used to reduce the impact of unmeasured variables like physical activity. To further ensure that all study participants were comparable at baseline, participants were screened for early signs of anemia prior to enrollment. Even though the WHO criteria were used for defining anemia, Hb <13 g/dl was used as a screening threshold to include children who were anemic and those at risk. Thus, participants with hemoglobin levels above 13 g/dL were excluded from the study, ensuring that only those who were anemic or borderline were included. This procedure helped to reduce baseline variability in iron status among participants and strengthened the internal validity of the effect of the intervention by ensuring that all participants started from a similar physiological status. Final Hb classification followed WHO standards (Figure 1).
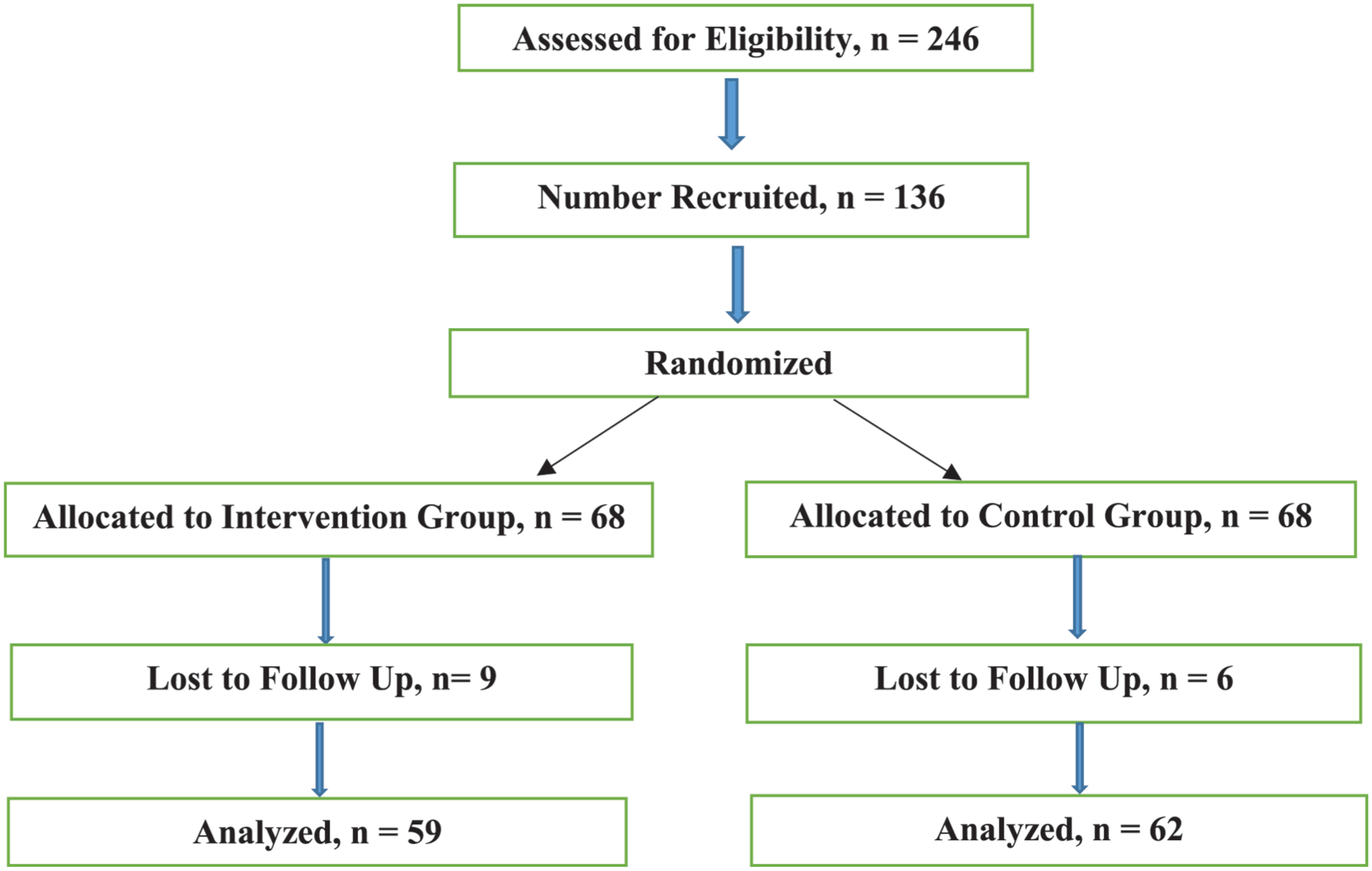
Flow diagram of the sample recruitment process.
Justification for Selection of biomarkers
Hemoglobin and serum ferritin were selected because they are widely recommended indicators for assessing anemia and iron status in population-based studies. Hemoglobin reflects functional anemia status, while ferritin provides an estimate of body iron stores. However, ferritin is an acute-phase reactant and may be elevated during inflammation; therefore, CRP-adjusted BRINDA correction was applied to improve validity.
Roasted Corn Flour Preparation
The ingredients used for the formulations are roasted corn flour produced from local corn from the Wa market; whole egg powder produced commercially using the spray-drying technique and with eggs of chickens from Emmppek Farms, Nigeria, Delta State; and a little sugar and salt to enhance the flavor and taste. The Tom Brown used in this study was made from locally produced corn. Yellow corn was purchased from the WA market, the bad ones carefully removed, and the remainder was roasted using dry heat in a gas-operated oven at a temperature of 150°C to 200°C. During roasting, the corn was stirred or turned to ensure even roasting and to avoid burning. The roasting process was deemed completed when the corn turned brown in color. The roasted corn was then allowed to cool down and milled into flour. 29
Egg Powder Preparation
The spray drying process involves the use of hot gas to convert liquid eggs into dry egg powder. The liquid feed (liquid egg) is prepared by passing it through pasteurization to retain its composition and consistency and ensure safety. It is atomized into droplets to increase the surface area for efficient drying. The nozzle atomizer approach is used where high-pressure nozzles are employed to create a fine spray. In the drying chamber, hot gas (air) is introduced where the temperature and flow rate of the hot gas are carefully controlled and the droplets are allowed contact with the hot gas, causing rapid evaporation of the moisture. The loss of moisture from the droplets results in solid particle formation with shapes and sizes that are influenced by factors such as feed properties, atomization, and drying conditions. The dried particles are then separated from the drying air using a cyclone separator, and the particles are collected. The dried egg powder is then collected and milled to obtain uniform particle size. Airtight, Ziplock-like bags are then used to store the powder to prevent moisture absorption and contamination. 30
Tom Brown Preparation, Sensory Evaluation and Acceptability Test
The Tom Brown was prepared with a formulation ratio of 100 g of roasted corn flour and 20 g of egg powder, respectively. The formulation ratio was scientifically arrived at after a sensory evaluation and acceptability study was conducted on the intervention product, the results of which are presented in another manuscript.
Study Intervention Delivery
The Tom brown was prepared daily by the researcher and his team, using a formulation ratio of 100 g of roasted corn flour to 20 g of egg powder. The prepared Tom Brown was then transported to the various schools and served to the participants by the team, who observed them consume it before departing to the next school to ensure compliance. The intervention group received 250 ml of fortified porridge three days per week for 24 weeks, while the control group received 250 ml of unfortified Tom Brown porridge. The trial was duly registered with the Pan African Clinical Trial Registry (registration number PACTR202408897182810).
For each batch, the porridge was prepared by mixing 2000 g of roasted corn flour and 400 g of egg powder with 4800 g of water. This yielded a per-serving formulation of 100 g roasted corn flour combined with 20 g of egg powder, with water added after the dry ingredients were thoroughly blended to ensure uniform distribution of micronutrients.
