
Abstract
Autism spectrum disorder is a neurodevelopmental disorder that is associated with problems in empathy. Recent research suggests that impaired control over self–other overlap based on motor representations in individuals with autism spectrum disorder might underlie these difficulties. In order to investigate the relationship of self–other distinction and empathy for pain in high-functioning autism and matched controls, we manipulated self–other distinction by using a paradigm in which participants are either imitated or not by a hand on a computer screen. A strong pain stimulus is then inflicted on the observed hand. Behavioral and physiological results in this study showed that overall affective responses while watching pain movies were the same in adults with high-functioning autism as in controls. Furthermore, controls showed higher affective responding after being imitated during the whole experiment, replicating previous studies. Adults with high-functioning autism, however, showed increased empathic responses over time after being imitated. Further exploratory analyses suggested that while affective responding was initially lower after being imitated compared to not being imitated, affective responding in the latter part of the experiment was higher after being imitated. These results shed new light on empathic abilities in high-functioning autism and on the role of control over self–other representational sharing.
Keywords
Introduction
Autism spectrum disorder (ASD) is a pervasive neurodevelopmental disorder characterized by abnormalities in social communication and interaction, and restricted and repetitive patterns of behavior, interests, or activities (Diagnostic and Statistical Manual of Mental Disorders (5th ed.) (DSM-5), American Psychiatric Association, 2013). Individuals with ASD often experience difficulties with daily interactions and communications, such as interpreting body language, reading facial expressions, and understanding others’ thoughts and desires (e.g. Centelles et al., 2013; Poljac et al., 2013; Senju, 2012). Research also suggests that individuals with ASD lack empathy (e.g. Baron-Cohen and Wheelwright, 2004; Lombardo et al., 2011; Minio-Paluello et al., 2009; Shamay-Tsoory et al., 2002), an aptitude defined as “the ability to form an embodied representation of another’s emotional state, while at the same time being aware of the causal mechanism that induced the emotional state in the other” (Gonzalez-Liencres et al., 2013). This claim has been supported by studies showing decreased self-reported empathy (Baron-Cohen and Wheelwright, 2004; Johnson et al., 2009; Lombardo et al., 2011; but see Dziobek et al., 2008; Rogers et al., 2007 for conflicting findings) and reduced inhibition of corticospinal excitability when observing pain (Minio-Paluello et al., 2009). However, other research has shown that findings on empathic abilities in ASD are not conclusive. For example, Krach et al. (2015) have shown that individuals with ASD showed impairments in observing complex vicarious social pain, while no deficits were found for vicarious physical pain tasks. The authors suggested that subjects with ASD might specifically be deficient in observing complex emotional experiences of others, and that this complexity is not always manipulated in simple experimental tasks. Additionally, Hadjikhani et al. (2014) have shown that emotional reactions to observing facial expressions of people experiencing pain are intact in ASD and that activation in areas associated with pain sharing is comparable to activation found in controls. Together, these studies suggest that there is not simply a general lack of empathy in individuals with ASD, but that the pattern of empathic responses in ASD is more complex.
Recently, it has been suggested that impairments in empathic abilities in ASD may be related to inadequate control over switching between self and other (Bird and Viding, 2014). This relates to the definition of empathy (see above), postulating that it is crucial to be able to distinguish self from other, that is, apprehend that the source of the emotional state lies with the other person, for adequate empathic responding. More specifically, Bird and Viding (2014) suggest that, when others are attended, abnormal self–other switching in ASD should result in an inability to draw distinctions between self and other. This in turn should lead to the observation that individuals with ASD are more affected by other’s emotional states than typically developing (TD) individuals. The latter hypothesis has been supported by research on self-reported empathic responding of individuals with ASD, showing reporting of more empathic responding upon viewing emotional distress in others (Rogers et al., 2007, for a review see Markram et al., 2007). Importantly, Bird and Viding (2014) suggest that the inability to distinguish between self and other that leads to empathic difficulties is based on motor representations. This idea of inadequate control over self–other representations has already been investigated in the motor domain, where echolalia (hyper-imitation of spoken words) and echopraxia (hyper-imitation of movements) are observed clinically, and lab-based research suggests that ASD might be characterized by an inability to control or inhibit imitative tendencies (hyper-imitation; Bird et al., 2007; Hamilton, 2013; Schunke et al., 2015; Sowden et al., 2016; Spengler et al., 2010). This is in contrast with research indicating lack/delay of automatic imitation (hypo-imitation; Cook et al., 2014; McIntosh et al., 2006; Oberman et al., 2009, for a review, see Williams et al., 2004), suggesting impaired processing of others’ actions per se rather than the control over the merging of these representations. However, research has thus far been inconclusive in the motor domain, with numerous studies failing to find evidence for hyper-imitation (Forbes et al., 2016; Gowen et al., 2008; Grecucci et al., 2013; Press et al., 2010; Sowden et al., 2016). Nevertheless, Bird and Viding (2014) propose that adequate distinction between self- and other-related motor representations is crucial for empathic responding and that impairments of empathic abilities in ASD are related to this process (see also De Guzman et al., 2016). Both increased (lack of distinction) and reduced (compensation mechanisms as a response to the lack of distinction) empathic responses could thus be the result of inadequate distinction between self–other-related motor representations. However, a direct test of the hypothesis that empathic abilities in ASD are related to impaired self–other distinction based on motor representations is still lacking.
