
Abstract
Dysfunctions of the human mirror neuron system have been postulated to underlie some deficits in autism spectrum disorders including poor imitative performance and impaired social skills. Using three reaction time experiments addressing mirror neuron system functions under simple and complex conditions, we examined 20 adult autism spectrum disorder participants and 20 healthy controls matched for age, gender and education. Participants performed simple finger-lifting movements in response to (1) biological finger and non-biological dot movement stimuli, (2) acoustic stimuli and (3) combined visual-acoustic stimuli with different contextual (compatible/incompatible) and temporal (simultaneous/asynchronous) relation. Mixed model analyses revealed slower reaction times in autism spectrum disorder. Both groups responded faster to biological compared to non-biological stimuli (Experiment 1) implying intact processing advantage for biological stimuli in autism spectrum disorder. In Experiment 3, both groups had similar ‘interference effects’ when stimuli were presented simultaneously. However, autism spectrum disorder participants had abnormally slow responses particularly when incompatible stimuli were presented consecutively. Our results suggest imitative control deficits rather than global imitative system impairments.
Introduction
Autism spectrum disorders (ASDs) are characterized by good performance on tasks requiring systemizing and the perception of detail (Baron-Cohen et al., 2009) and poor performance on tasks requiring an understanding of mental states, emotional processing and imitation (for reviews, see Vivanti and Hamilton, 2014; Williams et al., 2001, 2004). To imitate, observed actions are matched onto motor representations in the observer’s motor system (Rizzolatti and Craighero, 2004) via an action observation–execution matching (AOEM) system engaging, among other neuronal circuits, the mirror neuron system (MNS). The MNS, first described in primates, predominantly comprises the inferior frontal gyrus, the inferior parietal and superior temporal lobe (Iacoboni and Dapretto, 2006) and is closely linked to the anterior cingulated cortex and insula (Rizzolatti and Fabbri-Destro, 2008), areas that are implicated in empathy and understanding others’ emotions and intentions – some of the core problems in ASD. There is an ongoing debate whether MNS dysfunctions are relevant in the pathophysiology of ASD (Hamilton, 2013; Iacoboni and Dapretto, 2006). Results of studies addressing MNS functioning in ASD are conflicting. Some suggested global MNS dysfunction (review Iacoboni and Dapretto, 2006), whereas others suggested partial dysfunction, mainly related to the imitation of non-goal-directed actions (Hamilton, 2008), normal functioning (Bird et al., 2007) or even MNS hyperactivity, probably as a consequence of impaired imitative top-down control (Hamilton, 2008; Spengler et al., 2010). Incorporating data on behavioural, neurophysiological and neuroimaging studies examining the MNS in ASD, Hamilton (2008, 2013) suggested replacing the somewhat imprecise ‘broken mirror hypotheses’ in ASD with a more specific model of partially disturbed imitation in these patients. Thus, whereas ASD patients show normal performance and normal brain activity on meaningful ‘emulation’ tasks that involve a goal or object, that is, goal-directed imitation (for review, see Gowen, 2012), they have performance deficits predominantly in ‘imitation/mimicry’ tasks (Wild et al., 2012). The latter require spontaneous copying of low-level, kinematic features of an action that does not normally involve an object or goal, but are either meaningless gestures or facial expressions. Spontaneous mimicry of meaningless actions – also referred to as automatic imitation – appears to be lacking in ASD (Gowen, 2012; Hamilton, 2008; Wild et al., 2012).
To this end, we tested reaction times (RTs) of ASD patients and healthy controls in previously validated tasks involving the automatic imitation of simple, meaningless movements (Biermann-Ruben et al., 2008a; Jonas et al., 2010). In the main experiment, participants were asked to immediately respond to meaningless finger-lifting or dot movements with their corresponding finger. We hypothesized that in ASD, the observation of biological (finger) movements, known to lead to RT decreases in healthy controls, would not convey behavioural advantages reflecting their difficulty in mimicry tasks. In a more complex visuo-acoustic experiment, we used two different either contextually compatible or incompatible stimuli to study ‘interference effects’. These could result from co-activation of different elements within the MNS (Blakemore and Frith, 2005) and might therefore be attenuated if the MNS was organized abnormally. On the other hand, ‘interference effects’ are influenced by top-down control of imitation (Hamilton, 2008) and might therefore be more pronounced if such top-down control was reduced. Importantly, in the complex visuo-acoustic experiment, compatible and incompatible stimuli were either presented simultaneously or consecutively (Biermann-Ruben et al., 2008a). In healthy subjects, only simultaneous stimulus presentation led to significant interactions of the type of stimuli (finger vs dot) and compatibility effects indicating that such interaction crucially depends on the timing of the presentation (Biermann-Ruben et al., 2008a; Jonas et al., 2010). Abnormalities in the perception of stimulus onset synchrony/asynchrony have pre-viously been found in ASD in a flash-beep illusion paradigm (Foss-Feig et al., 2010). In addition, problems with respect to timing probably related to cerebellar abnormalities in ASD (Sears et al., 1994) may lead to abnormal ‘interference effects’ particularly with respect to the presentation mode (simultaneously or consecutively) in these patients.
Patients included in this study were thoroughly assessed clinically. RTs were related to neuropsychiatric measures to explore the relationship between behavioural data and neuropsychiatric (autistic) traits.
Methods
Participants
All clinical instruments described below were applied in both ASD participants and healthy controls. Neuro-psychiatric assessment included a Structured Clinical Interview for DSM-IV (Diagnostic and Statistical Manual of Mental Disorders, 4th ed.) Axis 1 and 2 disorders (SCID; Wittchen et al., 1997) and a standard neurological examination.
Healthy controls with manifest psychiatric diseases according to the SKID or neurologic comorbidities were excluded.
Initially, 24 patients with suspected ASD were assessed in a psychiatric interview using the ICD-10 criteria (International Statistical Classification of Diseases and Related Health Problems, 10th Revision; Dilling et al., 2008). Participants with the diagnostic categories F84.0 (criterion A (at least one criterion), B (at least six symptoms with two out of the first domain and one from each of the other two domains) and C had to be fulfilled) and F84.5 (categories A–D) were included. In nine of them, an additional third-party history by a family member or friends was included.
Of the 24 patients recruited for this study, 3 were excluded because suspected ASD was not confirmed. Another patient was excluded because he was left-handed (the experiment was designed for right-handed participants). One of the initially recruited 21 healthy controls was excluded because of present alcohol abuse.
