
Abstract
Background
Dichlorodiphenyltrichloroethane (DDT) and its metabolites (DDTs), such as dichlorodiphenyldichloroethylene (DDE) and dichlorodiphenyldichloroethane (DDD), are synthetic organochlorine pesticides with long environmental persistence. Although DDT has been phased out in many countries, DDE and DDD remain prevalent worldwide. Growing evidence links DDTs exposure to Alzheimer's disease (AD), though underlying molecular targets and mechanisms remain unclear.
Objective
In this study, we investigated molecular targets and pathways through which DDTs potentially induce AD using network toxicology combined with molecular docking techniques.
Methods
AD-related targets associated with DDTs were identified through bioinformatics searches. Key targets were selected via STRING protein-protein interaction analysis and Cytoscape, followed by signaling pathway enrichment analysis. Diagnostic efficacy was evaluated using ROC curve analysis and nomogram modeling based on GEO datasets. Molecular docking validated binding affinity between DDTs and core target proteins predicted by AlphaFold 3.
Results
We identified 1732 potential molecular targets linking DDTs exposure to AD. Pathway analysis revealed DDTs predominantly affect AD pathogenesis by modulating apoptosis, p53 signaling, TNF signaling, and IL-17 signaling pathways. STRING and Cytoscape analyses identified seven core targets. GEO dataset validation indicated RPL23, RPS6, and RPS8 as pivotal targets, with RPL23 having strongest predictive capacity. Molecular docking confirmed binding interactions between DDTs and RPL23, with binding energies of −7.2 kcal mol−1 for DDT, −6.5 kcal mol−1 for DDE, and −7.1 kcal mol−1 for DDD.
Conclusions
This research provides novel insights into neurotoxic mechanisms of DDT and its persistent metabolites DDE and DDD, and supports enhanced public health strategies for AD prevention.
Introduction
Dichlorodiphenyltrichloroethane, also known as DDT or 1,1,1-trichloro-2,2-bis(p-chlorophenyl) ethane, is a synthetic pesticide belonging to the family of organohalogen compounds. 1 Since the 1940s, it has been widely used for plant protection and infectious disease control, as well as in antifouling paints for ships. 2 In the natural environment, DDT is primarily transformed through biological and abiotic degradation into two key metabolites: dichlorodiphenyldichloroethane (DDD) via reductive dechlorination under anaerobic conditions, and dichlorodiphenyldichloroethylene (DDE) via dehydrochlorination under aerobic conditions. 3 However, due to the stable molecular structures of DDT and its derivatives (DDTs), particularly DDE and DDD, which are difficult to degrade further, they exhibit environmental persistence, further exacerbating the long-term nature of environmental pollution.4,5 Since the 1970s, many countries have banned the use of DDT, and in 2004, DDT was listed as a persistent organic pollutant in the Stockholm Convention on Persistent Organic Pollutants. 5 However, in some developing countries such as those in Africa, DDT remains selectively employed to control vector-borne disease transmission in efforts to reduce the malaria burden. 6 Despite legal restrictions on its use, DDT residues can still enter aquatic ecosystems through runoff, sewage discharge, or atmospheric deposition, accumulate in benthic organisms, and potentially biomagnify in aquatic food webs.2,7 In addition, due to their non-polar molecular structures, DDTs are highly lipophilic, making them prone to accumulation in organisms and producing biological toxicity, 8 thus posing potential ecological and health threats to wildlife and humans at higher trophic levels through trophic transfer. 9
Previous studies have shown that DDTs have strong toxic effects, which may be related to diseases such as pancreatic cancer, reproductive disorders, and Alzheimer's disease (AD).9,10 AD is an age-related neurodegenerative disease characterized by the gradual deterioration of cognition, function, and behavior. 11 Research shows that both genetic and environmental factors may influence the occurrence and development of AD, such as environmental toxins like DDTs which may increase the risk of AD. 12 For example, epidemiological studies indicate that the serum DDE levels in AD patients are 3.8 times higher than in the normal population. 10 Additionally, in SH-SY5Y cells, primary neurons, and wild-type C57BL/6J and 3xTG-AD mouse models, DDT exposure significantly increases amyloid-β protein precursor (AβPP) mRNA and protein levels, and by interfering with sodium ion channels, leads to neuronal hyperexcitation, thereby increasing the release of amyloid-β protein (Aβ) and the production of Aβ, thus promoting the progression of AD. 13 However, regarding the specific targets and toxicological mechanisms by which DDTs induce the development of AD, relevant research is still limited and requires more evidence to support.