We have recently developed a paradigm that allows investigating the effect of modulating self–other distinction on empathic responding when observing someone else in pain (empathy for pain; Singer et al., 2004), by introducing an imitation manipulation (De Coster et al., 2013). In this study, TD adults were either being imitated by a video-taped hand on screen or not prior to observing this hand on screen in pain. Using self-report and physiological measures, it was observed that being imitated by the hand on screen led to higher affective reactions to observing painful stimulation to the hand, and that decreased self–other distinction as a result of observing actions that are identical to one’s own mediated this effect. In the current study, this paradigm was used to test whether a manipulation of self–other distinction modulates empathy for pain in adults with high-functioning autism (HFA). Based on the observation that painful stimulation leads to changes in autonomic responses (Preston and De Waal, 2002), somatic responses (startle reflex and skin conductance, as indices of affective value and arousal, respectively) were used as indirect indices of empathy for pain. Importantly, this paradigm allows to directly investigate how control over self–other overlap of motor representations, that is, self–other distinction, affects empathy for pain in HFA.
If empathic responding in HFA would be generally decreased or increased in HFA, and not susceptible to social (self–other) manipulations, no influence of our imitation manipulation on empathy for pain in HFA should be observed. However, if—as suggested by Bird and Viding (2014)—inadequate self–other distinction lies at the basis of deficient empathic responding in ASD, a manipulation of this distinction using an imitation manipulation (decreasing self–other distinction) should result in a pattern of empathic responses in ASD that is different from controls. Thus, we predicted that (1) there would be an effect of our imitation manipulation on empathic responding in individuals with HFA and (2) that adults with HFA would show an amplification of the imitation effect, that is, hyper-empathic responses in response to the observed pain stimuli in the Imitation condition, based on the idea that adults with HFA have more problems in a situation where it is more difficult to distinguish self from other. Thus, this account predicts that empathic responding irrespective of a self–other modulation (i.e. overall empathic responding irrespective of the imitation manipulation) is not impaired in HFA. In sum, the aim of the current study was to gain insight into the complexity of empathic responses in ASD, by investigating whether these responses are modulated by a self–other distinction mechanism based on motor representations.
Materials and methods
Participants
A total of 20 adults with HFA and 20 TD adults aged 21–48 years, all right-handed, participated in the study in exchange for 20€. Adults with HFA (10F, 10M) were recruited via the Flemish Autism Association, while TD adults (10F, 10M) were recruited via the university pool of subjects or advertisements. Chi-square test and T-tests confirmed that no significant demographic differences existed between groups (see Table 1 for participant characteristics). Adults with HFA were required to have an official clinical diagnosis of ASD given by a multidisciplinary team or independent clinician (including autistic disorder, Asperger’s syndrome, and pervasive developmental disorder–not otherwise specified). Furthermore, their status as “high-functioning” was derived from their performance on a standardized cognitive assessment using the Kaufman 2 short form of the Wechsler Adult Intelligence Scale-III (WAIS-III, full scale IQ ⩾ 85; Wechsler, [1987] 1997; Minshew et al., 2005 for the use in adults with HFA). Participants in both groups did not report any additional neurological disorders as indicated by a history of neurological problems such as epilepsy and/or brain trauma and were matched on demographic measures of age (±5 years), handedness, gender, and IQ. The study was granted ethical approval by the local ethics committee at the Faculty of Psychology and Educational Sciences, and all participants provided written consent beforehand. Participants from both groups filled in self-report questionnaires measuring (social) autistic behavior: the Autism Questionnaire (AQ; Baron-Cohen et al., 2001) and the Social Responsiveness Scale–Adult version (SRS-A; Bölte et al., 2011; Constantino et al., 2003; Constantino and Todd, 2005). A total of 14 out of 20 participants with a formal clinical diagnosis of HFA completed the Autism Diagnostic Observation Scale–Module 4 (ADOS; Lord et al., 2000), administered by a trained researcher after the experimental phase (six participants were unable to return). Following liberal ASD inclusion criteria of other studies investigating HFA (e.g. Deschrijver et al., 2016; Lai et al., 2011; Magnee et al., 2008; Zwickel et al., 2011), adults with HFA scored above or one point below cut-off on one subscale of the ADOS and attained an ADOS score of minimum 6. Furthermore, all adults with HFA scored above the SRS-A clinical threshold (T-score ⩾ 76), while no TD adults met this criterion. While not all participants met ADOS criteria, research has shown that this is often the case for individuals with HFA and that this is even more so for female subjects (e.g. Lai et al., 2011). This is likely due to compensatory mechanisms and the fact that most of these individuals have received intensive training that improves social functioning. In total 13 out of 20 individuals with HFA were prescribed antidepressants (10), anti-anxiety drugs (8), and/or antipsychotics (3) at the moment of testing compared to only 4 out of 20 control participants who were prescribed antidepressants.
Participant characteristics for adults with high-functioning autism (HFA) and typically developing (TD) adults.
