
Abstract
Background
Alzheimer's disease (AD) is a common neurodegenerative disease, where neuroinflammation significantly influences its pathophysiology by driving the disease's pathological cascade. As a pro-inflammatory regulator, miR-125b-5p contributes to AD progression, though its precise role and mechanisms remain unclear.
Objective
We aims to identify mRNAs significantly regulated by pro-inflammatory miR-125b-5p in AD and uncover key neuroinflammatory pathways.
Methods
Target mRNAs regulated by miR-125b-5p were predicted using online databases and analyzed with two mRNA datasets to identify differentially expressed mRNAs (DEmRNAs). Enrichment analysis was conducted to explore their biological functions and pathways. The significance of DEmRNAs expression in AD-related inflammatory pathways was verified by the Wilcoxon test, predictive accuracy was assessed via area under the curves (AUCs), and novel mRNAs were identified through positive control analysis.
Results
A total of 613 miR-125b-5p target mRNAs were identified, and 44 DEmRNAs were detected to be regulated by miR-125b-5p in two datasets. The 44 target DEmRNAs associated with AD include three key pathways: insulin signaling (EXOC7, FLOT2, MKNK2), phosphatidylinositol signaling (IP6K1, MTMR3), and phospholipase D signaling (CYTH1, GAB2). Correlation analysis indicated strong correlations among 7 mRNAs, all showing significant differential expression, with AUCs above 0.5, confirming their predictive value. Three mRNAs (EXOC7, IP6K1, CYTH1) were identified as novel AD-related genes. MiR-125b-5p binding sites in the 3′-UTRs of these 7 mRNAs suggest their potential roles in AD-related inflammation and signaling pathways.
Conclusions
This study investigates the pro-inflammatory miR-125b-5p's role in the pathological processes of AD, highlighting its regulation of key target mRNAs and critical pathways.
Keywords
Introduction
Alzheimer's disease (AD), is a common progressive degenerative disease that affects the brain, resulting in impairment of behavior and cognition function.1,2 The incidence of AD is highest among the older adult. With the aging of the global population, the increasing prevalence of AD poses a great challenge to modern society, including social care, emotional toll on caregivers, financial impact for healthcare services, and has become a significant burden on global public and medical systems. 3 Pathological features of AD patients include intracellular neurofibrillary tangles (NFTs), consisting of the hyperphosphorylated microtubule-associated protein tau, and extracellular amyloid plaques, formed by the amyloid-β (Aβ) peptides. 4 But clinical advances based on these two characteristics, such as pathological research, disease treatment and drug development, have remained slow over the past few decades. Evidence suggests that neuroinflammation plays a key role in AD pathophysiology in addition to Aβ plaques and NFTs, and may promote many other signaling pathways in AD pathology. 5
Whole-gene transcriptome studies have shown that dysregulation of AD-related genes is critical for inflammatory responses. As major gene regulators, microRNAs (miRNAs) can help maintain transcriptome homeostasis.6,7 miRNAs are an endogenous class of small non-coding RNAs (snRNAs), which are involved in post-transcriptional gene regulation. miRNAs play a regulatory role on target messenger RNA (mRNAs) by disrupting and inhibiting the translation of mRNAs.8–10 About 70% of the miRNAs identified so far are expressed in the human brain, 11 and they are also stable in plasma and serum. 12 miRNAs can regulate gene silencing, and their target genes can participate in the pathophysiology of AD, such as PSEN1, PSEN2, BACE1, and APP. 12 Some studies have shown that abnormal regulation and alteration of specific miRNA may lead to the pathogenesis of AD.13,14 One study has observed that miR-125b, one of the plasma miRNAs, may be a useful marker in differentiating those affected by AD (from those who are not). 15 There is an existing base of research on miR-125b, which poses various theories on the role of miR-125b in the pathogenesis of AD. Overexpression of miR-125b has been shown to increase tau hyperphosphorylation and accelerate apoptosis in primary hippocampal neurons by negatively regulating p53. Although p53 is typically known for promoting apoptosis, in this context, its downregulation by miR-125b disrupts its protective role in preventing AD lesion formation, leading to enhanced neuronal apoptosis.16,17 Thus, subsequent effects such as impaired learning and memory function have been observed in mice studies.18,19 Similarly, in a study of APP/PS1 mice, Hong et al. found a positive association between miR-125b with cognitive decline. 20 Another clinical study found that miR-125b is upregulated in the brain of AD patients, whereas it is downregulated in serum. 21 Anyway, these observations seem to suggest miR-125b could be a potential biomarker of disease.
The other main evidence base suggests miR-125b may contribute to AD pathogenesis and progression via acting as a pro-inflammatory factor. Many studies have shown that miR-125b is involved in the progression of AD disease and can act as an important regulator of inflammatory processes. 22 Alexandrov et al. quickly analyzed the postmortem brains of AD patients, with the results indicating that miRNA-125b was not only significantly elevated in the brain tissue, extracellular fluid (ECF) and cerebrospinal fluid (CSF) of AD patients, but also regulated by pro-inflammatory cytokines. 23 Also, miR-125b is consistently upregulated under the transcriptional control of pro-inflammatory transcription factor NF-κB. The genetic regulatory information carried by miR-125b can be stably expressed in the peripheral blood through the blood-brain barrier (BBB).22,24 Another study observed that upregulated miR-125b in the human AD brain may lead to glial cell proliferation and non-homeostasis activation of inflammatory signals, aggravating neuroinflammation, and direct inhibition of miR-125b may reduce the effect of proinflammatory cytokines. 25 Additionally, miR-125b can act alone or synergistically with other pro-inflammatory miRNAs to stimulate inflammatory responses in AD pathology. Clinical data from human AD brains suggest that elevated miR-125b levels increase neuroinflammation and reduce complement factor H (CFH), which is associated with miR-155 neuronal release and the spread of inflammation in AD brains. 26 However, our purpose in this study was to screen out the target miR-125b-5p regulated AD candidate markers, not to explore the complete miRNA regulatory mechanism in the disease.
MiR-125b-5p is the mature, biologically active form of miR-125b. Specifically, miR-125b is a well-studied member of miRNAs family, initially transcribed as a long primary miRNA (pri-miR-125b). This precursor undergoes processing to form a hairpin-shaped precursor miRNA (pre-miR-125b) in the nucleus. Subsequent processing by the Dicer enzyme in the cytoplasm yields mature miRNAs. Among the mature forms, miR-125b-5p is derived from the 5′ arm of the pre-miRNA hairpin and is the biologically active form that participates in gene silencing. In contrast, the 3′ arm generates the miR-125b-3p form, which may have different target genes and functions. The expression of miR-125b-5p in the brain is associated with AD pathogenesis and may also have a role in regulating inflammatory responses. For example, in a cell line experiment, Li et al. using a luciferase reporting assay demonstrated that overexpression of miR-125b-5p may reduce Aβ-induced neurotoxicity in vitro by targeting BACE1, suggesting that miR-125b-5p may inhibit AD development by decreasing the expression of target genes. 27 Bazrgar et al. used miRNA chips to analyze mice microglia and immune cells and found miR-125b-5p was specifically expressed only in microglia. This was subsequently verified in fetal and adult microglia, and the results were consistent with those initially found in mice experiment. 28 Moreover, the upregulated expression of miR-125b-5p as an inflammatory snRNA in AD disease can be retrieved from the Non-coding RNAs in Inflammation (ncRI) database (http://www.jianglab.cn/ncRI/).29 The specific information of miR-125b-5p includes disease (Alzheimer's disease), expression direction (Upregulated), detection method (Microarray), throughput (High), and species (Human). The ncRI database contains experimentally proven snRNAs in inflammatory diseases, providing a comprehensive and reliable knowledge base for snRNAs and their role in inflammatory diseases, 29 further providing more favorable evidence for the role of miR-125b-5p as a pro-inflammatory factor.