The mineral analysis conducted on the egg powder-fortified Tom Brown and the calculations made revealed that 250 ml of the Tom Brown provided 6.66 mg. The average RDA for iron in children aged 5 to 12 years is 9 mg per day. Each serving of 250 ml of Tom Brown porridge to the children provided 6.66 mg of iron, which was equivalent to 74% of the RDA if consumed each day of the week. However, given that it was consumed 3 times per week, the average daily contribution was approximately 2.85 mg/day, equivalent to 32% of their daily RDA. With regard to energy content, the conversion factors for protein, fat and carbohydrate were used to calculate the energy contribution of the Tom Brown, yielding a total energy of 573 kcal per 250 ml serving a day, translating into about 34.7% of the average RDA of energy for children 5 to 12 years old. The total volume of Tom Brown was prepared daily by mixing 2000 g of roasted corn flour and 400 g of egg powder with 4800 g of added water. To estimate the daily protein contribution of Tom Brown, the total protein content in corn flour and egg powder was first measured in their dry form. This amount was then adjusted to account for the water used during cooking, resulting in a daily protein supply of 17.34 g. 31 In each batch, 20 g of egg powder was combined with 100 g of roasted corn flour to fortify the composite Tom Brown. The addition of iron, zinc, magnesium, and high-quality protein from the egg powder enhanced the overall nutritional profile of Tom Brown. To produce the porridge, water was added after the dry ingredients had been blended equally to complete the fortification. Every 250 ml serving provided approximately 6.66 mg of iron (74% of RDA), 17.34 g of protein, and 573 kcal, as determined by proximate and mineral analyses. This method ensured uniform distribution of micronutrients across servings. Further details of the analytical results and optimization of the formulation are presented in our previous publication 32
Outcome assessors, laboratory analysts, and data analysts were blinded to group allocation. Due to the nature of the food-based intervention, complete blinding of participants and caregivers was not feasible, as the fortified and unfortified Tom Brown could differ slightly in sensory characteristics. However, efforts were made to minimize performance bias by ensuring that both porridges were prepared and served in a similar manner, with comparable appearance, texture, and taste to reduce the likelihood of participants distinguishing between groups.
Compliance with the intervention was closely monitored throughout the study. The porridges were administered under supervision at school, and home follow-ups were conducted for absent pupils to ensure consistent intake and minimize missed doses. The intervention was also monitored for adverse effects. No changes were made to the study design, methods, or outcome measures after trial commencement, and no interim analyses or stopping guidelines were applied, as the trial proceeded as planned until completion.
Measurements
Blood samples from study participants were collected by trained laboratory technicians before and after the feeding intervention. The endline blood sample was collected after the 24 weeks of the Tom Brown feeding intervention. Again, the Hb of study participants was taken by trained nurses using the Hb meter, the HemoCue, at baseline and at endline. The collected blood samples were processed into serum and frozen at the Wa Municipal Hospital laboratory and later transferred to the UDS Tamale campus laboratory for analysis.
Laboratory Procedures
Determination of Ferritin and C-reactive Protein Concentrations
The study employed the standard sandwich ELISA technique to measure ferritin concentrations using commercially prepared kits, ELISA kits (Biobase Biotech [Melson Shanghai] Ltd., China), following established scientific protocols for immunoassay measurement. All procedures were carried out according to the manufacturer’s instructions. The serum ferritin concentration was determined using a microplate ELISA reader (Powean) at a wavelength of 420. The ferritin concentration in the samples was determined by comparing the optical density of the samples to the standard curve. 33 The same procedure but with different kits and reagents was used to measure C-reactive protein concentrations. The ELISA laboratory work was carried out at the University for Development Studies' Tamale campus central laboratory, Ghana. All measurements were done in duplicate.
Data Processing and Analysis
Data were cleaned, coded and analyzed using the Statistical Package for the Social Sciences (SPSS), version 21. The dataset was examined for accuracy, missing values, and outliers through frequency checks, boxplots, and descriptive summaries. All continuous variables, including hemoglobin (Hb), serum ferritin and C-reactive protein (CRP), were assessed to ensure they satisfy parametric assumptions. The Shapiro–Wilk test was used to check for normality in the baseline and endline values for each biomarker in both the intervention and control groups. Boxplots were also used to check for normality visually. Variables with P > .05 were considered normally distributed, allowing the application of parametric statistical procedures.
Descriptive statistics, in the form of means, medians, and standard deviations, were computed to summarize participant biomarker concentrations. Differences between the intervention and control groups at baseline and endline in Hb, ferritin and CRP levels were assessed using independent samples t-tests to compare whether mean biomarker concentrations differed significantly between groups before and after the intervention. Within-group changes over time were also assessed with paired-samples t-tests, and mean differences and corresponding P-values were reported to determine the statistical significance of changes attributable to the intervention. Cohen’s d was used for both between-group and within-group comparisons to measure the magnitude of the effect size due to the intervention. Cohen’s d was computed using pooled standard deviations for independent comparisons, while for paired analyses it was derived from the mean change divided by the standard deviation of the difference scores. Effect sizes were interpreted using established thresholds of 0.2 (small), 0.5 (medium), 0.8 (large), and >1.2 (very large). Graphical representations with boxplots were generated to visually depict group differences and trends across time. All statistical tests were two-tailed, with significance set at P < .05. Results were presented in tables and figures.
Ethical Considerations
Ethical approval was obtained from the Institutional Review Board, the Committee on Human Research, Publication and Ethics, College of Health Sciences, School of Medicine and Dentistry, Kwame Nkrumah University of Science and Technology, Ghana. Aside from that, approval was also obtained from the Regional Health Directorate, Upper West Region, and the Wa Municipal Education Directorate. The Municipal Director of Education sent an approval letter to inform the heads of the participating schools. The parents or guardians of the children provided informed consent, and the children themselves gave their assent. The researchers strictly adhered to confidentiality and anonymity throughout the study.
Results
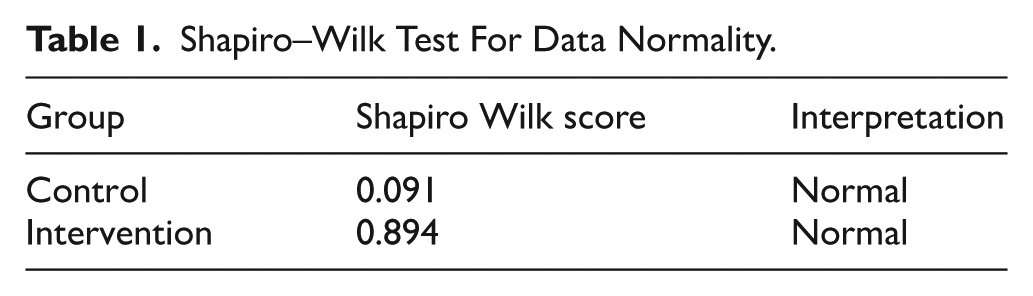
Before conducting comparative analyses, the normality of Hb, serum ferritin and C-reactive protein values at baseline and endline was assessed using the Shapiro–Wilk test, and the results showed that all outcome variables were normally distributed (Shapiro-Wilk P > .05) in both the control (P = .091 > .05) and intervention (P = .89 > .05), permitting the use of parametric tests (Table 1).
Shapiro–Wilk Test For Data Normality.
To further demonstrate the effect of the intervention on Hb levels, a graphical confirmation is provided in Figure 2, which indicates the distribution and change in Hb levels between baseline and endline for both study groups. The figure depicts that the intervention group observed measurable improvement, whereas the control group shows only a modest increase. This visual pattern thus supports the statistical findings in Table 2.
