
Abstract
Background
Alzheimer’s disease (AD) is a progressive neurodegenerative disorder that predominantly affects elderly individuals across the globe. While genetic, environmental, and lifestyle factors are known to influence the onset of AD, the underlying mechanisms remain unclear.
Objective
To elucidate the intricate interplay between metabolites and immune cell activation in the ethology of AD, and to determine their collective impact on AD risk.
Methods
We conducted a comprehensive analysis of genome-wide association studies data to examine the relationships between metabolites, immune cell phenotypes, and the risk of AD. Our study encompassed a comprehensive examination involving 731 distinct immune cell types, 1400 metabolites, and a large cohort comprising10,520 AD cases with 401,661 controls. We employed univariate Mendelian randomization to assess bidirectional relationships between metabolites and AD, metabolites and immune cells, as well as immune cells and AD. Subsequently, multivariate Mendelian randomization was then applied to evaluate the potential mediating role of immune cells on the relationship between metabolites and AD.
Results
Specific metabolites, the histidine/pyruvate ratio and homoarginine, were positively associated with the risk of AD, mediated by immune cells. Conversely, 4-hydroxycoumarin and glycolithocholate sulfate showed protective associations against AD. Immune cell markers, CD64 on monocytes and HLA DR on CD14+ CD16− monocytes were linked to higher AD risk, while CD33dim HLA DR+ CD11b− myeloid cells and HLA DR on CD8+ T cells were protective.
Conclusions
This study highlights the critical role of immune cells in the pathogenesis of AD, demonstrating how their interaction with specific metabolites influences disease risk.
Introduction
Alzheimer’s disease (AD) is a progressive neurodegenerative illness characterized by memory loss, cognitive impairment, and difficulty with daily living activities. The prevalence of AD is increasing as the global population ages, posing a significant threat to the elderly. 1 It currently affects more than 50 million individuals, a figure expected to soar to 150 million by 2050. 2 This disease not only exacts a profound emotional toll on patients and caregivers but also exerts a substantial burden on healthcare systems and economies. The pathological hallmarks of AD are the aggregation of amyloid-β (Aβ) plaques and the formation of tau protein aggregates. 3 Normally Aβ peptides are typically cleared from the brain, in AD they accumulate, leading to amyloid plaques that are diagnostic of the disease.3–5 These plaques disrupt neuronal function, potentially triggering neuroinflammation and oxidative stress, which exacerbate neuronal damage.6,7 The tau protein, crucial for microtubule integrity within neurons, becomes hyperphosphorylated in AD, detaching from microtubules and forming neurofibrillary tangles—a pathological hallmark that compromises neuronal integrity and impairs neural signaling. 8 Beyond the dysregulation of Aβ and tau, AD brains also exhibit additional pathologies such as neuronal loss, synaptic dysfunction, and cerebrovascular issues, which collectively contribute to the cognitive and behavioral decline.9–11 Current treatments include pharmacological interventions, lifestyle changes, and supportive care, yet they are limited in efficacy and do not halt disease progression.12–14 Medications aimed at symptom relief have diminishing benefits and can cause significant side effects. 14 Physical activity may slow cognitive decline by promoting neurogenesis and vascular health, but its impact varies due to individual genetic and environmental factors.15,16 The search for effective therapies is a critical challenge for modern society, with the efficacy of current interventions still a matter of debate due to the absence of robust clinical trial data. 17 The development of innovative treatments is imperative to address the escalating burden of AD.
In the pathophysiology of AD, metabolic pathway disorders are indeed a key factor, involving multiple biological pathways and molecular mechanisms. 18 These include amino acid metabolism, the glutamate-glutamine cycle, lipid metabolism, and energy metabolism.19,20 For example, glutamate, as the main excitatory neurotransmitter in the central nervous system, is closely related to neurodegenerative processes when its metabolism is disrupted. 21 In AD, glutamate accumulation can trigger excitotoxicity, damaging neurons and promoting the development of neurodegeneration.22,23 Changes in lipid metabolism, particularly the imbalance of cholesterol and phospholipid homeostasis, have been found to be associated with the accumulation of Aβ in the brain.24,25 Cholesterol plays a crucial role in the composition of cell membranes and is involved in the production and clearance of Aβ. 26 In AD patients, abnormal cholesterol metabolism may lead to excessive Aβ production or impaired clearance, contributing to the formation of Aβ plaques. 27 Energy metabolism disorders also play an important role in AD pathogenesis. 28 The brain, being an organ with high energy demands, primarily relies on glucose oxidation for energy production. 29 In AD patients, abnormal glucose metabolism leads to reduced energy production, affecting the normal function of neurons. Additionally, mitochondrial dysfunction is closely related to AD pathogenesis. 30 As the cell’s energy factory, mitochondrial dysfunction not only impacts energy metabolism but may also lead to oxidative stress and cell apoptosis, exacerbating neurodegenerative changes. 30 Beyond metabolic disorders, immune cells also play roles in the pathological process of AD. Monocytes, T cells, and B cells can cross the blood-brain barrier, aggravating neuroinflammation. 31 The breakdown of the blood-brain barrier makes it easier for these cells to enter the brain, where monocytes have the capacity to differentiate into microglia.32–34 In neurodegenerative diseases like AD, microglia may become hyperactive, exacerbating neural damage. 35 The interaction between microglia and T cells promotes the formation of an inflammatory environment, while T cells have a dual role in clearing pathogens and exacerbating neurodegenerative changes. 36 Understanding how these immune cells influence the immune response in AD is critical for developing new therapies.
Expanding upon the existing framework of knowledge, delving into the nuanced interplay and potential relationships between metabolites and immune cells in the pathogenesis of AD is of particular importance. While individual studies have provided in-depth analyses of specific metabolites and immune cells in isolation, the intricate interrelationships and synergistic effects within the context of AD’s progression remain a relatively unexplored domain. This study leverages genome-wide association studies (GWAS) data, utilizing Mendelian randomization (MR) analytical strategies, to scrutinize the potential causal associations between metabolites and immune cells in etiological landscape of AD. 37 Our central hypothesis is that select metabolites, through their modulatory effects on immune cell functions, may exert a significant influence on pathogenesis of AD. To empirically test this hypothesis, we will employ single nucleotide polymorphisms (SNPs) as instrumental variables, exploring the latent causal nexus between metabolites and immune cells in pathogenesis of AD via MR analysis. This study is designed to bridge the existing gap in our understanding of the pivotal role that specific metabolite, by impacting immune cell functionality, may play in the evolution of AD. The findings shed new light on the intricate interactions between metabolites, immune cells, and AD. This foundation may inform the development of strategies aimed at attenuating or potentially reversing the trajectory of AD, enhancing patient quality of life, and mitigating the socio-economic ramifications of the disease. Furthermore, our research endeavors to contribute salient scientific evidence that could underpin the innovation of therapeutic strategies for AD.