The function and specific mechanisms of pro-inflammatory miR-125b-5p in the development of AD remains unclear, but miRNAs can significantly control many targets and regulate neuroinflammatory signaling pathways. Therefore, this study has designed experimental protocols to analyze key mRNAs and signaling pathways regulated by pro-inflammatory miR-125b-5p. The overall workflow is shown in Figure 1: the target mRNAs of miR-125b-5p were predicted, and then target differentially expressed mRNAs (DEmRNAs) of miR-125b-5p were obtained by comprehensive association analysis with the blood mRNA expression profiles of AD patients. Subsequently, the potential biological function of target DEmRNAs was explored to search for the significant inflammatory pathways involved. The correlations of mRNAs enriched in the pathways were analyzed, the differential expression level and predictive ability were verified, and positive control analysis was performed, thus providing a new perspective for the pathological mechanism and treatment of AD caused by miR-125b-5p induced mRNAs and pathway changes.

The work flow of this study. The red frame indicates the ultimate purpose of this work: pro-inflammatory miR-125b-5p regulates key mRNAs and significant signaling pathways involved in the inflammatory pathological process of AD. DEmRNAs: differentially expressed mRNAs; GEO: Gene Expression Omnibus; GO: Gene Ontology; KEGG: Kyoto Encyclopedia of Genes and Genomes; PCC: Pearson correlation coefficient; ROC: receiver operating characteristic; AUC: area under the curve; AD: Alzheimer's disease.
Methods
Detection of target mRNAs of miRNA
Target mRNAs of pro-inflammatory miR-125b-5p were detected based on TargetScan database (https://www.targetscan.org/), 30 miRTarBase database (https://miRTarBase.cuhk.edu.cn/) 31 and miRDB database (https://mirdb.org/).32,33 These databases predict miRNA binding sites based on the complementarity between miRNA seed sequences and the 3′-UTR regions of mRNAs using different algorithms. During the prediction process, default parameters were applied for all databases. To enhance the accuracy of the predictions, only those target mRNAs identified in at least two databases were retained.
MRNA datasets
The GSE63060 and GSE63061 expression profiles (Experiment type: Expression profiling by array) 34 were download from the GEO database (http://www.ncbi.nlm.nih.gov/geo/). The data in the expression profiles came from blood samples taken from AD patients and control subjects. The GSE63060 expression profile included 249 samples (145 AD patients:104 control subjects), and GSE63061 expression profile included 275 samples (140 AD patients:135 control subjects). Specific information is listed in Table 1.
Specific information of the AD patients and subjects.

Analysis of the differentially expressed mRNAs (DEmRNAs) and target DEmRNAs
The R language “limma” package was used to standardize the data from AD patients and control subjects to identify significant DEmRNAs (False discovery rate, FDR < 0.05; |log2FC|>0). 35 Then the intersection of miR-125b-5p target mRNAs and DEmRNAs was detected, to obtain the target DEmRNAs regulated by miR-125b-5p. The hierarchical clustering method was used to cluster these target DEmRNAs, and the Euclidean distance method was selected as the measure of distance between samples (R language: “pheatmap” package). 36
Biological functional analysis and correlation analysis of target DEmRNAs
Gene Ontology (GO) analysis classifies genes to analyze their functions, including biological process, molecular function and cellular component. The Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis was conducted to identify the signaling pathways for gene enrichment. Target DEmRNAs were analyzed using GO (R language: “clusterProfiler” package) (p value < 0.05) 37 and KEGG pathways (R language: “pathview” package) (p value < 0.05) 38 to further understand the potential functions of genes and the signaling pathways involved. And the Pearson correlation coefficient (PCC) analysis (https://www.bioinformatics.com.cn) was used to investigate interaction relationships among DEmRNAs.
Validation of target DEmRNAs in signaling pathways
The Wilcoxon test (R language: “ggpubr” package) was used for boxplot analysis of the target DEmRNAs in the participating pathway, to detect whether the differential expression levels were significant in AD patients and controls subjects. The p value < 0.05 indicates the difference is statistically significant. 39 The predictive ability of target DEmRNAs was evaluated by a ROC curve, and the AUC was calculated by R language (“pROC” package). The AUC greater than 0.5 indicates that the predictive power of the target DEmRNAs is credible. 40
Positive control analysis
Comparing the significant and credible target DEmRNAs identified in this study with larger experimental or clinical AD genetic data could benefit better classification of both published and new identified genes. First, 75 genes were obtained from a large-scale AD related genetic study. This study is the largest to date, involving the genomes of 100,000 AD patients and 600,000 healthy people. 41 Subsequently, the existing gene databases DisGeNET (https://www.disgenet.org/home/), 42 Phenopedia (https://phgkb.cdc.gov/PHGKB/startPagePhenoPedia.action), 43 and Gene2Function (https://www.gene2function.org/search/web/) 44 were searched, which contain publicly available genes associated with AD. A total of 3397,866 and 284 AD-related genes were obtained from the three databases, respectively (Supplemental Table 3). And then integrated analysis obtained a total of 4012 genes for subsequent cross-analysis.
Results
Target mRNAs of pro-inflammatory factor mir-125b-5p
The number of target mRNAs of miR-125b-5p detected in the TargetScan, miRTarBase and miRDB databases was 931, 466 and 925, respectively. Based on the criterion that mRNAs exist in at least two databases, a total of 613 target mRNAs were detected, including 103 mRNAs in three databases, 23 mRNAs in TargetScan and miRTarBase databases, 466 mRNAs in TargetScan and miRDB databases, and 21 mRNAs in miRTarBase and miRDB databases (Figure 2(a)).