Thus, 20 ASD patients (9 males) were included in the study. They were aged between 20 and 48 years (mean age 32.6 years ± SD 8.15 years). In addition, 8 were diagnosed with high-functioning autism (4 males) and 12 with Asperger’s syndrome (5 males). The control group consisted of 20 healthy participants matched for age (33.1 ± 7.90 years; p = 0.830), gender (9 males) and education (each group: ‘no A level’ (n = 9), ‘A level’ (n = 2) and ‘university’ (n = 9)). IQ values did not differ between ASD (mean = 109.5 ± 14.28 ranging from 92 to 130) and control participants (mean = 106.2 ± 13.41 (92–136); U = 226.00, z = 0.706, p = 0.480). All participants were right-handed (except one ambidextrous) according to the Edinburgh Handedness Inventory (Oldfield, 1971). Of the ASD participants, 11 had comorbid psychiatric disorders at the time of the study (2 each had depression, OCD, ADHD, combined ADHD and OCD, combined ADHD, dysthymia and OCD, and 1 had ADHD and dysthymia). Two ASD participants with a history of chronic cannabis abuse were abstinent for at least 1 year prior to the study. Hearing was intact in every participant. No control but six ASD participants took psychiatric medication (two SSRI, one SSNRI, one opipramol, one promethazine, one fluvoxamine, pregabalin and methylphenidate).
All participants gave their written informed consent in accordance with the Declaration of Helsinki (1964). The study was approved by the Ethics Committee of the Medical Council of Hamburg (PV3506).
Neuropsychiatric tests
The German Multiple Choice Word Test (MWT-B; Lehrl, 2005) was used to calculate individual verbal IQ levels and the supermarket fluency task (Mattis, 1988) to evaluate semantic verbal fluency that is said to be impaired in ASD and to be associated with low processing speed (Spek et al., 2009). It might be associated with low processing speed in ASD in the present experiment because both processes likely engage the ventral premotor cortex. The Autism Quotient (AQ; Baron-Cohen et al., 2006), Empathy Quotient (EQ, Baron-Cohen and Wheelwright, 2004), Systemizing Quotient (SQ; Baron-Cohen et al., 2003) and the Reading the mind in the eyes test (Baron-Cohen et al., 2001) were used to quantify autistic trait severity. Each participant completed questionnaires assessing comorbidities potentially interfering with experimental results, for example, depression (Beck Depression Inventory (BDI); Beck et al., 1961), attention deficit hyperactivity disorder (ADHD; German self-assessment scale (ADHS-SB); Rosler et al., 2004) and obsessive–compulsive disorder (OCD; Yale–Brown Obsessive–Compulsive Scale (Y-BOCS); Goodman et al., 1989). Although no participant had a diagnosis of a tic disorder prior to the study, tics, a common comorbidity in ASD (Canitano and Vivanti, 2007) that could influence task performance (Jonas et al., 2010), were assessed using the modified Rush video protocol (Goetz et al., 1999). They were distinguished from stereotypes using criteria mentioned by Cath et al. (2011). The total Rush score and the Yale Global Tic Severity Scale (YGTSS; Leckman et al., 1989) were calculated. All clinical parameters were correlated with behavioural data.
Experimental set-up
Stimuli and set-up were presented with Presentation software version 14.1 (http://www.neurobs.com/). Participants were seated in front of a PC screen at a distance of 70 cm in a dimmed, acoustically shielded room and performed three behavioural experiments lasting a total of ≈35 min (Biermann-Ruben et al., 2008a; Jonas et al., 2010). Before each experiment, they completed a short training phase. Participants observed video clips showing movement sequences performed by either the index or the little finger of a left male hand (front view). Movements of the fingers are referred to as the ‘biological (socially biased) stimulus’. Additionally, red dots were attached to both index and little fingertips of the video-recorded hand. Dot movements are referred to as the ‘non-biological stimuli’ as they are not socially biased. Dot movements had an identical kinematic profile as finger movements to ensure comparability of stimuli used.
Participants were asked to constantly fixate a white cross located centrally arranged in equal distance from both fingertips. They were instructed to imitate the presented movement sequence as fast as possible using the corresponding fingers of their own right hand (placed in photoelectric barriers recording RTs), thereby creating a ‘mirror effect’ which typically increases MNS activation (Koski et al., 2003). The stimulus setting comprised movements of two different fingers in order to prevent expectancy effects.
Experiments
Experiment 1
Participants were instructed to imitate the following three different types of movement (TM) stimuli by lifting their corresponding index or little finger (Figure 1(a); Jonas et al., 2010):
(a) Finger = an isolated movement (lifting and lowering) of the presented index or little finger was shown = ‘biological’ stimulus;
(b) Dot = isolated movement (lifting and lowering) of presented red dot on either the index or littler fingertip = ‘non-biological’ stimulus (fingers do not move in this condition);
(c) Finger and dot = finger and dot are moving together.
