
Abstract
Background
Sex differences in the brain's structure, function, and response to interventions are increasingly recognized as critical factors in neurodegenerative diseases like mild cognitive impairment (MCI) and Alzheimer's disease.
Objective
This study investigated sex differences in brain activity among participants classified as having MCI (based on Montreal Cognitive Assessment scores) during exposure to a 40 Hz rhythmic light (RL) intervention versus a control RL condition.
Methods
Twenty-four participants in the MCI group (mean age = 74.5 years, SD = 9.06; 15 women, 9 men) and 16 age-matched healthy controls (mean age = 70 years, SD = 8.97; 9 women, 7 men) were exposed to both light conditions and underwent electroencephalography recordings, cognitive performance testing (2-back task), and a subjective assessment of sleepiness (Karolinska Sleepiness Scale).
Results
40 Hz RL increased gamma power significantly, indicating its potential to enhance brain function, especially in the MCI group. Men showed greater neural response and better cognitive performance under the 40 Hz RL while women (particularly in the MCI group) responded more strongly to the control. The sleepiness results were not significant.
Conclusions
There is compelling evidence that 40 Hz RL has sex-dependent effects on gamma oscillations and cognitive performance, particularly in individuals with MCI. Men may benefit more directly from gamma-enhancing interventions, whereas women may require alternative approaches that account for their unique neural responses to light. Further research with larger, more diverse populations is needed to confirm these findings and explore the potential for personalized therapeutic strategies based on sex and cognitive status.
Introduction
Brain oscillations, or rhythmic fluctuations in neural activity, play a critical role in cognitive processes and have been extensively studied in mild cognitive impairment (MCI). In individuals with MCI, disruptions in brain rhythms appear across several frequency bands, reflecting early neural alterations linked to cognitive decline.1,2 In healthy adults, sex differences in brain oscillations are typically modest, but in pathological states like MCI these differences become more pronounced. This could be due to protective factors or genetic susceptibilities that vary between sexes. 3
Light flicker stimulation is emerging as a promising non-invasive approach to enhance neural oscillations associated with cognition and alertness, particularly in individuals with MCI. White light flicker has been shown to entrain neural activity and modulate cognitive performance. 4 More specifically, studies indicate that 40 Hz flickering light can enhance neural entrainment and behavioral alertness in both animal models and humans.5–7 It is not known, however, whether the circadian system modulates some of these observed effects of white light flicker. The use of red light flicker, particularly at 40 Hz, will not affect the circadian system while still promoting gamma oscillatory activity linked to cognitive processing.5,6,8
Gamma oscillations play a crucial role in supporting higher cognitive functions, as they facilitate neural synchronization across brain regions, enabling efficient communication and processing of information.9,10 Likewise, theta oscillations are central to memory, learning, and attentional control. In the context of cognitive dysfunction, disruptions specifically in the 40 Hz gamma range have been associated with MCI, suggesting that these oscillations may serve as early markers of cognitive decline.5,11 Sex differences in these oscillations have also been observed, with one study suggesting that men have higher resting theta power 12 and others reporting stronger theta activation in women, possibly reflecting heightened arousal or engagement during cognitive tasks.13–15 Factors such as hormonal influences, structural brain differences, and functional connectivity patterns between sexes might contribute to distinct neural responses to rhythmic sensory stimulation. Understanding these sex-specific neural dynamics is crucial for tailoring cognitive stimulation therapies effectively.16–19 For instance, a study examining sex differences in amygdalohippocampal oscillations found that women displayed more prominent basal gamma activity compared to men, suggesting inherent differences in neural processing mechanisms. 20 Additionally, research on resting-state brain activity revealed that aging affects neural oscillations differently between sexes, with women showing higher theta event-related oscillation responses than men in certain brain regions during early adulthood, though these differences diminish with age. 21 These variations in neural oscillations could influence how each sex responds to interventions like 40 Hz flickering light.
The therapeutic potential of 40 Hz flicker stimulation was first demonstrated by Iaccarino et al., who found that exposure to visual gamma flicker reduced amyloid-beta deposition and improved neural synchrony in Alzheimer's disease models. 6 This work laid the foundation for investigating gamma-frequency stimulation as a non-invasive neuromodulatory approach for cognitive decline.7,8,19,20,22–25 While our study focuses on visually driven gamma entrainment, other forms of gamma-frequency-based stimulation, such as transcranial alternating current stimulation (tACS), transcranial magnetic stimulation (TMS), and photobiomodulation, have also been explored for their cognitive benefits. TMS and tACS can directly induce gamma oscillations in cortical networks, improving cognitive function by modulating neural excitability and connectivity.26–29 Similarly, photobiomodulation, particularly in the near-infrared spectrum, has shown promise in enhancing mitochondrial function and neuroprotection, albeit through different mechanisms than rhythmic flicker.30–32 However, visual 40 Hz stimulation engages the retinogeniculate-cortical pathway and has been shown to entrain cortical gamma-band activity through sensory pathways, producing oscillatory responses that are time-locked but not equivalent to spontaneously generated (endogenous) gamma rhythms. This sensory-driven entrainment provides a non-invasive means to modulate gamma-frequency network dynamics and offers a mechanistically distinct alternative to direct cortical stimulation.
Studies on working memory deficits in MCI also reveal distinct patterns between sexes. Women with MCI typically experience sharper declines in working memory,33,34 likely due to postmenopausal estrogen reduction, which may make them more vulnerable to these deficits. Estrogen's role in modulating dopamine pathways may also contribute to these slower reaction times and reduced accuracy in working memory tasks. 35 Men, meanwhile, tend to retain resilience in certain memory tasks but are often more vulnerable to deficits in spatial working memory, possibly due to structural and neurochemical differences. 36 These sex-based disparities extend beyond task performance and also impact reaction times and accuracy.
This preliminary study is a subset analysis of a study examining the broader effects of 40 Hz RL intervention in patients with MCI (National Institute on Aging, ORS-6195 GCO 21-0378-01). While the present contribution focuses on the critical aspect of sex differences in response to 40 Hz RL, the parent study is investigating the long-term therapeutic potential of this intervention in a population of MCI patients. The purpose of this subset analysis was to investigate the sex differences in response to 40 Hz RL intervention and thereby permit the tailoring of therapeutic responses between sexes.
Methods
Participants
The study's 40 participants included 24 individuals classified as having MCI based on their Montreal Cognitive Assessment (MoCA) scores (mean age = 74.5 years, SD = 9.06; 15 women, 9 men) and 16 age-matched healthy controls (mean age = 70 years, SD = 8.97; 9 women, 7 men). A power analysis based on pilot data determined that a sample size of 40 participants (accounting for 30% attrition) provides 95% power to detect significant differences in paired treatment conditions (active versus control rhythmic light) using a linear mixed-effects model. Given the within-subjects design, this sample size is sufficient to assess changes over time within each group (healthy controls and MCI). Aim 2, which is not reported in this manuscript, expands on this design with a planned sample size of 120 participants to evaluate treatment effects over an extended intervention period.