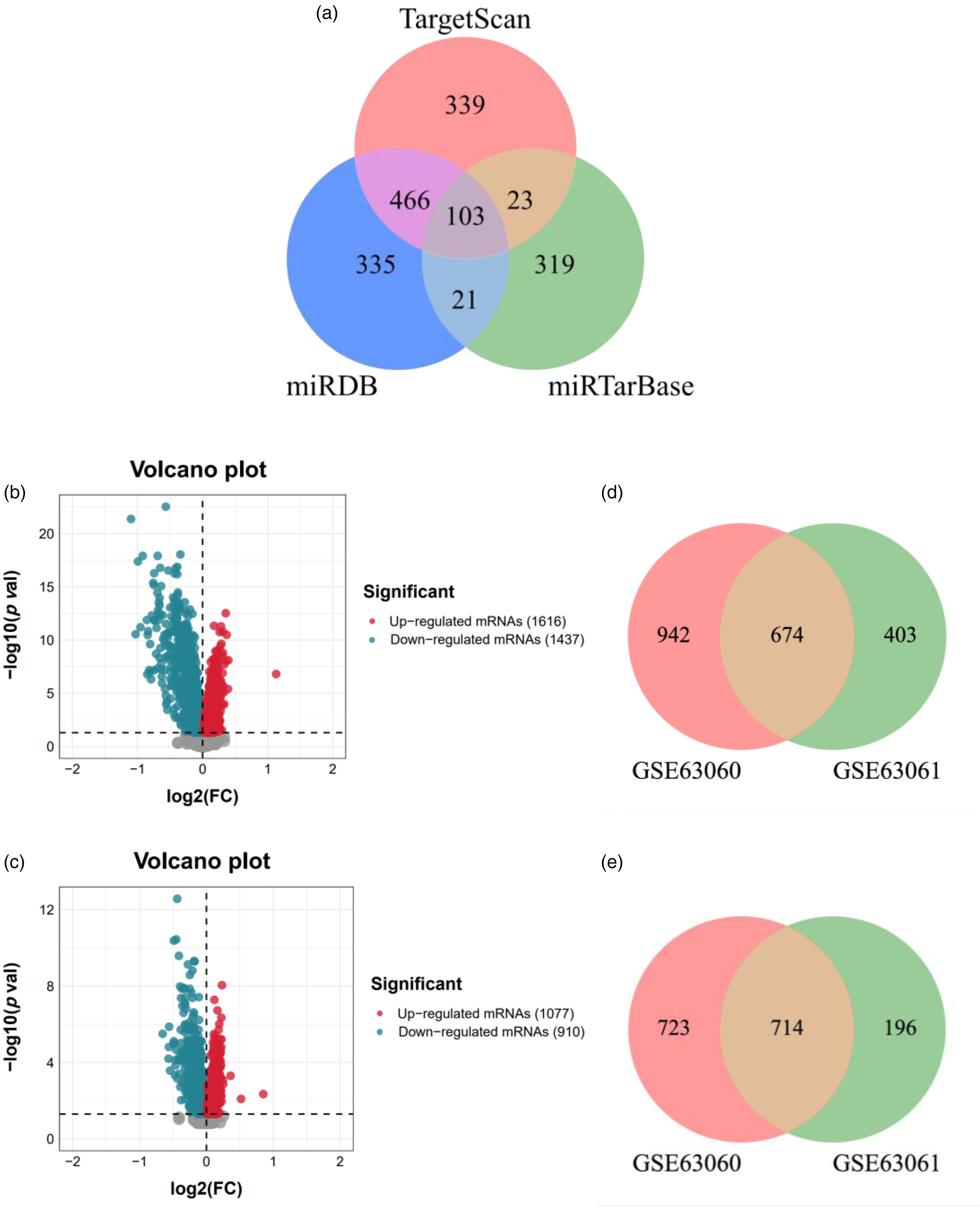
Target mRNAs of miR-125b-5p and DEmRNAs in gene expression profiles. (a) The Venn diagram of 613 target mRNAs. 103 target mRNAs (gray) exist in three databases, 23 target mRNAs (dark yellow) exist in TargetScan and miRTarBase databases, 466 target mRNAs (pink) exist in TargetScan and miRDB databases, and 21 target mRNAs (light blue) exist in miRTarBase and miRDB databases. Red circle represents the TargetScan database; Green circle represents the miRTarBase database; Blue circle represents the miRDB database. (b) Volcanic plot of DEmRNAs expression in the GSE63060 expression profile. A total of 3053 DEmRNAs, the red indicates 1616 upregulated DEmRNAs, and the blue indicates 1437 downregulated DEmRNAs. (c) Volcanic plot of DEmRNAs expression in the GSE63061 expression profile. A total of 1987 DEmRNAs, the red indicates 1077 upregulated DEmRNAs, and the blue indicates 910 downregulated DEmRNAs. (d) 674 common upregulated DEmRNAs and (e) 714 common downregulated DEmRNAs in the GSE63060 and GSE63061 expression profiles. The pink represents GSE63060 expression profile; The green represents GSE63061 expression profile.
DEmRNAs in the mRNA datasets
A total of 3053 DEmRNAs (1616 upregulated DEmRNAs and 1437 downregulated DEmRNAs) were detected in the GSE63060 expression profile, and 1987 DEmRNAs (1077 upregulated DEmRNAs and 910 downregulated DEmRNAs) were detected in the GSE63061 expression profile. Figure 2(b) and (c) show the volcano plot of all DEmRNAs in GSE63060 and GSE63061 dataset, respectively. In the GSE63060 expression profile, a total of 3053 DEmRNAs, the red indicates 1616 upregulated DEmRNAs, and the blue indicates 1437 downregulated DEmRNAs. In the GSE63061 expression profile, a total of 1987 DEmRNAs, the red indicates 1077 upregulated DEmRNAs, and the blue indicates 910 downregulated DEmRNAs. Meanwhile, 674 common upregulated DEmRNAs and 714 common downregulated DEmRNAs were detected in the two expression profile datasets (Figure 2(d) and (e)).