Methods
Study design
This investigation harnesses GWAS data from publicly accessible databases to meticulously dissect the plausible causal linkages among metabolites, immune cells, and AD, as illustrated in Figure 1. We have upheld stringent ethical standards throughout the research, with the dataset in question having received prior sanction from the Ethics Review Committee. The open-access attribute of the data also precluded the necessity for supplementary ethical clearance. The granular findings of our research are exhaustively documented within the manuscript and appended in the annexes for comprehensive reference.
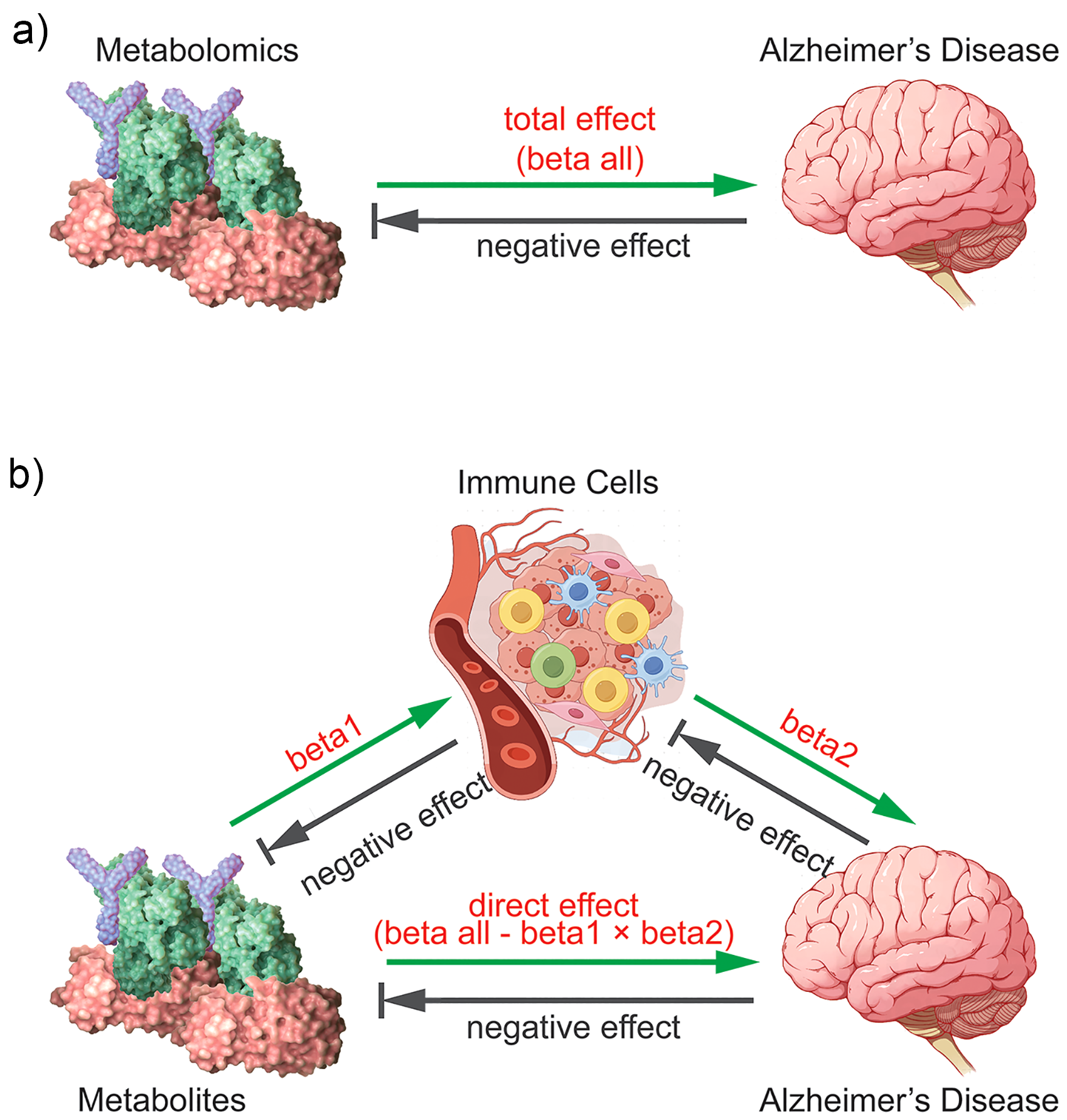
Schematic diagrams of the associations examined in this study. (a) The total effect between metabolites and AD. (b) The total effect is decomposed into: (i) beta1, the total effect of metabolites on immune cells, (ii) beta2, the total effect of immune cells on AD, and (iii) the direct effect is beta all – (beta1 × beta2). The proportions mediated are the ratios of beta1/beta all and beta2/beta all. The grey arrows indicate that reverse causality is not considered in this study, that is, the reverse impact from immune cells to metabolites or from AD to metabolites. These arrows are used to emphasize the unidirectional pathways in our analysis, namely the impact of metabolites on immune cells (beta1) and the impact of immune cells on AD (beta2).
In this investigation, we have utilized a bidirectional MR analytical framework, incorporating two independent and diversely representative cohorts to deeply excavate the latent causal connections between metabolites and immune cells within the etiological mechanisms of AD. The strategic utilization of SNPs as instrumental variables (IVs) marks a significant methodological innovation, augmenting our inferential rigor for uncovering causal pathways. 38 SNPs, serving as the quintessential indicators of genetic variance, are endowed with a stochastic genomic distribution that confers a pronounced methodological advantage. This feature is instrumental in neutralizing the influence of confounding factors, thereby ensuring the integrity and authenticity of our research outcomes.