Network toxicology is an innovative computational network-based approach that integrates bioinformatics, genomics, proteomics, and metabolomics to construct association networks among compounds, targets, and pathways. 14 This method is particularly adept at addressing complex interactions involving multiple components, diseases, and targets, and it translates intricate mechanisms into comprehensible visual representations. Consequently, network toxicology offers a robust framework for analyzing protein-protein interactions and predicting the molecular mechanisms through which toxins facilitate disease progression. AlphaFold3 is capable of predicting the interaction structures of complex molecular assemblies with high accuracy, without the need for prior structural information, and it can detail the binding interactions and conformational changes between toxins and biomolecules such as metabolic enzymes, receptors, and antibodies. 15 Molecular docking, on the other hand, simulates the spatial orientation and binding interactions of small molecule compounds with proteins at the atomic level, thereby providing valuable insights into potential therapeutic effects or adverse reactions. In the context of toxicology research, molecular docking can predict and clarify the interactions between toxins and biomolecules, thus uncovering toxicity mechanisms and their potential harm to organisms.16,17 This approach significantly enhances the efficiency of analyzing toxicity pathways mechanistically and establishes a groundbreaking molecular foundation for investigating the toxicity mechanisms of environmental toxins and pharmaceuticals, thereby expediting the toxicological risk assessment process.
Accordingly, this study utilizes network toxicology methodologies to delve into the intricate relationship between DDTs and potential targets associated with AD, with the objective of elucidating the mechanisms by which DDTs impact the onset and progression of AD. Additionally, it leverages external public datasets to further validate these potential targets, employs AlphaFold3 to predict the protein structures of these targets, and ultimately uses molecular docking techniques to explore the molecular interactions between DDTs and key potential target proteins. This integrated analysis seeks to clarify the molecular underpinnings of how DDTs influence the development and progression of AD and to pinpoint potential intervention sites that could alleviate their detrimental effects.
Methods
Network toxicological analysis of DDTs
The SMILES sequences of DDT, DDE, and DDD were retrieved from the PubChem database (https://pubchem.ncbi.nlm.nih.gov/). These sequences were then inputted into the ProTox 3.0 database (https://tox.charite.de) to obtain toxicity predictions. 18
Collection of DDTs targets
The PubChem database was initially searched to confirm the names and molecular formulas of DDTs (Figure 1A). The corresponding SDF file was uploaded to the Swiss Target Prediction website (http://www.swisstargetprediction.ch/), 19 and targets with a probability greater than 0 were selected. Additional targets were retrieved from the ChEMBL database (https://www.ebi.ac.uk/chembl/), 20 restricted to Homo sapiens, and from the Comparative Toxicogenomics Database (CTD, http://ctdbase.org/). 21 Targets from these sources were integrated, deduplicated, and standardized using the UniProt database 22 to compile the final DDTs target library.