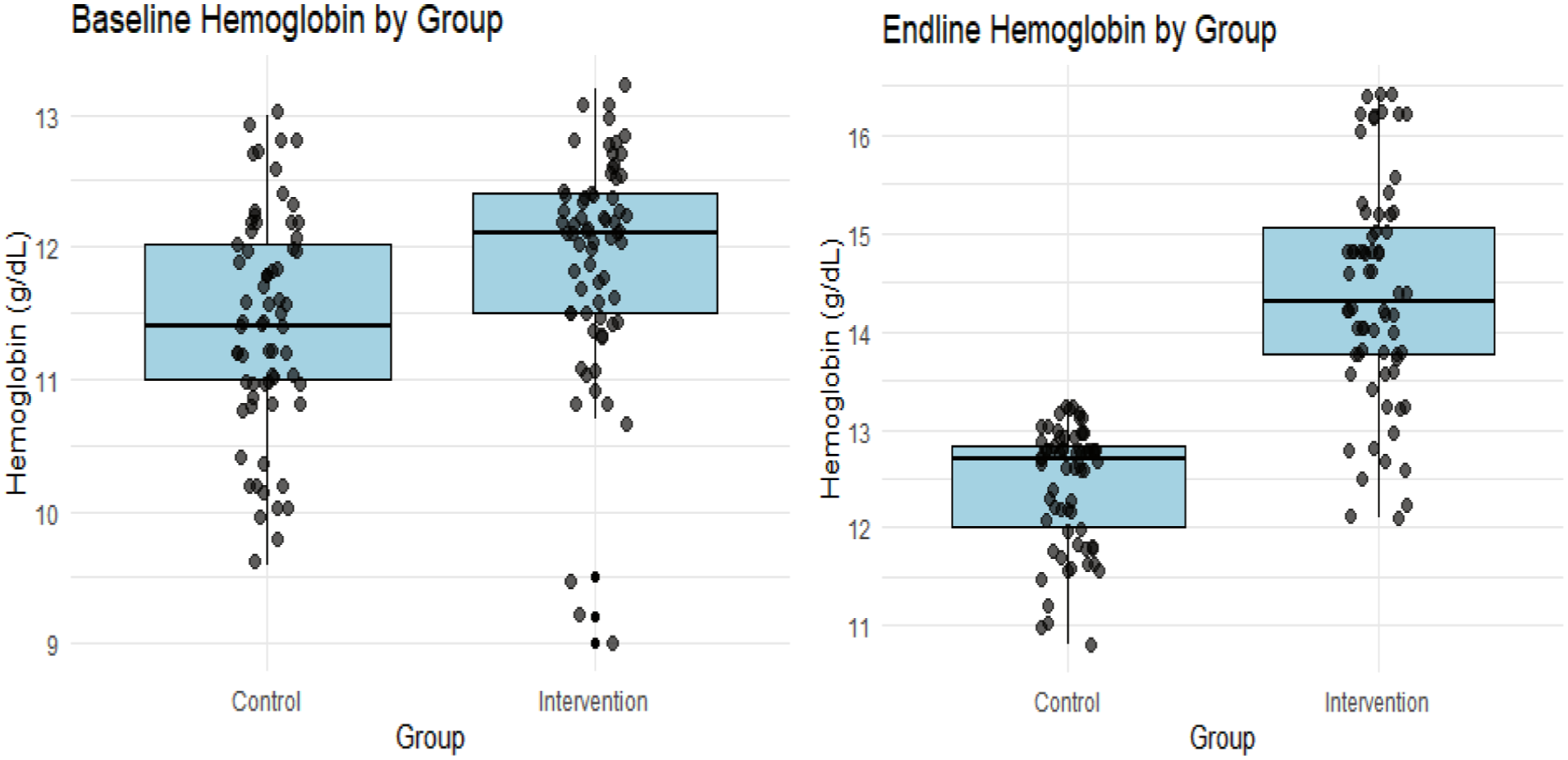
Baseline and endline hemoglobin comparison between control and intervention groups.
Between Groups Comparison of Hb at Baseline and Endline.
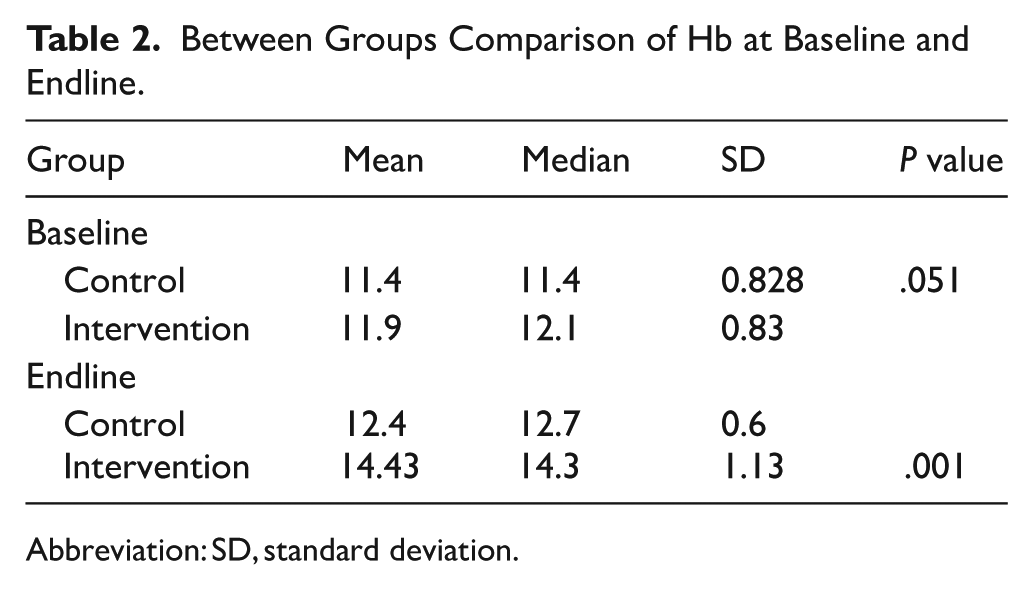
Abbreviation: SD, standard deviation.
For baseline comparison between groups, the results show that at baseline, the intervention group had a slightly higher mean Hb (11.9 g/dl) compared to the control group (11.4 g/dl), but the difference was not statistically significant (P = .051 > .05). This implies that the groups were comparable at baseline. The intervention group, however, recorded a much higher mean Hb (14.43 g/dl) at the endline compared to the control group (12.4 g/dl), and the difference was highly significant (P = .001 < .05). This demonstrates that the egg-powder-fortified Tom Brown produced a substantial improvement in Hb relative to the control group (Table 2).
The within-group comparison also revealed that the control group observed an increase in Hb, from 11.43 ± 0.85 to 12.53 ± 0.54, with a relatively small effect size (Cohen’s d = 0.519). The change was, however, not statistically significant (P = .051 > .05). The intervention group saw a higher jump in Hb, from 11.81 ± 0.83 to 14.67 ± 0.93, indicating a highly significant (P = .001 < .05) difference with a
Within Groups Comparison of Hb at Baseline and Endline.
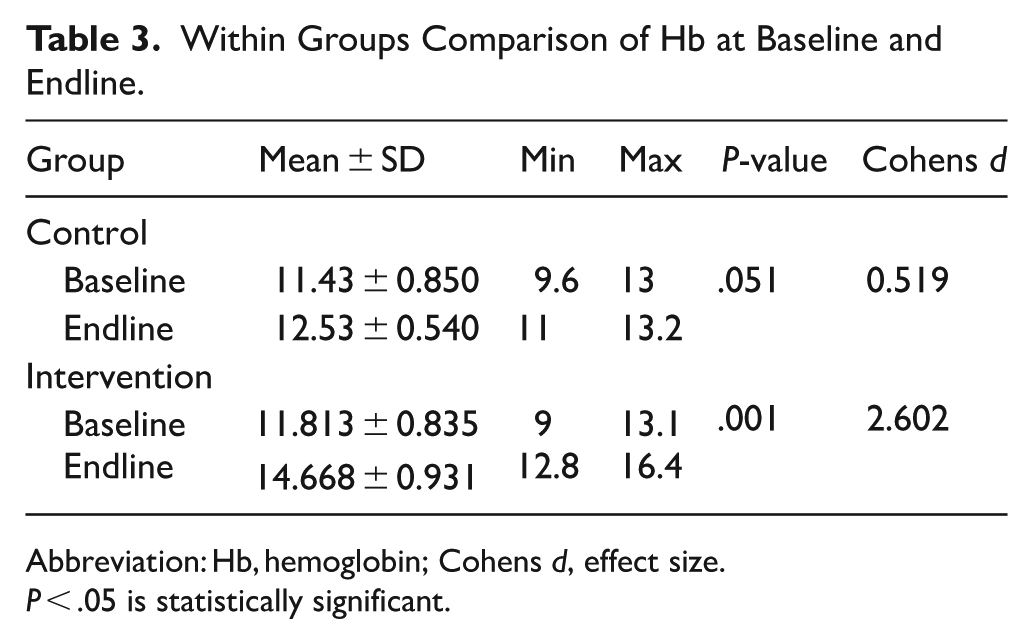
Abbreviation: Hb, hemoglobin; Cohens d, effect size.
P < .05 is statistically significant.
The ferritin data across groups were also normally distributed at both baseline and endline (P > .05), thus permitting the use of parametric analyses. Again, the virtual representations in Figure 3 further support the normality of the data.