Target DEmRNAs regulated by mir-125b-5p
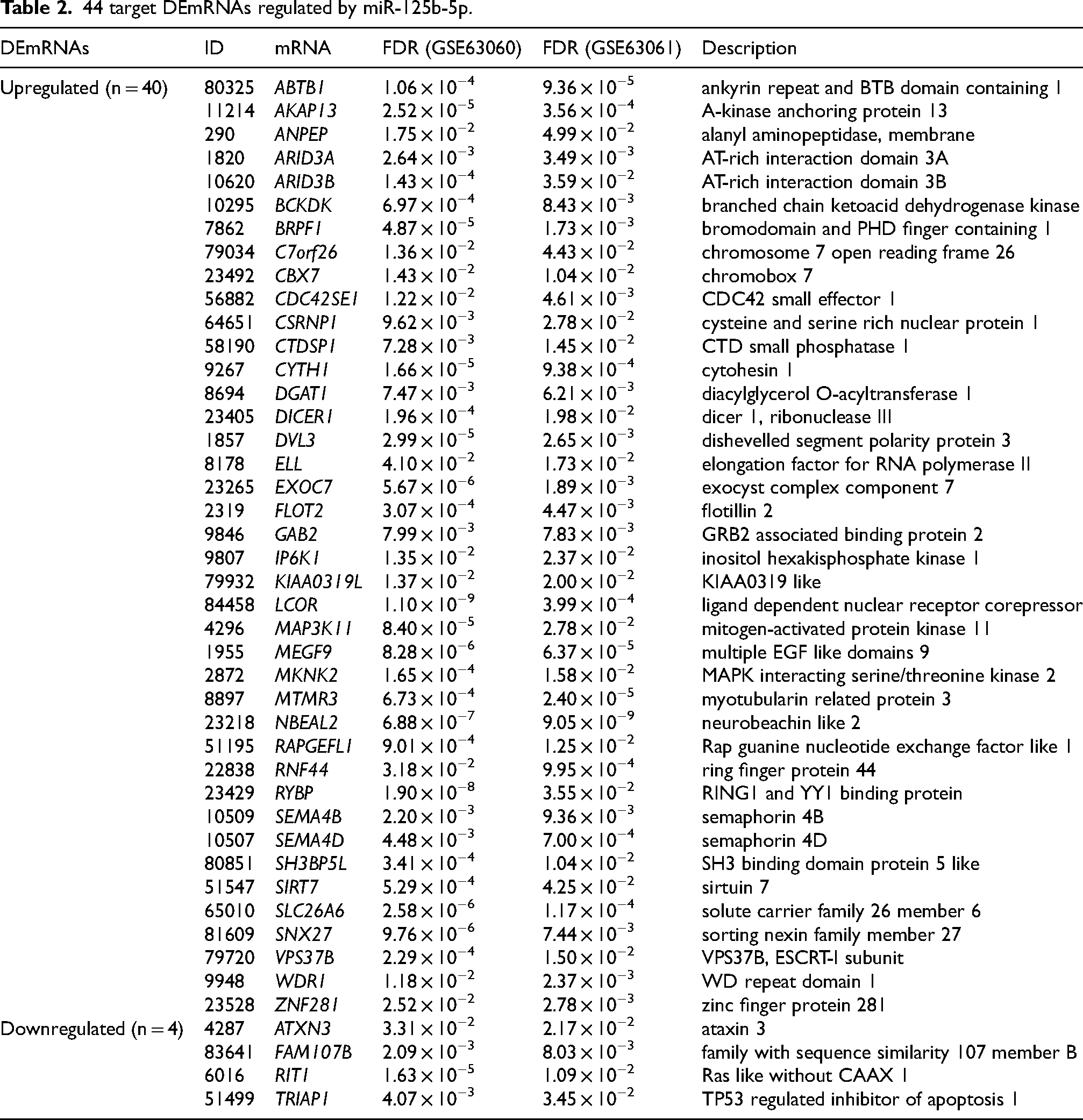
There were 40 upregulated target DEmRNAs (cross of 613 target mRNAs and 674 upregulated DEmRNAs) and 4 downregulated target DEmRNAs (cross of 613 target mRNAs and 714 downregulated DEmRNAs) in the target mRNAs dataset and DEmRNAs dataset (Figure 3(a) and (b)). Figure 3(c) shows the network diagram of miR-125b-5p regulating 44 target DEmRNAs. Orange triangle represents 40 upregulated target DEmRNAs, and green vee represents 4 downregulated target DEmRNAs. These target DEmRNAs are regulated by pro-inflammatory factor miR-125b-5p and may have potential roles in inflammation-related pathways. Table 2 counts the more information of 44 target DEmRNAs.
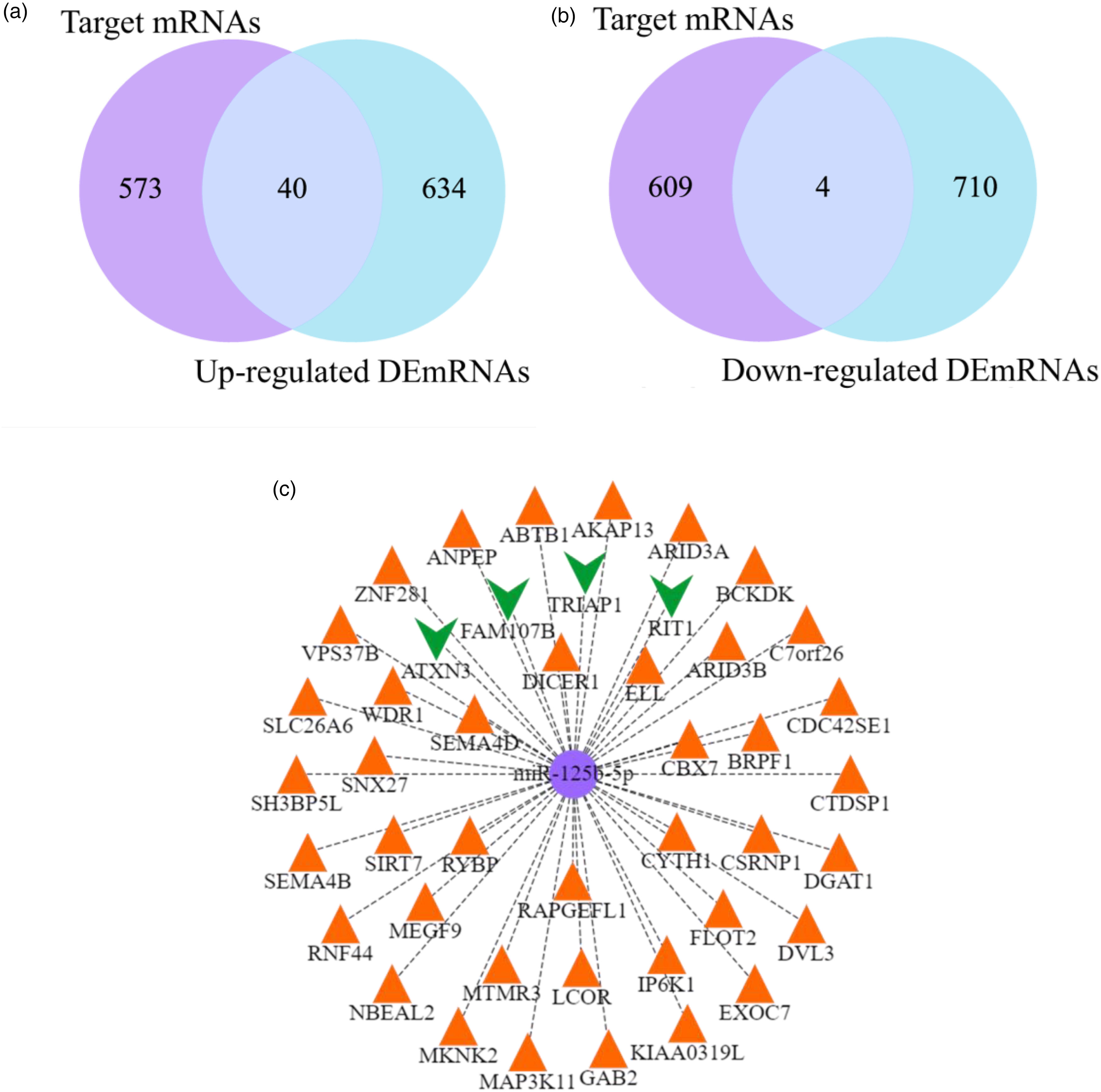
Target DEmRNAs and network diagram. (a) 40 upregulated target DEmRNAs, and (b) 4 downregulated target DEmRNAs. The purple represents the target mRNAs dataset, and blue represents the DEmRNAs dataset. (c) The network diagram of miR-125b-5p regulating 44 target DEmRNAs. Orange triangle represents 40 upregulated target DEmRNAs, and green vee represents 4 downregulated target DEmRNAs.
44 target DEmRNAs regulated by miR-125b-5p.