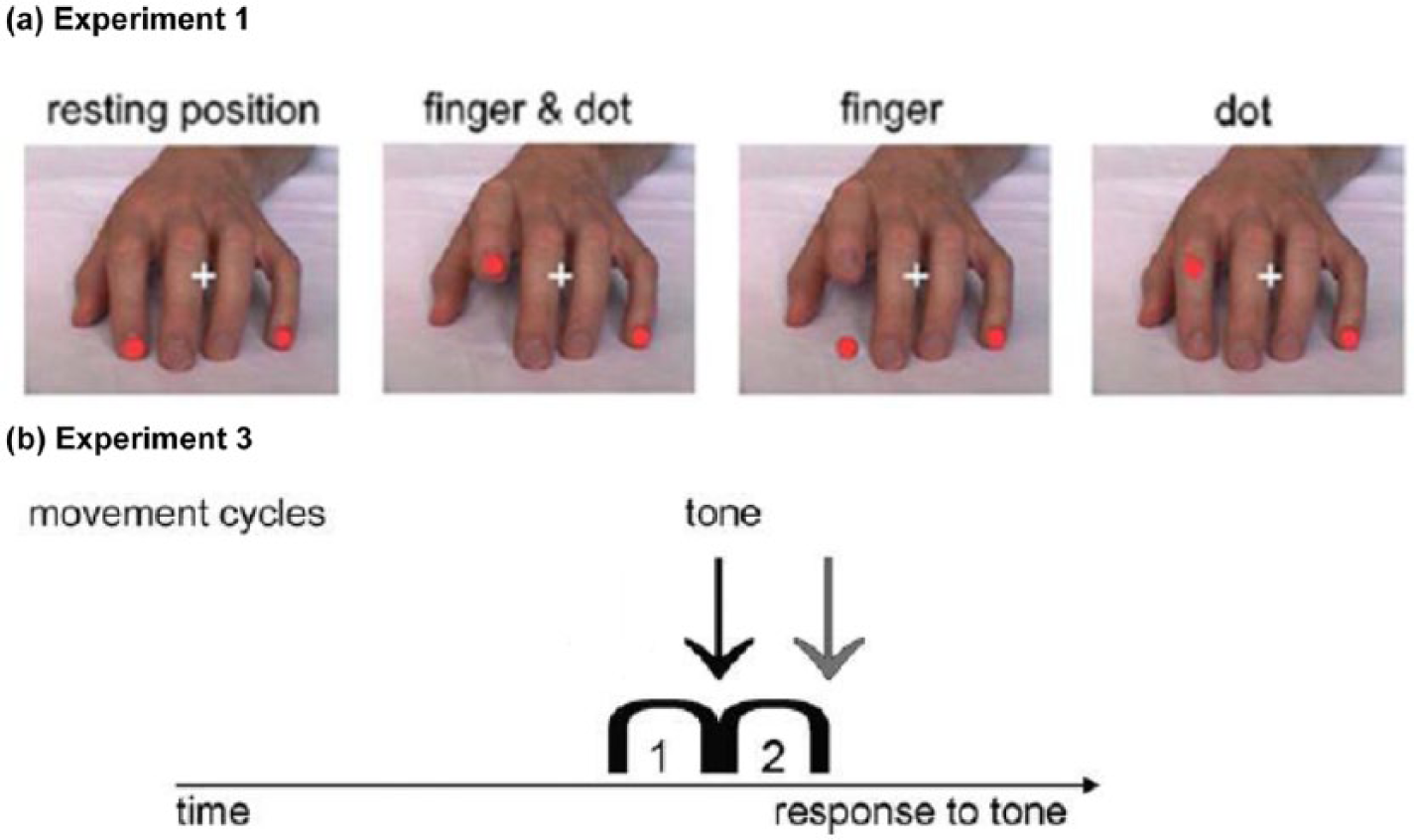
Visual cues in (a) Experiment 1 and combined cues in (b) Experiment 3. (a) Different types of movement at their maximum movement amplitude compared to the resting position (left) and (b) one arc represents one visual movement (lifting-lowering) stimulus. The arrows symbolize the onset of the tone in relation to the movement stimulus (black arrow: onset; grey arrow: offset).
Each type of movement (composed of 12 single pictures) lasted about 0.4 s and was presented randomly 24 times (= total of 72 trials). Before and after the movement, a static hand was presented (1.5 s). Inter-trial intervals (= black screen) lasted 2 s.
Experiment 2
This was a simple acoustic RT task (40 trials, each lasting 3.5 s; inter-trial interval = 2 s) where participants fixated the motionless, static hand and were asked to respond to two tones (500 or 1000 Hz) as quickly as possible. They were randomly assigned to respond to the low tone by lifting their index and to the high tone by lifting their little finger or vice versa. In four catch trials, only the static hand but no tone was given to ensure persistent attention to the task. This task always preceded Experiment 3 and served as a preparatory training for the acoustic part of the experiment.
Experiment 3
The third paradigm was used as previously described (Biermann-Ruben et al., 2008a; Jonas et al., 2010) and combined visual and acoustic stimuli. Participants viewed stimuli showing either two consecutive ‘finger and dot movements’ or two ‘dot movements’ (‘type of movement’: TM) and were instructed to watch the movements closely but only to react in response to a tone (see Experiment 2), but not to the movement on the screen. Movements displayed on the screen could be either compatible (different stimuli; same action) or incompatible (different stimuli; different action) with the participants’ ‘reactive’ movement instructed by the tone (Figure 1(b)). We subsequently refer to this aspect as ‘compatibility’ (Comp). Tones were presented either at the beginning (onset presentation, that is, simultaneously) or after completion of the second movement (offset presentation, that is, consecutively), which is referred to as ‘presentation mode’ (PM). This experiment comprised five blocks with 160 main and 40 catch trials (static hand without movement sequence or tone). Finger lifting and lowering (13 pictures, each with a duration of 32 ms) was preceded and followed by presentation of the static hand (1.5 s) and an inter-trial interval. For all three experiments, RT and error rates were recorded.
As previous studies showed that ‘finger’ and ‘finger and dot’ movements lead to similar RT in healthy subjects (Jonas et al., 2010), the ‘finger and dot’ movements in Experiment 3 are – for simplicity – equated with ‘finger’ movements to point to the difference between biological and non-biological movements.
Statistical analysis
Analyses were performed with IBM® SPSS® Statistics 20 (www.ibm.com/software/de/analytics/spss/products/statistics/).
RT represents the duration between stimuli onset and the subject’s lifting response. Results of index and little finger were averaged. RT outliers (RT< and >2.5 SD and RT <100 ms), omissions and false responses were excluded. Group comparisons of neuropsychiatric tests were analysed using independent t-tests or non-parametric Mann–Whitney tests. As we matched ASD-control couples for age, gender and education, we introduced the variable ‘pairs’ in our linear mixed model analyses (1:1 matching, both ASD and control subjects got the identical ‘pairs’ number). Fixed and random effects are given separately for each experiment below. Random effects were constructed using quantile–quantile (QQ) plots of the residuals. Including factors explaining variance heterogeneity as random effects, normal distribution of residuals was established. Competing models were judged using the Akaike information criterion. A diagonal covariance structure suitable for repeated measures was applied. In the final model, restricted maximum likelihood and Type III tests were used to estimate parameters. Finally, paired comparisons of estimated marginal means were conducted.
For error rates measured as counts, we generally used negative binomial regressions models. Because we expected controls to make fewer errors, we also used zero-inflated models which are available in the R package pscl (http://www.r-project.org/; Zeileis et al., 2008). Model selection was based on likelihood ratio tests (nested models) or Vuong’s test for non-nested models (see also Karazsia and Van Dulmen, 2008). The Spearman coefficient was used to correlate RTs, error rates and clinical parameters.