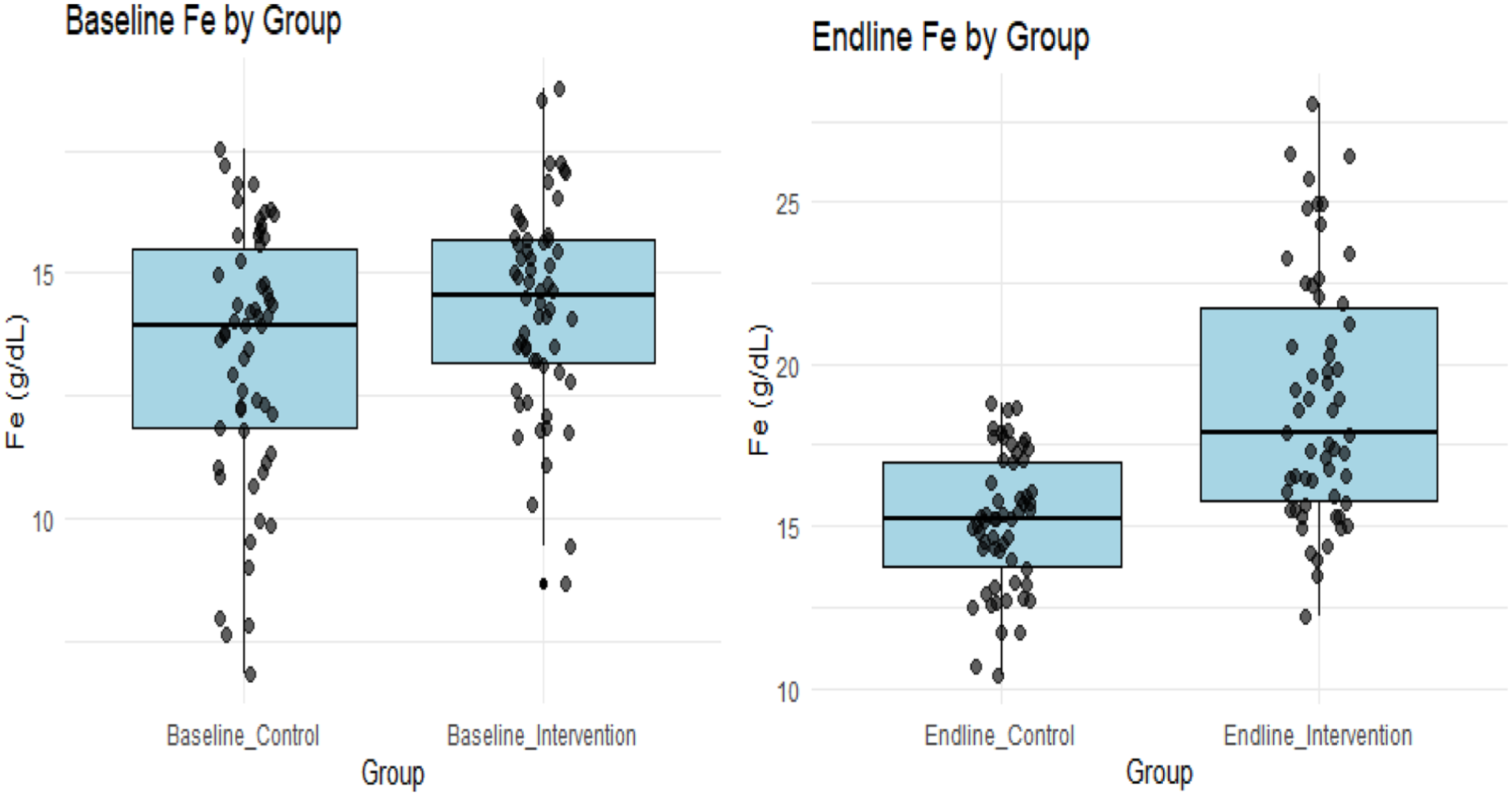
Baseline and endline ferritin comparison between control and intervention groups.
Further analysis indicated that at baseline, ferritin levels did not differ significantly between groups (P = .077 > .05). However, the intervention group at the endline had a
Between Groups Raw Serum Ferritin (Fe) Comparison at Baseline and Endline.
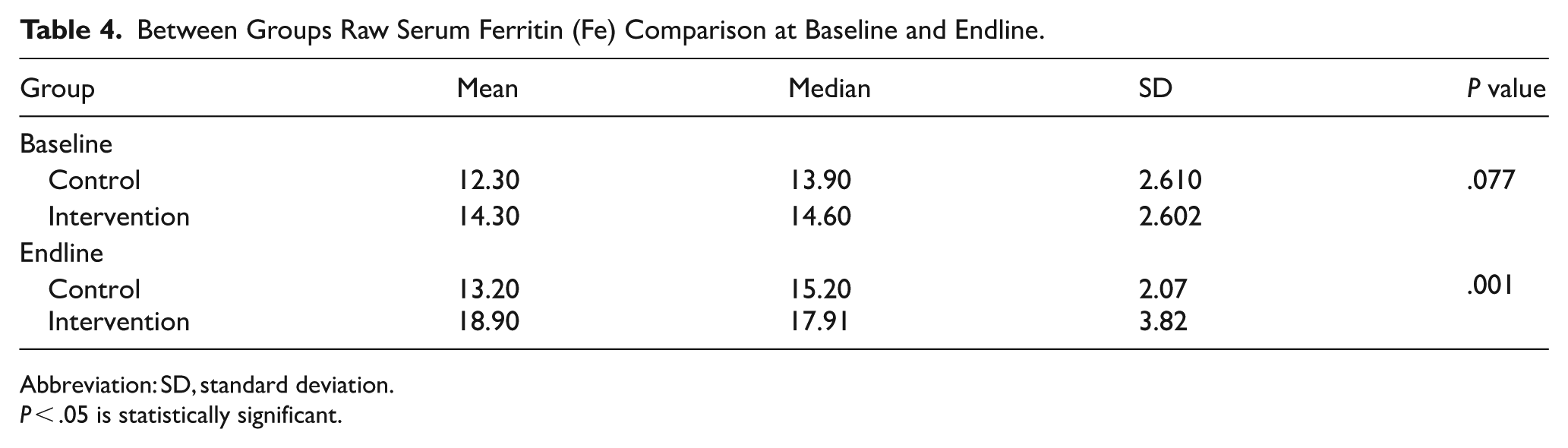
Abbreviation: SD, standard deviation.
P < .05 is statistically significant.
The serum ferritin (Fe) comparison of baseline and endline within groups demonstrated that serum ferritin showed an observable but statistically insignificant increase (P = .052) among the control group, with an appreciable effect size (d = 0.699). The intervention group, however, experienced a highly significant (P = .001 < .05) rise in ferritin, with a substantial effect size (d = 2.640) as displayed in Table 5 below.
Within Groups Raw Serum Ferritin Comparison at Baseline and Endline.
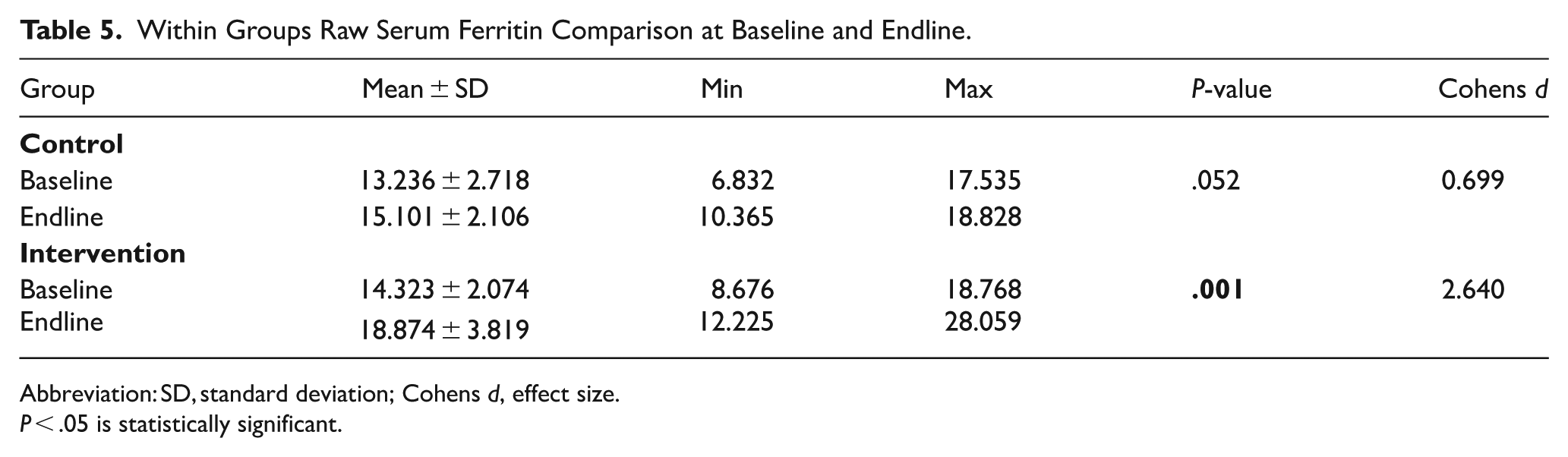
Abbreviation: SD, standard deviation; Cohens d, effect size.