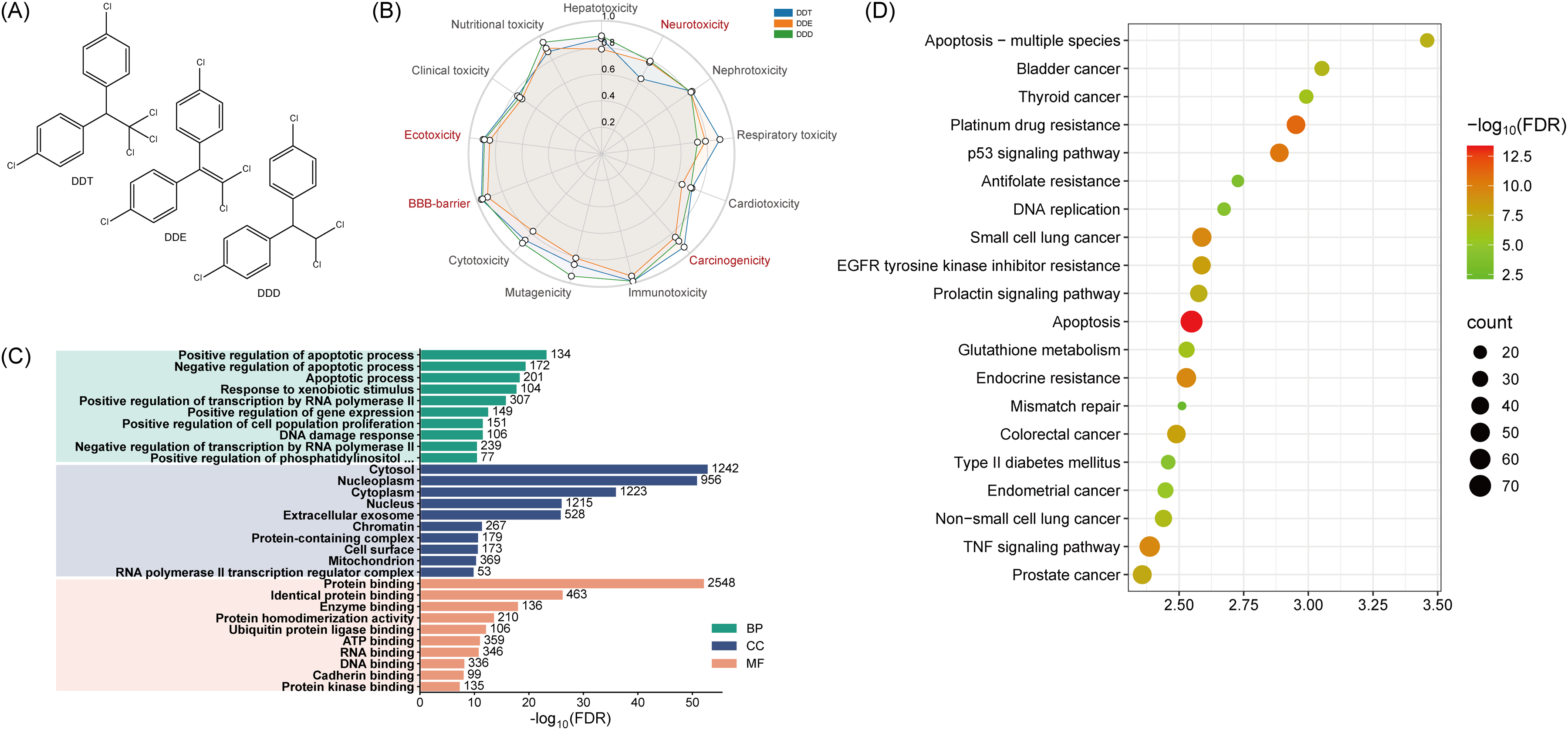
Molecular structure of DDTs (A), toxicity distribution map (B) and enrichment results of predicted targets (C, D).
GO and KEGG pathway analysis
Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) analyses were conducted using the DAVID database (https://david.ncifcrf.gov/) to identify pathways associated with the targets. 23 The analysis focused on biological processes, molecular functions, and cellular components to elucidate key signaling pathways. GO and KEGG enrichment results were visualized using the Weishengxin platform (http://www.bioinformatics.com.cn).
Target construction of diseases
Disease target libraries were constructed using the GeneCards database (https://www.genecards.org/) with keywords such as “Alzheimer's disease”. 24 Genes with scores above the median were selected from the resulting targets to ensure high relevance to AD.