Results
Neuropsychiatric assessment
As expected, ASD participants scored higher in autism questionnaires and comorbidity scores for depression, ADHD and OCD and showed poorer performance in the reading mind and supermarket task. Tics occurred more often in ASD participants. Details of neuropsychiatric assessment are given in Table 1.
Neuropsychiatric assessment.
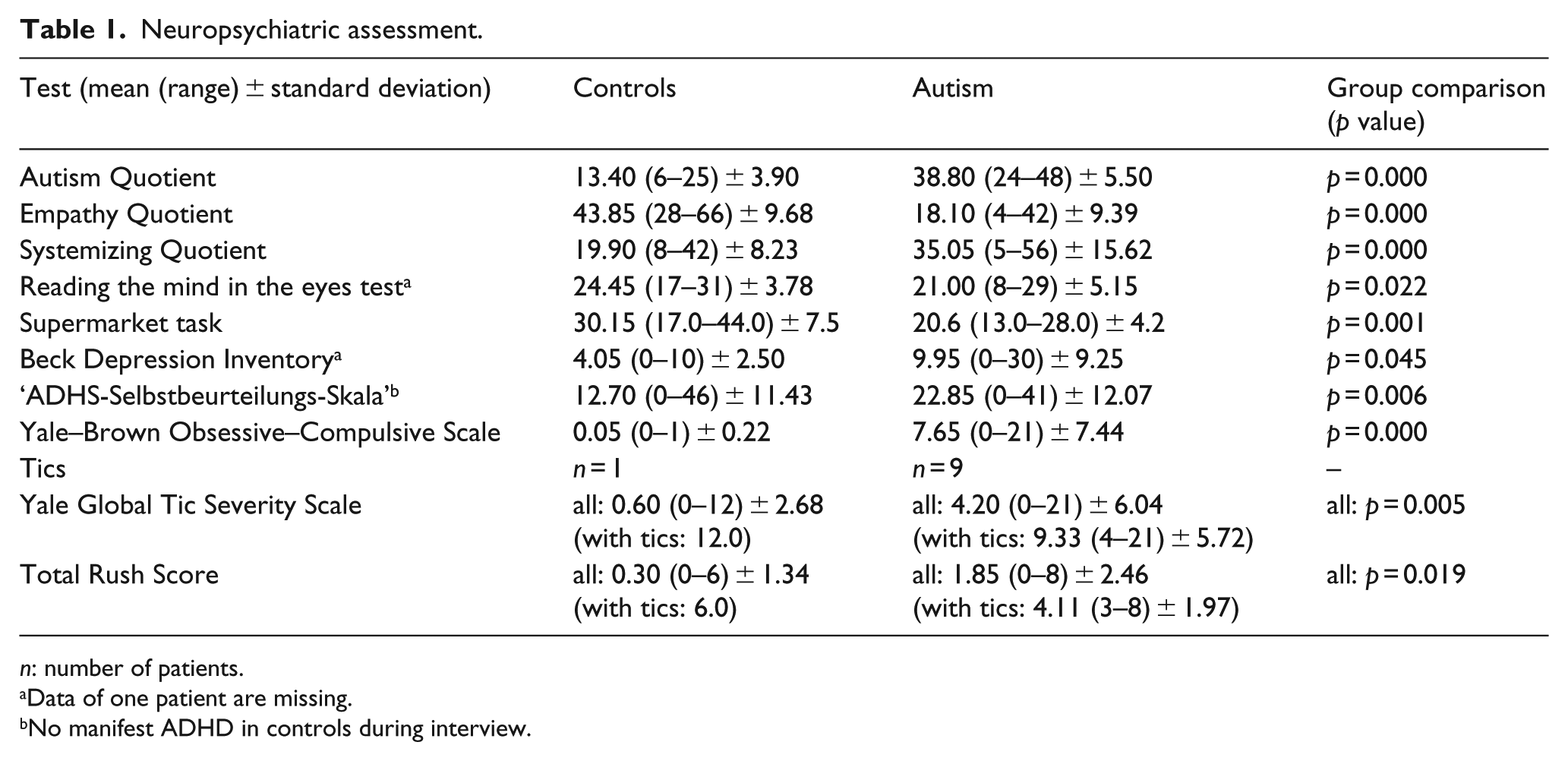
n: number of patients.
Data of one patient are missing.
No manifest ADHD in controls during interview.
Experiment 1
A mixed model analysis ‘RTs’ by ‘TM’ (finger vs dot vs finger and dot) and ‘group’ (controls vs ASD) was performed. Fixed effects were ‘group’, ‘TM’ and their interaction. ‘Group’ in conjunction with ‘pairs’ was also used as random effect to retain the matched ASD-control structure. Below, the estimated means of RTs and their confidence intervals (CI) are given. There was no interaction, but a significant main effect of ‘group’ (p = 0.006) with paired comparisons showing faster total mean RTs in controls (391 ms) compared to ASD participants (518 ms; difference = 127 ms (38–215 ms)). There was an effect of ‘TM’ (p < 0.001) with slower RTs to dot (476 ms) as compared to finger (443 ms; difference = 33 ms (25–42 ms); p < 0.001) and finger and dot movement stimuli (445 ms; difference = 32 ms (20–42 ms); p < 0.001). RTs to finger or combined finger and dot stimuli were similar (p = 0.599). This ‘biological advantage’ applied to both groups (Table 2, Figure 2).
Error rates and reaction times in Experiment 1.