Informed consent was obtained from all participants in accordance with experimental protocols approved by the Institutional Review Board of the Icahn School of Medicine at Mount Sinai, New York. All participants were aged ≥55 years and free from sleep disturbances, as determined by Pittsburgh Sleep Quality Index (PSQI) scores <5. 37 The PSQI includes an item assessing breathing difficulties during sleep. Those reporting breathing difficulties underwent further evaluation using the Sleep Apnea Disorder Questionnaire (SDQ-SA). The PSQI has been extensively validated as an effective tool for screening sleep disturbances and was instrumental in confirming that participants met the required sleep health criteria for inclusion. Furthermore, the PSQI does not directly assess Activities of Daily Living (ADL); however, it includes items related to daytime dysfunction, such as difficulty staying awake during daily activities or reduced enthusiasm to complete tasks, which may indirectly reflect functional impairments. Inclusion in the MCI group was based on MoCA scores ranging from 17 to 25. 38 Controls had MoCA scores > 25.
All participants, including both control subjects and those in the MCI group, underwent a thorough phone screening to assess their initial eligibility. Additionally, participants in the MCI group were carefully evaluated during the screening to confirm their diagnosis and eligibility for the study. This screening reviewed self- or caregiver reports of each participant's medical history, current and previous treatment regimens, and current medication use. Excluded from the study were those with suicidal tendencies, recent psychotropic medication changes, major organ failure, uncontrolled disorders like hypertension or diabetes, obstructive vision conditions, prior cataract surgery with specific lenses, severe sleep apnea, restless leg syndrome, or the presence of another brain disease accounting for their cognitive status. Those with histories of severe epilepsy, photosensitivity dermatitis, progressive retinal disease, or permanently dilated pupils were also excluded. Furthermore, individuals with a history of alcohol use disorder or current excessive alcohol consumption were excluded. Alcohol intake was not explicitly measured, though the PSQI includes a question that allows participants to report other factors contributing to poor sleep. No participants reported alcohol consumption to aid sleep. There were no biological markers collected for this study aim.
Study design
This two-week study exposed participants to two experimental conditions for 60 min over the course of two sessions. The sessions took place on two different weeks, each preceded by a one-week period during which participants recorded a diary of their sleep and wake times.
Experimental conditions
The study's two experimental conditions (i.e., 40 Hz RL and control RL condition) were delivered via custom-built luminaires housing three light-emitting diode (LED) strings (153 total LEDS) housed in a 50 cm × 10 cm × 6.4 cm (h × w × d) polyvinyl chloride enclosure covered by a frosted acrylic diffuser. The bottom edge of the luminaire was elevated 20 cm above the surface of the table facing the seated participants, 60 cm from their eyes. The LED strips were driven by a programmed microcontroller (Uno model, Arduino, New York, NY, USA) and the temporal pattern of the RL stimuli was driven by a second microcontroller (BuckPuck 3023-D-E-700 model, LEDdynamics, Randolph, VT, USA). Both devices were virtually identical, and the participants were not informed of which condition they were experiencing. The active 40 Hz RL intervention presented the stimulus at a frequency of 40 Hz (set as the square wave of a 50% duty cycle, 12.5 ms on and 12.5 ms off). The control RL presented the stimulus at randomly varying frequencies of 30 Hz and 50 Hz, specifically avoiding the frequency of 40 Hz. Both the 40 Hz RL and control RL emitted circadian-ineffective red light (λpeak = 620 nm), which lies well beyond the human circadian system's peak spectral sensitivity of 460 nm.39,40
This light spectrum was selected to avoid evoking a circadian system response at the targeted illuminance level of 60 lx (measured at participants’ eyes), thereby limiting any observed effects to the stimulus's frequency and not to the spectral properties of the light itself.
Experimental protocol
Both participant groups were randomly assigned to start the experiment with one of the two experimental conditions (either the 40 Hz RL or the control RL) followed by the other in counterbalanced order (Figure 1). All participants experienced both conditions. Each session started at 14:00 with a 10-min adaptation to dim ambient room lighting providing an illuminance of 15 lx measured at the participants’ eyes. The session time was based on our previous work (Sahin and Figueiro). 5 Additionally, afternoon scheduling improved participant compliance by accommodating work and other commitments, resulting in higher data quality and reliability. The adaptation period was in turn followed by a 15-min baseline assessment of the study's behavioral measures: working memory (2-back task task) and subjective sleepiness (Karolinska Sleepiness Scale [KSS] questionnaire). The 60-min RL conditions were then delivered in the otherwise darkened room, followed by a second assessment of working memory and subjective sleepiness. Both behavioral measures assessments were conducted in the dim room lighting employed for the 10-min adaptation period (see Figure 1). EEG was recorded at three time points: before (T1), during (T2) the light exposure; and post light exposure (T3), however, no EEG data were collected during the working memory task itself.

The experimental procedure, showing the room lighting during the experiment and the timing of the light exposure period, the EEG recordings (T1–T3), and the administration of the behavioral measures (working memory [2-back task] and subjective sleepiness [KSS questionnaire]). EEG was recorded at two time points: before (T1) and during (T2) the light exposure; however, no EEG data were collected during the working memory task itself. A third post-exposure EEG recording taken at T3 (grayed out in this figure) was not included in the present analyses.
EEG recordings were performed at three time points (T1–T3) per session: T1 at baseline (before the light exposure) and before administration of the WM and KSS, T2 during the final 5 min of the light exposure, and T3 at the session's end (after the light exposure, but before the administration of WM and KSS). Thus, no EEG recordings were obtained during the KSS or WM administration. WM and KSS were administered before the T1 time point and again after the light intervention, but prior to the T3 time point. To accurately assess sex differences in response to the 40 Hz RL, we analyzed the change in electrophysiological activity between T1 (baseline) and T2 (during the light exposure) only. Focusing on this difference permitted us to isolate the light interventions’ effects more precisely, as it would account for individual variability in pre-intervention neural activity and ensure the observed changes could be directly attributed to the 40 Hz RL. This method also minimized post-exposure confounds (e.g., fatigue or external factors) that might have influenced the recording at T3.