Functional enrichment analysis of target DEmRNAs
The GO and KEGG enrichment analysis were used to further study the function and enrichment pathway of target DEmRNAs (Supplemental Tables 1 and 2). The GO analysis includes biological process, cellular component, and molecular function, and visualized by Figure 4(a)–(c). Biological process mainly contained the establishment of lymphocyte polarity, histone H3 deacetylation and platelet formation; Cellular component mainly contained the cortical actin cytoskeleton, cortical cytoskeleton and cell cortex; Molecular function mainly contained the Rho GTPase binding, semaphorin receptor binding and guanyl-nucleotide exchange factor activity. KEGG analysis revealed three signaling pathways, namely, insulin signaling pathway (mRNAs: EXOC7, FLOT2, MKNK2), phosphatidylinositol signaling system (mRNAs: IP6K1, MTMR3), and phospholipase D signaling pathway (mRNAs: CYTH1, GAB2), the visualized result is shown in Figure 4(d). These three signaling pathways are all critical in regulating various cellular processes, including inflammation. Given that the above seven mRNAs are involved in these pathways, it suggests that there may be a link between their expression and inflammatory responses, highlighting their relevance in understanding the molecular mechanisms underlying inflammatory diseases.

Go and KEGG enrichment analysis of target DEmRNAs. (a) Biological process; (b) Cellular component; (c) Molecular function; (d)KEGG enrichment pathways.
High correlation of target DEmRNAs in the pathways
From the above analysis results, it can be seen that seven target DEmRNAs (CYTH1, EXOC7, FLOT2, GAB2, IP6K1, MKNK2, MTMR3) are involved in AD-related signaling pathways, so the next step is to focus on analyzing these key genes. Figure 5 shows the relationships among seven target DEmRNAs. CYTH1 had the highest positive correlation with EXOC7 (PCC = 0.93), and also showed a high positive correlation with FLOT2 (PCC = 0.9), GAB2 (PCC = 0.86), IP6K1 (PCC = 0.91), MKNK2 (PCC = 0.88) and MTMR3 (PCC = 0.89). Similarly, EXOC7 was highly positively correlated with FLOT2 (PCC = 0.94), GAB2 (PCC = 0.86), IP6K1 (PCC = 0.95), MKNK2 (PCC = 0.9) and MTMR3 (PCC = 0.92). FLOT2 was highly positively correlated with GAB2 (PCC = 0.94), IP6K1 (PCC = 0.96), MKNK2 (PCC = 0.93) and MTMR3 (PCC = 0.96). GAB2 had the highest positive correlation with MTMR3 (PCC = 0.95), and also showed highly positively correlated with IP6K1 (PCC = 0.92) and MKNK2 (PCC = 0.93). IP6K1 was highly positively correlated with MKNK2 (PCC = 0.95) and MTMR3 (PCC = 0.96). MKNK2 was highly positively correlated with MTMR3 (PCC = 0.94). The correlation analysis reveals the potential high correlation among these seven target DEmRNAs.
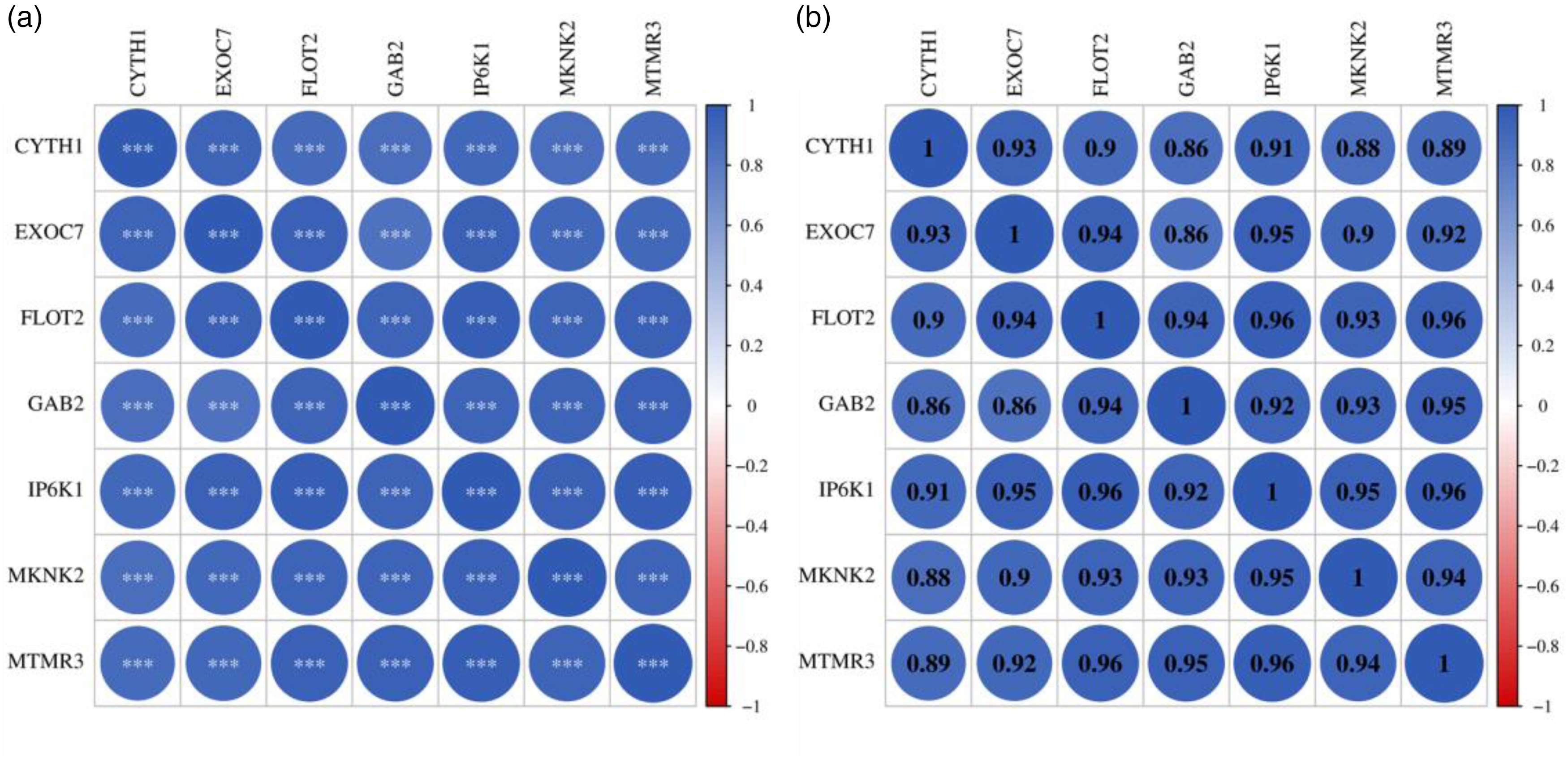
Correlation analysis among seven target DEmRNAs. Each cell contains (a) p-value and (b) PCC. *p < 0.05; **p < 0.01; ***p < 0.001. The coefficient of correlation is presented by a coloring scheme from red (negative correlation) to blue (positive correlation) while white represents an absence of correlation.