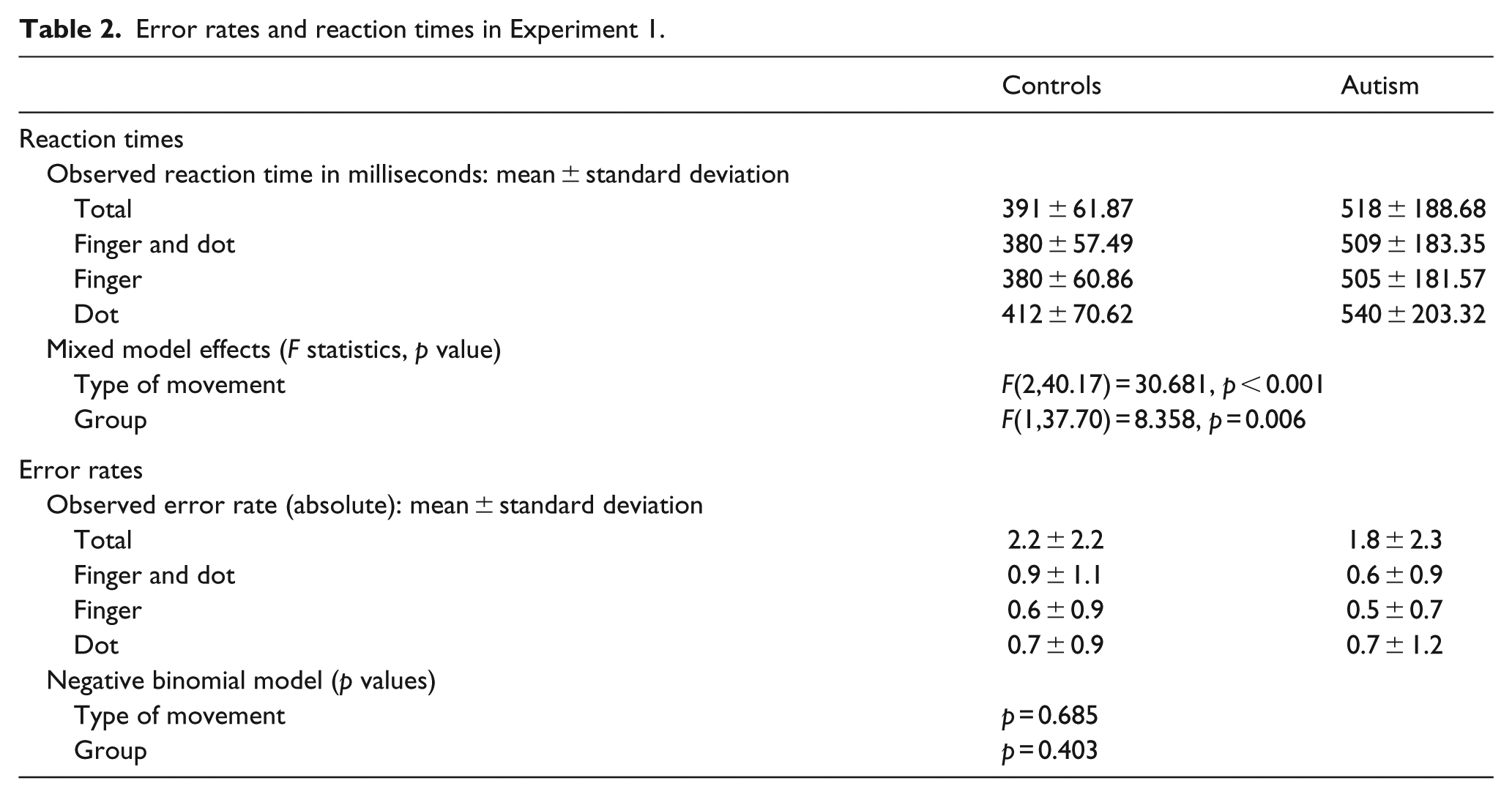
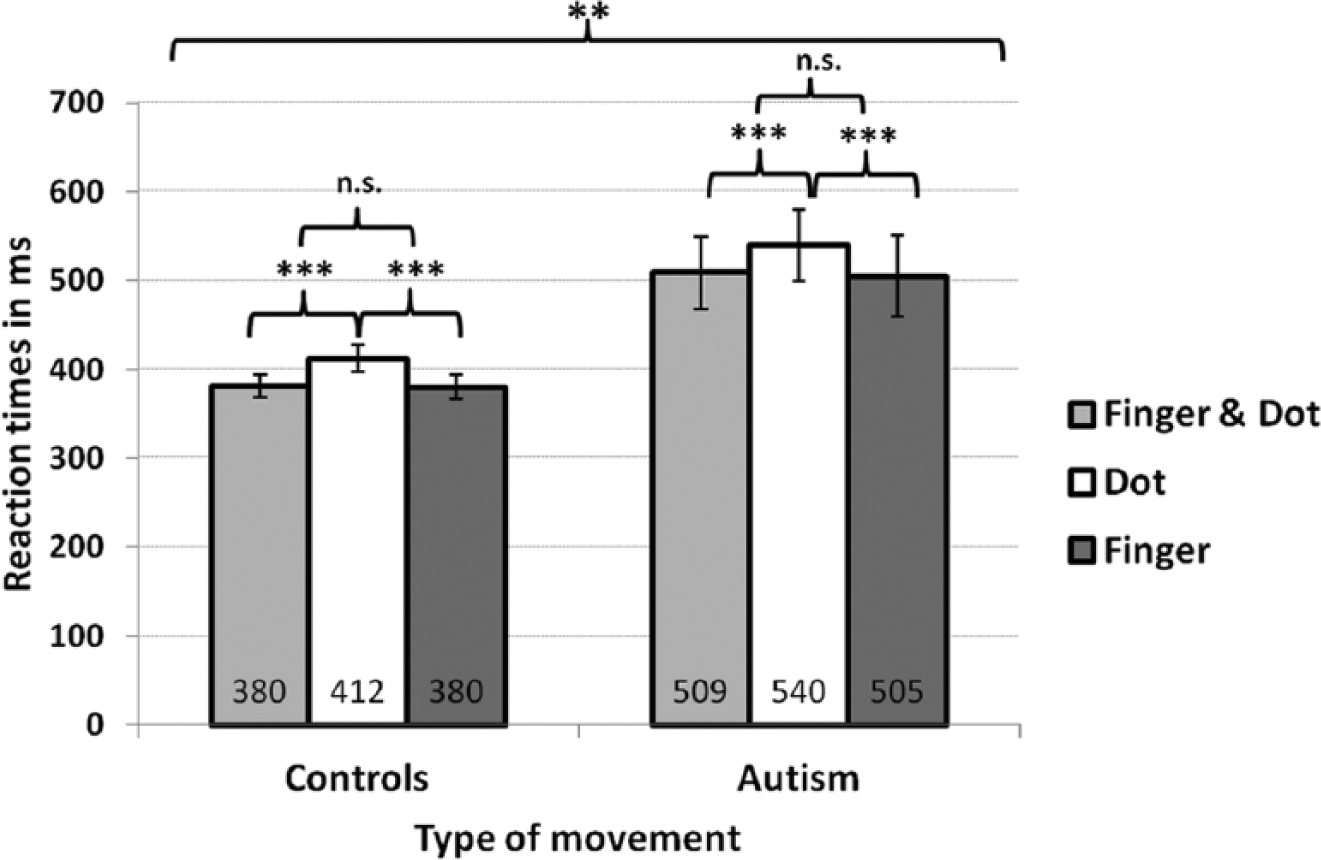
Reaction times in Experiment 1.
