
Abstract
Background
Evidence supports the neuroprotective effects of physical activity, either in experimental animal models or humans. However, the biological mechanisms by which physical exercise modulates dementia and Alzheimer's disease (AD) progression are still unclear.
Objective
This study investigated whether long-term (6 months) of voluntary wheel running induces neuroprotective effects in the pathogenesis of AD in middle-aged (8 months) female mice, focusing on energy metabolism.
Methods
A genetic mice model of AD (3xTg-AD) that performed wheel running presented changes in body metabolism and muscle oxidative profile, as well as restored discriminative and non-associative retention memories, evaluated by novel object recognition and open field tasks, respectively.
Results
In the hippocampus, these mice exhibited reduced levels of amyloidogenic AβPPβ fragment, phospho-Tau protein and phospho-Akt (activated form), without changes in phospho-AMPK (activated form). In addition, hippocampal mitochondria presented a restored respiratory function, characterized by lower coupling degree and weak contribution from complex I found in 3xTg-AD mice.
Conclusions
The results demonstrated that voluntary exercise improves cognitive parameters and biochemical hallmarks of AD, modulates Akt activation and enhances mitochondrial energy metabolism in hippocampus of middle-aged 3xTg-AD female mice, thereby reinforcing the neuroprotective role of physical exercise and the involvement of mitochondria in the etiology of the AD.
Introduction
Alzheimer's disease (AD) is currently a global challenge, accounting for 60%–80% of dementia cases, with more than 55 million people affected worldwide. 1 Characterized by neurodegenerative disorder associated with learning, memory, and communication deficits, AD remains a disease without a treatment capable of halting its progression. 2
The primary pathological marker of AD is the production and deposition of extracellular amyloid-β peptides (Aβ) and the intracellular aggregates of hyperphosphorylated tau proteins.3,4 The production of Aβ occurs through the sequential cleavage of amyloid-β protein precursor (AβPP) by β-secretase (BACE) and γ-secretase enzymes. 5 However, the complete understanding of the triggers that increase Aβ production and deposition, as well as tau aggregation, remains debatable. Metabolic dysfunctions are probable candidates, given their identification as risk factors for AD, including diabetes, obesity, and physical inactivity.6,7 Mitochondrial metabolic dysfunction is also implicated in the etiology of AD,8–13 reinforced by growing evidence demonstrating co-localization of AβPP (wild-type or mutant AβPPSwed) and Aβ with mitochondria in postmortem AD brains, AD animal and cell models. 14 However, the detailed mechanism of this relationship is still elusive.15,16
Physical exercise has emerged as a potential therapy that can reduce the prevalence of AD by indirectly modifying risk factors 17 and directly reducing Aβ production and deposition and tau aggregation.18,19 In a meta-analysis study conducted by Hamer and Chida, 20 it was suggested that physical exercise in both genders and across different training regimes was associated with a 28% reduction in dementia and a 45% reduction in AD. Therefore, this study aimed was to investigate whether long-term (6 months) voluntary wheel-running induces neuroprotective effects in the pathogenesis of AD in middle-aged (8 months) 3xTg-AD female mice, focusing on energy metabolism. This genetic mice model of AD present a set of neurodegenerative biomarkers, such as early long-term memory impairment (4 months), age-related Aβ plaques and tau phosphorylation species in hippocampal regions. 21 Here we analyzed cognitive parameters (discriminative memory and habitual memory), the levels of pathological AD hallmarks (AβPP-C99 amyloidogenic fragments and phospho-Tau protein), body and muscle metabolisms, the activation levels of key metabolic sensors enzymes (Akt and AMPK) and mitochondrial energy metabolism in the hippocampus. Our results demonstrated that voluntary exercise improves cognitive and biochemical parameters of AD and modulate Akt activation and mitochondrial energy metabolism in the hippocampus of middle-aged 3xTg-AD female mice.
Methods
Animals
All experiments were approved by the Animal Care and Use Committee of the School of Pharmaceutical Sciences of Ribeirão Preto, USP (Protocols no. 22.1.292.60.9). Female 3xTg-AD mice (B6; 129-Psen1tm1Mpm Tg (AβPPSwe, tauP301L) 1L fa/MmJax; 24 weeks old) and background strain B6129SF2/J (wide-type control; WT) were provided by the Center of animals from the School of Pharmaceutical Sciences of Ribeirão Preto, USP. The animals were maintained at the vivarium of the School of Pharmaceutical Sciences of Ribeirão Preto, housed in groups of 4–6 per cage (size 18 × 33.5 × 41 cm, H × B × L) under 20 ± 2° C, in a 12-h light/dark cycle with free access to standard chow and drink water.
Voluntary wheel running and experimental design
After acclimatization, 8-month-old mice (both wild-type [WT] and 3xTg-AD) were housed either without access to a running wheel (sedentary, SED) or with free access to a running wheel (voluntary wheel running, VWR) measuring 20 cm in diameter and 8 cm in width (Bragança Wired Exerciser, São Paulo, SP). Together, the group of mouse per cage (4–6 animals) moved the wheel an average of 12–18 km/day, without differences between WT and 3xTg-AD. After 4 months, the animals underwent an Incremental Load Test and Indirect Calorimetry (see Figure 1A) to verify the exercise adaption of each animal. Over the subsequent 2 months, mice were subjected to cognitive tests in the following sequence: the Open field (OF), Novel object recognition (NOR), and Active Avoidance (AA) tests. At 14 months of age, mice were euthanized, and the brain, heart, and soleus muscle were excised. Half of the hippocampus was promptly utilized for respirometry assays, while the remaining tissues were stored at −80°C.