Data sources
The cornerstone of our study’s dataset is a wide-ranging and demographically representative sample of the European population, intricately linked to the study of AD. Our data is derived from the most comprehensive GWAS conducted to date, with a special acknowledgment to the Finngen project (https://www.finngen.fi/en) for their contribution of G6_Alzheimer SNPs data. 39 The G6_Alzheimer dataset is meticulously constructed based on the International Classification of Diseases (ICD-10) system, and it takes into account age, gender, and other potential confounding factors to provide us with a comprehensive perspective on the genetic factors of a range of diseases, including early-onset AD and late-onset AD. 39 This dataset comprises 10,520 individuals with clinically validated AD diagnoses and 401,661 controls, reinforcing the study’s methodological stringency and the veracity of its findings. Adhering to the highest international clinical diagnostic standards, the collection of AD patient samples has yielded a dataset of exceptional quality and scope. This dataset is instrumental in conducting an in-depth analysis of the genetic risk factors associated with AD, as well as their interaction with the disease’s developmental trajectory.
For our investigation, metabolite data were curated from the GWAS Catalog (https://www.ebi.ac.uk/gwas/), a preeminent database celebrated for its rich depository of associations between genetic variations and phenotypic manifestations. 40 The dataset we accessed, spanning accession numbers GCST90199621 to GCST90201020, offers a thorough resource that meticulously outlines an array of metabolite categories alongside the kinetics of their metabolic interconversions. 41 This dataset is inclusive of a multitude of metabolite classes, such as amino acids, carbohydrates, cofactors and vitamins, entities pertinent to energy metabolism, lipids, nucleotides, partially characterized molecules, peptides, and xenobiotics. It encapsulates the diversity of individual metabolite species in addition to the sophisticated interplay of their metabolic conversions, including the intricate relationships and transformations among amino acids, carbohydrates, cofactors and vitamins, energy, lipids, nucleotides, and xenobiotics.
The comprehensive dataset of immune cell statistics was extracted from the GWAS Catalog, spanning accession numbers GCST90001391 to GCST90002121, and is centered on a sample cohort of 3757 individuals hailing from European populations, ensuring ample breadth and representation within the study’s demographic scope. 42 Encompassing 731 distinct immune cell types and an approximate count of 22 million SNPs, this dataset has been meticulously adjusted for key covariates including age, age squared, and gender to bolster the precision and reliability of the study’s results. The assessment of immune cell profiles was conducted using diverse methodologies, including median fluorescence intensity values based on surface antigen expression (accession number: GCST90001701 to GCSG90001962, GCST90001979 to GCSG90002070, and GCST90002087 to GCST90002121), morphological parameters (accession numbers: GCST90001963 to GCST90001978, GCST90002071 to GCST90002086), and relative cell counts (accession numbers: GCST90001391 to GCST90001700).
Instrumental variable selection
Our analytical approach in this study is underpinned by a battery of rigorously crafted methodologies designed to elucidate the potential causal interconnections between AD, metabolites, immune cells, and specific health outcomes. We initiated our investigation by identifying SNPs with substantial associations to the exposure variables, applying a highly conservative significance threshold of p < 1 × 10−5 to guarantee a formidable relationship between the instrumental variables and the exposures in question. 43 To uphold the integrity of the instrumental variable set and to preclude confounding arising from linkage disequilibrium (LD), we adhered to a strict selection regimen facilitated by PLINK software. 43 The LD criterion for the analysis pertaining to AD and metabolites was established at r2 < 0.01, with a physical distance criterion set at 10,000 kilobase pairs. When immune cells were under investigation as exposure variables, we calibrated the LD criterion to r2 < 0.1, with a reduced physical distance criterion of 500 kilobase pairs to better reflect genetic patterns within distinct biological contexts. 44 The assessment of the instrumental variables’ efficacy was anchored in the LD reference panel data from the 1000 Genomes Project, meticulously designed to represent the genetic tapestry of a European super-population, thereby providing a solid genetic substratum for our analysis. 45 These methodological choices have been instrumental in assuring the genetic fidelity and reliability of our selected instrumental variables. Moreover, we undertook F-statistic computations for each instrumental variable to gauge its efficacy. Throughout this evaluation, we maintained a keen vigilance for SNPs with F-statistics below the 10 threshold, identifying them as weak instrumental variables that could skew the analytical outcomes. 46 As such, we were poised to exclude these SNPs to reinforce the statistical significance, robustness, and credibility of our causal inferences.
Sensitivity analysis
In our MR analysis, the inverse variance weighted method was instrumental in identifying significant causal effects. To fortify the robustness of our findings against conceivable breaches of MR assumptions, we meticulously applied a quartet of distinct evaluative methods. These include the MR Egger method for probing potential pleiotropy 13 ; the Weighted median approach to attenuate the effects of outliers 47 ; the Simple mode, aligned with the most frequently occurring effect size estimate 13 ; and the Weighted mode, which amalgamates effect magnitude with statistical significance 48 ; This multifaceted appraisal strategy is employed to substantiate the fidelity and steadfastness of our analytical conclusions. Furthermore, we incorporated the MR-PRESSO method, detected via the heterogeneity examination, to reinforce the analytical rigor. Additionally, we employed Cochran’s Q statistic alongside funnel plots to meticulously appraise the heterogeneity among SNPs effect estimates, ensuring the integrity of our genetic inferences.45,49
Results
Association of genetically predicted metabolites with AD risk
In this study, the application of inverse variance weighted analysis has brought to light significant correlations between specific metabolites and AD. Our data indicate a pronounced positive association between homoarginine and the metabolite ratio of histidine/pyruvate with AD, which may underpin their potential to exacerbate the disease’s onset and progression (Figure 2(a)). In contrast, the negative correlations observed between glycolithocholate sulfate and 4-hydroxycoumarin with AD could signify their potential protective influence or a retarding effect on disease advancement (Figure 2(a)). To reinforce the veracity of these preliminary observations, we executed a comprehensive assessment through four diverse MR methods, including inverse variance weighted, MR-Egger, weighted median regression, and MR-PRESSO, each corroborating the metabolite-AD nexus from unique analytical angles. Supplemental Tables 1 and 2 delineate the statistical data and interpretive insights of these analyses, furnishing a solid evidentiary foundation for our research. Importantly, our MR analysis has successfully excluded the possibility of reverse causality between the genetically predicted values of these metabolites or ratios and AD, thereby precluding the confounding impact of extraneous factors on our results (Figure 2(b) and Supplemental Table 2). The symmetrical distribution within funnel plots (Supplemental Figures 1 and 5) and the non-significant intercepts from the MR-Egger tests (Supplemental Figures 2 and 6) provide no evidence of publication bias or directional pleiotropy, thus fortifying our confidence in the study’s outcomes. The execution of single SNP analyses (Supplemental Figures 3 and 7) and leave-one-out sensitivity analyses (Supplemental Figures 4 and 8) has further mitigated the impact of any particular SNP on the collective estimates, substantiating the robustness and integrity of our findings.