A similar mixed model analysis including comorbidity test scores of ADHS-SB, BDI and Y-BOCS as covariates did not reveal any significant effects. Thus, comorbidities were omitted in subsequent analyses.
A negative binomial regression model showed no differences in error rates (see Supplement 1) for ‘group’ (p = 0.40) or ‘TM’ (p = 0.69). RTs and error rates did not correlate.
No group differences were found concerning the learning effect in the preceding training phase. In Experiment 1 only, RT variance in ASD was greater and scatter plots revealed seven ASD participants with aberrant high RTs. We found no pattern in epidemiologic or neuropsychiatric tests mentioned above characterizing this subgroup (see Supplement 2).
Experiment 2
A mixed model analysis ‘RTs’ by ‘group’ was performed with ‘group’ as fixed and in conjunction with ‘pairs’ also as random effect. Although mean RTs differed numerically between controls (421 ms (373–469 ms)) and ASD participants (475 ms (427–523 ms), difference =54 ms), there was no significant effect of ‘group’ (p = 0.112). For error counts, we did not find group differences (controls: 2.2 errors/ASD: 2.8; p = 0.493; see Supplement 3). RTs and error rates did not correlate.
Experiment 3
We set up a linear mixed model for RTs using ‘PM’ (onset vs offset), ‘Comp’ (compatible vs incompatible), ‘TM’ (finger vs dot) and ‘group’ (controls vs ASD) as fixed effects. The interaction of ‘Comp’ and ‘group’ was identified as the main explanation for variance heterogeneity and was included as random effect resulting in normal distribution of residuals. For ‘PM’, ‘Comp’ and ‘TM’, repeated measures were conducted.
Analysis of RTs revealed no significant fourfold interaction. The interaction ‘PM*Comp*TM’ was significant (F(1, 210.38) = 5.185, p = 0.024) indicating that RTs in both groups vary in response to finger and dot movement stimuli as a function of stimulus compatibility and temporal presentation. Paired comparisons of estimated means and their CI revealed the following: there was an overall ‘interference effect’ (= faster RTs in compatible compared to incompatible trials) for finger and dot movement stimuli in the onset (finger: difference = 59 ms (28–90 ms), p < 0.001/dot: difference = 29 ms (2–60 ms), p = 0.066) and offset mode (finger: difference = 37 ms (4–70 ms), p = 0.030/dot: difference = 40 ms (7–72 ms), p = 0.018). Onset (513 ms) outpaced offset stimuli (531 ms) significantly only in the compatible finger movement condition (difference = 18 ms (3–32 ms), p = 0.020). This ‘simultaneity effect’ was absent in incompatible or dot conditions. For the onset mode, there was a ‘biological advantage’ with faster RTs to compatible finger (513 ms) compared to compatible dot (531 ms) movement stimuli (difference = 18 ms (4–31 ms), p = 0.014). In contrast, the onset incompatible finger (572 ms) condition led to slower RTs as compared to the dot condition (560 ms, difference = 12 ms (1–24 ms), p = 0.035). The offset mode did not show any significant differences.
Focusing on group differences, we found that the three-way interaction ‘PM*Comp*group’ was significant (F(1, 211.41) = 8.115, p = 0.005) indicating that groups differed with respect to compatible and incompatible stimuli in the on- and offset condition. In paired comparisons of estimated means, ASD participants had slower RTs in all experimental conditions compared to controls (all p values ⩽ 0.020, Figure 3(a)). Controls showed a ‘simultaneity effect’ with faster RTs to compatible onset (477 ms) compared to compatible offset (495 ms) stimuli (difference = 17 ms (3–32 ms), p = 0.022). This was not true for incompatible stimuli or the ASD group. When responses (to either finger or dot movements) were combined, ASD participants revealed a marginally significant ‘interference effect’ in the onset (difference incompatible–compatible = 39 ms (4–81 ms), p = 0.072) and a significant ‘interference effect’ in the offset condition (difference = 53 ms (10–97 ms), p = 0.017). By contrast, controls showed this ‘interference effect’ only in the onset mode (difference = 49 ms (7–92 ms), p = 0.022).
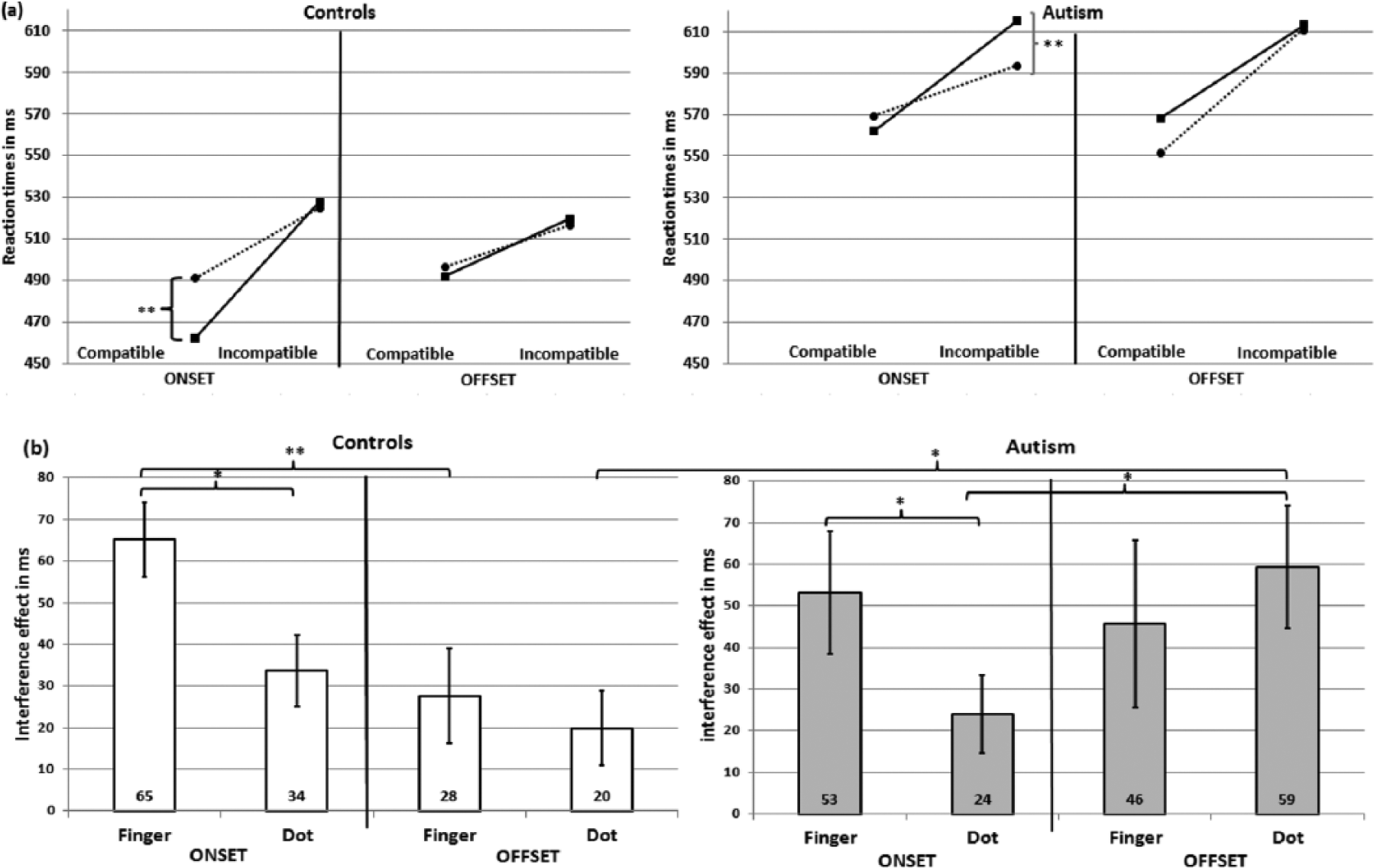
Mean reaction times in Experiment 3: (a) reaction times in the eight experimental conditions for both groups (controls left, autism right). Presentation mode (onset/offset), compatibility (compatible/incompatible) and type of movement (finger (F; solid line)/dot (D, dashed line)) are presented separately and (b) interference effects (difference incompatible–compatible trials) in response to finger and dot movement conditions are given in the onset and offset conditions for both the control (left) and autism group (right).