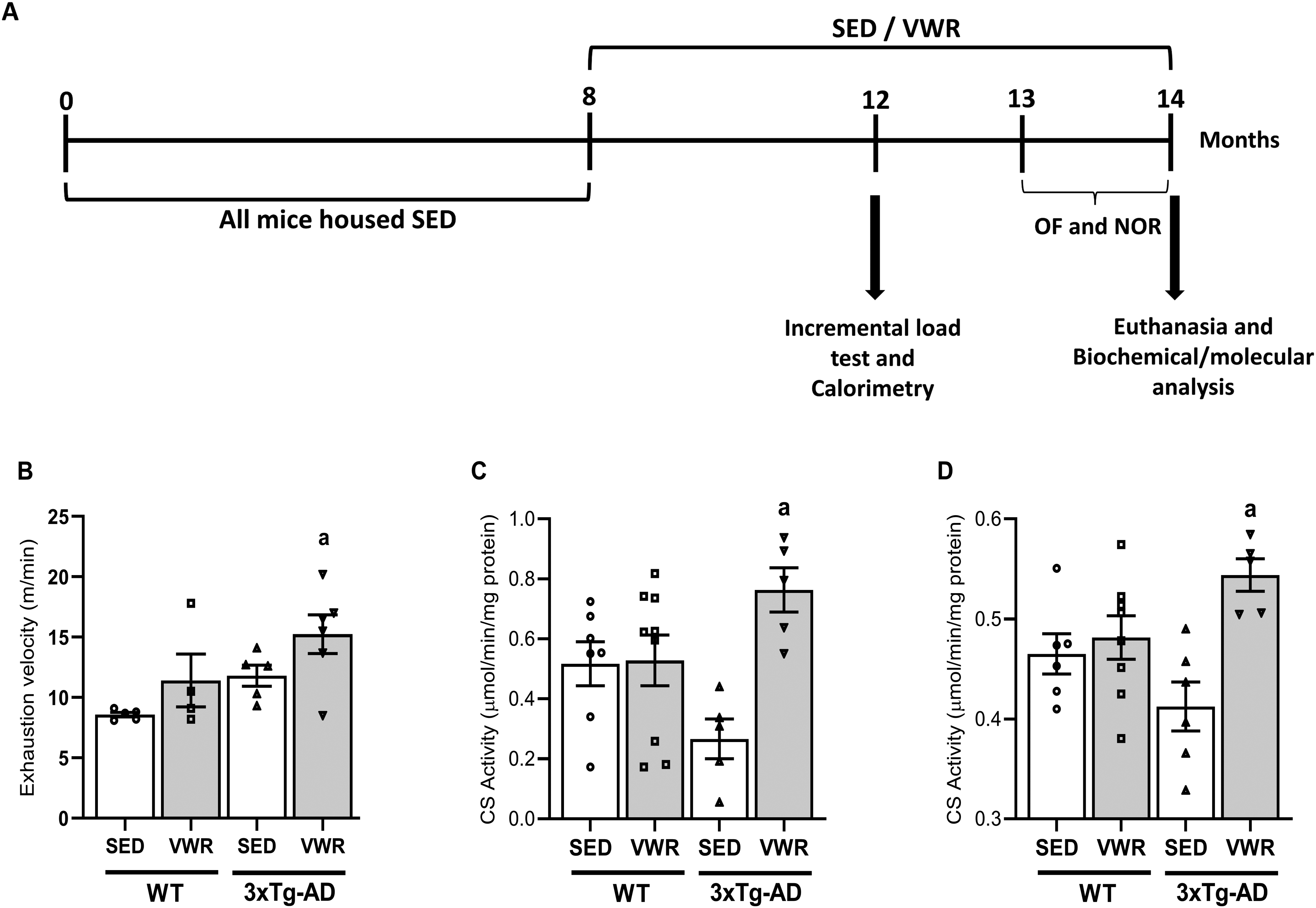
Six months of VWR improves the oxidative metabolism of middle-aged 3xTg-AD female mice. (A) Experimental timeline of 8-months-old WT and 3xTg-AD sedentary (SED) or voluntary wheel running (VWR) housing for 6 months, followed by behavioral tests: Open Field (OF), Novel Object Recognition (NOR), and Active Avoidance (AA) tasks. (B) Incremental load test. (C) Citrate synthase (CS) activity in heart muscle. (D) CS activity in soleus muscle. Data are expressed as the mean ± SEM; a = p < 0.05 versus Wt_SED (one-way ANOVA, Tukey's post-hoc test); N = 8 (WT_SED), 7 (WT_VWR), 6 (3xTg-AD_SED), and 6 (3xTg-AD_VWR).
Incremental load test
The test was conducted on a treadmill (AVS Special projects®), initially set at 9 m/min velocity with a 20% incline and with increments of 2 m/min every 2 min until the mice reached voluntary exhaustion. Exhaustion was defined as the point at which the animals touched the end of the treadmill five times within a 1-min interval. 22
Citrate synthase activity
Soleus and heart muscles were homogenized in 250 mM Tris-HCl buffer (pH 7.4), 2 mM EGTA, and 250 mM sucrose. The Bradford method 23 (Bio-Rad, CA, USA) was used to determine protein concentration. The citrate synthase activity was measured following the increase in 5,5′-dithiobis (2-nitrobenzoic acid) absorbance at 412 nm in the presence of acetyl-CoA, according to previous protocol. 24 Reactions were adjusted to fit in a 96-well microplate platform, using 10 µg of protein/well.
Indirect calorimetry
Mice were placed individually in a hermetic chamber connected to an indirect calorimetry system (Oxylet Physiocage™, PanLab, Barcelona, Spain) with free access to chow and drink water. After 2 h of habituation, oxygen consumption (VO2) and CO2 release (VCO2) were recorded for 24 h. Respiratory quotient (RQ) was calculated as VCO2/VO2 and used to estimate macronutrient oxidation. 25 Energy expenditure was calculated using the following formula: EE = [3.85 + (1.232 × RQ)] × VO2 × 1.44.
Open-field (OF) task
This task evaluates motor performance in the training section and non-associative memory in the retention test session. Habituation to the open field was carried out in an acrylic arena (46 × 46 × 46 cm) surrounded by infrared sensors, responsible for detecting the animal's position throughout the monitoring period. Thus, it was possible to calculate the average speed rate and the total distance traveled throughout the experiment using dedicated software (Insight, Ribeirão Preto, Brazil). The animals were placed in the center of the arena to explore it for 5 min (training session) and, after 24 h, they were again subjected to a similar open field session (test session). The total distance covered and rearing in both sessions were counted. The decrease in the total distance covered and the number of rearings between the two sessions were used as measures of the retention of habituation. 26 The arena was cleaned with 30% ethanol and water between each session.