P < .05 is statistically significant.
Upon adjustment for inflammation through the BRINDA approach, using C-reactive protein (CRP), ferritin concentrations improved significantly, from 11.27 ± 1.62 at baseline to 20.30 ± 4.11 µg/L at endline (mean change +9.03 ± 3.20 µg/L, paired t-test P < .001) in the intervention group. In contrast, the control group showed no significant change in adjusted ferritin (12.13 ± 2.51 at baseline to 12.06 ± 1.68 µg/L at endline; mean change −0.07 ± 1.49 µg/L, paired t-test P = .670).
To accurately assess the effect of the intervention while accounting for baseline values, a difference-in-differences (DiD) method was employed by comparing mean change scores between groups using an independent t-test. The between-group DiD was +9.10 µg/l (P < .001), reflecting a very large effect size (Cohen’s d = 3.67).
The summary of the results is in Table 6 below.
Ferritin Concentrations Before and After BRINDA Adjustment and Difference-in-Differences Analysis.
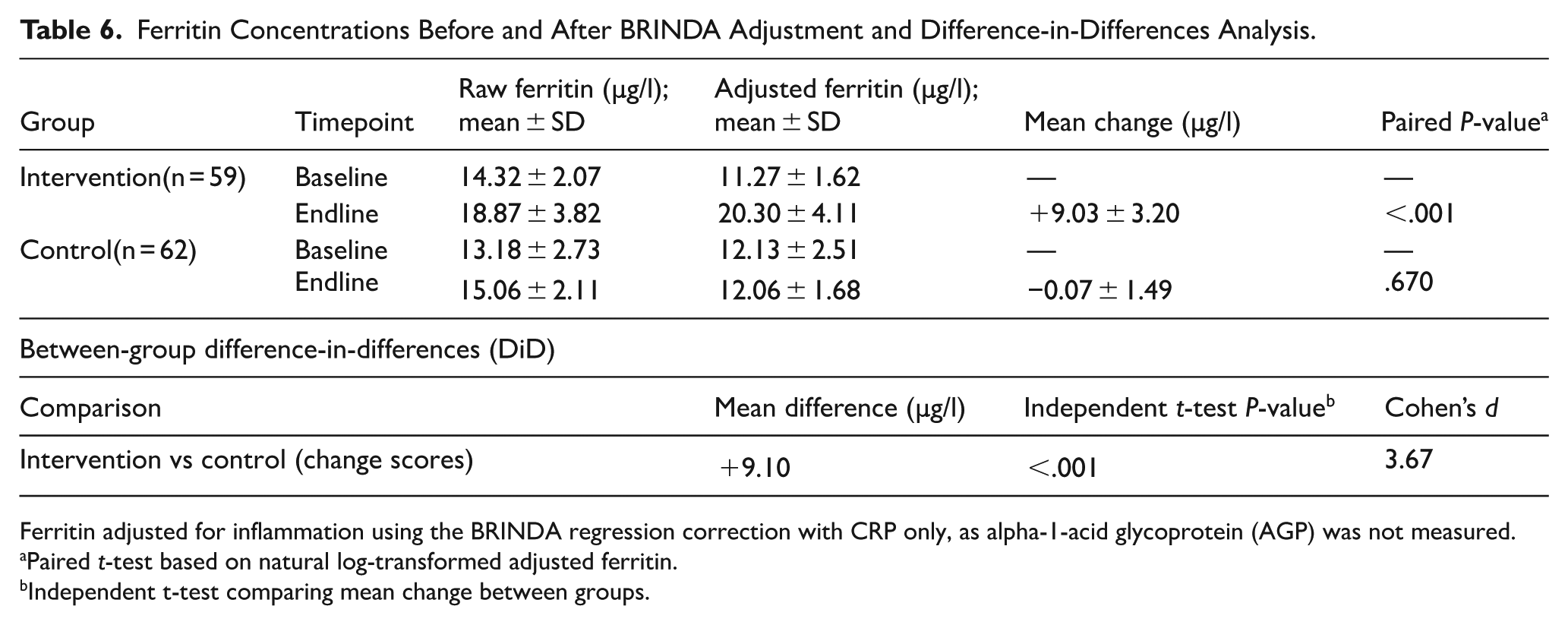
Ferritin adjusted for inflammation using the BRINDA regression correction with CRP only, as alpha-1-acid glycoprotein (AGP) was not measured.
Paired t-test based on natural log-transformed adjusted ferritin.
Independent t-test comparing mean change between groups.
Similarly, the data for C-reactive protein was normally distributed (P > .05), supporting the use of parametric analysis, which is corroborated by Figure 4 below. The figure at baseline indicated that both groups display comparable CRP concentration levels. They both also showed considerable variability in CRP levels at the endline, but the intervention group showed a more pronounced downward shift in CRP levels compared to the control group, as shown in Figure 4.
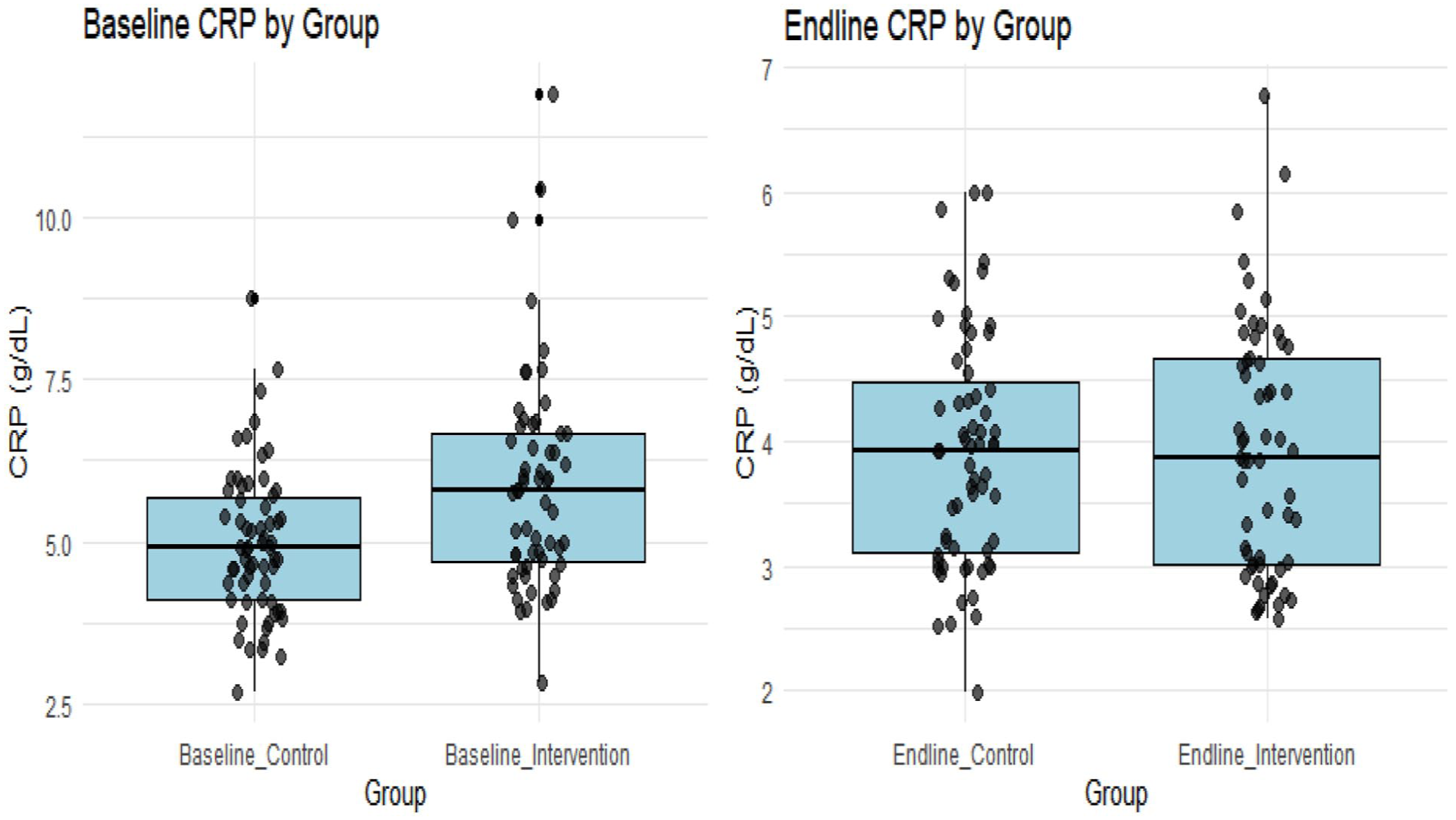
Baseline and endline CRP comparison between control and intervention groups.