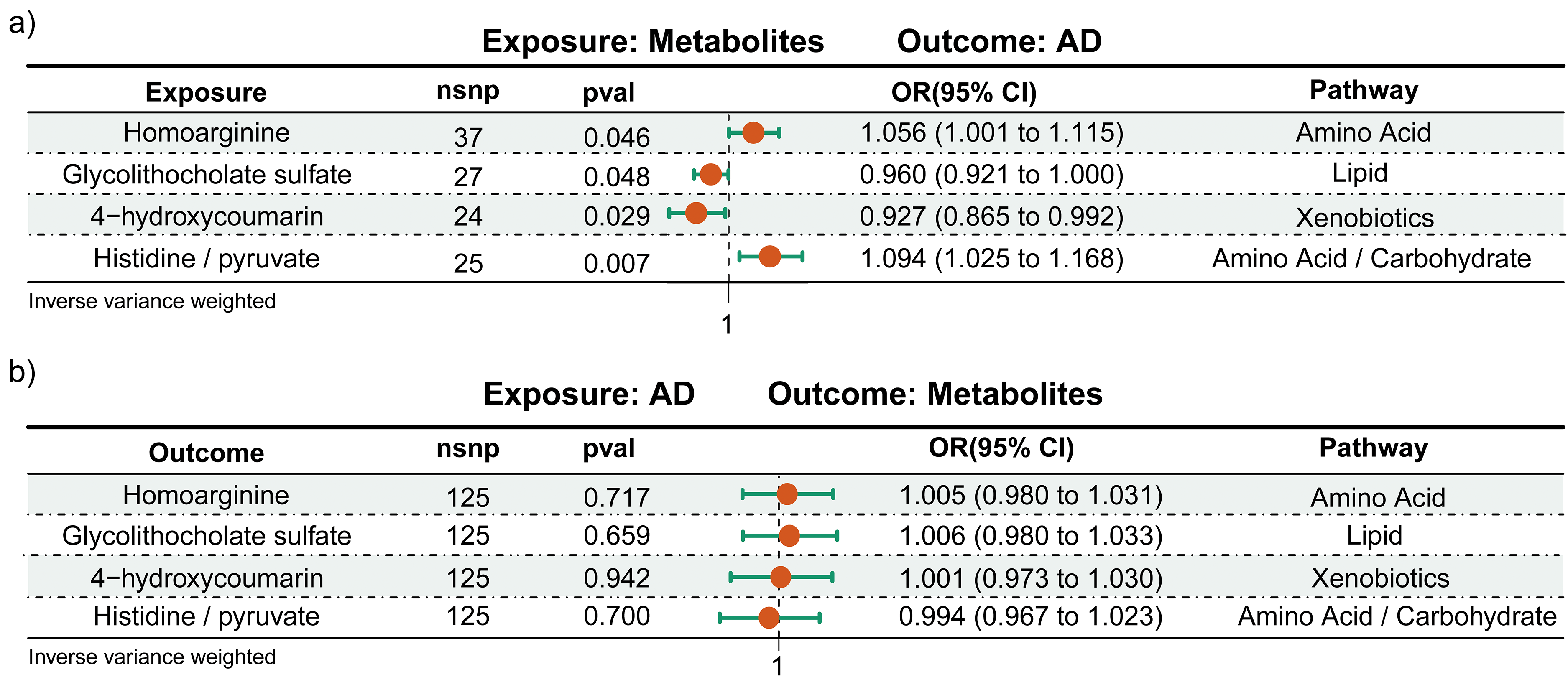
The bidirectional MR estimates exploring the relationship between metabolites and AD. (a) Illustrates the analysis where metabolites are regarded as the exposure, with AD as the outcome. (b) Depicts the analysis with AD as the exposure and metabolites as the outcome. nsnp: number of single-nucleotide polymorphism; pval: p value; OR: odd ratio; CI: confidence interval.
Association of genetically predicted metabolites with immune cells
Post the application of multiple testing corrections, genetic instrumentality linked four specific metabolites to associations with four categories of immune cells (Figure 3 and Supplemental Table 3). Within the framework of inverse variance weighted analysis, the genetically forecasted histidine/pyruvate ratio manifested significant positive correlations with the expression of CD64 on monocyte, and homoarginine was correlated with significant positive associations with HLA DR on CD14+ CD16− monocyte. On the contrary, 4-hydroxycoumarin was found to have significant inverse correlations with the proportion of CD33dim HLA DR+ CD11b− %CD33dim HLA DR+, and glycolithocholate sulfate was correlated with significant inverse relationships with HLA DR on HLA DR+ CD8+ T cells (Figure 3(a)). Of particular importance, the MR analysis did not uncover any evidence of reverse causality between the genetically predicted values of these metabolites or their ratios and the immune cells in question, effectively ruling out the potential influence of confounding factors on our results (Figure 3(b) and Supplemental Table 3).
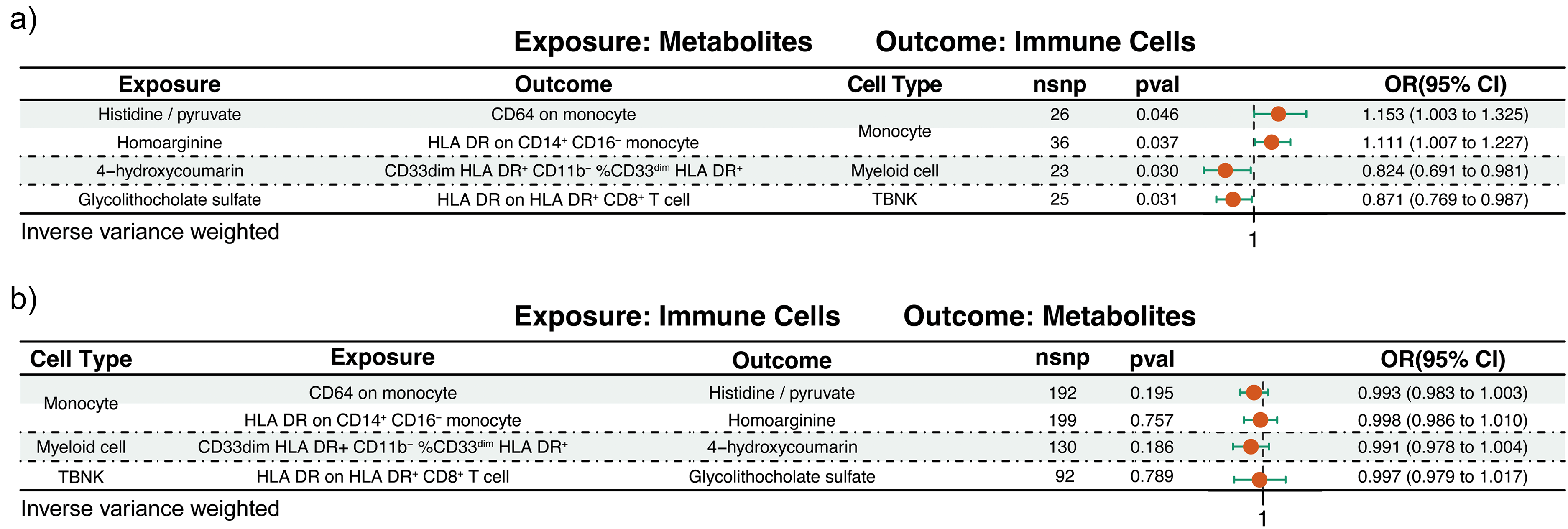
The bidirectional MR estimates of the association between metabolites and immune cells. (a) Metabolites serve as the exposure with immune cells as the outcome. (b) Represents the immune cells serve as the exposure and metabolites are the outcome. nsnp: number of single-nucleotide polymorphism; pval: p value; OR: odd ratio; CI: confidence interval.