Novel object recognition (NOR) task
The NOR task evaluated discriminative short-term memory. 27 The mice were habituated to the empty arena (without any objects) for 10 min per 3 consecutive days. Twenty-four hours after the habituation days, during the training phase, the mice were subjected to two identical objects (shape and size) at two adjacent corners of the arena, each 10 cm away from the side of the walls. Each time, the mice were placed in the middle of the box, facing away from the objects, and they were allowed to explore the objects for 5 min. After a delay of 15 min, the test phase was conducted for 5 min. One previously introduced object was replaced by an object of the same size but different shape and color in the same places. Exploration time was analyzed for the arrangement of identical objects from the training phase (exploration time) and different objects from the test phase (discrimination time). A recognition index was calculated and reported as the ratio TB/(TA + TB) (TA = time exploring familiar object A, and TB = time spent exploring new object B). Sniffing or touching the object with the nose or front paws was defined as exploration.28,29
Immunoblotting technique
The hippocampus and cortex were homogenized in RIPA buffer (50 mM Tris-HCl, pH 7.5, 150 mM NaCl, 1% Triton, 0,01% SDS, 1.2 mM EDTA, 0,5% deoxycholic acid) containing a 1% protease inhibitor cocktail (Sigma-Aldrich, P2714), and protein concentration was determined by the Bradford method. 23 In next step, the samples were centrifuged at 20.000 × g for 90 min at 4°C. Twenty micrograms (20 μg) of total protein were electrophoretically separated in 10% and 14% SDS-polyacrylamide gel (100 V, 90 min) and then transferred onto PVDF membrane (0.45 μm pore size; GE Healthcare Life Science) in a tank blotting system (100 V, 120 min). Membranes were blocked with a 5% casein solution for 1 h and incubated overnight (4°C) under agitation with the antibodies: AβPP-C99/AβPP-C83 (1:1000, Merck, A8717), α-actin (1:3000, DSHB, 224-236-1), phospho-Tau (1:1000, Millipore, AB8672), Tom20 (1:1000, Cell Signaling, D8T4N), anti-mouse secondary (1:5000, Cell Signaling, 7076S), anti-rabbit secondary (1:5000, Cell Signaling, 7074S), tTau (1:1000, Sigma, SAB4501821), Akt (pan) (1:1000, Cell Signaling, C67E7), phosphor-Akt (1:1000, Cell Signaling, D9E); AMPK (1:1000, Cell Signaling, D63G4), phospho-AMPK (1:1000, Santa Cruz, 33524), and Oxphos (1:1000, Abcam, Ab 110413). After three washes with TBS-T, the membranes were incubated with appropriate HRP-conjugated secondary antibodies (1:5000 in 1% BSA) for 1 h at room temperature. For detection, we used a chemiluminescence reaction kit. Immunoreactive protein bands were visualized in the ChemiDoc MP System (BioRad) and analyzed by the system's software (ImageLab 5.2.1). The image result for each band was normalized against the α-actin level (internal control) of the respective sample.
Respirometry
Fresh biopsies of the bilateral hippocampus (4–7 mg wet weight) were hand dissected in cubes (∼2 × 2 × 2 mm, 4–7 mg w.w.) and permeabilized in 1 mL ice-cold BIOPS solution (50 mM K+ -MES, 20 mM taurine, 0.5 mM dithiothreitol, 6.56 mM MgCl2, 5.77 mM ATP, 15 mM phosphocreatine, 20 mM imidazole, and 10 mM Ca-EGTA buffer, pH 7.1) containing saponin (0.01%) for 10 min. Then, after 12–15 min after mice euthanasia, biopsies were placed in the Oroboros chamber (Oxygraph-2k, Oroboros, Innsbruck, Austria) containing 2.1 mL air-saturated MiR05 (0.5 mM EGTA, 3 mM MgCl2, 60 mM K-lactobionate, 20 mM taurine, 10 mM KH2PO4, 20 mM HEPES, 110 mM sucrose, 1 g/L albumin, pH 7.1) at 37°C, with 300 rpm stirring, and the O2 consumption rates were monitored. Mitochondrial respiratory States were determined as follow: non-phosphorylation (State L), in the presence of 10 mM glutamate (NADH-linked substrate, N); phosphorylation supported by N (State P-N), after addition of 1 mM ADP, or supported by N and succinate (State P-NS), after addition of 10 mM succinate; noncoupled or at maximal respiratory capacity supported by NS (State E-NS), after 2 additions of 1 µM carbonyl cyanide m-chlorophenyl hydrazone (CCCP, uncoupler), or supported by S (State E-S), after addition of 1 µM rotenone (complex I inhibitor). Lastly, 5 mM malonate and 2 mM NaCN, inhibited complexes II and IV respectively, and residual oxygen consumption (ROx) was determined. Rox rates were subtracted from every respiratory state. 30
Statistics
The results are expressed as mean ± SEM and were analyzed by two-way analysis of variance followed by Tukey's post-hoc test for multiple comparisons. Values of p ≤ 0.05 were considered statistically significant in all cases. The software used was Graph Pad Prism version 8.0 (GraphPad Software, Inc., San Diego, CA, USA, 2007).
Results
Six months of voluntary wheel running (VWR) improves aerobic capacity and the molecular marker of oxidative metabolism in soleus and cardiac muscles in middle-aged 3xTg-AD female mice
The animals were submitted to the incremental load test to verify if 6 months VWR improves in vivo aerobic capacity in middle-aged female mice. Figure 1B shows that both mice groups (WT and 3xTg-AD) with wheel access (exercised, VWR) presented higher exhaustion velocity compared to their respective pairs without wheel access (control condition, considered “sedentary” for comparative means, SED), in which the 3xTg-AD exercised mice (VWR) demonstrated the highest significant level. In addition, muscle oxidative capacity in these mice was checked by measuring citrate synthase activity, a classical biomarker of mitochondrial content. 31 Hearts (Figure 1C) and Soleus (Figure 1D) muscles of 3xTg-AD exercised mice presented higher CS activity levels than 3xTg-AD under sedentary conditions.
Six months of VWR reduces the basal but not active energy expenditure in middle-aged female mice
These animals were submitted to the indirect calorimetry to verify if 6 months VWR changes body metabolic parameters in middle-aged female mice. Figure 2 shows that during the active period of the day (12 h dark) no differences in energy expenditure (EE) (Figure 2A) were observed between mice groups (WT and 3xTg-AD) with or without wheel access. However, when the mice were resting (12 h light), 3xTg-AD_SED mice presented elevated EE compared to both WT groups (Figure 2B) and VWR reduced EE in both WT and 3xTg-AD mice. Despite this, the groups of mice had similar body weights (g): WT_SED = 27.94 ± 0.87; WT_VWR = 27.83 ± 1.46; 3xTg-AD_SED = 28.5 ± 0.97 and 3xTg-AD_VWR = 29.3 ± 0.54 (p > 0.05). Respiratory quotient (RQ) was similar between all mice groups under resting or active periods (Figures 2C, D), indicating no changes in energy substrate oxidation.