Significant expression and reliable predictive ability of target DEmRNAs in the pathways
The boxplot was used to represent the differential expression levels of target DEmRNAs in the three pathways. Figure 6(a)–(g) shows the expression levels of genes CYTH1 (p = 5.1 × 10−5), EXOC7 (p = 6.5 × 10−6), FLOT2 (p = 1.5 × 10−5), GAB2 (p = 4.3 × 10−5), IP6K1 (p = 7.7 × 10−4), MKNK2 (p = 1.5 × 10−5), and MTMR3 (p = 2.3 × 10−5), p < 0.05 indicates that the differences of these genes are significant. In addition, it was observed from the Figure 6 that both the AD group and the control group showed significant two-part data, which may be due to the use of two different microarray datasets, which can introduce variability in gene expression levels. This variability is a result of differences in normalization and scaling between the datasets. Figure 7(a)–(g) shows the AUC result, the AUC values of CYTH1, EXOC7, FLOT2, GAB2, IP6K1, MKNK2, and MTMR3 were 0.6027, 0.6143, 0.6097, 0.6037, 0.5852, 0.6097, and 0.6073, respectively. AUC > 0.5 indicates that the predictive ability of these genes is reliable.
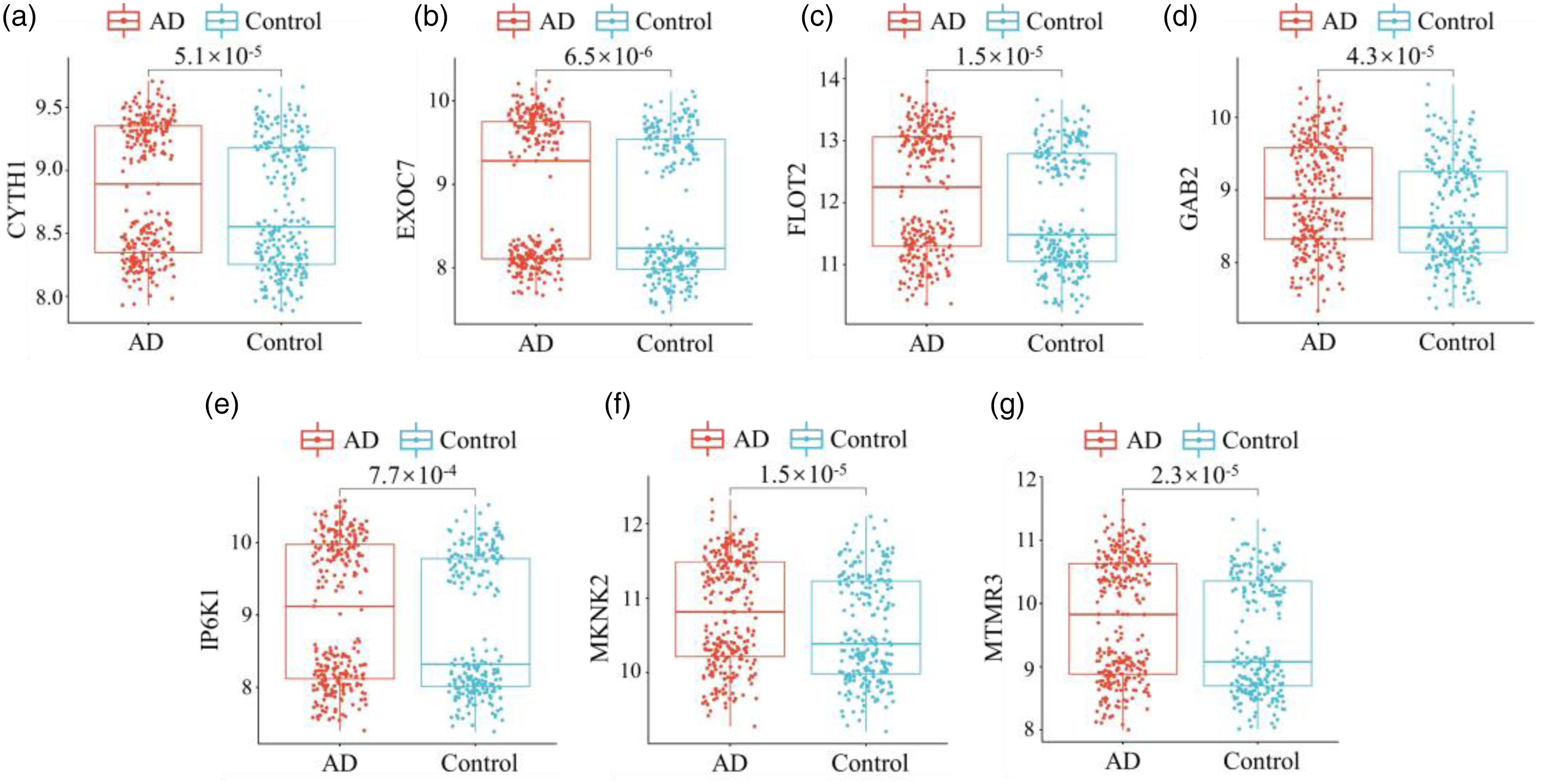
Boxplot of target DEmRNAs expression levels. (a) CYTH1: p = 5.1 × 10−5; (b) EXOC7: p = 6.5 × 10−6; (c) FLOT2: p = 1.5 × 10−5; (d) GAB2: p = 4.3 × 10−5; (e) IP6K1: p = 7.7 × 10−4; (f) MKNK2: p = 1.5 × 10−5; (g) MTMR3: p = 2.3 × 10−5. p < 0.05 indicates the difference is statistically significant.

Predictive ability of target DEmRNAs. (a) CYTH1: AUC = 0.6027; (b) EXOC7: AUC = 0.6143; (c) FLOT2: AUC = 0.6097; (d) GAB2: AUC = 0.6037; (e) IP6K1: AUC = 0.5852; (f) MKNK2: AUC = 0.6097; (g) MTMR3: AUC = 0.6073. AUC > 0.5 indicates that the predictive ability is reliable.
Newly identified mRNAs in AD
Comparative analysis results are shown in Figure 8. Among the 7 core target DEmRNAs found in the key pathway of this study, one gene (GAB2) has been reported and included in the existing database, and four genes (GAB2, FLOT2, MKNK2, MTMR3) have been reported to be associated with AD. Three genes (EXOC7, IP6K1, CYTH1) can be considered as newly discovered AD related genes (Table 3).