To reinforce the credibility of our analytical findings, we implemented a series of bias detection protocols. The funnel plots’ symmetry (Supplemental Figures 9 and 13) is indicative of an absence of significant publication bias. Additionally, the MR-Egger intercept tests (Supplemental Figures 10 and 14) did not uncover any indications of directional pleiotropy, further bolstering our conviction in the results. In our quest to appraise the robustness of the outcomes, we executed single SNP analyses (Supplemental Figures 11 and 15) and conducted leave-one-out sensitivity analyses (Supplemental Figures 12 and 16) to preclude any excessive impact that particular SNPs might have on the collective estimates. The consistency of the results from these sensitivity analyses, devoid of any notable anomalies, substantiates the integrity of our findings.
Association of genetically predicted immune cells with AD
In our rigorous application of the primary inverse variance weighted analytical method, we identified significant correlations linking genetically predicted immune cell phenotypes to the incidence and progression of AD. Specifically, the expression of HLA DR on CD14+ CD16− monocyte (OR = 1.023, 95% CI = 1.009–1.037) and CD64 on monocyte (OR = 1.009, 95% CI = 1.001–1.018) were both positively correlated with AD. Moreover, the presence of CD33dim HLA DR+ CD11b− %CD33dim HLA DR+ (OR = 0.988, 95% CI = 0.977–0.999) and HLA DR on HLA DR+ CD8+ T cell (OR = 0.982, 95% CI = 0.965–1.000) emerged as protective factors against the development of AD (Figure 4(a)). To reinforce the credibility of these preliminary observations, we executed an integrated analysis utilizing four diverse MR methodologies, each validating the immune cell-AD associations from unique perspectives and providing a solid empirical foundation for our study (Supplemental Table 5). Importantly, our MR analysis did not detect any reverse causality between the genetically predicted values of these immune cells and AD, effectively precluding the potential impact of confounding factors on our findings (Figure 4(b) and Supplemental Table 6).
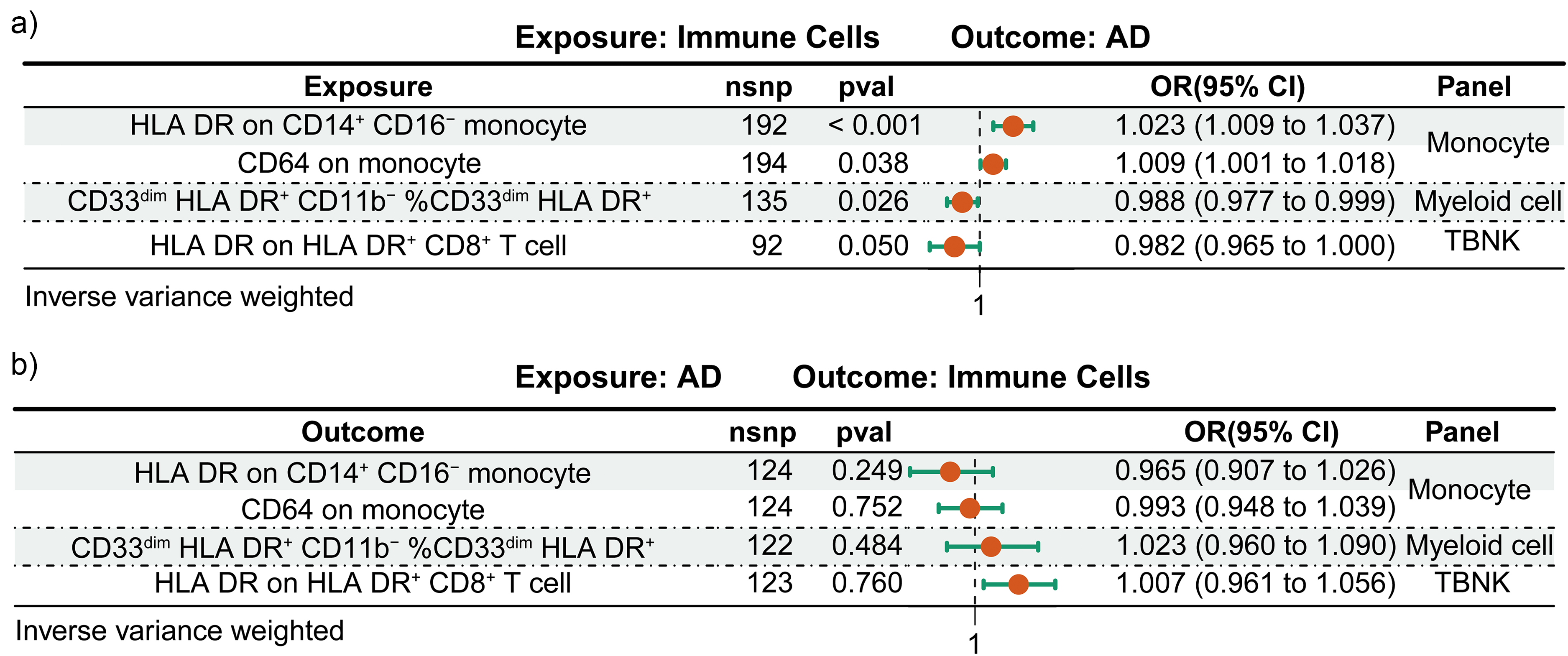
The bidirectional MR estimates of the link between immune cells and AD. (a) Illustrates the analysis where immune cells are the exposure, with AD as the outcome. (b) Reverses the relationship, showing AD as the exposure and immune cells as the outcome in the MR analysis. nsnp: number of single-nucleotide polymorphism; pval: p value; OR: odd ratio; CI: confidence interval.