Six months VWR reduces body metabolic rates in the resting period of the day in middle-aged female mice. (A) Energy expenditure (EE) during the 12 h dark. (B) EE during the 12 h light; a = p < 0.05 versus WT_VWR; b = p < 0.05 versus 3xTg-AD_VWR; c = p < 0.05 versus WT_SED (Two-way ANOVA, Tukey's post hoc). C) Respiratory quotient (RQ) index during the 12 h light. (D) RQ during the 12 h dark. Data are expressed as the mean ± SEM; N = 9 (WT_SED), 9 (WT_VWR), 7 (3xTg-AD_SED), and 6 (3xTg-AD_VWR).
Six months of VWR recovers discriminative short-term and non-associative long-term memory in middle-aged 3xTg-AD female mice
After finding evidence that VWR has improved parameters of aerobic capacity and metabolism in 3xTg-AD middle-aged female mice, we tested if 6 months of VWR improved discriminative memory and performance related to retention of habituation, submitting the mouse groups to new object recognition (NOR, Figure 3A-C) and open field (OF, Figures 3D, E) tasks, respectively. Figure 3A presents the similar exploratory indexes (%) in training session of all mice groups. As expected, 3xTg-AD mice submitted to control condition (3xTg-AD_SED) presented lower exploratory index (Figure 3B) and time spent (Figure 3C) on NOR compared to control WT animals, demonstrating an expressive impairment of short-term discriminative memory. WT animals (SED and VWR) also showed a common characteristic of the normal adaptive learning process of habituation in the OF arena, in which the number of rearing (Figure 3D) and total distance traveled (Figure 3E) during the Test session is smaller than Training session. However, 3xTg-AD_SED presented an opposite profile, demonstrating a significant deficit of habituation memory. All these memories and learning parameters appeared fully recovered in 3xTg-AD mice with wheel access. Despite this, the VWR did not reverse the decreased total distance exploration observed in 3xTg-AD, a behavior associated with increased innate anxiety. 32
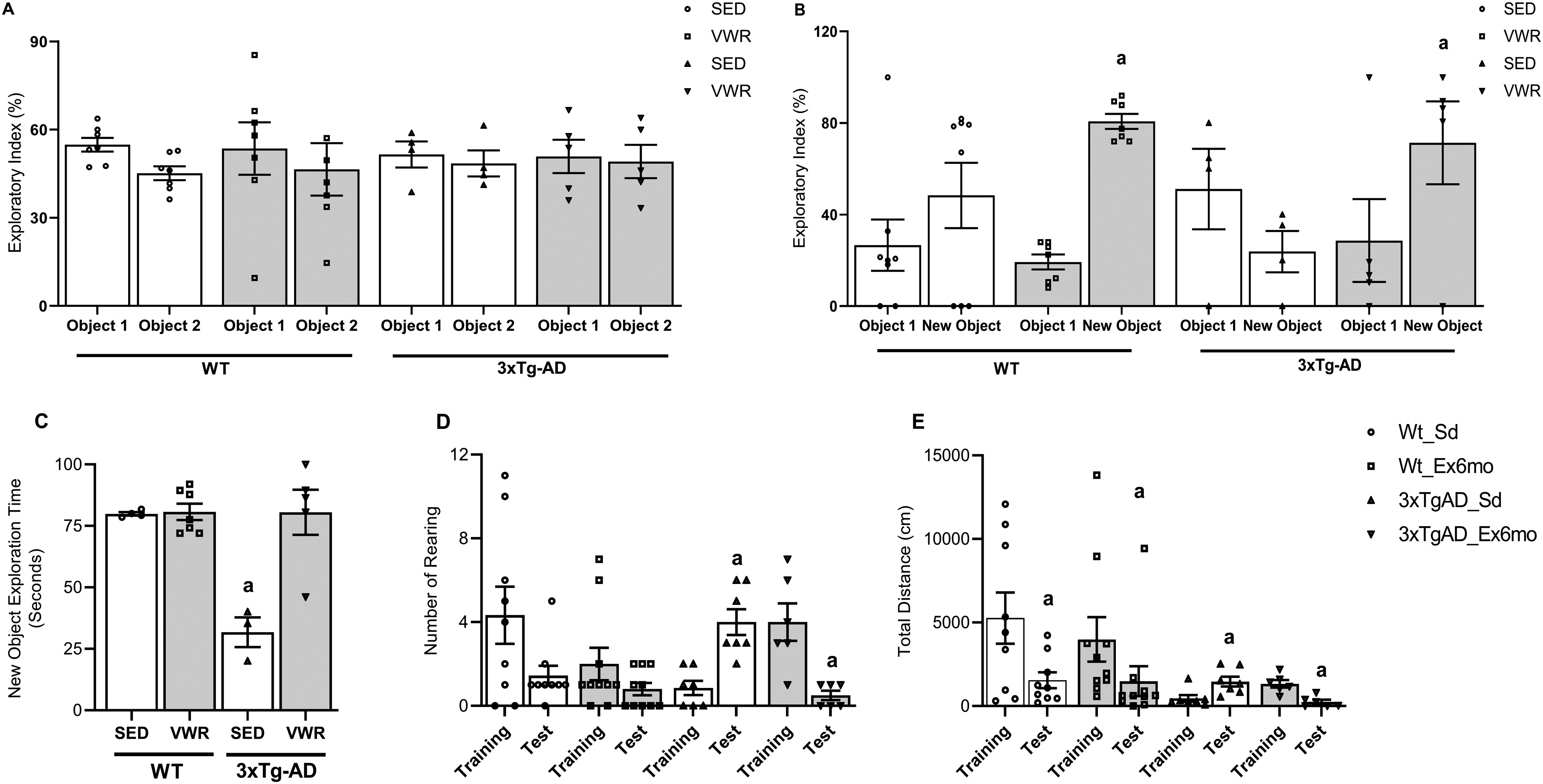
Six months of VWR improves short-term memory and learning habituation of middle-aged 3xTg-AD female mice. (A) Percentual of exploratory behavior in the initial familiarization session in the novel object recognition (NOR). (B) Percentual of exploratory behavior of the new object in the test session in NOR; a = p < 0.05 versus object 1 (familiar) of the same group (unpaired Student's t-test). (C) Time expenditure in NOR; a = p < 0.05 versus WT_SED, WT_VWR, and 3xTg-AD_VWR (Two-way ANOVA, Tukey's post hoc). (D) Number of rearing in open field (OF) arena; a = p < 0.05 versus training session of the same group (unpaired Student's t-test). (E) Total distance traveled in OF arena; a = p < 0.05 versus training session of the same group (unpaired Student's t-test). Data are expressed as the mean ± SEM; N = 8 (WT_SED), 7 (WT_VWR), 6 (3xTg-AD_SED), and 6 (3xTg-AD_VWR).