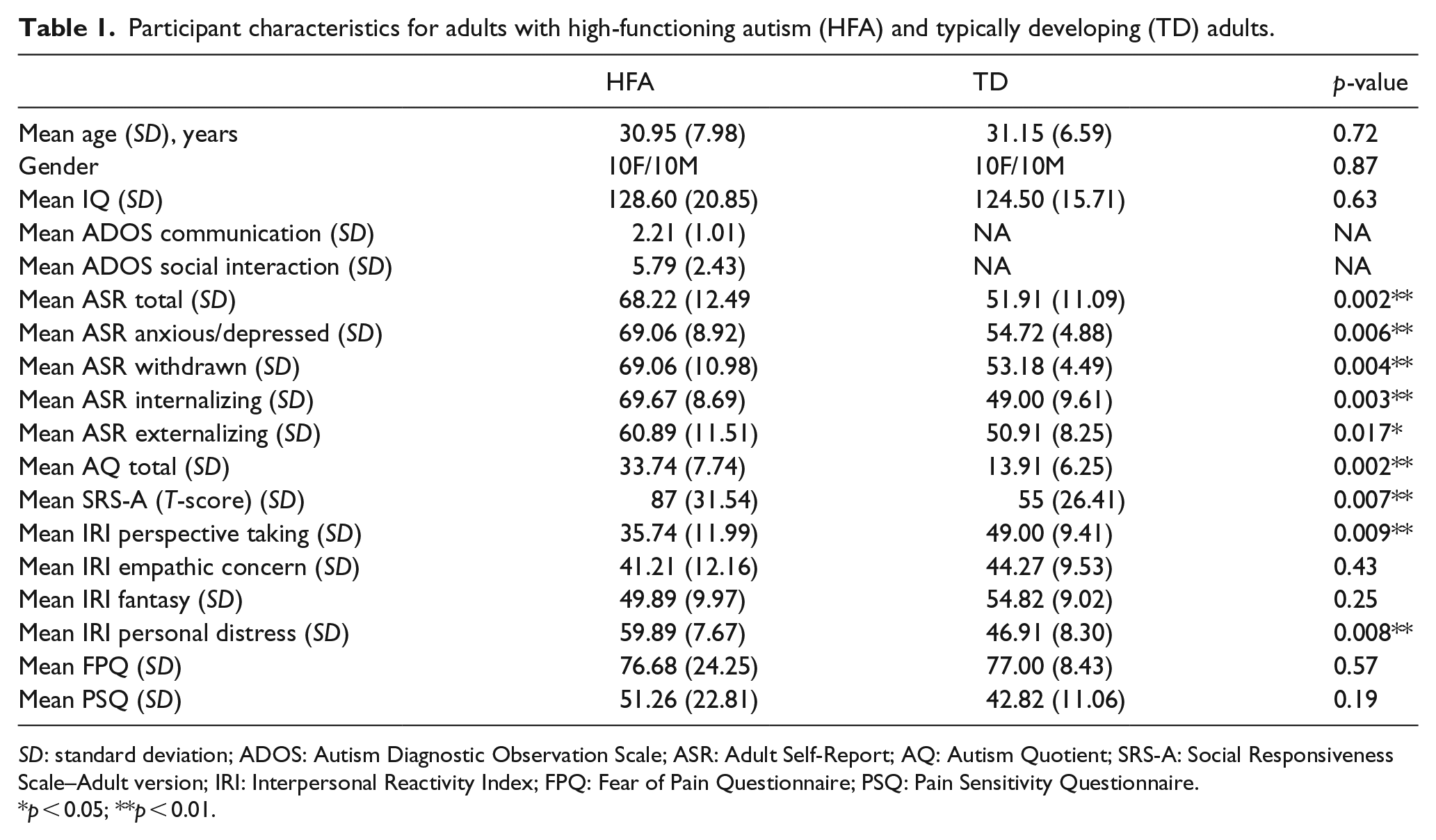
SD: standard deviation; ADOS: Autism Diagnostic Observation Scale; ASR: Adult Self-Report; AQ: Autism Quotient; SRS-A: Social Responsiveness Scale–Adult version; IRI: Interpersonal Reactivity Index; FPQ: Fear of Pain Questionnaire; PSQ: Pain Sensitivity Questionnaire.
p < 0.05; **p < 0.01.
Stimuli and apparatus
Stimulus material consisted of three types of 720 × 576 video-clips: a hand in a resting position, simple finger movements (for the action phase of the task), and pain movies showing a hand receiving pain stimulation (e.g. paper makes a papercut in the hand, nail is knocked into the hand with a hammer; for a complete overview of the pain movies in the pain perception phase, see De Coster et al., 2013).
In the resting state video clip, a right hand with palm down and fingers slightly spread was shown, matching the position of the right hand of participants placed on the response box. This video remained on screen in between presentation of the other videos in order to assure continuous observation of a right hand on screen.
During the action phase of the experimental task, participants carried out simple finger movements of the index, middle, ring, or little finger. These finger movements were recorded with a custom-built response device using light sensors. This device allowed us to use finger lifting movements of participants as triggers for the presentation of the appropriate finger movement video. Temporal resolution was optimized (see the “Procedure” section) so that participants immediately viewed a video-taped finger movement on screen after initiating a finger movement with their own hand. For example, in an imitation block, the lifting of an index finger resulted in the presentation of the index finger lifting video, while the middle, ring, or little finger lifting video was shown in a non-imitation block. All finger movement clips had a total duration of 2000 ms.
The perception phase of the experimental task consisted of the presentation of one of nine pain movies in which painful stimulation was applied to the hand on screen. All movies had a total duration of 8000 ms.
Subjective reports
During the experiment, four behavioral questions were presented after each pain movie, to measure explicit reactions to observing the hand in pain: “How unpleasant do you think the other person found the pain stimulation?,” “How intense do you think the other person experienced painful sensations?,” “How unpleasant did you find the pain stimulation yourself?,” and “How intense did you experience painful sensations yourself?.” Furthermore, a Dutch translation of the scale of Batson et al. (1987) was used, presenting seven items measuring two types of emotional responses. These items inquired about the subjective feelings of participants while viewing painful stimulation, with four items referring to self-oriented feelings (personal distress; “While viewing the painful stimulation of the other person I felt worried/distressed/anxious/sad”) and three items referring to other-oriented feelings (concern; “While viewing the painful stimulation of the other person I felt understanding/empathetic/compassionate”). As such, questions referring to the observed painful situations could be divided into two categories: self versus other.