To affirm the veracity of our analytical results, we executed an exhaustive array of bias detection strategies. Employing funnel plots (Supplemental Figures 17 and 21), we assessed for publication bias, with the plots’ symmetry indicating an absence of notable bias in published research. Subsequently, the MR-Egger intercept test (Supplemental Figures 18 and 22) was utilized to probe for potential pleiotropy, with the findings not exhibiting any overt evidence of pleiotropy. Moreover, to ensure that our conclusions are not disproportionately affected by any SNP, we conducted rigorous SNP-level analyses (Supplemental Figures 19 and 23) and applied a leave-one-out sensitivity analysis (Supplemental Figures 20 and 24). The consistency of these analyses, which revealed no SNP to induce a substantial bias in the overall estimates, further fortifies the robustness and validity of our research outcomes.
Immune cells as mediators of the association between metabolites and AD
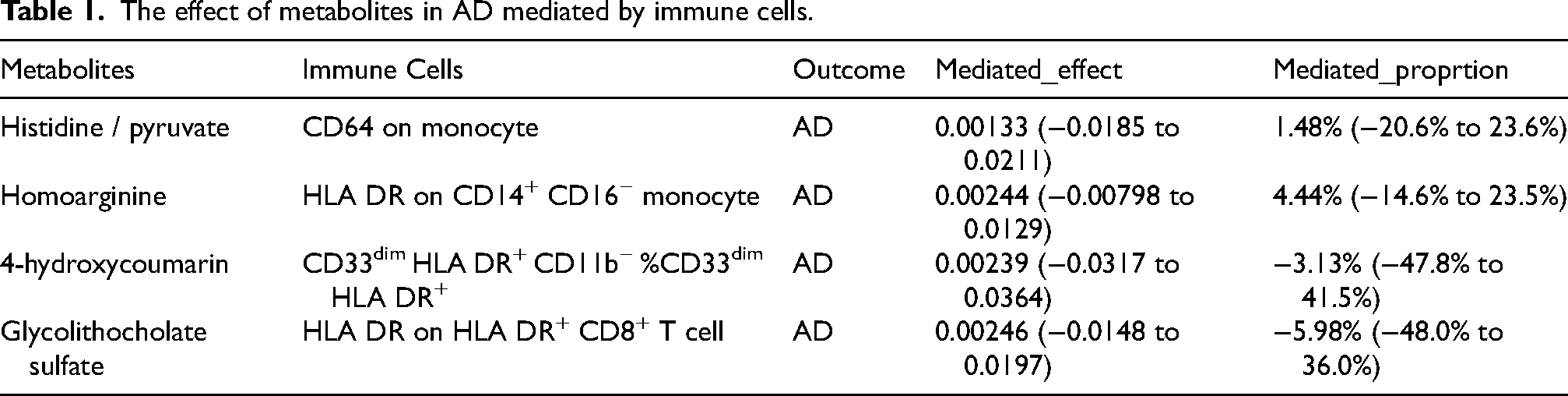
In our examination of immune cells as mediators in the metabolic pathway to AD, we identified significant associations. Specifically, an increase in the histidine/pyruvate ratio was found to be correlated with CD64 on monocyte, a factor that is associated with an elevated risk of AD (mediation effect ratio: 51.48%, 95% CI = −20.6%–23.6%). Furthermore, higher levels of homoarginine were associated with HLA DR on CD14+ CD16− monocyte, which is linked to an increased AD risk (mediation effect ratio: 4.44%, 95% CI = −14.6%−23.5%). Conversely, an increase in 4-hydroxycoumarin was found to be associated with CD33dim HLA DR+ CD11b− %CD33dim HLA DR+, a factor that seems to confer protection against AD (mediation effect ratio: −3.13%, 95% CI = −47.8%–41.5%). Similarly, an increase in glycolithocholate sulfate was linked to HLA DR expression on HLA DR+ CD8+ T cell, which is suggested to have a protective effect in the context of AD (mediation effect ratio: −5.98%, 95% CI = −48.0%–36.0%) (Table 1, Supplemental Table 7).
The effect of metabolites in AD mediated by immune cells.
Discussion
Within the pathophysiological framework of AD, alterations in metabolite levels are of paramount importance, ranging from amino acids and fatty acids to glucose, ketone bodies, the byproducts of neurotransmitter metabolism, and an array of signaling molecules.23,50–53 These entities are integral to the maintenance of cerebral energy homeostasis and the stability of the intracellular environment, and they exert influence over immune cell behavior and function in both direct and indirect manners. Despite the depth of existing research on individual metabolites and immune cells, the nuanced interactions and potential linkages between them in the context of AD’s pathogenesis represent a domain that has yet to be thoroughly investigated. Through the application of MR analysis, we explored the correlation between metabolites and AD, with a focus on establishing whether the causal relationships are mediated by immune cells. Our study’s outcomes underscore a significant correlation between the genetically predicted levels of five metabolites and an increased risk of AD, with this association being realized through the intermediary of immune cells. Furthermore, the genetically predicted levels of two metabolites are found to confer a protective effect against AD, an effect that is intricately connected to the regulatory influence of immune cells.