Six months of VWR reverts tau phosphorylation in hippocampus in middle-aged 3xTg-AD female mice
After finding evidence that VWR has improved parameters of metabolism and memory in middle-aged 3xTg-AD female mice, next we verify in these animals if 6 months of VWR alters the levels of Tau phosphorylation (p-Tau), a widely considered pathological hallmark in AD.33,34 The analyses of Tau by western blotting (Figure 4) showed that VWR reduced the elevated levels of p-Tau relative to total Tau (t-Tau) (Figure 4B) or α-actin (Figure 4C) in hippocampus of 3xTg-AD mice, to the levels of WT mice (SED or VWR). Notably, the cortex of middle-aged 3xTg-AD female mice did not present significant p-Tau levels compared to WT, and wheel access exerts no effect (Figure 4D, E).

Six months VWR reduces the levels of tau phosphorylation in hippocampus of middle-aged 3xTg-AD female mice. (A) Western blotting representative image showing anti-Tau phospho (pTau), anti-Tau (total Tau, tTau) or anti-actin stainings in hippocampus of WT and 3xTg-AD sedentary (SED) or voluntary wheel running (VWR). (B) pTau/tTau band ratio in hippocampus; a = p < 0.05 versus WT_SED, WT_VWR, 3xTg-AD_VWR; (C) Band intensities of pTau relative to α-actin in hippocampus; a = p < 0.05 versus 3xTg-AD_SED, 3xTg-AD_VWR; (D) Western blotting representative image showing anti-pTau, anti-tTau or anti-actin stainings in cortex. (E) pTau/tTau band ratio in cortex; (F) Band intensities of pTau relative to α-actin in cortex. Two-way ANOVA, Tukey's post hoc. Data are expressed as the mean ± SEM; N = 8 (WT_SED), 7 (WT_VWR), 6 (3xTg-AD_SED), and 6 (3xTg-AD_VWR).
Six months of VWR reduces the accumulation of amyloidogenic aβpp-c99 fragments in hippocampus in middle-aged 3xTg-AD female mice
In line with Tau phosphorylation, next we verify if 6 months VWR alters the amyloid cascade of AD in these animals. The starting point is the AβPP cleavage by BACE1, producing a 99-residue C-terminal peptide (AβPP-C99), that is subsequently cleaved to Aβ protein by γ-secretase. 35 As expected, the analyses of AβPP-C99 fragments by western blotting demonstrated that the hippocampus (Figure 5A, B) and cortex (Figure 5E, F) of middle-aged 3xTg-AD female mice have ∼ 4- and 2-fold higher AβPP-C99 content than WT mice under control conditions (SED), respectively. Despite the non-amyloidogenic AβPP-C83 fragments, generated from AβPP cleavage by α-secretase, also elevated in 3xTg-AD animals (Figures 5A, C, E, G), the higher AβPP-C99/C83 ratios (Figures 5D, H) indicates a more intense amyloidogenic pathway in hippocampus and cortex of these animals. Surprisingly, middle-aged 3xTg-AD female mice with access to a running wheel presented a reduction in AβPP-C99 and AβPP-C83 content, and a significant reduction in AβPP-C99/C83 ratio only in hippocampus; in cortex, VWR reduced AβPP-C83 fragments but not the AβPP-C99/C83 ratio.
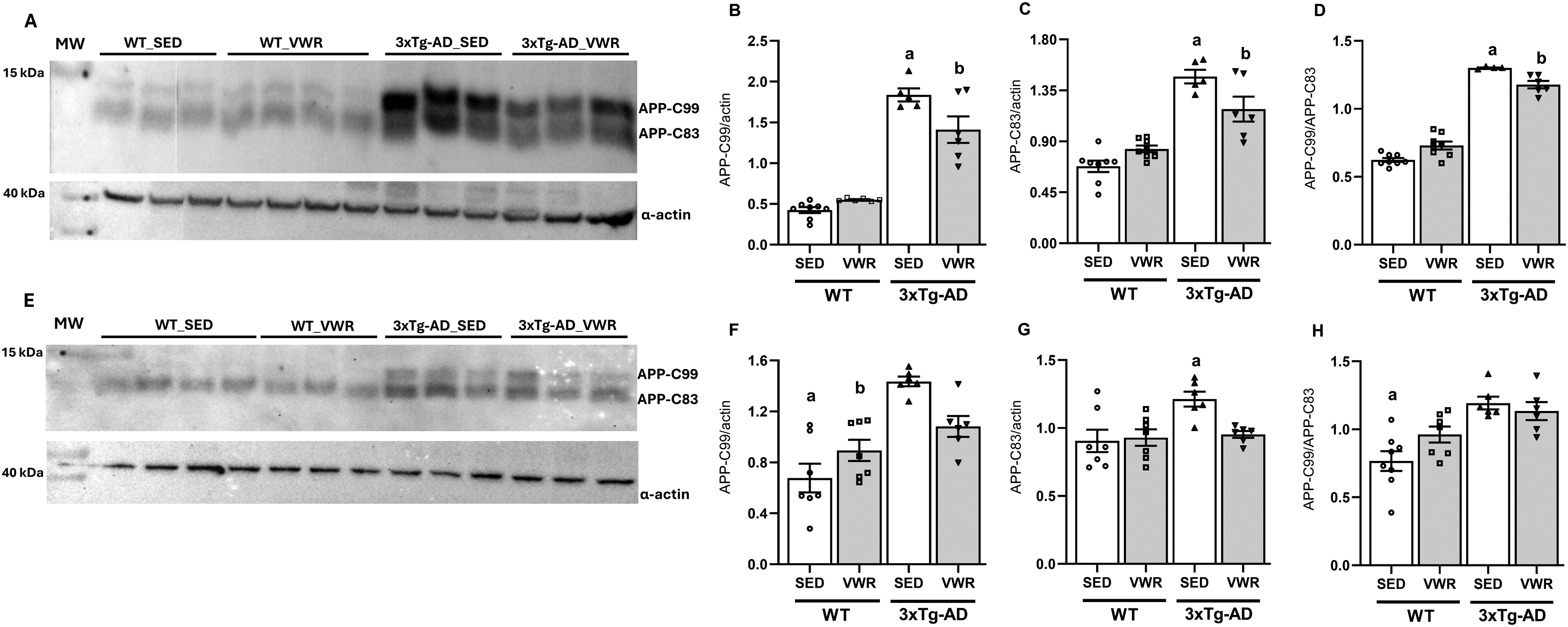
Six months of VWR reduces the levels of APP-C99 and APP-C83 fragments in hippocampus and cortex of middle-aged 3xTg-AD female mice. (A) Western blot representative image showing anti-APP (APP-C99 or APP-C83 fragments) and anti-actin stainings in hippocampus of WT and 3xTg-AD sedentary (SED) or voluntary wheel running (VWR). (B) Band intensities of APP-C99 band relative to α-actin in hippocampus; a = p < 0.05 versus WT_SED, WT_VWR, 3xTg-AD_VWR; b = p < 0.05 versus WT_SED, WT_VWR. (C) Band intensities of APP-C83 relative to α-actin in hippocampus; a = p < 0.05 versus WT_SED, WT_VWR, 3xTg-AD_VWR; b = p < 0.05 versus WT_SED, WT_VWR; (D) APP-C99/APP-C83 band ratio in hippocampus; a = p < 0.05 versus WT_SED, WT_VWR, 3xTg-AD_VWR; b = p < 0.05 versus WT_SED, WT_VWR. (E) Western blot representative image showing APP-C99, APP-C83 and α-actin expression levels in cortex. (F) Band intensities of APP-C99 relative to α-actin in cortex; a = p < 0.05 versus 3xTg-AD_SED, 3xTg-AD_VWR; b = p < 0.05 versus 3xTg-AD_SED. (G) Band intensities of APP-C83 relative to α-actin in cortex; a = p < 0.05 versus WT_SED, WT_VWR, 3xTg-AD_VWR; (H) APP-C99/APP-C83 band ratio in cortex; a = p < 0.05 versus 3xTg-AD_SED, 3xTg-AD_VWR; Two-way ANOVA, Tukey's post hoc. Data are expressed as the mean ± SEM; N = 8 (WT_SED), 7 (WT_VWR), 6 (3xTg-AD_SED), and 6 (3xTg-AD_VWR).