EEG data collection and processing
The EEG recordings were obtained using the Brain Vision actiCHamp 32 channel cap, which employs electrode placement using the 10–20 system (Supplemental Figure 1). The EEG signal obtained during each experimental session was sampled using a 32-electrode EEG amplifier (actiCHamp 32, Brain Vision, Garner, NC, USA) and impedances were checked to remain below 5 kΩ with a 2500 Hz sampling rate and a band pass filter applied in the range of 0.15–200 Hz. Offline data processing was performed using EEGLAB, an open-source MATLAB toolbox (MATLAB R 2020a, The Math Works Inc. Natick, MA, USA). Data were resampled to 128 Hz, and an additional band-pass filter was applied in the range of 0.1–55 Hz. The processed data were then plotted and carefully inspected by a trained experimenter for the manual rejection of artifacts (e.g., eye blinks, eye movement, teeth clenching, body movement, channel noise, electrocardiogram artifacts). The default reference electrode is placed at FCz according to the 10–20 EEG system. This placement minimizes eye movement (electrooculographic) artifacts, which are more prominent at frontal sites, and reduces muscle (electromyographic) contamination from the jaw and neck. Additionally, FCz provides a balanced signal representation, as it is equidistant from most electrodes, ensuring a neutral and symmetrical reference point across the scalp.
Average Fourier cross-spectral matrices were computed for the following frequency bands: delta (2–3 Hz), theta (4–7 Hz), alpha (8–12 Hz), beta1 (13–18 Hz), beta2 (19–21 Hz), beta3 (22–30 Hz), and gamma (31–44 Hz). PSD plots and representative raw traces were also created for 40 Hz RL, the control RL condition, and a no-stimulation baseline. This comparison highlights each condition's spectral signature and the neural engagement induced by the RL.
Behavioral measures
Working memory
In the 2-back task 41 participants were presented with a visual sequence of uppercase letters (i.e., total of 15 letters in the set) displayed one by one (named “trials”) on a 15-inch laptop computer screen. Each trial was presented for a maximum of 760 ms, and participants were given 2000 ms (the intertrial interval) to decide whether the present trial matched the trial presented two trials ago. If the two trials matched, participants pressed the “m” key on the computer keyboard as instructed prior to the experimental session. Accurate responses were highlighted in green and inaccurate responses were highlighted in red for the remainder of the trial. Each assessment period was composed of three blocks of 15 trials (i.e., 45 total trials per period), and the following letters were used in all test blocks: A, B, C, D, E, H, I, K, L, M, O, P, R, S, and T. The 2-back task outcomes included reaction time (in milliseconds) and total accuracy (percentage of correct responses).
Subjective sleepiness
Participants reported KSS 42 scores twice per session, at baseline and following the light exposure (see Figure 1). Participants indicated how sleepy or alert they felt on a scale ranging from 1 to 9, where 1 represented “very alert,” 3 represented “rather alert,” 5 represented “neither alert nor sleepy,” 7 represented “sleepy, but no difficulty remaining awake,” and 9 represented “very sleepy, fighting sleep, an effort to remain awake.”
Data and statistical analysis
Power spectral density (PSD) analysis
The artifact-free data obtained from the processing step were analyzed using a fast Fourier transform (averaged window of 5 s with 50% overlap) to calculate power (µV²) for all frequency bands. All power values in the Supplemental Material are given in µV² values. The initial PSD signal distribution along the range of frequencies of the brain activity was computed using the Spectopo function as implemented in EEGLAB using Welch's method 43 with a window length of 512 points, a (f) length of 1024 points, and no overlapping. The power was then calculated for 32 channels and the seven frequency bands noted above. An additional electrode, POZ, was derived by averaging the values from electrodes Pz and Oz, resulting in 33 electrode measures.
In this analysis, we examined each electrode individually in addition to evaluating the overall average across all electrodes to account for the spatial specificity of brain activity. EEG signals can vary across different scalp regions, reflecting underlying neural processes associated with distinct cognitive functions. Focusing on individual electrodes permits the capture of localized effects that might be masked in a global average. Significant changes at specific electrodes provide valuable insights into targeted areas of brain activity allowing the mapping of intervention effects to specific neural regions. This approach is widely applied in EEG research to discern focal changes in brain activity, offering a detailed perspective on the intervention's impact across different cortical areas.
In addition, given the potential impact of prolonged light stimulation on EEG power spectral characteristics, we examined changes in the aperiodic (1/f) component of the EEG signal to assess whether fatigue after 55 min of RL exposure influenced spectral offset and exponent. Specifically, we aimed to determine whether observed changes in the PSD were due to broadband spectral shifts or genuine neural oscillatory activity. To achieve this, we employed a 1/f decomposition analysis using a MATLAB-based algorithm to extract the exponent and offset of the power spectrum. To quantify 1/f slope of the EEG PSD, for each dataset, the PSD was computed via the Spectopo function, yielding power values across frequencies. These power values were converted from decibels to linear scale, and both the frequency and power data were transformed to logarithmic scales. A linear regression was then performed on the log-transformed data, with the slope of the resulting line representing the 1/f component of the PSD. This procedure was repeated for all datasets, and the individual slopes were averaged to obtain a group 1/f slope. Additionally, a group-averaged log-log PSD plot was generated, displaying PSD data points alongside the fitted 1/f line. This approach allowed us to isolate oscillatory power changes from aperiodic spectral contributions and evaluate whether differences in gamma-band activity (40 Hz) were driven by specific neural entrainment rather than global spectral scaling effects. We analyzed data separately for each group (healthy controls, MCI) under both the 40 Hz RL and control RL conditions.
Electrode clusters were determined based on the topographical distribution of neural responses to 40 Hz rhythmic visual stimulation. Given that gamma-band entrainment occurs primarily in early visual cortical areas, we identified regions of interest over occipital and parietal regions, where, based on reports from the literature, strongest phase-locked responses to flicker stimulation were expected.44,45 Cluster-level correction was performed using the 8 anatomical regions (Frontal, Fronto-central, Central, Temporal, Centro-parietal, Parietal, Parieto-occipital, and Occipital). For each of the 33 electrodes (32 electrodes + POZ) within the cluster, the absolute value of the mean 40 Hz power was calculated at both baseline (T1) and during stimulation (T2). This approach was used to ensure consistency in the magnitude of the power estimates across participants. To quantify changes in response to each light condition while accounting for individual variability, the difference in absolute mean 40 Hz power between T2 and T1 was computed for each participant and electrode (i.e., T2 – T1). The resulting difference scores, which could be positive or negative, reflect the direction and magnitude of change from baseline and served as the dependent variable in the linear mixed-effects models. Light condition (40 Hz RL versus control RL) was assessed in the linear mixed models as the main exposure variable. The difference in 40 Hz power between T1 and T2 was modeled while evaluating interactions between light condition, sex, and group, adjusting for age. Thus, rather than analyzing each light condition separately, the model directly compared 40 Hz RL to control RL to assess their differential effects on 40 Hz power changes. Since multiple comparisons were already addressed at the cluster level, no additional adjustments were applied at the individual electrode level to avoid excessive Type II errors. Statistical significance was accepted at p < 0.05. Analyses were conducted in R (R version 4.4.1).