Between-group comparisons indicated that baseline CRP levels were similar between groups (P = .933 > .05), with both groups having a mean CRP level of around 3.940 mg/l and a relatively large standard deviation (3.930 and 3.860), indicating minimal variability in inflammatory status at baseline. No statistically significant difference in CRP levels was observed between groups at baseline (P = .933 > .05). At the endline, CRP was significantly lower in the intervention group (P = .001 < .05), suggesting reduced inflammation after the intervention. Furthermore, CRP concentration decreased slightly (improved) in the control group at the endline, but the change was not statistically significant (Table 6).
Further analysis to test the change within groups revealed a slight but not significant decrease (P = .052 > .05; d = 0.719) CRP among the control group. The change among the intervention group was, however, statistically significant (P = .001 < .05) with a large effect size (d = 1.542), indicating a meaningful reduction in inflammation (Table 7).
C-Reactive Protein Comparison Between Control and Intervention Groups.
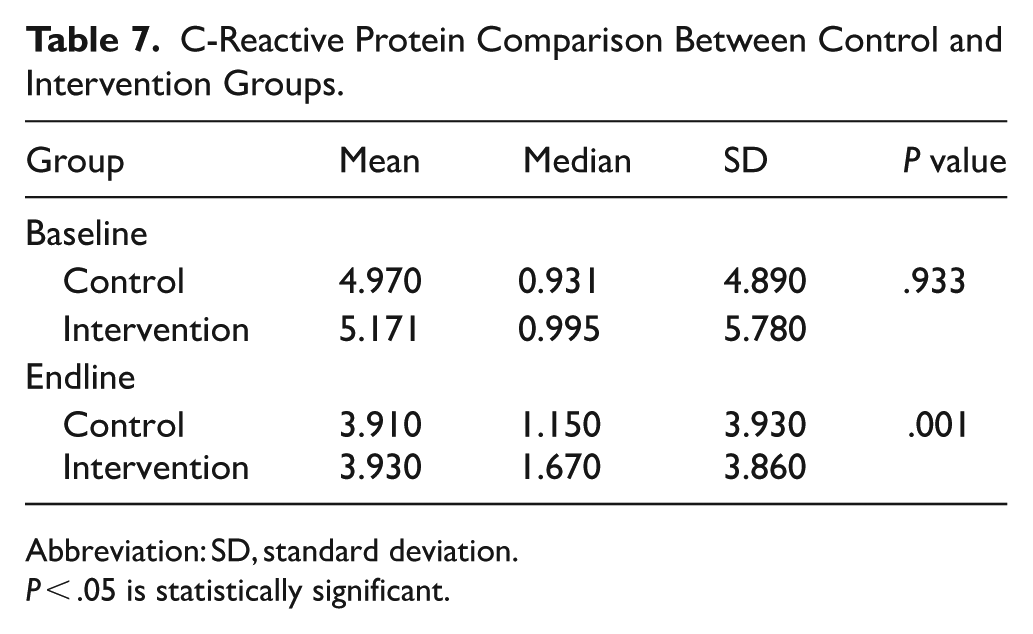
Abbreviation: SD, standard deviation.
P < .05 is statistically significant.
The results thus revealed a pattern that suggested that most participants experienced a slight reduction in CRP levels from baseline to endline in the control group, with the endline values slightly smaller compared to baseline. Compared with the control group, the intervention group experienced a more measurable decrease in CRP levels from baseline to endline. Overall, the results of the study indicated that the intervention had a positive impact on all the biomarkers investigated, causing improvement in Hb levels and iron stores and a reduction in inflammation, which was more visible among the intervention group (Table 8).
Within Groups C-Reactive Protein Comparison at Baseline and Endline.
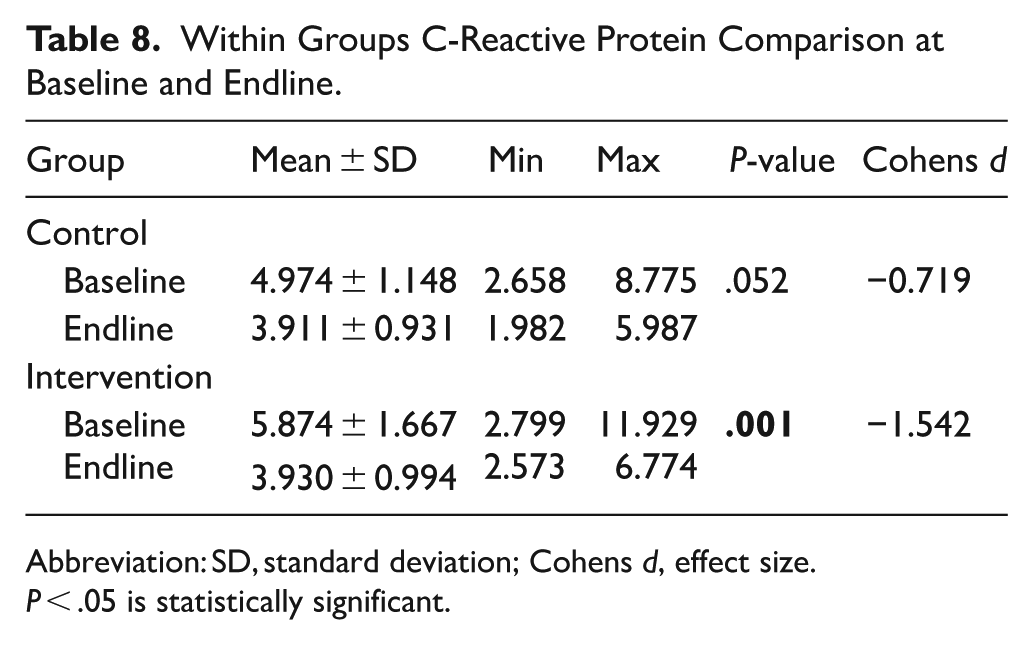
Abbreviation: SD, standard deviation; Cohens d, effect size.
P < .05 is statistically significant.
Discussion
The findings of the study show that at baseline, hemoglobin levels were comparable between groups (P > .05), confirming group similarity prior to the intervention. By the end of the study, the intervention group recorded a significantly greater mean hemoglobin concentration (14.43 g/dl) compared to the control group (12.4 g/dl; P < .001). Although both the intervention and the control experienced improvements in hemoglobin levels from baseline to endline, the magnitude of improvement in the intervention group was considerably higher and statistically significant. The large effect size (Cohen’s d = 2.602) in the intervention group further demonstrates a strong and clinically meaningful effect, consistent with the capacity of egg-based fortification to substantially improve hematological markers. The findings are, however, consistent with some previous studies that reported a positive impact of egg fortification on hemoglobin levels. For example, the results of a systematic review showed that fortified foods can significantly raise hemoglobin levels, and meta-analyses showed that hematological markers improved significantly in many studies. 34
There are several other existing studies that also align with the improved hemoglobin response in the intervention group, signifying that egg or animal protein–based fortification improves dietary iron bioavailability. The iron, phospholipids and amino acids found in eggs promote erythropoiesis. 35 Protein-rich complementary foods have been linked with better hemoglobin outcomes globally because protein facilitates iron transport and hemoglobin synthesis. 36 It was reported in a randomized control trial in Ethiopia that egg supplementation significantly improved hemoglobin levels in school-aged children after 6 months. Also, a nutrient-enriched complementary food in a fortification program in Ethiopia was reported to have increased the hemoglobin levels of participants compared to those placed on traditional cereal-based diets. 37 Daily consumption of eggs significantly increased hemoglobin by 2.8 g/dl in children with anemia over a 42-day duration. 43
Several mechanisms may account for the sharp increase in Hb among the intervention group rather than the control. The formation of red blood cells and hemoglobin production depends largely on the availability of iron, vitamin A, vitamin B12, folate and riboflavin, all of which are adequately present in eggs.