Procedure
Participants were asked to place the four fingers of their right hand on a custom-made response box. As soon as the video-taped right hand appeared on screen (resting state movie), subjects were instructed to voluntary move a randomly chosen finger that was placed on the response box. Immediately after movement of one of the subjects’ fingers (delay = 0 ms), a movie was shown in which the hand on screen performed the same or a different movement for imitation and non-imitation blocks, respectively. After 20 movements (all imitative or all non-imitative), one of the pain movies was immediately presented. After a pain movie, participants had to rate the 11 behavioral statements on a scale from −5 to +5. This sequence of events was repeated 36 times, combining each of the nine pain movies with both the Imitation and Non-imitation conditions and with a startle and no startle condition. Presentation of these different trial types was randomized, with the limitation that imitation and non-imitation blocks (with and without startle noise) were presented equally frequent during the first and last 18 blocks of the experiment. During the pain clips, a burst of white noise of 95 dB(A) was presented after 4000 ms via headphones in only 50% of the cases in order to avoid predictability of the occurrence of this startle probe (Hawk and Cook, 2000). See Figure 1 for a schematic overview of the procedure.

Design of the experiment. Participants see a hand on screen that either imitates or does not imitate their finger movements (lifting of index, middle, ring, or little finger). After 20 movements, the hand on screen receives a painful stimulus. During and after this pain perception phase, physiological responses and self-reports are measured, respectively.
At the end of the experiment, the Kaufman 2 short form of the WAIS-III (Minshew et al., 2005) was conducted in order to estimate IQ. This short form of the WAIS-III, including the subtests Vocabulary, Similarities, Block Design, and Picture Completion, has been shown to be adequately predictive for total IQ scores. Furthermore, Minshew et al. (2005) have shown that the Kaufman 2 is the most adequate short form for adults with ASD. IQ scores did not differ significantly between both groups (p > 0.05).
Finally, participants were given a set of questionnaires (additional to the AQ and SRS-A described above) to complete at home. This set included the Interpersonal Reactivity Index (IRI; Davis, 1980) as a measure of dispositional trait empathy, The Fear of Pain Questionnaire III (FPQ-III; McNeil and Rainwater, 1998) as a measure of fear of experiencing pain in different situations, and the Pain Sensitivity Questionnaire (PSQ; Ruscheweyh et al., 2009) as a measure of sensitivity for painful stimulation. Furthermore, since ASD shows high comorbidity (Joshi et al., 2013), participants filled in the Adult Self-Report (ASR; Achenbach and Rescorla, 2003) measuring problematic behavior during the past 6 months, providing information on internalizing versus externalizing behavior, such as anxiety, depression, and attention problems, and providing exclusion criteria for controls (e.g. no depression, anxiety, and attention problems were allowed). It has to be noted, however, that the current questionnaires have not been validated in a (Dutch-speaking) ASD population and should thus be interpreted with caution, especially since self-report for socio-emotional states is less reliable in ASD (Hill et al., 2004). See Table 1 for group characteristics and statistical tests.
Electrophysiological recording and analyses
Psychophysiological signals were registered with a Biopac MP150 System and digitalized using AC1001 (AcqKnowledge Software for Windows with Electronic Manual (Biopac Systems, Inc.)).
Startle blink reflex
The startle eye blink reflex was measured according to Blumenthal et al.’s guidelines (2005). Two small Ag/AgCL electrodes (5 mm) were placed over the orbicularis oculi muscle of the left eye, while a ground electrode was placed in the middle of the forehead. The raw electromyography (EMG) signal was amplified with a gain of 5000, filtered with a hardware band pass filter of 0.5–500 Hz, and digitally sampled at 1000 Hz, later offline rectified and integrated with psychophysiological analysis (PSPHA) (De Clercq et al., 2006). The magnitude of the eye blink amplitude was computed as the subtraction of the mean rectified baseline value (0–20 ms after probe onset) from the rectified peak value in the 21- to 120-ms interval after probe onset. Trials on which baseline values deviated more than 2.5 standard deviation (SD) from the mean baseline value of the subject were eliminated. Finally, reflex magnitudes were converted to T-scores across trials on a within-participant basis to adjust for between-participant differences in response and baseline EMG magnitude (Funayama et al., 2001).
Skin conductance
Skin conductance was measured as an index of autonomic functioning that has been shown to be responsive to negative emotional stimuli (Bradley et al., 2001). Skin conductance was measured using a constant voltage (0.5 V) and two Ag/AgCL electrodes with a diameter of 8 mm. The electrodes were filled with conductive gel and were attached on the thenar and hypothenar eminences of the left hand. Skin conductance was digitized at 10 Hz for the entire duration of the pain movie (8000 ms). Using PSPHA, skin conductance responses were calculated as the difference between the highest and the lowest value in this 8000-ms time window. In order to normalize the data, skin conductance amplitudes were square root transformed prior to analysis (Dawson et al., 2000).
Data analysis
Due to equipment failure, one participant was excluded from all psychophysiological analyses, and one additional participant was excluded for blink modulation data only (both in the HFA group). No outliers were removed from either group. Cronbach’s α for all measures, in both conditions, for both groups were all >0.70.
The effect of being imitated on empathy for pain
We performed a linear mixed-model analysis using the lme4 package in R (R Foundation for Statistical Computing, 2015) using a 95% confidence interval (CI) for all statistical tests and restricted maximum likelihood to estimate model parameters. Individual subjects were included as random effects in the model. The dependent variables were subjective reports, that is, self- and other-related items, and physiological measures, that is, startle blink magnitude and skin conductance response. These variables were simultaneously modeled as a function of a fixed within-subjects factor Condition (Non-imitation = 0, Imitation = 1) and a fixed between-subjects factor group (TD = 0, HFA = 1). The interaction Condition × Group was subsequently modeled. Post hoc comparisons were performed using planned contrasts, applying Holm–Bonferroni correction to correct for multiple testing.