Source construction, whole brain analysis
Standardized low resolution electromagnetic tomography (sLORETA) 46 was employed to calculate standardized current density across 6239 voxels in the gray matter and hippocampus of the Montreal Neurological Institute reference brain. This method offers robust localization of potential neural sources based on a linear weighted sum of scalp-recorded electrical potentials. sLORETA operates under the assumption that adjacent voxels exhibit maximally similar electrical activity, thereby enhancing spatial accuracy in source estimation. 47 Images generated by sLORETA, with a spatial resolution of 5 mm, provided comparative data on sex differences within MCI and healthy control groups. Voxel-wise randomization tests, built into sLORETA and utilizing 5000 permutations, were conducted for statistical non-parametric mapping. These tests were corrected for multiple comparisons to improve the reliability of findings. The following comparisons were conducted in whole-brain analyses: (1) sex differences regardless of group affiliation (MCI or control), (2) sex differences in response to 40 Hz RL, and (3) sex differences in response to the control RL. The threshold for statistical significance was set at p < 0.05.
It is important to note that, in our analysis, we did not include a 20 Hz steady-state visual evoked potential (SSVEP) in the control RL condition. Instead, the control RL consisted of 30 Hz and 50 Hz light flicker. Neither of these frequencies produce a direct harmonic at 40 Hz: 30 Hz stimulation does not generate a 40 Hz harmonic, as its primary harmonic would be at 60 Hz; likewise, 50 Hz stimulation also does not introduce a 40 Hz component, with its primary harmonic occurring at 100 Hz. By ensuring that 20 Hz was not used in the control RL condition, we effectively mitigate the risk of spurious 40 Hz harmonics confounding the analysis. Furthermore, each control frequency was analyzed separately to avoid averaging effects that could obscure frequency-specific responses. This approach strengthens the validity of our findings by ensuring that any 40 Hz response observed is specific to the 40 Hz intervention and not an artifact of harmonic contamination.
Behavioral measures
The differences in baseline (T1) versus post-exposure (T3) values for working memory reaction time, working memory accuracy, and subjective sleepiness (KSS scores) were calculated to evaluate changes (expressed as means and standard deviation [SD]) in response to the two light conditions. Because the measures were repeated within subjects, linear mixed models were used to assess whether these differences varied by light condition. Possible interactions between light condition, sex, and group were also assessed. All models were adjusted for age. Statistical significance was accepted at p < 0.05. Analyses were conducted using R (R version 4.4.1).
Results
Power spectral density
Difference in 40 Hz power between the two light conditions
All 33 electrodes were analyzed. However, C4 was selected for reporting as it exhibited the most significant findings. There was a significant difference in 40 Hz power between the two light conditions at the C4 electrode site, with an increase in power density under the 40 Hz RL compared to the control RL (mean difference, 40 Hz RL versus control RL: 2.52 [SD 10.9] versus −2.50 [SD 7.7], p = 0.046, Figure 2). No other significant differences were identified (Supplemental Table 1).
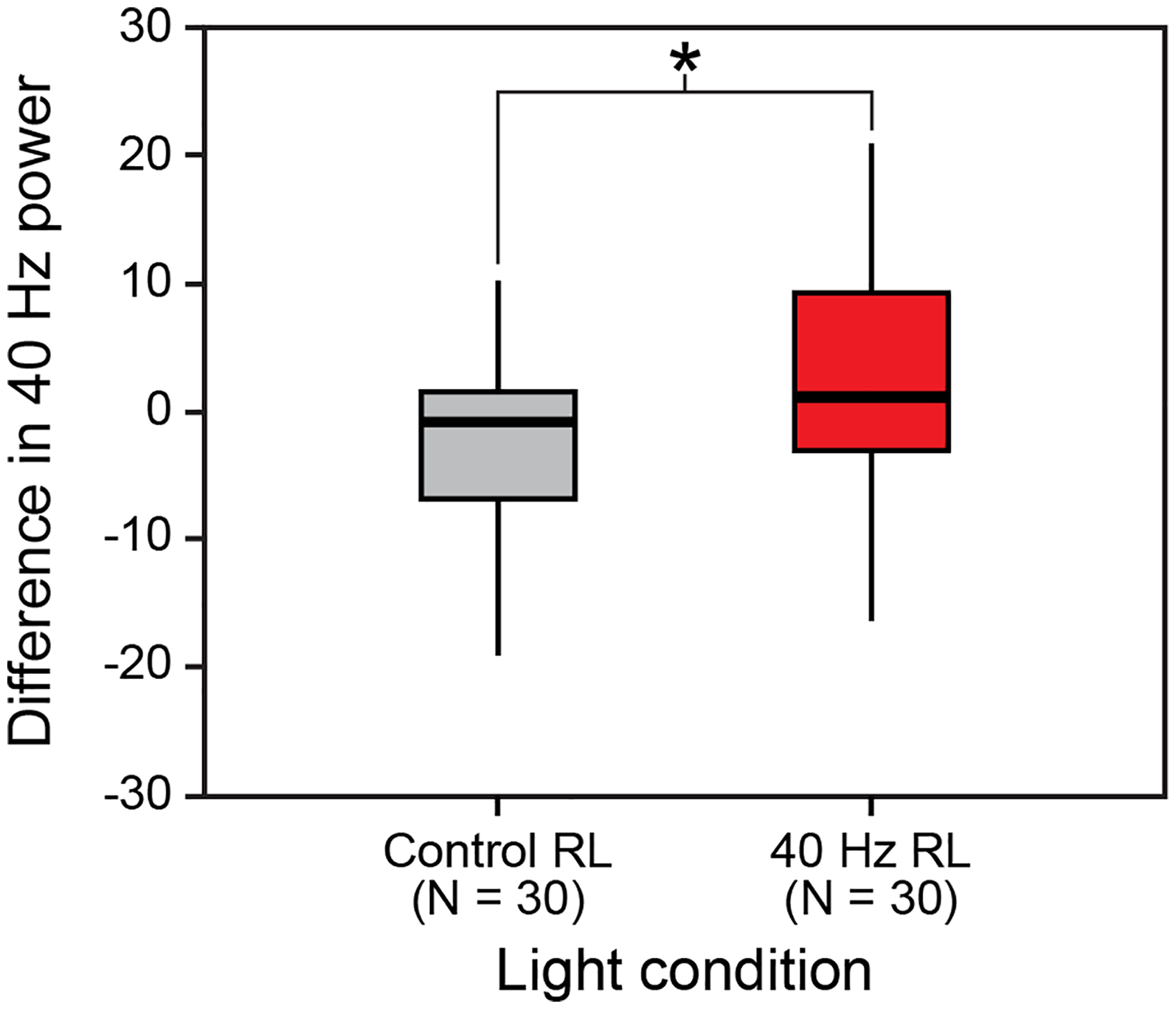
Differences in 40 Hz power at electrode C4 by light conditions.