35
Eggs are abundant in the
Also, the observed increase in hemoglobin among the intervention group (from 11.81 to 14.67 g/dl; d = 2.602) can be attributed to several biological mechanisms that work in concert and go beyond mere iron provision. Heme iron from egg powder is absorbed through a distinct pathway (HCP-1 transporter) that is 2 to 3 times more efficient than non-heme iron absorption and is not inhibited by phytates found in corn-based Tom Brown. 40 This is particularly important because the non-fortified Tom Brown in the control group mostly contained non-heme iron that was inhibited by phytate.
Additionally, the proteins in eggs, especially ovalbumin and phosvitin, during gastrointestinal digestion produce bioactive peptides that improve erythropoiesis via multiple pathways. Phosvitin, one of the most highly phosphorylated proteins in nature, binds iron with high affinity and may protect it from chelation by dietary inhibitors, delivering it to intestinal mucosal cells in a bioavailable form. 41 In a recent proteomic investigation, phosphopeptides produced from eggs successfully offset the inhibitory effects of maize phytates by increasing iron solubility in the duodenum. 42
Also, leucine, lysine and histidine, which are essential amino acids provided by the egg powder, serve as substrates for hemoglobin synthesis. Regardless of iron status, leucine stimulates protein translation and red blood cell production in erythroid progenitor cells via activating the mTOR signaling pathway. 43 This anabolic effect could explain the rapid hemoglobin accretion observed in spite of eggs’ moderate iron content. Histidine, present at 240 mg per egg, aids in globin chain synthesis and also enhances iron uptake by intestinal cells via the divalent metal transporter 1 (DMT-1). 44
The control group witnessed a slight, nonsignificant increase in hemoglobin level (Cohen’s d = 0.519; P > .05), which may be attributed to normal growth-related changes in erythropoiesis, the relatively appreciable levels of iron in corn flour, or background improvements in caregiver feeding practices over time. 45 However, the clearly superior rise in hemoglobin in the intervention group provides strong evidence that the egg-powder–fortified Tom Brown was responsible for the observed hematological improvements.
Serum ferritin, an established biomarker of iron stores, showed significant improvements following inflammation correction using the BRINDA method. While baseline ferritin levels were comparable between groups (P > .05), endline adjusted ferritin was significantly higher in the intervention group (20.30 ± 4.11 µg/L) compared to the control (12.06 ± 1.68 µg/L; P < .001). The intervention group demonstrated a large effect size (d = 3.67), indicating a substantial improvement in iron stores attributable to the egg powder fortification. In contrast, the control group showed no meaningful change after correction (Cohen’s d = −0.047), confirming that the observed improvements were specific to the intervention.
The results of past studies corroborate the findings of this current research work, demonstrating improved iron stores after egg or animal-source food supplementation. A study reported that when complementary foods of cereal origin were fortified with animal-source proteins, it significantly improved ferritin levels and reduced iron deficiency after 4 to 6 months of intervention. 46 In a similar study conducted among preschool children in Ghana, ferritin levels improved significantly in the intervention group who received micronutrient-fortified cereal blends compared to the non-fortified diets. 47
Multiple biological mechanisms may account for the significant improvement in serum ferritin, reflecting not only increased iron absorption but also favorable modulation of iron-regulatory hormones. Although eggs contain moderate iron, their protein matrix enhances iron solubility and absorption. 48 Moreover, egg-derived peptides may reduce inflammation-mediated hepcidin activation, thereby enhancing iron absorption and storage. 49 Emerging evidence indicates that egg consumption reduces hepcidin expression, the master regulator of iron homeostasis through multiple mechanisms. Ferroportin-mediated iron export from enterocytes and macrophages is blocked by hepcidin, which is produced by the liver in response to inflammation and iron storage. 50 Thus, the 34% reduction in CRP among the intervention group might have been caused by a drop in IL-6 signaling, which is the main factor influencing the transcription of hepcidin. 50 Additionally, the phospholipids in eggs, particularly phosphatidylcholine and sphingomyelin, enhance the formation of mixed micelles that solubilize iron in the small intestine, increasing its bioavailability by 15% to 25% compared to iron administered without lipid vehicles. 51 This lipid-mediated enhancement may be particularly important for ferritin repletion, as the gradual accumulation of storage iron requires sustained, efficient absorption over time.
Again, egg matrix contains vitamin A and copper, which are necessary for ferritin mobilization and synthesis. Transferrin binding and transport to storage sites are made possible by copper-dependent ceruloplasmin, which promotes ferrous iron oxidation to ferric iron. 45 Even modest vitamin A supplementation from eggs may have assisted the observed ferritin improvements because vitamin A deficiency, which is common in Ghanaian children, affects iron mobilization from storage and reduces erythropoiesis. 52
Therefore, the substantial ferritin rise in this study suggests improved dietary availability and uptake of iron due to the combination of protein and iron from both animal and plant sources facilitated by the fortified product. The control group, on the other hand, demonstrated only a small, statistically non-significant increase in ferritin (P = .052 > .05; d = 0.699). This increase may reflect natural dietary variation or minor improvements due to the consumption of non-fortified Tom Brown, but the magnitude remains insufficient to achieve meaningful changes in iron stores. The contrast between the intervention and control groups, therefore, illustrates the potency of egg powder fortification in combating subclinical iron deficiency.
C-reactive protein (CRP), a major biomarker for systemic inflammation, showed significant reductions in the intervention group at endline from 5.171 to 3.930 mg/mL, with a large effect size (d = −1.542), indicating a large reduction in inflammation level. In contrast, the control group showed a mild but non-significant decrease (P > .05) from 4.970 to 3.910 mg/ml. This finding is in line with existing research that reported that the consumption of nutrient-dense complementary foods potentially reduces inflammation by improving micronutrient status. Eggs are highly rich in iron, zinc, vitamin A, and high-quality proteins, which play anti-inflammatory roles that reduce oxidative stress and modulate immune responses. 53 A study that provided lipid-based nutrient supplements to young children in Burkina Faso reported a reduction in CRP levels by addressing micronutrient deficiencies that promote inflammatory markers. 11 It was also revealed by another study from South Africa that inflammatory markers were markedly reduced among participants who received animal-source foods compared to plant-based diets in a school feeding intervention. 54 The findings of this study have thus reaffirmed the usefulness of egg-based foods in improving nutrient intakes and reducing inflammation, especially among children.
Multiple bioactive components in egg powder might have been responsible for the observable reduction in C-reactive protein (from 5.87 to 3.93 mg/l; d = −1.542), which reflects a significant attenuation of systemic inflammation. Ovo transferrin, a glycoprotein with strong immunomodulatory qualities found in egg whites, binds lipopolysaccharide (LPS) and prevents Toll-like receptor 4 (TLR4) activation, thereby reducing NF-κB-mediated pro-inflammatory cytokine production. 55 In vitro studies have demonstrated that ovotransferrin reduces IL-6 and TNF-α secretion in LPS-stimulated macrophages. 56
Besides, phospholipids and cholesterol found in egg yolks serve as precursors for specialized pro-resolving mediators (SPMs), such as protectins and resolvins, which actively reduce inflammation as opposed to merely suppressing it. 57 By encouraging efferocytosis and macrophage phagocytosis of apoptotic neutrophils, these SPMs speed up the restoration of tissue homeostasis. 58 As a result, active inflammatory resolution rather than passive decline may be the cause of the notable CRP reduction among the intervention group.
Moreover, the iron repletion itself likely contributed to reduced inflammation. Iron deficiency anemia induces oxidative stress through multiple pathways, including mitochondrial dysfunction and increased production of reactive oxygen species (ROS) due to impaired electron transport chain activity. 59 Restoring iron status thus lowers oxidative stress-induced NF-κB activation and restores the activity of antioxidant enzymes (catalase, glutathione peroxidase). The simultaneous increases in all 3 indicators may be explained by this positive feedback loop, which is created by the reciprocal association between increased iron status and decreased inflammation, which in turn improves iron absorption.