Construction of protein–protein interaction (PPI) networks
The intersection of DDTs targets and AD targets was determined using the Venn website (http://bioinformatics.psb.ugent.be/webtools/Venn/). Overlapping targets were used to construct a PPI network via the STRING database (https://cn.string-db.org/), 25 with Homo sapiens selected as the species and an interaction score of ≥0.7. The network was visualized and analyzed in Cytoscape, where topological properties were calculated. The CytoHubba plugin identified the top 20 significant genes using the maximal clique centrality (MCC), edge percolated component (EPC), maximum neighborhood component (MNC), and degree algorithms. The MCODE plugin detected significant subclusters. Core genes were determined by intersecting the targets from the MCODE module with those predicted by the four CytoHubba algorithms.
Core gene diagnostic modeling in AD
The diagnostic potential of the core gene for AD was evaluated using the GSE138260 dataset. This dataset comprised postmortem brain tissue samples from patients with typical AD, diagnosed according to the IWG-2 criteria, and from neurologically and psychiatrically healthy controls. After quality control, 17 AD cases and 19 controls were included in the analysis. The expression levels of the core gene were compared between AD and normal groups, and receiver operating characteristic (ROC) curves were constructed to assess diagnostic accuracy. A nomogram model was developed to further evaluate the clinical diagnostic value of core gene alterations. The nomogram was visualized in R 4.5.0 utilizing packages such as tidyverse 2.0.0, rms 8.0–0, and pROC 1.8.5. Group comparisons were performed using the Wilcoxon test, with statistical significance set at p < 0.05.
Molecular docking
Molecular docking was performed to analyze the interactions between DDTs and core target proteins. Ligand structures were retrieved from PubChem, and core gene sequences were obtained from UniProt. The three-dimensional structures of the proteins were predicted using AlphaFold 3 (https://alphafoldserver.com/). 15 The ligand small molecules were docked with the predicted protein structures using CB-Dock 2 (http://cao.labshare.cn/cb-dock/), 26 and the results were visualized. Additionally, LigPlus was employed to obtain and visualize the 2D ligand-protein binding interactions.
Results
Toxicity analysis of DDTs
The organ-specific toxic effects of DDTs (hepatotoxicity, neurotoxicity, nephrotoxicity, respiratory toxicity, and cardiotoxicity) and its toxic endpoints (carcinogenicity, immunotoxicity, mutagenicity, cytotoxicity, blood-brain barrier (BBB), ecotoxicity, clinical toxicity and nutritional toxicity) were assessed using the PROTox platform (Figure 1B). The results indicated that the three DDTs consistently exhibited active effects on neurotoxicity, cardiotoxicity, ecotoxicity, and BBB permeability. This demonstrates a pronounced neurotoxic potential and an ability to disrupt the BBB, strongly suggesting an elevated risk of adverse neurological effects.
To investigate the potential biological impacts of DDTs, we thoroughly screened targets sourced from the Swiss Target Prediction website, ChEMBL and CTD databases, generating a substantial dataset. These targets were merged, deduplicated, and normalized using the UniProt database, revealing 3575 potential targets specifically associated with DDTs induced disease. These targets underwent functional annotation through DAVID, focusing on Homo sapiens, for GO and KEGG pathway enrichment. This process generated 159 KEGG entries and 1772 GO entries, comprising 1197 biological processes (BP), 239 cellular components (CC), and 336 molecular functions (MF).
As depicted in Figure 1C, the BP category was primarily in apoptotic modulation (both activation and suppression) and xenobiotic response. The CC category was mainly linked to cellular compartments such as the cytosol, nucleoplasm and cytoplasm. The MF annotations were centered on protein binding, identical protein binding, enzyme binding, and protein homodimerization activity. Figure 1D showed various disease signaling pathways, including the apoptosis p53 and TNF signaling pathways.
Despite methodological differences in annotation, the results consistently implicated several neurotoxicity associated biological processes and pathways, particularly apoptosis and inflammatory response, all of which are known to regulate cell survival and inflammation. Additionally, previous research supports an established association between DDTs exposure and AD, 12 with BBB dysfunction noted in various central nervous system (CNS) diseases, including AD. 27 In summary, DDTs exhibited notable neurotoxicity, and in conjunction with its potential AD risk, we conducted a network toxicology analysis to further understand whether DDTs increase the risk of AD.