Below are the PSD traces of the baseline condition (Figure 3(a)) (pre intervention), and during the 40 Hz RL (Figure 3(b)) and control RL (Figure 3(c)) for all subjects. As expected, the dominant spectral component appears in the low-frequency bands, particularly in the theta (4–8 Hz) and alpha (8–12 Hz) ranges, which are characteristics of resting state brain activity. During the 40 Hz RL condition, a clear increase in spectral power at 40 Hz is observed across both MCI and control participants (Figure 3(b)). Similarly, during the control RL condition, a clear increase in spectral power at 30 Hz and 50 Hz is observed (Figure 3(c)). The comparison highlights the effectiveness of rhythmic light in modulating brain oscillations.
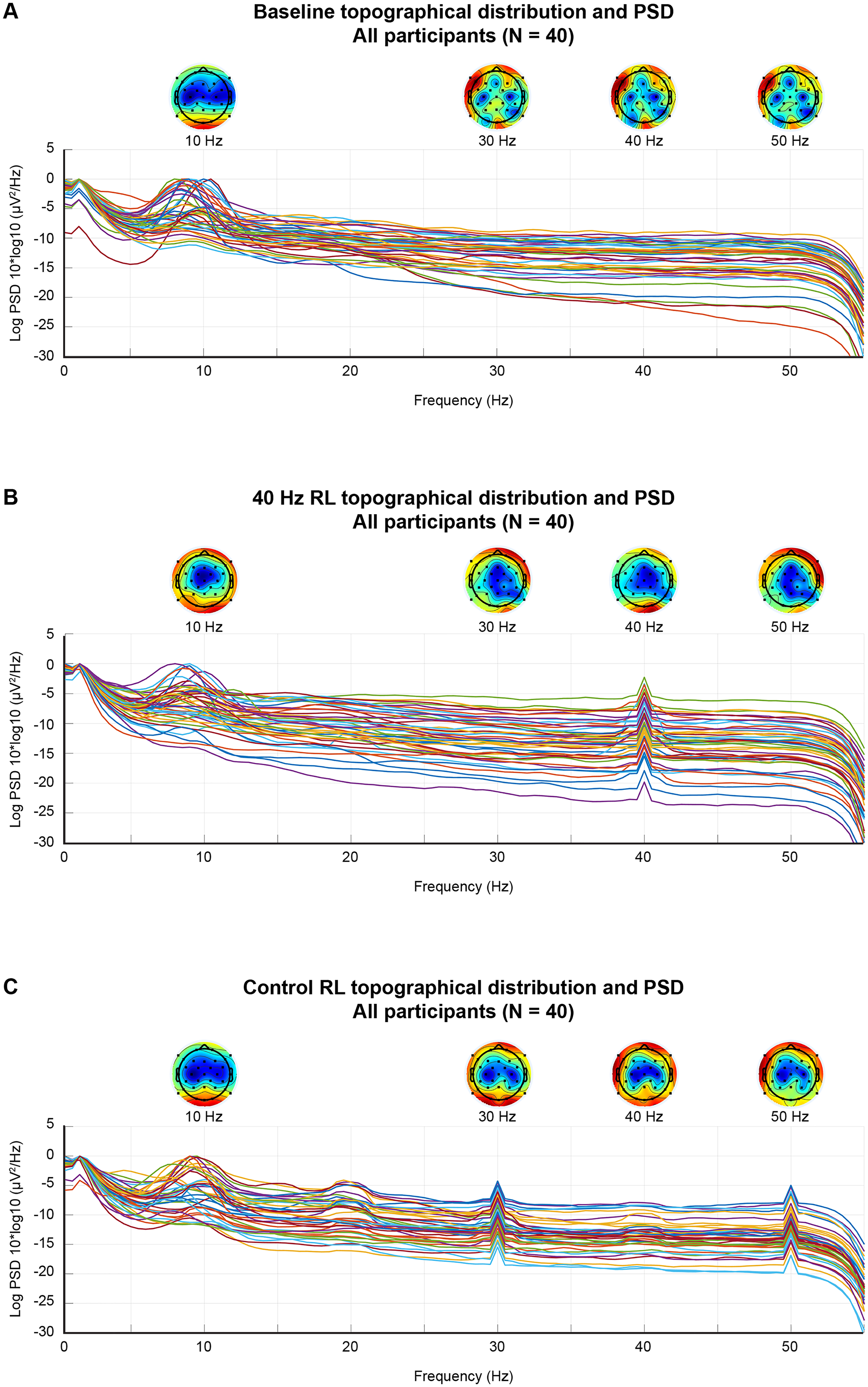
Power spectral density (PSD) and scalp topography at: (a) baseline, (b) during the 40 Hz RL condition, and (c) during the control RL condition. The PSD logs display mean log-transformed power spectra across all subjects. Topographic maps represent scalp power distribution at 10 Hz (alpha), 30 Hz (low beta), 40 Hz (gamma), and 50 Hz (high gamma), aligned above their corresponding frequency bins. Power is averaged across EEG channels and subjects.
1/f analysis
Figure 4(a) and (b) display the log-log PSD plots with extracted 1/f slopes under both light conditions in the healthy control group. Under 40 Hz RL, the 1/f slope was −0.28118 (Figure 4(a)), whereas in the control RL condition, the 1/f slope was −0.2478 (Figure 4(b)). There was a slightly steeper 1/f slope in the 40 Hz RL condition suggesting greater attenuation of higher frequencies relative to lower frequencies. However, the difference between conditions remains modest, indicating that fatigue is unlikely to be a confounding factor. If fatigue were a primary driver of spectral changes, a progressive steepening of the 1/f slope across time would be expected. Instead, the control RL condition exhibited a shallower 1/f slope, suggesting a flatter power spectrum compared to the 40 Hz RL condition, possibly reflecting differential brain activity modulation under different stimulation conditions.

Aperiodic spectral structure (1/f) in control and MCI subjects during rhythmic and control light exposure. (a–d): group averaged PSD plotted on log-log axes to characterize the aperiodic (1/f) component of neural activity during light stimulation. (a) 1/f spectral slope in control subjects during 40 Hz RL condition, a marked deviation from the 1/f trend is observed at 40 Hz, consistent with frequency-specific entrainment, (b) under control RL, control subjects exhibit a smooth 1/f spectral profile with no prominent frequency-specific deviations, (c) in subjects with MCI, 40 Hz RL results in a subtle flattening of the 1/f slope near the gamma range, suggestive of attenuated neural entrainment, (d) MCI subjects under the control RL show a typical 1/f distribution, consistent with a lack of frequency-specific engagement.
Figure 4(c) and 4(d) show the PSD plots for MCI participants. Under 40 Hz RL, the 1/f slope was steeper at −0.35478 (Figure 4(c)), compared to −0.24446 in the control RL condition (Figure 4(d)). This substantial difference suggests stronger suppression of broadband power under 40 Hz RL, indicating increased neural entrainment effects in MCI participants. The steeper slope under 40 Hz RL is indicative of a greater suppression of low-frequency activity (e.g., delta and theta bands), likely reflecting an altered compensatory mechanism in MCI. In contrast, the control RL condition produced a shallower slope, indicative of a broader spectral power distribution and reduced neural entrainment.