VWR reduces the activation of insulin signaling intermediary Akt in hippocampus of middle-aged 3xTg-AD female mice
After finding evidence that VWR improved parameters of metabolism, memory, and markers of AD in middle-aged 3xTg-AD female mice, we next investigated whether 6 months of VWR changes the activation levels of metabolic sensors AMP-activated protein kinase (AMPK) and protein kinase B (Akt) in hippocampus. Both kinases play several vital roles in neuronal metabolism and function: AMPK is a sensor of low ATP concentration and regulates essential metabolic pathways, such as glycolysis and mitochondrial biogenesis,36,37 and cellular processes of autophagy, ion flow and neuronal excitability; 38 while Akt regulates glucose uptake and utilization, as well as protein synthesis, neurotransmitter signaling and synaptic plasticity. 39 The levels of phosphorylated-AMPK (pAMPK, Figure 6A, B) and phosphorylated-Akt (pAkt Ser473, Figure 6C, D) in hippocampus of middle-aged 3xTg-AD female mice were similar compared to WT, indicating the similar levels of activations at sedentary conditions. However, in mice with wheel access (VWR) the levels of p-Akt were decreased while pAMPK were not changed, compared to animals without wheel access (SED). These results indicate that the reduction in AKT activation in the hippocampus induced by VWR was unrelated to intracellular energetic changes.

Six months of VWR reduces the activation of insulin signaling intermediary Akt in hippocampus of middle-aged female mice. (A) Western blotting representative image showing anti-AMPK phospho (p-AMPK) or anti-actin stainings in hippocampus of WT and 3xTg-AD sedentary (SED) or voluntary wheel running (VWR). (B) Band intensities of p-AMPK relative to α-actin in hippocampus; (C) Western blotting representative image showing anti-Akt phospho (p-Akt Ser473) or anti-actin staining in cortex. (D) Band intensities of p-Akt relative to α-actin in cortex; a = p < 0.05 versus WT_SED, 3xTg_AD_SED; Data are expressed as the mean ± SEM; Two-way ANOVA, Tukey's post hoc. N = 8 (WT_SED), 7 (WT_VWR), 6 (3xTg-AD_SED), and 6 (3xTg-AD_VWR).
Six months of VWR reversed the higher mitochondrial coupling degree in hippocampus in middle-aged 3xTg-AD female mice
As mitochondrial dysfunction is a common finding in several in vivo and in vitro models of AD, 40 next we investigated if VWR modulated energetic mitochondrial function in middle-aged 3xTg-AD female mice. Respiratory rates were monitored by HRR in permeabilized fresh biopsies of hippocampus, a technique that maintain the native cellular environment and interactions (mix of cell types and extracellular matrix), providing a more physiologically relevant assessment of mitochondrial function, reflecting the in vivo situation more accurately. Respiratory rates in the states of non-phosphorylation (State L, without ADP), phosphorylation (State P, after ADP addition) or at maximal respiratory capacity (State E, after CCCP uncoupler addition), in the presence of NADH-linked (or complex I) substrate (N, glutamate) alone or with complex II substrate (S, succinate) (Figure 7A). Respiratory rates at State P were similar between all mice groups, independently of the respiratory substrate (Figures 7C, D). However, State L respiration was lower in 3xTg-AD_SED (Figure 7B), leading to an increased P/L ratio (with substrates N, for complex I) (Figure 7H) compared to WT_SED, which indicates a higher coupling degree in these mitochondria. Respiratory rates at State E presented no differences between all mice groups when sustained by substrates for both complexes (NS, Figure 7E), but after complex I inhibition by rotenone, State E sustained by succinate (S) appeared elevated in 3xTg-AD_SED (Figure 7F). Still, in these animals the lower rates at NS-S pathway control efficiency (State E: NS - S), Figure 7G) demonstrate a low contribution of complex I relative to complex II in respiration, indicating a relative complex I inhibition in these mitochondria. The limitation of OXPHOS capacity by the phosphorylation system, expressed by the P/E control ratio, was similar between all mice groups (Figure 7I). The wheel access (VWR) reversed all mitochondrial changes found in hippocampus of 3xTg-AD animals, and in WT animals reduced State L respiration, without changes in coupling degree (P/L ratio).