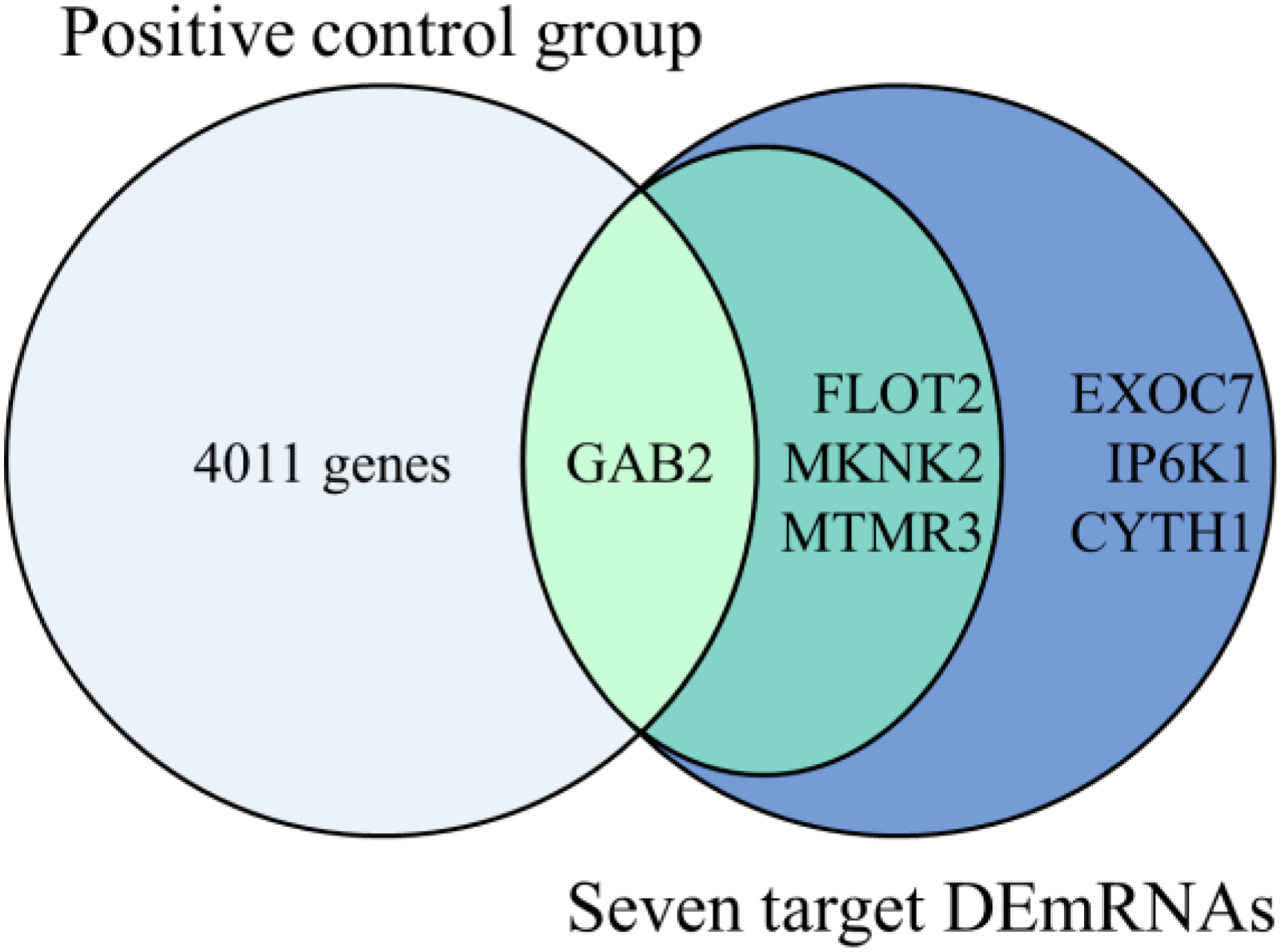
Comparative analysis result. Inclusion in database: GAB2; Published: GAB2, FLOT2, MKNK2, MTMR3; Newly identified: EXOC7, IP6K1, CYTH1.
Newly identified mRNAs in AD.

Prediction of mir-125b-5p binding sites within the target mRNA 3′UTRs
Through bioinformatics analysis, this study identified miR-125b-5p binding sites in the 3′-UTR regions of target mRNAs, including CYTH1, EXOC7, FLOT2, GAB2, IP6K1, MKNK2, and MTMR3 (Figure 9). The results showed that miR-125b-5p binding sites are present in the 3′-UTR of mRNAs, and these target mRNAs may play significant roles in AD-related inflammatory responses and other related biological processes.

miR-125b-5p pairing regions with mRNAs 3′ UTR.
Discussion
Pro-inflammatory miR-125b-5p can induce the expression of multiple target mRNAs and regulate neuroinflammatory signaling pathways. However, its function and specific mechanism in the development of AD is still not clear. Therefore, key mRNAs and important pathways related to AD inflammation regulated by pro-inflammatory miR-125b-5p were identified by integrating target mRNAs and mRNA expression profile datasets in this work.
The study first predicted 613 target mRNAs of miR-125b-5p, then the target mRNAs were correlated with DEmRNAs in the GSE63060 and GSE63061 expression profiles, and 44 target DEmRNAs were obtained. Through the analysis of these target DEmRNAs pathways, three important signaling pathways related to AD pathology were found, namely, insulin signaling pathway) (mRNAs: EXOC7, FLOT2, MKNK2), phosphatidylinositol signaling system (mRNAs: IP6K1, MTMR3) and phospholipase D signaling pathway (mRNAs: CYTH1, GAB2). The correlation analysis reveals the potentially high correlations among 7 mRNAs. Subsequently, it was verified that the differential expression levels of 7 mRNAs in AD patients and control subjects were all significant (p < 0.05), and the AUC curves were all greater than 0.5, indicating that the predictive ability of mRNAs was credible. These seven target mRNAs are regulated by miR-125b-5p and show upregulation, which may be due to several contributing factors. Firstly, feedback mechanisms might play a significant role. The upregulation of target mRNAs could trigger a compensatory increase in miR-125b-5p expression, but miR-125b-5p may not be able to fully suppress the elevated levels of its targets, leading to the co-upregulation observed. Additionally, the regulatory effects of miRNAs are context-dependent, and specific cellular environments in the brains of AD patients, such as inflammatory responses or other stress-related factors, may alter miR-125b-5p-mediated regulation, resulting in the simultaneous upregulation of miR-125b-5p and its targets. Thirdly, while miR-125b-5p is upregulated, it may only partially inhibit its target mRNAs, allowing them to be upregulated as well. This partial inhibition could be due to other competing factors or the relative abundance of the target mRNAs. Finally, the regulation of miR-125b-5p and its targets may differ between the brain and serum. While miR-125b-5p is downregulated in serum, its upregulation in the brain might reflect a tissue-specific response to the pathological environment of AD. Moreover, miR-125b-5p binding sites exist in the 3′-UTR of these 7 mRNAs, and these target mRNAs may play significant roles in AD-related inflammatory responses and other related pathways. Finally, positive control analysis revealed three mRNAs (EXOC7, IP6K1, CYTH1) in this study that could be considered newly identified AD-related mRNAs, which were unpublished and not present in existing AD-related gene databases.