Table 1 provides a summary of the 1/f slope comparisons across conditions and groups.
1/f slope by group and condition. MCI participants showed the greatest broadband suppression under 40 Hz rhythmic light (RL), indicating strong neural entrainment. Slope values were shallower in control conditions and in healthy controls, reflecting weaker spectral modulation.

Differences in 40 Hz power across electrodes by light conditions and sex
There was a significant sex difference in 40 Hz power at the electrodes Fz (p = 0.028), F3 (p = 0.027), CP5 (p = 0.036), CP6 (p = 0.047), T8 (p = 0.004), Cz (p = 0.046), P4 (p = 0.010), F4 (p = 0.019), O2 (p = 0.021), C4 (p = 0.005) (Figure 5), with no significant variation by group. At these sites, men generally exhibited a greater increase in 40 Hz power density under the 40 Hz RL compared to control RL. In contrast, women showed either a slightly larger increase in power density with the control RL or a similar increase under both conditions. For example, at electrode T8, men showed a greater mean difference in 40 Hz power with 40 Hz RL versus the control RL (5.14 [SD 10.0] versus −10.4 [SD 16.5]), while women exhibited similar increases under both conditions (control RL: 1.63 [SD 12.5] versus 40 Hz RL: 1.13 [SD 11.4]). No other significant differences were identified (Supplemental Table 2).

Differences in 40 Hz power across electrodes that showed significant difference by light conditions and by sex.
A significant sex difference by group was observed at electrodes FC1 (p = 0.043), FC5 (p = 0.049), and Fp2 (p = 0.022) (Supplemental Table 3). The common trend across these electrodes indicates that men participants in the MCI group generally experienced greater increases in 40 Hz power under the 40 Hz RL compared to the control RL. In contrast, women and men healthy controls tended to show either smaller increases or even reductions in 40 Hz power with the 40 Hz RL relative to the control RL. Specifically, at electrode FC1, women in the MCI group showed similar reductions in 40 Hz power under both conditions (40 Hz RL: −2.42 [SD 9.22] versus control RL: −2.83 [SD 5.56]), whereas men in the MCI group demonstrated a greater reduction with the control RL compared to the 40 Hz RL (40 Hz RL: −2.68 [SD 14.7] versus control RL: −20.9 [SD 24.7]). In comparison, women healthy controls had minimal changes (40 Hz RL: −0.403 [SD 6.15] versus control RL: −0.0293 [SD 3.09]) and healthy control men also showed a smaller reduction (40 Hz RL: −5.62 [SD 5.38] versus control RL: −1.19 [SD 4.87]).
Furthermore, at electrode FC5, women in the MCI group had a slight decrease in 40 Hz power with the 40 Hz RL and a greater decrease under the control RL (40 Hz RL: −0.900 [SD 7.48] versus control RL: −2.79 [SD 14.6]). Men in the MCI group experienced an increase under the 40 Hz RL but a marked decrease with the control RL (40 Hz RL: 1.78 [SD 13.2] versus control RL: −15.1 [SD 20.8]) (Figure 6(a)). For the women healthy controls, the 40 Hz RL was associated with in an increase (40 Hz RL: 5.05 [SD 15.3] versus control RL: −2.42 [SD 2.80]), while men healthy controls showed a slight reduction under both conditions (40 Hz RL: −1.54 [SD 11.3] versus control RL: 1.25 [SD 6.01]). At electrode Fp2, women in the MCI group exhibited a mild reduction under the 40 Hz RL but an increase under the control RL (40 Hz RL: −1.38 [SD 8.47] versus control RL: 3.85 [SD 10.4]). Men in the MCI group had a significant increase under the 40 RL light and a large reduction with the control RL (40 Hz RL: 10.4 [SD 7.22] versus control RL: −8.57 [SD 25.9]). Women healthy controls showed minor reductions (40 Hz RL: −1.07 [SD 8.37] versus control RL: 1.09 [SD 3.91]). Men healthy controls showed a mild reduction with the 40 Hz RL and a small increase under the control RL (40 Hz RL: −2.25 [SD 9.86] versus control RL: 0.393 [SD 4.87]).

Statistical nonparametric maps showing voxel-wise differences in whole-brain activity between women and men, computed using standardized low-resolution electromagnetic tomography (sLORETA). Blue represents regions with significantly lower current density in women relative to men; red/yellow indicates higher current density in women. All comparisons are between women and men within each specific condition and group. Sex-based differences in cortical current density under rhythmic light (RL) stimulation across theta and gamma frequency bands. sLORETA results show frequency-specific differences in cortical activation patterns between women and men, under both 40 Hz RL and control RL, stratified by group and condition. (a) Under 40 Hz RL, the total sample of 24 women (compared to total sample of 16 men) demonstrated a significant decrease in theta activity irrespective of group (MCI or control). Decreased activity localized to the right medial frontal gyrus, posterior cingulate cortex, and paracentral lobule. (b) Under 40 Hz RL, 24 women (compared to 16 men) also showed a significant increase in gamma activity, primarily in the left lingual gyrus and parahippocampal gyrus. (c) Under control RL, 24 women (compared to 16 men) showed a decrease in theta activity especially in the left middle frontal gyrus, pre- and post-central gyri, inferior parietal lobule, and cingulate gyrus, across all participants. (d) In the MCI group under 40 Hz RL, 15 women exhibited lower theta activity than 9 men, localized to the right middle frontal gyrus, paracentral lobule, and posterior cingulate cortex. (e) In the MCI group under control RL, 15 women showed reduced theta activity compared to 9 men centered in the left precentral gyrus. (f) In the control group under control RL, 9 women showed increased gamma activity, relative to 7 men, primarily in the middle occipital gyrus.
No other significant differences were identified (see Supplemental Table 3).
Source construction: whole brain analysis
General sex differences irrespective of group affiliation
Irrespective of group affiliation (MCI versus healthy control), women generally demonstrated both a decrease in theta frequency band activity and an increase in gamma frequency band activity (t = 4.47, p = 0.01) under the 40 Hz RL. This decrease in theta activity occurred in the frontal lobe, localized mostly to the right medial frontal gyrus, posterior cingulate cortex, and extending slightly to the paracentral lobule (Figure 6(a)). The increase in gamma frequency band activity spread out more to the left hemisphere, within the lingual gyrus of the occipital lobe and the parahippocampal gyrus of the limbic lobe (Figure 6(b)). Similarly, compared to men during the control RL, women demonstrated a decrease in theta band activity (t = −3.12, p = 0.001) that was widespread to the left hemisphere, extending to left middle frontal gyrus, pre-and -post central gyrus, inferior parietal lobule, and the cingulate gyrus (Figure 6(c)).