Exploratory analyses of delayed effects
During a formal debriefing of the HFA group after the experiment, several participants revealed that they felt to have been (too) strongly affected by the pain videos in the beginning of the experiment. According to these participants, this effect was attenuated over the course of the experiment by developing strategies to cope with this sensation (e.g. diverting attention away from the stimuli). Thus, exploratory analyses were performed using the same aforementioned linear mixed model, adding Time of the experiment as a continuous covariate (36 time points, one for each block) and its interactions with the other factors to the model. However, these analyses were performed post hoc and as such are exploratory. Thus, caution in interpretation of these tests is warranted, and replication of these effects is necessary.
Correlational analyses
Correlation analyses were performed between questionnaire scores on the AQ for the whole population (Hoekstra et al., 2007), as well as ADOS scores for the HFA group. Correlations were calculated using the difference between both conditions (Condition effect: Imitation–Non-imitation), where a positive effect of Condition indicates stronger empathic responding in the Imitation condition, while the reverse is true for a negative effect of Condition. Additionally, correlations were also performed for the Imitation and Non-imitation conditions separately. Bonferroni correction was used to control for multiple testing.
Results
The effects of being imitated on empathy for pain
Subjective reports: other-related items
For other-related items, neither Condition nor Group had a significant effect on self-reports, β = 0.11, t = 1.16, p = 0.35 and β = 0.21, t = 0.43, p = 0.41, respectively. No significant interaction was observed between Group and Condition, β = 0.09, t = 0.18, p = 0.55 (see Figure 2(a)).

Mean scores (range from −5 to +5) in the Imitation and Non-imitation conditions in both groups, for (a) other- and (b) self-related items referring to pain. Error bars are standard errors of the mean
Subjective reports: self-related items
Analysis of self-related items revealed a significant effect of Condition on self-related self-reports, β = 1.14, t = 2.20, p < 0.05. This indicates higher scores in the Imitation (M = −0.61, SD = 0.30) compared to the Non-imitation condition (M = −0.80, SD = 0.32). Neither an effect of Group, β = 0.12, t = 1.07, p = 0.21, nor an interaction effect was observed, β = −0.15, t = 0.18, p = 0.75 (see Figure 2(b)).
Blink modulation
No effect of Group was observed, β = 0.13, t = 1.14, p = 0.26. A significant effect of Condition, β = 2.28, t = 2.15, p < 0.05, was observed, indicating higher startle blink magnitude in the Imitation condition (M = 50.52, SD = 1.94) compared to the Non-imitation condition (M = 48.49, SD = 1.82). Furthermore, a significant interaction between Group and Condition was observed, β = 3.18, t = 2.84, p < 0.01. Planned contrasts showed that higher startle magnitude in the Imitation (M = 51.38, SD = 2.10) compared to the Non-imitation condition (M = 46.97, SD = 0.53) was only observed in the TD group, β = 2.32, t = 2.34, p < 0.01. No difference between the Imitation (M = 49.67, SD = 1.30) and Non-imitation conditions (M = 50.02, SD = 1.32) was observed in the HFA group, β = 0.87, t = 0.89, p = 0.47 (see Figure 3).

Mean blink magnitude in the Imitation and Non-imitation conditions in both groups. Magnitude is expressed as within-subjects T-scores. Error bars are standard errors of the mean.
Skin conductance
An effect of Group was not observed, β = 0.03, t = 0.19, p = 0.96. A significant effect of Condition, β = 2.12, t = 2.74, p < 0.05, indicating higher skin conductance responses in the Imitation condition (M = 0.36, SD = 0.03) compared to the Non-imitation condition (M = 0.28, SD = 0.11) was shown. Additionally, the interaction effect between Group and Condition was significant, β = 4.28, t = 3.01, p < 0.01. Planned contrasts revealed that higher skin conductance responses in the Imitation (M = 0.33, SD = 0.01) compared to the Non-imitation condition (M = 0.23, SD = 0.11) was only observed in the TD group, β = 3.11, t = 2.39, p < 0.01. No difference was observed in the HFA group, β = 0.12, t = 1.02, p = 0.25, between the Imitation (M = 0.39, SD = 0.01) and Non-imitation condition (M = 0.34, SD = 0.14) (see Figure 4).

Mean skin conductance in the Imitation and Non-imitation conditions in both groups. Skin conductance responses are expressed as the difference between the highest and the lowest value in a specified time window. Error bars are standard errors of the mean.
Exploratory analyses of delayed effects
Subjective reports: other-related items
When including Time and its interactions into the analysis, an effect of Time was observed, β = −2.23, t = −2.05, p < 0.05, indicating a decrease of other-related self-reports over time (see Figure 5(a)).

Mean scores (range from −5 to +5) in the Imitation and Non-imitation conditions for both groups, for (a) other- and (b) self-related items referring to pain when including time as a continuous covariate. Shaded areas indicate 95% CI intervals. Raw data mean values are also depicted.
Subjective reports: self-related items
An interaction between Group and Time was observed, β = −2.18, t = −2.01, p < 0.05. A negative effect of Time was only present in the HFA group, β = −3.28, t = −2.24, p < 0.05, indicating a decrease of self-related reports over time for individuals with HFA only, while no effect of Time was observed for TD, β = −0.11, t = −1.57, p = 0.21 (see Figure 5(b)).