The insulin signaling pathway is the main signal transduction pathway of insulin physiological function, which is related to the regulation of metabolic, neuroprotective and cognitive dysfunction. More and more evidence show that AD patients have symptoms of insulin signaling pathway disorder, suggesting that the insulin signaling pathway may be closely related to the incidence of AD. 51 Insulin can enter the central nervous system cross the BBB, then bind to the insulin receptor (IR) causing cerebrovascular dysfunction and cognitive deficits. 52 This process is involved in glial cell stimulation and inflammation, 53 glial cells and cytokine mediated inflammatory cascades in the brain are the basis of the pathogenesis of AD. Moreover, insulin can regulate the secretion of inflammatory factors through astrocytes and microglia in response to inflammatory factors. 54 Studies have found that insulin administration can reduce chronic neuroinflammation in the brain and improve memory function. 55 In addition, studies have shown that miRNAs promote AD progression by targeting mRNAs through insulin signaling pathways. For example, miR-98 induces AD-like disorders by targeting insulin-like growth factor 1 (IGF1), 56 and miR-206 enhances inflammation and Aβ release by targeting IGF1 in microglia cell. 57
The FLOT2 gene in the insulin signaling pathway belongs to the flotillin family, and flotillin is involved in signal transduction and membrane protein interaction. Relevant study shows that flotillin is associated with the pathogenesis of AD, which may be related to APP endocytosis and Aβ-induced neurotoxicity. 46 In a clinical trial, the results showed that the serum flotillin protein level can be used as one of the blood markers for the detection of brain amyloid deposition and early diagnosis of AD. 47 In addition, the increase of flotillin-2 observed in the brain of older adults in clinical studies may trigger the internalization of amyloid-β protein precursor (AβPP) and affect Aβ metabolism, thus predisposing the older adult to AD. 48 Another mRNA, MKNK2 (MAPK interacting serine/threonine kinase 2), belongs to the protein kinase superfamily, which mediates cell cycle, apoptosis and angiogenesis. An in-vitro experiment showed that MNKN2 regulates autophagy by targeting miR-125b. 49 Autophagy is a non-selective degradation process that can remove abnormally aggregated proteins to maintain cell homeostasis. Autophagy dysfunction is also closely associated with the pathophysiology of AD. Increased autophagy around dystrophic neuritis has been observed in the AD models and postmortem brains of AD patients, suggesting that defects in autophagy can induce the progression of AD. 58
Phosphonoinositol-3 kinase (PI3K) pathway is the main signal transduction pathway of insulin signaling pathway, 59 and PI3K belongs to the phosphatidylinositol signaling system. PI3K is related to inflammation and pathology in AD. Studies have shown that PI3K inhibition reduces activity and memory function in AD mice, while increasing insulin resistance, 59 PI3K pathway may be a new target for regulating AD inflammation. 60 MTMR3 (myotubularin related protein 3) is found in the PI3K pathway, and an in-vitro study has shown its overexpression can also induce autophagy. 50 Another important pathway is the phospholipase D (PLD) signaling pathway. PLD, as a key component of a variety of signaling and metabolic pathways, plays a role in autoimmune central nervous system inflammation. PLD1 inhibitors are feasible therapies for neuroinflammatory diseases. 61 Studies have reported that PLD1 is upregulated in the mitochondrial part of the brain in AD patients, 62 which regulates AβPP transport and γ-secretase complex processing through its interaction with presenilin 1 (PS1).63,64 In vivo experiments, PLD2 has been shown to improve memory deficits and promote synaptic protection in AD mice, 65 and PLD3 has been associated with amyloid plaques and cognitive function in AD. 66 The protein encoded by the gene GAB2 (GRB2 associated binding protein 2) is the primary activator of PI3K in response to receptor activation. GAB2 silencing can inhibit the expression of inflammatory factors in response to various inflammatory stimuli. A clinical genetic association analysis identified GAB2 as an AD susceptibility gene, expressed in pathologically vulnerable areas of the brain. 45
Therefore, it can be found that insulin signaling pathway, phosphatidylinositol signaling system, PLD signaling pathway are intricately related to target mRNAs, which in turn are closely related to AD pathology. miR-125b-5p regulates target mRNAs and participates in important AD signaling pathways, which may induce AD inflammation. Surprisingly, this study found that MKNK2 and MTMR3 may induce autophagy response, and autophagy defects can also promote the progression of AD. MKNK2 and MTMR3 are enriched in the insulin signaling pathway and phosphatidylinositol signaling system, respectively. Studies have shown that miR-125b-5p is related to autophagy. For example, one in vitro study demonstrated that silencing miR-125b-5p can protect human macrophages from bacterial infection by promoting apoptosis and inhibiting inflammation by targeting DRAM2 (DNA damage-regulated autophagy modulator 2). 67 Another clinical study found that downregulation of miR-125b-5p in patients with systemic lupus erythematosus can inhibit autophagy by targeting the UVRAG gene. 68 In addition, bioinformatics studies in this study showed that the AD-related gene MKNK2 can regulate autophagy through targeted regulation of miR-125b. 49
In our following work, we will further use the gene chip data to explore the potential relationship between autophagy related mRNAs and AD pathway, which would be helpful to demonstrate the cascade reaction between autophagy, inflammation and AD, provide new clues for the research on the mechanism of AD, and provide a basis for the induction of autophagy as a new therapeutic strategy. However, there are some limitations to this study. At present, we have screened newly mRNAs through traditional bioinformatics analysis methods and verified the mRNAs through data analysis. Due to the limitations in our laboratory, the validation experiments cannot be conducted at this stage, but is planned in the future work. In addition, this study did not provide disease staging for AD patients, resulting in performance of subgroup analyses based on disease staging not currently applicable.
Conclusions
In this study, we identified key mRNAs and signaling pathways regulated by pro-inflammatory miR-125b-5p by integrating the target mRNAs and mRNA expression profile datasets of miR-125b-5p. We demonstrated that miRNA and mRNAs pathways interact and are closely related to AD inflammation. This study provides new evidence for the role of miR-125b-5p as a pro-inflammatory factor in AD pathology.
Supplemental Material
sj-xlsx-1-alz-10.1177_13872877241297178 - Supplemental material for Exploring the role of miR-125b-5p as a pro-inflammatory factor in Alzheimer's disease pathology
Supplemental material, sj-xlsx-1-alz-10.1177_13872877241297178 for Exploring the role of miR-125b-5p as a pro-inflammatory factor in Alzheimer's disease pathology by Wenjia Liu, Sophia Chen, Xin Rao, Xiaodong Chen, Liyang Yu, Jiangtao Zhang, Jiong Chen and Bohan Cui in Journal of Alzheimer's Disease
Footnotes
Acknowledgments
The authors thank the supervision and control of this research program from Dr Jiangtao Zhang and Dr Jiong Chen (Department of Geriatrics, Tongde Hospital of Zhejiang Province).
Author contributions
Wenjia Liu (Conceptualization; Formal analysis; Methodology; Writing – original draft); Sophia Chen (Data curation; Visualization; Writing – original draft); Xin Rao (Conceptualization; Methodology; Project administration); Xiaodong Chen (Methodology; Project administration; Supervision); Liyang Yu (Investigation; Supervision); Jiangtao Zhang (Investigation; Supervision); Jiong Chen (Supervision); Bohan Cui (Methodology).
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the National Natural Science Foundation of China (Grant No. 62101171), Natural Science Foundation of Zhejiang Province (Grant No. LQ22F010015), and National Key R&D Program of China (Grant No. 2023YFC2412001).
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data availability
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.