Sex differences in response to 40 Hz RL
During the 40 Hz RL, women in the MCI group demonstrated decreased activity (compared to men in the MCI group) in the right middle frontal gyrus, extending to the paracentral lobule and posterior cingulate cortex (t = −6.27, p = 0.02) for theta frequency band (Figure 6(d)). Among the healthy control group, there was no significant difference demonstrated between women and men (t = 5.69, p = 0.37) during the 40 Hz RL condition.
Sex differences in response to control RL
During the control RL, women in the MCI group demonstrated decreased activity (compared to men in the MCI group) centered to the left precentral gyrus of the frontal lobe (t = −4.82, p = 0.02) for the theta frequency band (Figure 6(e)). Additionally, in the healthy control group, women demonstrated an increase in gamma band activity (t = 2.80, p = 0.03) in the middle occipital gyrus compared to men (Figure 6(f)).
Working memory
Differences between the two light conditions
Overall, participants demonstrated improvements in working memory accuracy and a reduction in reaction time between baseline and post-exposure. Specifically, participants showed a significantly greater increase in accuracy in response to the control RL compared to the 40 Hz RL (p = 0.011) (Figure 7(a)). The difference in reaction time between the light conditions was not significant (Figure 7(b)).

Differences (baseline versus post-exposure) in working memory accuracy and reaction time. (a) Comparison of working memory accuracy irrespective of group affiliation between the 40 Hz RL and control RL. (b) Comparison of working memory reaction time irrespective of group affiliation between the 40 Hz RL and control RL. (c) Comparison of working memory accuracy between sexes and light conditions irrespective of participant group affiliation. (d) Comparison of working memory reaction time between sexes and light conditions irrespective of participant group affiliation. The results were based on 34 subjects (20 MCI and 14 healthy controls) with complete data on behavior measures. The error bars represent standard deviation. Statistically significant (p < 0.05) differences are indicated by an asterisk.
Differences in 40 Hz gamma power across electrodes by light conditions and sex
There was a significant interaction between light condition and sex for working memory accuracy (p = 0.045). Men showed a greater increase in accuracy under the 40 Hz RL compared to the control RL (Figure 7(c)). In contrast, women exhibited a larger increase in accuracy with the control RL compared to the 40 Hz RL (see Figure 7(c)). While these results highlight a sex difference in response to light conditions, this effect was not consistent across groups (non-significant three-way interaction, p = 0.65, Supplemental Figure 2). No significant sex differences were observed for reaction time overall (Figure 7(d)) or between groups (Supplemental Figure 2).
Subjective sleepiness
The participants overall showed greater sleepiness (i.e., greater KSS scores) under the control RL compared to the 40 Hz RL, but the difference was not statistically significant (Supplemental Figure 3A). There were no significant differences for this measure based on sex (Supplemental Figure 3B–D).
Discussion
Our findings demonstrated differences in neural responses to 40 Hz RL, with sex variations in PSD changes, 1/f slope characteristics, and working memory performance. These results align with previous studies that highlight differential neural mechanism underlying gamma oscillations and cognitive function across sexes. 48 Specifically, findings of our study showed that men consistently demonstrated greater increases in 40 Hz power under the 40 Hz RL compared to women, which has profound implications for understanding neuroplasticity and brain activity differences between the sexes, especially in the context of MCI.
Power spectral density analysis revealed a significant increase in 40 Hz power at the C4 electrode under the 40 Hz RL compared to the control RL condition, particularly in men. The pronounced response at electrode C4, positioned over the right motor and somatosensory cortices, brings attention to the role of this region in processing rhythmic sensory input and integrating gamma-band activity related to both exogenous (stimulus-driven) and endogenous (internally generated) oscillatory mechanisms. Sensory gamma, which is elicited by rhythmic external stimulation, has been shown to facilitate perceptual integration and attention modulation. 49 Meanwhile, intrinsic gamma oscillation in the sensorimotor cortex is closely associated with motor planning, execution, and cognitive control processes.50,51 The significant response at C4, an electrode overlying the right sensorimotor cortex, is consistent with prior studies indicating that gamma oscillations in this region are particularly sensitive to rhythmic sensory input and motor-related processes,50,52,53 and further implicate that the enhanced 40 Hz power at C4 under 40 Hz RL may reflect established mechanisms of gamma modulation, more pronounced in men, in the sensorimotor system, rather than solely being an artifact of stimulation. Furthermore, the significant differences in 40 Hz power across other multiple electrode sites, including Fz, T8, and P4, further suggest that 40 Hz RL modulates brain oscillations in a spatially distinct manner, which may relate to underlying neurophysiological sex differences. For example, women displayed distinct patterns of PSD changes at electrode T8, where gamma power was slightly higher under the control RL or similar across both light conditions. Electrode T8 is located over the right temporal cortex, which is linked to verbal memory encoding and retrieval and in women often shows resilience through alternative neural pathways or compensatory mechanisms such as verbal memory strategies. 3
Estrogen may play a role here, as it supports neural plasticity and is known to modulate the hippocampus and related memory areas. 35 Thus, a plausible argument is that as estrogen levels decline in women with MCI, their reliance on gamma synchronization for cognitive compensation under 40 Hz RL stimulation may decrease. Instead, women may depend more on alternative, pre-existing compensatory mechanisms that involve the temporal cortex—an area linked to verbal memory and processing.
Findings of the 1/f component of the PSD provided further insight into the nature of gamma-band changes under each of the RL conditions. In healthy controls, the 1/f slope was only slightly steeper under the 40 Hz RL compared to the control RL condition, suggesting minimal broadband suppression and selective neural response to rhythmic stimulation. In contrast, MCI participants exhibited substantially steeper 1/f slope under the 40 Hz RL, indicating greater suppression of broadband power. As previously noted, women tend to exhibit greater neuroplasticity in response to cognitive challenges, potentially due to changes in estrogen-mediated neuromodulation. Crucially, the increased broadband suppression under 40 Hz RL in MCI participants does not appear to be driven by fatigue. If fatigue were a significant factor, we would expect a progressive steepening of the 1/f slope across time in all conditions. Instead, the control RL condition resulted in a shallower 1/f slope, indicating less broadband modulation. This reinforces the conclusion that 40 Hz RL selectively engages neural circuits in a way that is distinct from general fatigue-related spectral shifts.
Research indicates that estrogen influences synaptic plasticity differently in men and women, with estrogen receptor signaling contributing to sex specific neural adaptations. 54 The role of estrogen receptor beta (ERβ) in modulating synaptic strength and dendritic architecture further supports the idea that variations in estrogen levels influence cortical excitability and plasticity. Given that broadband suppression is thought to reflect shifts in the balance of excitatory and inhibitory (E/I) activity, 55 sex differences in E/I balance may underlie individual variability in response to 40 Hz RL. Given that broadband suppression is thought to reflect E/I balance, and women exhibit greater compensatory network engagement under cognitive load, it is plausible that 40 Hz RL elicits distinct broadband suppression patterns across sexes. This provides a possible mechanistic explanation for the lateralized gamma increases observed in women and their potential compensatory reliance on different neural circuits compared to men.