Six months of VWR reversed the altered mitochondrial respiration in hippocampus of middle-aged 3xTg-AD female mice. (A) Representative figure of respiratory monitoring by HRR. (B) Respiratory rates at State L (Leak) supported by complex I substrates glutamate 10 mM (N); a = p < 0.05 versus WT_VWR, 3xTg-AD_SED; (C) Respiratory rates at State P (oxidative phosphorylation) supported by NADH-linked substrates (N). (D) Respiratory rates at State P supported by N and 10 mM succinate (NS). (E) Respiratory rates at State E (maximal electron transfer) supported by N and 10 mM succinate (NS). (F) Respiratory rates at State E supported by succinate in the presence of 2 μM rotenone (S); a = p < 0.05 versus WT_VWR, 3xTg-AD_VWR; (G) NS-S pathway control efficiency (State E: NS - S); a = p < 0.05 versus WT_SED; WT_VWR, 3xTg-AD_SED; (H) P/L control ratio. a = p < 0.05 versus WT_VWR, 3xTg-AD_VWR; (I) P/E control ratio. Two-way ANOVA, Tukey's post hoc. Data are expressed as the mean ± SEM; N = 8 (WT_SED), 7 (WT_VWR), 6 (3xTg-AD_SED) and 6 (3xTg-AD_VWR).
In addition, the content of mitochondrial proteins was analyzed by western blotting. Hippocampus of middle-aged 3xTg-AD female mice did not present significant differences in the contents of Tom20, a marker of mitochondrial mass, or RC components (from Complex I to IV) compared to WT under sedentary conditions. The wheel access seemed to exert antagonistic effects on mitochondrial content in each strain, as seen by the significantly high Tom20 in WT_VWR compared to 3xTg-AD_VWR mice (Figure 8).
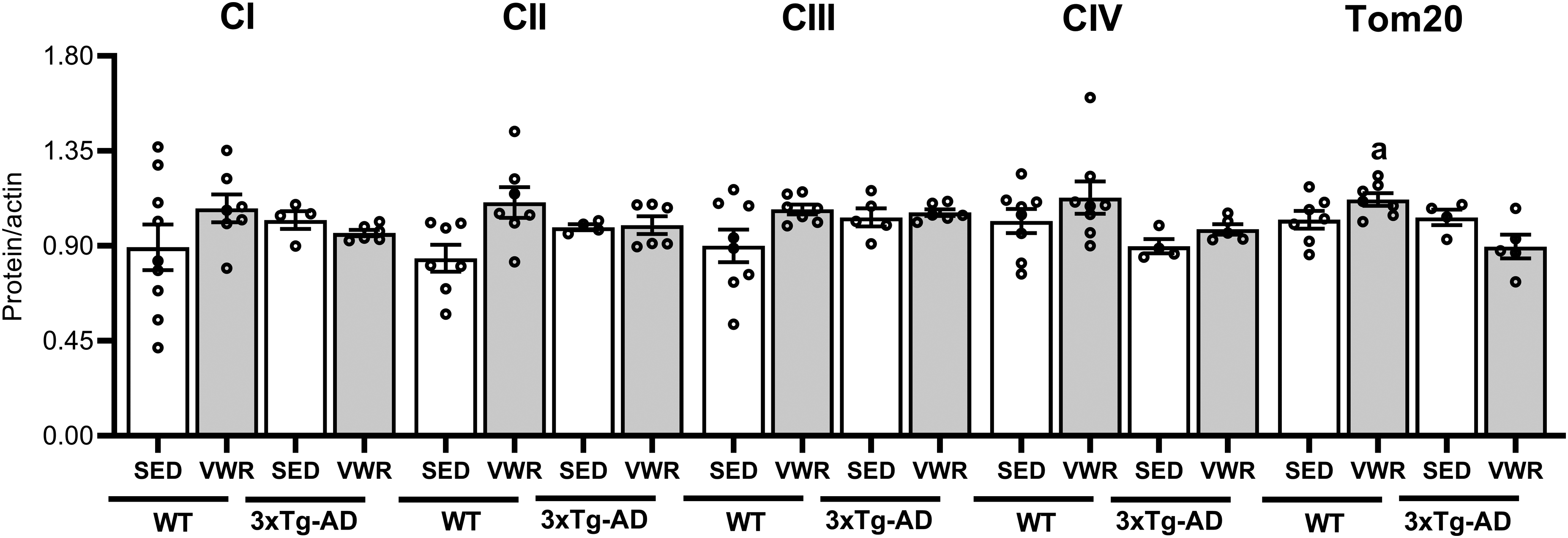
Six months of VWR exerts antagonistic effects on mitochondrial content in hippocampus of middle age wt and 3xTg-AD female mice. Quantification by western blotting of mitochondrial proteins of respiratory chain complexes (CI, CII, CIII and CIV) and Tom20 normalized by α-actin. WT and 3xTg-AD sedentary (SED) or voluntary wheel running (VWR). a = p < 0.05 versus 3xTg-AD_VWR. Data are expressed as the mean ± SEM; Two-way ANOVA, Tukey's post hoc. N = 8 (WT_SED), 7 (WT_VWR), 6 (3xTg-AD_SED), and 6 (3xTg-AD_VWR).
Discussion
This study aimed to investigate whether 6 months of voluntary exercise induces neuroprotective effects in the pathogenesis of AD in 8-month-old female mice, focusing on energy metabolism. Together, the results demonstrate that VWR improved the parameters of body and muscle metabolisms, memory and learning, biochemical markers of AD (p-Tau and AβPP-C99), modulated Akt activation and mitochondrial coupling degree in hippocampus of middle-aged 3xTg-AD female mice. These beneficial effects of VWR were observed during a stage of significant cognitive decline in the 3xTg-AD female mice, which is first apparent at 6 months and worsens significantly with aging. 41
Our results shown that the expressive impairments in discriminative and the non-associative retention memories, evaluated by NOR and OF tasks, respectively, were fully recovered in middle-aged 3xTg-AD mice that performed 6 months of wheel running. The protective effects of exercise against cognitive decline have been demonstrated in various training protocols and AD models.18,19,42 For example, in young (7-week-old) AβPP/PS1 transgenic mice submitted to 9 weeks of mandatory treadmill running (5 days/week, 20–60 min/day at 10 m.min−1, with increments of 10 min per day), the exercise increased the learning and memory performances associated with amygdala and hippocampus, assessed by Pavlovian fear conditioning task. 43 Long-term voluntary running (4 months) in different periods of daily access to a running wheel (0, 1, 3, and 12-h), beginning in 14-month-old Tg2576 mice, was able to reduce behavioral dysfunction, evaluated by OF and Y-maze, with a better response with 3-h daily exercise. 18 Nevertheless, in male AβPP/PS1 mice 4 months of voluntary exercise improved cognitive function in young (9-weeks old) but not in middle-aged (6-month old) animals, analyzed by Morris water maze test. 44 Possibly, this different result may be related to sex differences in underlying mechanisms, with females responding better to exercise. 45 In addition to aerobic exercise, 9-month-old 3xTg mice underwent a resistance training protocol and showed improved cognitive performance assessed by the NOR and Y-maze tests. 46 Other studies, including resistance exercise, have shown neuroprotective effects related to a decrease in amyloid load and plaques, an increase in neurotrophic factors, and an improvement in synaptic transmission.47–49 Thereby, the protective effects of exercise are still up for debate, but seem to be dependent on the exercise program, including regime (voluntary versus mandatory), type, start time, volume and intensity, type of AD model, and molecular condition of the disease, as well as the cognitive task used to evaluate.