Blink modulation
When including Time into the mixed model, the three-way interaction effect between Group, Condition, and Time proved to be significant, β = 3.16, t = 2.57, p < 0.01. The interaction between Condition and Time was significant in the HFA group only, β = 3.27, t = 2.04, p < 0.05, and not in the TD group, β = 0.07, t = 0.87, p = 0.34. Follow-up tests in the HFA group showed that the effect of Time was reversed for both conditions. For the Imitation condition, scores increased significantly over Time, β = 2.19, t = 3.01, p < 0.05, while scores in the Non-imitation condition decreased over Time, β = −1.81, t = −2.18, p < 0.05. Figure 6 suggests that this led to a reversal of the effect of being imitated in the beginning of the experiment (looking at the first 18 blocks), that is, lower scores after being imitated compared to not being imitated, β = −2.67, t = −2.17, p < 0.05. At the end of the experiment (looking at the last 18 blocks), scores were higher after being imitated (although not significantly), replicating the pattern found in the TD group, β = 1.57, t = 1.54, p = 0.12.

Mean blink magnitude in the Imitation and Non-imitation conditions in both groups when including Time as a continuous covariate. Magnitude is expressed as within subjects T-scores. Shaded areas indicate 95% CI intervals. Raw data mean values are also depicted. p-values were calculated over the first and last 18 blocks of the experiment separately.
Skin conductance
When including Time, the three-way interaction effect between Group, Condition, and Time was significant, β = 3.23, t = 3.09, p < 0.01. The interaction between Condition and Time was significant in the HFA group only, β = 3.31, t = 2.86, p < 0.05, and not in the TD group, β = 0.17, t = 0.75, p = 0.49. In the HFA group, the effect of Time increased in the Imitation condition, β = 2.29, t = 2.61, p < 0.05, and decreased significantly in the Non-imitation condition, β = −2.14, t = −2.07, p < 0.05. Figure 7 indicates that the pattern found for skin conductance responses was similar to the pattern found for blink modulation when comparing scores in the Imitation and Non-imitation conditions, with lower scores after being imitated in the beginning of the experiment, β = −2.03, t = −1.87, p = 0.08, and higher scores after being imitated at the end of the experiment, β = 2.16, t = 1.91, p = 0.07.

Mean skin conductance in the Imitation and Non-imitation conditions in both groups when including Time as a continuous covariate. Skin conductance responses are expressed as the difference between the highest and the lowest value in a specified time window. Shaded areas indicate 95% CI intervals. Raw data mean values are also depicted. p-values were calculated over the first and last 18 blocks of the experiment separately.
Correlational analyses
Correlations are presented in Table 2. No significant correlations, corrected for multiple testing, were found for any of the dependent variables when looking at effects of our primary analyses. However, because of the strong effect of Time in our exploratory analyses, additional correlations were calculated between the questionnaire scores and effects for both halves of the experiment separately. These analyses revealed a significant negative correlation between AQ scores and the Condition effect for skin conductance responses during the first half of the experiment, r = −0.42, p < 0.001, indicating less empathic responding in the Imitation compared to the Non-imitation condition for individuals with higher scores of autism severity as measured with the AQ. No significant correlation was found for the second half of the experiment (p > 0.21). When looking at the Imitation and Non-imitation conditions separately for skin conductance responses, a significant negative correlation was found between the Imitation condition and AQ scores in the first half of the experiment only, r = −0.33, p < 0.001, suggesting that the correlation with the Condition effect was driven by reduced empathic responding in the Imitation condition for individuals with higher AQ scores. For startle magnitude responses, only a negative correlation was observed between the AQ scores and the Imitation condition in the first half of the experiment, r = −0.24, p < 0.001. All other correlations did not survive multiple testing corrections.
Correlations between questionnaire scores and the Condition (Imitation–Non-imitation), Imitation, and Non-imitation effect of the experiment.

AQ: Autism Quotient; ADOS: Autism Diagnostic Observation Scale
Correlations were calculated for the overall experiment and the first 18 and last 18 blocks of the experiment separately. Note that ADOS correlations are for individuals with high-functioning autism (HFA) only.
p < 0.05; **denotes correlations that survive multiple testing correction.
The role of alexithymia
While we did not measure alexithymia in this study, we retrieved scores from item B5 of the ADOS–Module 4 (for the HFA group only), an item representing communication of own feelings and thus closely related to the construct of alexithymia. Higher scores on this item, suggesting more problems with describing own emotions and feelings, were related to a stronger reversal of the Imitation effect (lower scores after being imitated) for skin conductance responses in the beginning of the experiment, r = −0.44, p < 0.001. This correlation was similar to the negative correlation found for AQ scores. To examine the unique contribution of AQ, we calculated a partial correlation between AQ scores and the reversal effect, partialling out item B5. The correlation, though reduced, remained significant, rp = −0.27, p < 0.01.
Discussion
Individuals with ASD have been reported to show abnormalities in empathy (Baron-Cohen and Wheelwright, 2004; Lombardo et al., 2011; Minio-Paluello et al., 2009; Shamay-Tsoory et al., 2002). However, the nature and origin of these deficits have been debated extensively. The aim of this study was to address the question whether empathic responding in ASD is modulated by control over self–other overlap, that is, self–other distinction, based on motor representations as recently suggested by Bird and Viding (2014). To achieve this aim, we used a validated paradigm, in which reactions to observing someone else in pain are heightened after being imitated compared to not being imitated, due to decreased self–other distinction when being imitated (De Coster et al., 2013). A group of adults with HFA and age- and gender-matched TD adults observed a hand on screen in pain subsequent to being imitated by this hand on screen or not. First, overall, adults with HFA showed similar responses on affective indices of seeing someone else in pain compared to TD adults, irrespective of being imitated or not. This indicates that no general deficit in empathy for pain was present in these adults with HFA. Second, while the pattern of TD adults replicated previous findings showing higher empathy for pain after being imitated in both explicit and implicit measures, adults with HFA showed a distinct pattern of results. On first inspection, startle blink magnitude and skin conductance responses, as measures of autonomous nervous functioning, revealed no influence of being imitated compared to not being imitated on empathy for pain. However, when conducting exploratory post hoc analyses looking at the effect of time, it was observed that the influence of being imitated on empathy for pain changed over time for adults with HFA only. While responses in adults with HFA initially showed a reversed influence of being imitated on empathy responses (smaller responses in the Imitation compared to the Non-imitation condition), the pattern later in time was similar to the pattern seen in TD adults (larger responses in the Imitation condition). Furthermore, while subjective reports on items related to own experiences when observing pain were higher in the Imitation compared to the Non-imitation condition for both groups, scores decreased significantly over time for adults with HFA only.