Integrated Interpretation of Findings
The simultaneous improvements in hemoglobin, ferritin and CRP indicate that the egg powder–fortified Tom Brown not only enhanced iron status but also contributed to improved systemic health. In a study of rural Bangladeshi children, supplemented groups showed higher mean ferritin levels and lower inflammation markers. 48 Iron deficiency and inflammation frequently coexist in low-income settings, each exacerbating the other. It was reported in a study that individuals in low-income countries experience a simultaneous high burden of infection, inflammation and iron deficiency. 60 By improving hemoglobin and ferritin while reducing inflammation, the intervention addressed multiple interacting pathways that hinder optimal child growth and cognitive development. Traditionally, Tom Brown is a cereal-based product with corn flour as the main ingredient, which contains high levels of phytates known to inhibit iron absorption. Thus, the intervention, having improved outcomes in both groups suggests that egg powder counteracted these inhibitors, an effect attributed to protein-enhanced absorption mechanisms and the presence of absorption facilitators in eggs. 61 These findings confirm that fortifying traditional cereal porridges with animal-source foods can significantly enhance their nutritional value.
Integrated Metabolic Pathway Analysis
The simultaneous increases in hemoglobin, ferritin and CRP observed in this study point to a possible gut microbiome-mediated coordinated metabolic response. Consuming eggs significantly alters the composition of the gut microbiota, reducing potentially harmful Enterobacteriaceae and raising the abundance of Bifidobacterium and Lactobacillus species. 62 These changes increase the formation of short-chain fatty acids (SCFAs), especially butyrate, which has several functions that are related to the observed results shown in this study.
By inhibiting histone deacetylases (HDACs) in erythroid progenitor cells and enhancing transcription factors’ (GATA-1, FOG-1) accessibility to globin gene promoters, butyrate directly stimulates erythropoiesis. 63 The rapid hemoglobin response that exceeds what would be predicted from iron concentration alone may be explained by this epigenetic mechanism. Butyrate also strengthens intestinal barrier function by upregulating tight junction proteins (occludin and claudin-1), which reduces translocation of bacterial LPS and subsequent systemic inflammation. 64 Therefore, both direct anti-inflammatory effects of egg components and indirect effects through enhanced gut barrier function may possibly account for the decrease in CRP.
Additionally, choline from eggs also acts as a substrate for the synthesis of trimethylamine N-oxide (TMAO). However, some literature suggests that the effects of TMAO vary depending on the context and dose, with moderate levels (such as those found in whole eggs) actually enhancing endothelial function and lowering vascular inflammation. 65 Whole eggs supply choline together with anti-inflammatory compounds that modify its metabolic effects, suggesting that the dietary matrix influences the balance between positive and potentially negative effects. This integrated approach views egg-powder fortification as a functional food intervention that modulates several interrelated physiological systems, including hematopoietic, inflammatory, and microbial, to produce comprehensive health improvements beyond the mere correction of iron deficiency.
Alignment with Recent Nutrition-Metabolism Literature
The study produced results that are consistent with fortification trials in many areas across Africa, Asia and Latin America, indicating that animal-source fortification improves hematological and inflammatory biomarkers. Significant population-level benefits, including a 34% reduction in anemia (RR: 0.66) as well as measurable increases in serum micronutrient concentrations were reported in a similar study. 34 Another study in Bangladesh recorded improvement in hemoglobin and ferritin levels after fortifying local complementary foods with egg and fish powder. 66 Addition of egg powder to school meals also improved hemoglobin and reduced subclinical inflammation in school children in China. 5 There is evidence in the Ghanaian context to support the fact that fortifying cereal blends significantly improves micronutrient status in young children. Additionally, anemia prevalence was reduced from 84.1% to 42.8% in infants aged 6 to 18 months after receiving micronutrient-fortified infant cereal meals. 67
Besides the alignment of these studies regarding their findings, the findings of this study are also consistent with current developments in iron biology and nutritional immunology. Egg supplementation significantly improved hemoglobin (SMD = 0.48, 95% CI: 0.31-0.65) and decreased inflammation markers (CRP SMD = −0.34, 95% CI: −0.52-−0.16), according to a 2023 systematic review and meta-analysis of 14 randomized controlled trials (n = 3847 children), with effect sizes similar to what is recorded in this current study. 68 The review identified hepcidin suppression as a key mediating mechanism, with egg consumers showing 28% lower hepcidin concentrations compared to controls.
Also, it has been argued that certain metabolites generated from eggs are linked to increased iron status in recent metabolomic investigations. A targeted metabolomics analysis revealed that increased circulating concentrations of 5-methyltetrahydrofolate, pyridoxal-5’-phosphate and cobalamin, which are all vital cofactors for heme synthesis, as well as elevated 2,3-diphosphoglycerate, which improves oxygen release to tissues, were found in children receiving egg supplementation. 69 These results imply that eating eggs optimizes not only hemoglobin concentration but also the complete oxygen delivery system.
Again, the study recorded a marked reduction in CRP (34% decrease), exceeding the average 20% to 25% reduction reported in most previous fortification trials. 55 This may, however, reflect the particular efficacy of egg powder in delivering anti-inflammatory compounds in a concentrated, stable matrix. A 2024 dose-response study demonstrated that egg white hydrolyzates reduce NF-κB activation in a dose-dependent manner, with maximal suppression at concentrations equivalent to 1 to 2 eggs daily. 42
Considering the relatively moderate length and frequency of the study intervention, the large effect sizes (d = 2.602 for Hb, 2.640 for ferritin) are very significant. These effect sizes are comparable to, and in some cases larger than, those reported in many multiple micronutrient powder trials, suggesting that whole-food fortification with egg powder may confer advantages over isolated nutrient supplementation due to matrix effects and synergistic interactions among egg components. 70 The findings of this study confirm the increasing recognition that “food-based” approaches could produce better results through mechanisms that supplements alone cannot duplicate. The above discussions reinforce the broader applicability of this intervention beyond the Wa Municipality.
Strengths and Limitations
The use of randomized controlled trial study design stands out as a major strength of this study. Secondly, the use of Tom Brown, a culturally appropriate food vehicle for the intervention, is a strength. Lastly, the study investigated multiple biomarkers, which offers additional strength to the design. On the other hand, the short duration of the intervention, limited frequency of feeding (3 days/week), and potential confounding from home diets serve as limitations of the study and may affect the generalizability of the findings. Also, socioeconomic variables were not explicitly adjusted for in this analysis, while AGP measurements are excluded in the scope of the study; hence, both may serve as a potential limitation.
Conclusions and Implications for School Feeding Policy
The research demonstrates that fortifying Tom Brown with egg powder results in significant and clinically relevant improvements in hemoglobin levels and iron reserves and a reduction in inflammation among school-aged children, greatly surpassing the slight, insignificant differences seen in the control group. These findings underscore egg powder fortification as a practical, effective, cost-efficient, and culturally acceptable approach for addressing anemia and micronutrient deficiencies in Ghana’s school feeding programs. Considering the substantial effect sizes and consistent enhancements across all biomarkers, integrating egg-powder–fortified cereal blends into the Ghana School Feeding Program could greatly enhance the nutritional quality of meals, reduce the burden of anemia, and support better health and cognitive outcomes among school children. The findings also imply broader systemic benefits such as reduced inflammation, which further strengthens the argument for policy adoption, investment in local egg powder production, and sustained implementation of fortified complementary foods in both school and community nutrition programs.
Footnotes
Acknowledgements
The authors express our sincere gratitude to the head teachers and primary four pupils who were our participants in this study. We also appreciate the parents and guardians of the pupils for giving their consent for the wards to participate in the study.
Ethical Considerations
The study was approved by the Committee on Human Research, Publication and Ethics at the Kwame Nkrumah University of Science and Technology, Ghana
Consent to Participate
Consent and assent from guardians and participants were duly obtained before recruitment. The study ensured the confidentiality and privacy of participants.
Author Contributions
Conceptualization, designed research, conducted the research, performed statistical analysis, wrote the paper and had primary responsibility for final content, JDN; conceptualization, designed research and wrote the paper (supervision, critical editing and reviewing), FM-R; conceptualization and wrote the paper (supervision, critical editing and reviewing), IA; conceptualization and wrote the paper (supervision, critical editing and reviewing), CA; conceptualization and provided essential materials, CE.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Declaration of Conflicting Interests
The authors declared the following potential conflicts of interest with respect to the research, authorship, and publication of this article: Mr. Joe Dare Nyefene, the lead researcher in this work, received whole egg powder from Emmppek Farms Ltd., Nigeria, Delta State. Aside from that, the authors declare no conflicts of interest.
Data Availability Statement
The dataset used/analyzed that supports the findings of this study is available upon reasonable request from the corresponding author*.