Cognitive improvements in the context of AD are frequently associated to reductions of pathological hallmarks, such as Tau phosphorylation.50–53 Such effects induced by exercise in 3xTg-AD mice were previously found in adult (4-months old) and aged (24-months old) mice, submitted to 3 months of mandatory treadmill running (5 days/week, 5 min at 5 m.min−1, 20 min at 10 m.min−1, 5 min at 5 m.min−1). 19 Here, in addition to diminished Tau phosphorylation, we show for the first-time reduced levels of amyloidogenic AβPP-C99 fragments in hippocampus of middle-aged 3xTg-AD female mice that performed voluntary wheel running. A known feature of this AD model is the overproduction of AβPP-C99, driven by expression of the mutant human AβPP (AβPPSwe - Swedish double mutation; K670N, M671L), 54 a mutation occurring at the amino terminal of Aβ that strongly increases BACE1 affinity for AβPP. 55 In addition to AβPP-C99, non-amyloidogenic AβPP-C83 fragments were diminished in 3xTg-AD VWR, indicating that BACE1 and α-secretase cleavages were reduced. However, the diminished AβPP-C99/C83 ratio in 3xTg-AD mice engaging in voluntary wheel running suggests that the amyloidogenic pathway undergoes a more pronounced decrease than the non-amyloidogenic pathway. This shift promoted by VWR was not found in the cortex of these animals, highlighting the potential protective impact of physical exercise specific in the hippocampus, at the initial pathogenesis of AD. A reduction in the AβPP-C99/C83 ratio was also found in the streptozotocin (STZ)-induced AD rat model after 4 weeks of treadmill exercise (5 days/week, 30 min/day). 56
AβPP processing products can enter the cell and, even as pTau, accumulate in mitochondria via a mitochondrial import mechanism, triggering mitochondrial dysfunction; in turn, mitochondrial function and bioenergetic intermediates, such as reactive oxygen species (ROS), and NAD+/NADH, can modulate AβPP processing and trafficking pathways (for review).15,57 Our results showed that VWR could hamper this feedback loop, by reversing the mitochondrial complex I inhibition and higher coupling degree found in the hippocampus in middle-aged 3xTg-AD female mice, and reducing AD pathological hallmarks. Substantial data support a role for pTau, AβPP, and Aβ in modulating mitochondrial function in several AD models,58–62 inducing respiratory impairment by directly influencing the activities of RC complexes, such as complex I and III, 63 also leading to increasing ROS generation 60 specifically in hippocampus of 3xTg-AD mice, Aβ accumulation, and mitochondrial respiratory dysfunction start early (3 months old) and increase in an age-dependent way 61 Aβ and pTau interacts with VDAC1, as found in the brains from AD patients and AD models, in which is observed mitochondrial dysfunction.58,63 In the context of reduced complex I activity, the higher coupling degree found in hippocampus of middle aged 3xTg-AD female mice can be suggested as an adaptive mechanism to improve efficiency of oxidative phosphorylation, since the metabolic sensor AMPK did not show any changes in cell energy levels (AMP/ATP ratio). However, a high coupling degree linked to slow leak respiration is associated with greater ROS formation, since under these conditions the mitochondrial membrane potential is high and electron flow rate is low, increasing the probability of an electron reducing oxygen to ROS before it reaches complex IV. 64
AMPK signaling is an essential modulator of mitochondrial biogenesis and metabolism, 37 and its role in AD is intriguing. 65 AMPK activation by pharmacological or dietary interventions has reduced66–68 or increased.69,70,13,71 AD hallmarks, mainly BACE1 mRNA and Aβ levels. However, the improvements in AD markers in middle aged 3xTg-AD mice promoted by VWR demonstrated here were not associated with activating this kinase (pAMPK) in the hippocampus. On the other hand, the high energy metabolic sensor Akt 72 was decreased in the hippocampus of mice with wheel access, characterizing a specific effect of VWR. In AD, reductions in Akt activation have been shown to promote beneficial effects, such as protecting against Aβ-induced toxicity in mutant PDK1K465E/K465E neurons 73 and increasing α-secretase AβPP cleavage. 74 Taken together, physical exercise's metabolic regulations involved in neuroprotection is still debatable, but it appears to involve a finely-tuned balance between catabolic and anabolic pathways, with a pronounced tendency towards a low-energy state condition.
In summary, the results presented here demonstrated that long-term voluntary wheel running improved phenotypic characteristics of AD (response in memory tasks), reduced amyloidogenic pathway (AβPP-C99) and molecular marker of AD (pTau), and restored mitochondrial metabolism in hippocampus of middle-aged female mice, genetic model of AD. These effects could be mediated by reductions in Akt activation (pAkt) unrelated to changes in the intracellular energy levels (pAMPK). Then, the present work reinforces the neuroprotective role of physical exercise and mitochondrial involvement in the etiology of AD.
Footnotes
Acknowledgments
The authors are deeply thankful to Prof. Dr Germán G. Sgro for their insightful advice and assistance throughout the course of Immunoblotting Technique, and Alcides Pereira for technical assistance.
Author contributions
Jonathas Rodrigo dos Santos (Conceptualization; Data curation; Investigation; Methodology; Writing – original draft); Carlos Rocha Catalão (Formal analysis; Methodology; Writing – review & editing); Anderson Vulczak (Methodology); Ana Elisa Azzolini (Data curation; Methodology; Supervision); Luciane Carla Alberici (Conceptualization; Data curation; Funding acquisition; Writing – original draft; Writing – review & editing).
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work received financial support from the São Paulo Research Foundation (FAPESP—2018/10089-2 to L.C.A) and Coordination for the Improvement of Higher Education Personnel - Brazil (CAPES) - Finance Code 001. LCA has a National Council for Scientific and Technological Development (CNPq) Research Fellowship.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data availability
The data supporting the findings of this study are available on request from the corresponding author. The data are not publicly available due to privacy or ethical restrictions.