First, the findings of this study clearly suggest that adults with HFA and TD adults show equal, increased affective responses when observing someone else in pain irrespective of being imitated or not. This is in line with research indicating no general empathy deficit in ASD, either by showing similar empathy for pain responses in ASD compared to healthy controls (e.g. Hadjikhani et al., 2014) or by suggesting that abnormalities in empathy for pain in ASD are dependent on modulating factors (e.g. Krach et al., 2015). However, research has been inconclusive regarding the underlying mechanisms of (deficits of) empathic responding in ASD. More specifically, there has been extensive debate concerning self–other representational sharing mechanisms, with research suggesting both impaired (e.g. Baron-Cohen and Wheelwright, 2004; Lombardo et al., 2011; Minio-Paluello et al., 2009; Shamay-Tsoory et al., 2002) and intact (e.g. Hadjikhani et al., 2014) self–other representations in ASD. Bird and Viding (2014), and more recently De Guzman et al. (2016), however, argue that adequate empathic responding is specifically dependent upon control over self–other representations, rather than these representations per se. Furthermore, these authors claim that deficient control over these self–other representations might lie at the roots of empathy deficits in ASD.
An important aim of this study was to investigate this idea put forward by Bird and Viding (2014), by testing whether a manipulation of self–other distinction based on motor representations had an influence on empathy for pain in adults with HFA. Our primary analysis looking at the influence of being imitated on empathy for pain, however, showed no difference between empathic responding after being imitated compared to not being imitated in HFA. At first glance, these findings seem to suggest that adults with HFA, in contrast to TD adults who did show a difference between both conditions, were not susceptible to the modulation of self–other distinction. This finding would support the idea that empathic responses in HFA are not modulated by this social manipulation and would contradict the hypothesis that self–other distinction is a crucial variable. Interestingly, a more fine-grained exploratory post hoc analysis of our data looking at the effect of time on the modulation of being imitated on empathic responses led to completely different conclusions. This post hoc analysis seemingly supports the idea of compromised self–other distinction in adults with HFA. Here, we showed that being imitated does have an influence on empathy for pain in HFA, but that the pattern of this influence is different over time for adults with HFA compared to TD adults. In TD adults, we replicated previous results by showing increased affective responding after being imitated over the whole course of the experiment. In contrast, adults with HFA showed an increase of empathic responding over time in the Imitation condition and a decrease over time in the Non-imitation condition. This resulted in a reversed influence in the Imitation compared to the Non-imitation condition in the beginning of the experiment and a more typical influence of being imitated on empathy for pain at the end of the experiment. These results, contrary to our primary analyses, suggest that participants in the HFA group were very sensitive to the imitation manipulation, but responded to the manipulation in a counterintuitive manner, showing an initial reversal of the empathic responding pattern. It is important to note, however, that these analyses were conducted post hoc and were primarily based on qualitative subjective reporting of our participants with HFA (e.g. “I found it difficult to deal with the painful stimuli and the matching movements in the beginning of the experiment.”). As such, these effects need to be interpreted with caution and are in need of replication. Nevertheless, we believe that these findings are supportive of compromised control over self–other merging and its link to empathic responding in HFA. We speculate that the reversal of the effect of being imitated on empathy for pain in the beginning of the experiment reflects a coping mechanism in response to an abnormal decrease of self–other distinction that may be perceived as too intense in HFA. Indeed, several adults with HFA informally reported heightened and uncomfortable experiences while perceiving pain in combination with being imitated, as well as individual strategies and mechanisms to deal with these heightened sensations (e.g. averting attention away from the imitating hand). Importantly, individual reports indicated that these strategies were specifically apparent during the first part of the experiment. We believe that these coping mechanisms led to the counterintuitive finding of a reversed influence of being imitated on empathy for pain in the beginning of our study. Additional subjective reports of participants suggested that habituation over time took place, eliminating the need for coping mechanisms (sometimes using additional strategies such as reminding oneself that the hand on screen was not real), which would explain the change of responses over time. This seems to be supported by the observation that ratings on subjective self-reports after observing the hand in pain decreased over the course of the experiment for adults with HFA only, although this decrease was not specific for the Imitation condition. The idea of hypersensitivity of adults with ASD to (novel) sensory stimuli—such as the pain videos in this study, selectively in the first part of the experiment—has been put forward by other research (Palmer et al., 2013; Paton et al., 2012; Pellicano and Burr, 2012; Van Boxtel and Lu, 2013) and notions such as the “intense world theory” (Markram et al., 2007; Markram and Markram, 2010). The idea of coping, however, needs to be explored in future research given that we were unable to systematically explore this notion due to the post hoc nature of the finding. For example, measuring eye gaze as an index of diverting attention away from the presented stimuli could be examined to verify whether and how an attention-diversion coping strategy affects the current effects. Additionally, future research is necessary to systematically investigate the idea that these coping mechanisms change over time and are a result of self–other distinction difficulties (e.g. looking at neural substrates related to control over self–other sharing such as the temporoparietal junction (TPJ) and medial prefrontal cortex (mPFC); Spengler et al., 2010). Interestingly, the idea of changes over time of socio-cognitive abilities (such as perspective taking) as a result of training of control over self–other overlap has been put forward in previous research (Santiesteban et al., 2011), as well as the ideas that imitation and synchrony improve emotion inference and cognitive empathy (Köhne et al., 2016a, 2016b), improved spontaneous object play (Ingersoll and Gergans, 2007; Ingersoll and Schreibman, 2006), gesture imitation (Ingersoll et al., 2007), and spontaneous imitation (Ingersoll, 2010). Finally, it is important to note that self-report and physiological responses in the current experiment did not show similar results concerning modulation by our imitation manipulation. However, we believe this might have been related to the nature of self-reports and the design of this study (see De Coster et al., 2013), as well as the fact that self-reports and psychophysiological recording are not always associated with HFA (Cook et al., 2013; Fan et al., 2013; Hill et al., 2004).