The histidine/pyruvate ratio is a significant biomarker reflecting the balance of histidine and pyruvate in the blood, increasingly valued in biomedical research for its insights into metabolic and health statuses. As an essential amino acid, histidine underpins diverse biological processes, including cell proliferation, DNA repair, and serves as a precursor for numerous enzymes and hormones. 54 The decarboxylation of histidine to histamine, facilitated by histidine decarboxylase, is central to immune modulation, gastric enzyme secretion, and neural signal transmission.55,56 Thus, histidine and its metabolites are pivotal in physiology and may offer biomarkers for diagnostics or targets for therapeutic intervention. Pyruvate, emerging as a keystone of glycolytic metabolism, is both a gateway to the tricarboxylic acid (TCA) cycle and, anaerobically, a converter to lactate for cellular energy, although histidine’s metabolism does not yield pyruvate directly, its derivative, acetyl-CoA, is a participant in the TCA cycle, impacting energy metabolism and overall metabolic balance through complex metabolic pathways and enzymatic synergies.4,57 In the realm of AD research, the role of metabolic anomalies as pivotal contributors to disease development is well-established.58,59 The metabolic signatures of AD-afflicted individuals markedly contrast with those of healthy counterparts, possibly reflecting shifts in the levels of amino acids and metabolites. 59 Despite a dearth of direct associations between the histidine/pyruvate ratio and AD in the current literature, incipient studies hint at a link between this ratio and the propensity for AD.60–63 Our research contributes to this discourse by demonstrating that the histidine/pyruvate ratio may serve as a risk factor for AD, particularly in relation to CD64 on monocytes. These immune cells potentially alter the metabolic pathways of histidine and the synthesis of pyruvate through specific signal transduction mechanisms, implicating themselves in the pathogenesis of AD. This revelation not only deepens our grasp of metabolic perturbations in AD but also paves the way for the exploration of innovative diagnostic markers and therapeutic strategies.
Homoarginine, bearing structural resemblance to arginine, operates within biological systems with a sophistication that extends beyond initial insights. 64 It evinces robust antibacterial capabilities against a spectrum of microorganisms, including Staphylococcus aureus, Escherichia coli, and Candida albicans, and its roles in human physiology are both broad and significant.64,65 As a direct substrate for nitric oxide (NO) synthase, homoarginine is integral to the synthesis of NO, a signaling molecule of pivotal importance in vasodilation, neurotransmission, and immune response.64,66 The regulated generation of NO is essential for homeostatic balance, and its mis-regulation, through overproduction or imbalance, can lead to neuroinflammatory and oxidative stress pathways that are particularly critical in neurodegenerative diseases such as AD.67,68 Our research posits homoarginine as a notable risk factor in the pathogenesis of AD. Within the pathological paradigm of AD, homoarginine’s involvement as a risk factor is particularly associated with immune responses on HLA DR on CD14+ CD16− monocytes. These monocytes, pivotal in neuroinflammation, exhibit the propensity to migrate to inflammatory sites and differentiate into macrophages or dendritic cells, further engaging in immune responses. The activation of these immune responses is intricately linked to the underlying pathogenic processes of the disease.69–71
In addition, the presence of HLA DR on monocytes equips them with the capacity to present antigens to T cells, thereby triggering the adaptive immune response. 72 This indicates that homoarginine participates in innate immunity and potentially modulates adaptive immunity by influencing monocyte antigen presentation. The pleiotropic functions of homoarginine highlight its integral role in immune modulation, neuroinflammation, and oxidative stress. Advancing our comprehension of the mechanisms by which homoarginine operates in physiological and pathological states could lead to the discovery of new therapeutic strategies for neurodegenerative conditions such as AD. Targeting homoarginine levels or its metabolic pathways could offer a means to regulate inflammation associated with AD. Nonetheless, the precise mechanisms of homoarginine action warrant further investigation. It is essential to explore how homoarginine influences NO synthesis under varying physiological and pathological conditions and to assess its effects on monocyte antigen-presenting functions. Furthermore, the interplay between homoarginine and other immune cells and signaling molecules, along with its potential contributions to AD, requires in-depth study. To encapsulate, status of homoarginine as a versatile non-protein amino acid has unveiled novel research perspectives and pioneering therapeutic avenues in antimicrobial defense, immune modulation, neuroinflammation, and oxidative stress. A profound understanding of its mechanisms could not only foster the development of innovative treatments for neurodegenerative diseases like AD but may also inform the creation of new antimicrobial pharmaceuticals.
Our study preliminarily discloses that 4-hydroxycoumarin, a ubiquitous natural compound within the plant kingdom, could be instrumental in the protective measures against AD, with the potential involvement of CD33dim HLA-DR+ CD11b− myeloid cells, specifically pointing towards plasmacytoid dendritic cells (pDCs), in this mechanism. 73 4-hydroxycoumarin exhibits a spectrum of biological activities, including anti-inflammatory, antioxidant, anticoagulant, and antibacterial effects.74,75 Although no direct correlation between 4-hydroxycoumarin and AD has been documented, its anticoagulant function suggests a possible novel method for decelerating AD progression by reducing thrombotic risks and ameliorating cerebral microcirculation. Given that cerebrovascular diseases and blood flow modifications are recognized factors in the advancement of AD, 4-hydroxycoumarin may positively modulate these elements through its anticoagulant attributes. 76 Nonetheless, there is a paucity of direct evidence delineating the specific therapeutic or preventative mechanisms of 4-hydroxycoumarin in AD. As key immunomodulators, pDCs secrete Type I interferons, a cytokine with the capacity to quell inflammatory responses and regulate the broader immune system. 77 Thus, pDCs may exert indirect effects on the trajectory of AD by tuning the inflammatory environment. In aggregation, we postulate that 4-hydroxycoumarin might augment the immunoregulatory role of pDCs, indirectly impacting AD treatment through enhanced blood circulation. Nevertheless, the intricate mechanisms of 4-hydroxycoumarin and pDCs in AD are in the early stages of exploration and require further in-depth basic and clinical research to uncover their interrelationships and therapeutic applicability. These studies will be instrumental in the comprehensive assessment of the value of 4-hydroxycoumarin and pDCs in AD therapeutics and will provide a scientific basis for the development of future treatment modalities.
Glycolithocholate sulfate, a bile acid conjugate, is a fundamental component of the secondary bile acid metabolism, exerting a significant influence on lipid absorption within the digestive process. The liver synthesizes primary bile acids, which are then further altered by the gut microbiota. 78 This compound is crucial for modulating the gut microbiota balance, enhancing an intestinal milieu that promotes the growth of beneficial microbes such as Bifidobacteria and Lactobacilli, which are pivotal for the generation of short-chain fatty acids essential for gut integrity.79,80 Additionally, glycolithocholate sulfate contributes to the suppression of detrimental bacterial strains, thereby aiding in the sustenance of a diverse gut microbiota. Glycolithocholate sulfate plays an indispensable role in the activation of intestinal immune cells, particularly dendritic cells and T cells. 81 Dendritic cells, as proficient antigen-presenting entities, are responsible for the sequestration and processing of antigens in the gut, followed by migration to the mesenteric lymph nodes to stimulate T cell activation, a crucial step for the immune response. 82 HLA-DR (MHC class II molecules), predominantly expressed on dendritic cells and macrophages, functions to present antigens to CD4+ T cells, initiating an immune cascade. It is noteworthy that CD8+ T cells may also exhibit HLA-DR under certain conditions, typically associated with chronic infections or autoimmune disorders. 83 In the AD context, the expression of HLA-DR on CD8+ T cell might be correlated with the genesis of neuroinflammation. 84 These cells possess the ability to recognize and respond to aberrant brain proteins, such as amyloid-beta and tau, potentially participating in their clearance or exacerbation of inflammatory responses.61,85 Our research indicates that glycolithocholate sulfate could act as a safeguard against the advancement of AD, with the engagement of HLA-DR on CD8+ T cell assuming a critical role. These discoveries not only enhance our comprehension of the pathophysiological underpinnings of AD but also present opportunities for the development of novel therapeutic interventions targeting the gut microbiota-immune system axis. However, it is essential to recognize that these findings are preliminary and warrant further exploration across broader subject populations and with varied research methodologies to ascertain their scientific validity and generalizability.