To further investigate this ‘interference effect’, we analysed its magnitude (= difference between incompatible and compatible trials) for both finger and dot movement stimuli in the onset and the offset modes (Figure 3(b)) using Wilcoxon tests for inner group and Mann–Whitney tests for between-group comparisons. In the onset mode, both groups revealed a greater ‘interference effect’ for biological finger movement stimuli compared to dot movement stimuli (controls: z = −2.389, p = 0.017/ASD: z = −2.091, p = 0.037), whereas in the offset mode the ‘interference effect’ of finger and dot was similar (controls: z = −0.709, p = 0.478/ASD: z = 1.008, p = 0.313). Controls, but not ASD participants, showed a greater ‘interference effect’ for the finger when stimulus presentation was simultaneous as compared to consecutive presentation (controls: z = −3.099, p = 0.002, ASD: z = −0.709, p = 0.478). In contrast, ASD participants had ‘interference effects’ in both conditions. In contrast to controls (z = −1.307, p = 0.191), ASD participants even showed a greater ‘interference effect’ for the dot when stimulus presentation was consecutive as compared to simultaneous (z = 2.165, p = 0.030). This indicates that in ASD, only consecutive stimulus presentation interfered with both biological and non-biological conditions. Comparing both groups, only the ‘interference effect’ during the offset dot condition differed significantly being larger in the ASD group (U = 277.00, z = −2.083, p = 0.037).
There was no significant interaction including ‘TM’ and ‘group’ indicating a similar processing of biological and non-biological stimuli in both groups. Nevertheless, we exploratory used paired comparisons of estimated means for each of the performed eight conditions to investigate minor differences between RTs to finger as compared to dot movement stimuli (Figure 3(a)). Whereas controls benefited from the presentation of finger as compared to dot movement stimuli in the compatible onset condition (difference = 26 ms (11–41 ms), p = 0.002), ASD participants showed a reversed pattern in the incompatible offset condition in which RTs to dot movement stimuli were shorter than those to finger movement stimuli (difference = 21 ms (7–35 ms), p = 0.003).
Using a negative binominal model, we found error rate differences between both groups (p < 0.001) in general with higher error counts in ASD (mean = 1.2 (1.0–1.5)) compared to controls (0.5 (0.4–0.7)), but no effect of ‘PM’, ‘Comp’, ‘TM’ or their interactions (see Supplement 4). RTs and error rates were not correlated.
Correlations
Correlations between RT and clinical parameters were performed mainly to explore potential relationships between behavioural results and severity of symptoms. All significant correlations are given in Table 3. Most importantly, total mean RTs in ASD in all three experiments correlated negatively with the supermarket task indicating that ASD participants with faster RTs perform better in the supermarket task.
Significant correlations (Spearman’s coefficient rs and p values) of reaction times per experiment (upper part) and interference effects (= difference of incompatible–compatible trials) in the onset and offset conditions (lower part) with clinical parameters.

Data of one patient are missing.
Discussion
This study aimed at contributing to the frequently disputed hypothesis of a MNS dysfunction in ASD by using three behavioural experiments addressing its functioning. The main findings of the present series of experiments are (1) that, contrary to our initial hypotheses, behavioural advantages conveyed by the observation of biological (finger) movements attributable to the human MNS did not differ between groups arguing against a general MNS dysfunction in ASD and (2) that interference of compatibility effects and type of stimuli depended on the presentation mode (onset vs offset) in healthy controls but less so in ASD. The latter suggests impaired control of imitation, particularly with respect to the timing of presented stimuli, in these patients.
Response patterns in Experiment 1 were identical in both groups with faster RTs in response to biological finger compared to non-biological dot movement stimuli (see also Brass et al., 2000) indicating that the processing of biological stimuli, mainly mediated by temporo-occipital, superior temporal and ventral motor/premotor areas (Biermann-Ruben et al., 2008b) as parts of the human MNS is not impaired in ASD. This lends support to the notion that there is no general dysfunction of the MNS in ASD (Hamilton, 2009, 2013). Moreover, these results also show that even spontaneous copying of low-level, kinematic features of an action that are not goal-directed, that is, automatic mimicry, is not necessarily abnormal in ASD, at least in an experimental context we used.