This pattern of compensatory reliance is further supported by whole brain sLORETA analysis, which revealed that women exhibited a general decrease in theta power and an increase in gamma power, particularly in the middle frontal and parahippocampal gyri, regions involved in cognitive processing and memory consolidation. The increase in gamma activity observed in women, especially in areas like the occipital and limbic lobes, suggests an adaptive response that supports visual processing and memory encoding, helping to compensate for a reduced reliance on theta activity in the frontal regions. Interestingly, the lateralization of activity also differs between men and women, with men showing stronger gamma synchrony in the right hemisphere (electrode C4) and women demonstrating more distributed or left-hemisphere gamma increases in response to the 40 Hz RL.
This lateralization effect is consistent with established differences in neural organization between sexes, with women more frequently exhibiting bilaterally distributed or left-lateralized cognitive function, while men tend to rely more on right –hemisphere-dominated spatial and memory tasks. 56 Furthermore, the observed increase in right-hemisphere gamma in men under the 40 Hz RL suggest that this stimulation pattern may preferentially enhance cognitive functions by reinforcing right-hemisphere network engagement. 57 This finding opens potential avenues for optimizing RL interventions to target hemisphere-specific networks based on sex, possibly improving cognitive resilience in men through tailored right-lateralized light therapies.
In working memory tasks, men showed greater accuracy gains under the 40 Hz RL while women showed greater improvement under the control RL. The higher gamma power observed in men under the 40 Hz RL suggests that this frequency preferentially enhances their cognitive function by promoting gamma synchronization within these networks.58,59
These results align with biological predispositions and highlight a novel avenue for potential future designs of adaptive light stimulation and treatment personalization. The uniqueness of the control RL stimulation in women compared to 40 Hz RL lies in their ability to engage broader and distributed neural networks, leveraging compensatory mechanisms aligned with women brain organization and hormonal influences. For example, at 30 Hz, the transition between high beta and low gamma activates pathways involved in cognitive functions like attention and emotional regulation, enhancing temporal-lobe processing linked to verbal memory. Meanwhile, 50 Hz stimulation boosts high-gamma activity, fostering synaptic plasticity in occipital and visual areas, which are more robust in women compared to men. In contrast, 40 Hz stimulation focuses narrowly on gamma synchronization, benefiting men more due to their reliance on right-hemisphere networks for spatial and memory tasks, whereas women depend more on bilateral activation and verbal strategies. Taken together, these findings open avenues for sex-specific light interventions in MCI, with a particular focus on electrode site and hemispheric lateralization. Sex-specific differences in neural organization, compensatory strategies, and hormonal influences strongly suggest that tailored light stimulation frequencies—control light with varying low and high frequencies for women, and targeted frequencies for men—could optimize cognitive outcomes in MCI patients.
Implications for personalized medicine
Overall, this study highlights the importance of personalized therapeutic strategies in treating MCI, tailored to sex-specific neurological profiles and hemispheric vulnerabilities. A key premise is that 40 Hz RL may enhance cognitive function in men by boosting right-hemisphere synchrony and regulating excessive theta activity at critical sites like C4 and Fz. In women, however, alternative approaches may be more effective. Given women's reliance on left-hemisphere or bilateral networks for memory and cognitive processing, interventions stimulating both hemispheres and selectively targeting left-lateralized regions may yield better outcomes. Further research could explore amplifying cognitive benefits in women with MCI by combining 40 Hz RL with hormone modulation therapies, or through careful control of the light's spatial and temporal characteristics—for example, by presenting light flickering at different frequencies (i.e., low beta, high gamma), or by presenting a flickering light to one eye only via monocular or hemifield stimulation, ensuring that the participant's gaze is fixed centrally and only stimulating the designated visual field based on the participant's sex.
Conclusion
This study provides strong evidence for sex differences in response to 40 Hz RL, with men showing greater improvements in gamma power, particularly in the MCI group. These findings open possibilities for developing tailored brain stimulation protocols and cognitive training regimens that address the distinct cognitive needs of men and women as they age. Future research should focus on understanding the underlying neurobiological mechanisms driving these differences, as well as exploring therapeutic approaches tailored to each sex, particularly in neurodegenerative conditions such as MCI.
Limitations
Several limitations must be considered. First, the generalizability of the results is limited by the study's sample size and demographic homogeneity. Future research should include larger, more diverse populations to validate the observed sex differences. Second, while the role of estrogen and hormonal fluctuations is acknowledged, further investigation is needed to understand how hormonal changes throughout the women lifespan impact brain oscillatory patterns and treatment responsiveness. Despite the study's promising results, a direct cause and effect relationship between brain oscillations and cognitive decline cannot be forged. Longitudinal research is necessary to track these oscillatory changes over time and their value for predicting cognitive outcomes. Additionally, the study included non-balanced sex groups, which may have influenced the observed differences. No distinction was made between amnestic and multi-domain MCI subtypes, which could impact the generalizability of findings within the MCI group. In addition, a key limitation of the current study is the absence of an occluded 40 Hz stimulation condition, which prevents definitive exclusion of frequency-specific stimulus-related electrical artifacts in the EEG signal. Although we applied shielding and included a matched luminance control, 40 Hz stimulation could still introduce artifacts. While our findings are consistent with neural gamma entrainment, future studies should include occluded or shielded stimulation conditions to validate the neural origin of observed 40 Hz EEG responses.
Lastly, individual chronotype differences were not accounted for, despite their potential influence on neural responses to RL stimulation. Future studies should incorporate these factors to refine the interpretation of treatment effects and improve the robustness of findings.
Supplemental Material
sj-docx-1-alz-10.1177_13872877251361061 - Supplemental material for Preliminary study of sex-based neural responses and cognitive benefits of a 40 Hz rhythmic light intervention in mild cognitive impairment
Supplemental material, sj-docx-1-alz-10.1177_13872877251361061 for Preliminary study of sex-based neural responses and cognitive benefits of a 40 Hz rhythmic light intervention in mild cognitive impairment by Ola A Alsalman, Weixin Li and Mariana G Figueiro in Journal of Alzheimer's Disease
Footnotes
Acknowledgments
The authors would like to express our sincere gratitude to all participants, including the mild cognitive impairment group members and control group members, for their invaluable contribution to this study. We also extend our thanks to the coordinators for their dedicated efforts in data collection and recruitment, which were essential to the success of this research.
Ethical considerations
The study was approved by the Institutional Review Board of the Icahn School of Medicine at Mount Sinai, New York (STUDY-21-00331).
Consent to participate
All participants in this study provided written informed consent prior to participation and any study procedures.
Author contribution(s)
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the National Institute on Aging (award 5R01AG072762).
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data availability statement
Anonymized data supporting the findings of this study will be made available from the corresponding author upon reasonable request.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.