This scholarly article provides a comprehensive examination of the critical role that immune cells play in the metabolic influences on the progression of AD, illuminating the intricate relationship between metabolic dysregulation and AD pathogenesis, and highlighting the pivotal function of immune cells within this complex interplay. Utilizing GWAS data and employing a bidirectional Mendelian Randomization analytical strategy, this study has revealed significant correlations between specific metabolites and immune cells. These findings offer novel perspectives for dissecting the immunopathological mechanisms at the heart of AD. While the study has achieved significant advancements, it acknowledges inherent limitations. The focus has been predominantly on established metabolites, including amino acids, lipids, and energy metabolism products, with less exploration of other potentially influential metabolites in AD. The precise mechanisms through which metabolites modulate immune cell functionality, whether directly or indirectly, remain obscure and demand more intensive and rigorous research. Although the study has applied a variety of methods to assess immune cell characteristics, there is an identified need to broaden the diversity and representativeness of the samples to ensure the wider applicability of the conclusions. Looking forward, several avenues of research are ripe for further exploration. It is crucial to broaden the scope of research to include a wider array of metabolite classes and to involve a more extensive and diverse sample cohort, which is essential for a more comprehensive understanding of the metabolomic profile in AD. A deeper elucidation of the molecular mechanisms underlying the interaction between metabolites and immune cells is fundamental to unraveling the etiology of AD and to constructing a robust theoretical framework for the development of innovative therapeutic approaches. Moreover, investigating the causal relationships between metabolites, immune cells, and AD may uncover potential biomarkers and therapeutic targets. Ultimately, conducting clinical trials to empirically substantiate the therapeutic efficacy of metabolites and immune cells in practical settings is crucial for devising strategies aimed at decelerating or halting the progression of AD.
Through the meticulous identification of the intricate connections between metabolites and immune cells that are intimately linked to AD risk, our analysis has significantly advanced our comprehension of the intricate pathophysiology of AD. These discoveries not only suggest potential new therapeutic targets but also pave the way for the development of innovative, targeted treatments for AD. Moreover, the variations in these metabolites and immune cells hold the promise of serving as biomarkers for the prediction of AD risk and facilitating early diagnosis, which is crucial for enhancing patient management and refining treatment strategies.
Conclusion
To encapsulate, this scholarly work offers profound insights into the pivotal function of immune cells amidst the metabolic influences on the progression of AD, as elucidated through GWAS data and a sophisticated bidirectional MR analysis strategy. The study uncovered notable correlations between specific metabolites and immune cells, with the histidine/pyruvate ratio and homoarginine exhibiting positive correlations linked to an elevated risk of AD, and 4-hydroxycoumarin and glycolithocholate sulfate showing negative correlations, suggesting a protective role against the disease. These findings accentuate the critical involvement of immune cells in AD pathogenesis and provide fresh vantage points for understanding the underlying immunopathological mechanisms. Further analysis of the study posits immune cells as conduits interlinking metabolites and AD, identifying specific immune cell subsets, such as CD64 on monocyte and immune responses on HLA DR on CD14+ CD16− monocyte, as potentially pivotal in the disease’s pathogenesis by modulating the metabolic and functional attributes of metabolites. The expression within CD33dim HLA DR+ CD11b− myeloid cell and on HLA DR on HLA DR+ CD8+ T cell may also be implicated in protective mechanisms against AD, unveiling new directions for the development of advanced therapeutic strategies.
Supplemental Material
sj-docx-2-alz-10.1177_13872877241313140 - Supplemental material for Immune cells: Mediators in the metabolites and Alzheimer’s disease
Supplemental material, sj-docx-2-alz-10.1177_13872877241313140 for Immune cells: Mediators in the metabolites and Alzheimer’s disease by Erdong Zhang, Fengqiu Dai, Ling Tao, Yanqin Chen, Tingting Chen and Xiangchun Shen in Journal of Alzheimer's Disease
Supplemental Material
sj-pdf-3-alz-10.1177_13872877241313140 - Supplemental material for Immune cells: Mediators in the metabolites and Alzheimer’s disease
Supplemental material, sj-pdf-3-alz-10.1177_13872877241313140 for Immune cells: Mediators in the metabolites and Alzheimer’s disease by Erdong Zhang, Fengqiu Dai, Ling Tao, Yanqin Chen, Tingting Chen and Xiangchun Shen in Journal of Alzheimer's Disease
Footnotes
Acknowledgments
We sincerely thank the Finngen project for providing the key dataset for Alzheimer’s disease, which is crucial for the progress of our research. The authors are grateful to the original studies for sharing the GWAS data on immune cells and metabolites used in this project.
Author contributions
Erdong Zhang (Conceptualization; Data curation; Formal analysis; Project administration); Fengqiu Dai (Conceptualization; Project administration; Writing – original draft); Ling Tao (Data curation; Formal analysis); Yanqin Chen (Data curation; Formal analysis); Tingting Chen (Writing – original draft); Xiangchun Shen (Writing – review & editing).
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by the Doctoral Scientific Research Foundation of Guizhou Medical University [grant number YJ20066], the National Natural Science Foundation of China Cultivation Project of Guizhou Medical University [grant number 20NSP051], the Scientific and Technological Project of Guizhou Province [grant number ZK (2022)398, ZK (2023)336], and the Fund of Science and Technology Project of Guizhou provincial health commission [grant number Gzwkj2023-564].
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data availability
The data supporting the findings of this study are openly available in Finngen database (https://www.finngen.fi/en) and GWAS Catalog (![]() ).
).
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.