Similarly, in the complex, visuo-acoustical set-up (Experiment 3) with temporally (simultaneous/consecutive) and contextually (compatible/incompatible) varying stimuli, there were no significant interactions including the factor ‘TM’ indicating that biological and non-biological movements are generally processed similarly in both groups. In accordance with previous studies, both healthy controls (Brass et al., 2000) and ASD participants (Bird et al., 2007; Spengler et al., 2010) showed an ‘interference effect’ (= faster RTs to compatible compared to incompatible trials). Responses to the imperative tone were facilitated by simultaneous presentation of compatible visual stimuli further supporting the main finding that automatic imitation of meaningless stimuli is normal in ASD.
During simultaneous stimulus presentation (onset), the ‘interference effect’ was larger in response to biological stimuli in both groups. In the offset condition, there was no difference between the ‘interference effects’ in response to finger and dot movement conditions in either group. However, here, strong ‘interference effects’ in response to both finger and dot movement stimuli were present in ASD but not in controls. Thus, only ASD participants are still affected by the visual stimulus preceding the imperative tone. Intriguingly, in ASD, both the biological finger and the non-biological dot movement resulted in a large ‘interference effect’ in the offset mode. The latter was even larger in the offset as compared to the onset mode. The visual input preceding the imperative tone seems to affect responses in ASD regardless of the type of movement. Thus, perhaps not only the inhibition of imitative behaviour but also the inhibition of other acquired sensory-motor reactions (e.g. reaction to dot) is altered in ASD.
Our results further indicate that healthy controls are more capable to discriminate two consecutively applied stimuli, whereas ASD participants still perceive them as bound together. Recent findings suggested that the temporal window in which visual-acoustical stimuli are perceived as bound might be larger in ASD (Foss-Feig et al., 2010). Such problems of temporal discrimination might be related to cerebellar pathology and dysfunction in ASD (Sears et al., 1994).
Interestingly, in the onset mode, finger movement stimuli led to behavioural advantages in compatible trials in healthy controls but caused slowing of RT in incompatible trials in ASD. This response pattern is reminiscent of that previously described in Gilles de la Tourette syndrome patients that are characterized by the presence of vocal and motor tics and echophenomena (Ganos et al., 2012; Jonas et al., 2010). In these patients, it has been argued that motor-related areas responsive to action observation (and echophenomena) have a higher ‘tonic’ activation, which is compensated for by increased inhibition. Down-regulation of overactivity might be particularly important to suppress echophenomena in potentially ‘echogenic’ situations as in Experiment 3, in which biological movements are not to be imitated but nevertheless can trigger involuntary imitation. Such responses (in compatible trials) may be particularly prone to inhibition in Tourette patients reducing behavioural advantages of biological stimuli in compatible trials. On the other hand, overactivity of an action observation matching system in Tourette patients may likewise lead to stronger (uninhibited) ‘interference effects’ in incompatible trials with biological stimuli.
Imitative response tendencies including echolalia/echopraxia also appear to be increased in ASD, which might be caused by an insufficient top-down control of imitative behaviour (Hamilton, 2008; Spengler et al., 2010). This imitative control deficit might lead to stronger interference effects in ASD as the tendency to respond to biological stimuli might then predominate the participants’ capacity to inhibit their motor actions. Alternatively, adaptation to a deficit of imitation related to a developmental delay in children with ASD could lead to exaggerated imitative response tendencies in adults with ASD (Biscaldi et al., 2013).
In this study, slow RTs in ASD did not correlate with error rates, that is, they cannot be explained by a speed-accuracy trade-off, which might be related to the fact that the task per se was not very demanding.
RTs were slower, albeit non-significantly, in ASD in Experiment 2 consistent with a previous study in ASD children (James and Barry, 1983) suggesting a global impairment of sensorimotor processing. Responses of ASD patients were significantly slower though in Experi-ments 1 and 3. Responses involving visuo-motor processing might be more impaired than those mediated by other, for example, audio-motor systems. Indeed, there are studies demonstrating deficits in the use of observed motor information to plan one’s own motor actions in ASD (review Becchio and Castiello, 2012). Also, in keeping with previous work, slower motor performance is a possible explanation (Biscaldi et al., 2013). The poorer performance in the supermarket task as an indicator for slow verbal fluency which has been found to be associated with slow processing speed (Spek et al., 2009) and its negative correlations with RTs in all three experiments could also point in the same direction. In addition, overall slow RTs of ASD participants in both visuo-motor experiments might result from ‘behavioural confusion’ induced by the counterintuitive, mirrored presentation of visual stimuli (observing left while responding with right hand; Avikainen et al., 2003) although in healthy controls this ‘mirror effect’ increases MNS activation (Koski et al., 2003). Finally, attentional deficits have to be considered as a confounder. Attention to both social and non-social stimuli is impaired in ASD (Anderson et al., 2006; Rice et al., 2012). Thus, it is conceivable that reduced attention to stimuli (measured, for example, by fixation duration) could lead to decreased brain activation and increased RTs.
Our study has limitations. Selecting a group of high-functioning adult ASD participants assured limited influence of potential confounders on behavioural data. On the other hand, results cannot be generalized to the larger population of ASD patients with cognitive impairment. We did not specifically assess visual abilities. There was no evidence of visual impairment in any participant but because the task relied on vision, it is important to note that no vision assessments were completed. Some ASD participants were taking psychoactive medications; the potential impact of this cannot be completely discounted, but we feel influence of medication is limited. The number of patients was relatively small. However, behavioural data appear to be robust and not minimal. Although ASD prevalence is four to five times higher in boys (CDC, 2014), in our study the number of male and female participants was nearly equal. This can be explained by a higher number of female volunteers and by the fact that two of the four participants who had to be excluded were male. As our participants were well selected and gender comparisons in our study did not reveal any differences, we think it is unlikely that the sex distribution has biased our results. Finally, future studies should also include children and adolescents with ASD because behavioural abnormalities described here might be age dependent.
To conclude, given normal advantages conveyed by biological stimuli and physiological ‘interference effects’ during simultaneous stimulus presentation in ASD, our study suggests that their AOEM system is not globally dysfunctional. In contrast, data presented here suggest that processing of successive visual and acoustic stimuli is impaired in ASD, which may be related to reduced top-down control causing hyperimitative behaviour and/or cerebellar pathology resulting in timing problems.
Footnotes
Declaration of conflicting interests
The authors declare that there is no conflict of interest.
Funding
This work was supported by the Else Kröner-Fresenius-Stiftung [grant 2011_A37] and by the European Union [EU grant FP7-ICT-270212 and H2020-641321].