This study has some important limitations. First, research has shown that emotional impairments and empathic deficits in ASD are due to the comorbidity with alexithymia, described as difficulties with identifying and describing own feelings (Nemiah et al., 1976), rather than ASD per se (Bird et al., 2010; Cook et al., 2013; Heaton et al., 2012). Further studies have indicated considerable impairments in own emotional processing and reporting in individuals with ASD, indicating less reliable self-reports and pointing toward a possible distinction between self-reported and other measures of empathic responding (Hill et al., 2004; see Bird and Cook, 2013 for a review). A major limitation of this study is the absence of an alexithymia measure. However, additional correlational analyses using item B5 of the ADOS–Module 4 as a proxy of alexithymia indicated that although part of the findings might be related to alexithymia rather than ASD, ASD symptomatology still had a unique role in explaining the current findings. This was evidenced by a reduced but still significant correlation between AQ scores and the reversal effect when partialling out item B5. It is important to note, however, that item B5 of the ADOS does not cover alexithymia fully, and that it is tightly linked to ASD severity given that it is measured as part of the ADOS module (although not part of its total score). Thus, future research is needed to further investigate the role of alexithymia in self–other distinction and empathy for pain in ASD. As a second limitation, it has to be noted that the physiological responses observed during the pain perception phase might have carried on residual activation from the action phase of the experiment, especially given that skin conductance has been shown to be a slow response (e.g. Geuter et al., 2014). As such, it is necessary to conduct further research and investigate what is happening during the action phase, that is, being imitated versus not being imitated, to gain insight into the timing and origin of the observed effects. Finally, this study was conducted with a group of high-functioning participants with ASD of which some did not reach cut-off scores on the ADOS (indicating mild ASD presentation). Future research is needed to explore the current effects and claims with adults with more severe ASD symptomatology. Other interesting avenues for future studies include ideas based on intriguing research by Rütgen et al. (2015a, 2015b) that has recently shown that empathy for pain is grounded in first-hand experience of pain and that this involves opioid neurotransmitter systems. Interestingly, acting in synchrony and social bonding have also been linked to this system (Cohen et al., 2010). Since the current paradigm brings together empathy and synchrony (of which being imitated is a special case) literature, it would be interesting to investigate whether a link between imitation, opioids, and empathy for pain exists, and whether individuals with autism show a delayed response of this system. Importantly, this points to the possible clinical applications that might result from further research based on the current findings. Second, the current paradigm has also been related to agency (De Coster et al., 2014), suggesting that being imitated induces control over the hand on screen. Again, this line of research has shown conflicting results concerning sense of agency in ASD with studies pointing to preserved (David et al., 2008; Williams and Happe, 2009) versus abnormal sense of agency (Grynszpan et al., 2012; Sperduti et al., 2014; Zalla et al., 2015; see Zalla and Sperduti, 2015 for a review). The relationship between self–other distinction, empathy, and agency, however, is still unclear.
In sum, this study aimed to investigate the effects of self–other merging based on motor representations on empathic responding in HFA. While initial analyses suggested that decreasing self–other distinction by being imitated did not seem to influence affective responding to painful stimulation in HFA (contrary to controls), secondary analyses showed that the effect of imitation was strongly modulated by time. Adults with HFA showed an increase of empathic responding over time in an Imitation condition and a decrease over time in a Non-imitation condition. Exploratory analyses suggested that this resulted in reduced responding after being imitated in the first part of the experiment, and that this pattern normalized over the course of the study. These results provide important insights into the role of time when looking at the influence of self–other distinction on empathy in HFA, suggesting that training of this mechanism over time might be an interesting avenue for future remediation therapies. However, these findings were based on exploratory analyses and more direct investigation of this account is necessary, as well as the idea that coping mechanisms related to abnormal control over self–other sharing might underlie several abnormalities observed in HFA. Nevertheless, the current findings are an important first step in pointing out the relevance of these ideas in ASD research on empathic responding.
Supplemental Material
AUT701268_Lay_Abstract – Supplemental material for The effect of being imitated on empathy for pain in adults with high-functioning autism: Disturbed self–other distinction leads to altered empathic responding
Supplemental material, AUT701268_Lay_Abstract for The effect of being imitated on empathy for pain in adults with high-functioning autism: Disturbed self–other distinction leads to altered empathic responding by Lize De Coster, Jan R Wiersema, Eliane Deschrijver and Marcel Brass in Autism
Footnotes
Acknowledgements
The authors would like to thank Febe Busseniers for her help in recruitment of participants and collecting data.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by Research Foundation Flanders Grant No. FWO10/ASP/321 to the first author.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.