Identification and enrichment analysis of core targets for DDTs action in AD
To identify the core pathogenic target genes associated with DDT, DDD, and DDE, we retrieved 3585 genes related to these compounds from the GeneCards database. These genes were then analyzed to create a Venn diagram (Figure 2A), which illustrates the overlapping and unique genes among DDT, DDE, and DDD. Subsequently, to determine the core targets relevant to both these compounds and AD, we intersected the genes associated with DDT, DDD, and DDE with those associated with AD, identifying 1732 common genes, which we defined as the core targets (Figure 2B). We then performed enrichment analysis on these core targets to explore their potential biological functions and mechanisms. For the GO enrichment results, we selected the top 10 entries in each category and presented them in a bubble chart (Figure 2C). For the KEGG enrichment analysis, we selected the top 20 enriched pathways and illustrated them in a Figure 2D.

Associations between Alzheimer's disease (AD) and DDTs. (A) Venn diagram of predicted targets for the three DDTs. (B) Venn diagram of hub targets shared between AD and DDTs. (C) GO enrichment analysis of hub targets. (D) KEGG enrichment analysis of hub targets.
As shown in Figure 2C, the BP category was primarily associated with responses to xenobiotic stimuli, regulation of apoptosis (including both promotion and inhibition), and positive regulation of gene expression. The CC category mainly included cytosol, extracellular exosomes, cytoplasm, and nucleoplasm. The MF category was predominantly related to protein binding, identical protein binding, and enzyme binding. In the KEGG enrichment analysis, pathways such as Apoptosis, p53 signaling, TNF signaling, and IL-17 signaling were significantly enriched (Figure 2D). These findings suggest that apoptosis and immune responses may play crucial roles in the pathogenesis of AD associated with DDT, DDD, and DDE.
Protein interaction network construction of core targets and identification of hub genes
To identify the hub genes involved in the effects of DDTs on AD, we utilized the STRING database to construct a PPI network for 1732 core targets and imported the results into Cytoscape software for further analysis. Next, we used the “MCC, EPC, MNC and Degree” algorithms from the “cytoHubba” plugin to calculate the key genes, and ranked the top 20 genes of each algorithm as the key genes of that algorithm (Figure 3A). Finally, the MCODE plugin in Cytoscape was used to cluster these core target genes. This integrated approach identified seven hub genes: RPS3, RPS6, RPS8, MED1, RACK1, RPL23, and RPS27A (Figure 3B, C). These seven hub genes were defined as central to DDTs’ role in AD and were used for subsequent analyses.

Construction of protein–protein interaction (PPI) networks and identification of core targets. (A) Top 20 genes identified by four algorithms in Cytoscape. (B) Intersection plot of genes from the four algorithms. (C) Core PPI network obtained by MCODE, with genes common to all algorithms highlighted in red.
Construction of a diagnostic model for hub genes in AD
In the present study, we statistically analyzed the expression of the seven hub genes based on the AD-related dataset GSE138260. Using the gene expression profile data from the dataset, we analyzed the potential targets screened. As depicted in Figure 4, RPL23, RPS6 and RPS8 were significantly down-regulated in the AD group (p < 0.05), while the other four hub genes did not show significant differential expression. Therefore, we first evaluated the diagnostic value of these three significant genes using ROC curves. As shown in Figure 4D, the RPL23 model demonstrated the highest discriminative ability (AUC = 0.817), followed by RPS6 (AUC = 0.771) and RPS8 (AUC = 0.718). Subsequently, we constructed a Nomogram model based on RPL23, RPS6, and RPS8. As shown in Figure 4E, this risk prediction framework converted the expression values of RPL23, RPS6, and RPS8 into quantifiable disease probabilities and emphasizing the cumulative effect of these genes, and RPL23 shows the higher point scale. Overall, the highest AUC for RPL23 corresponds to its higher point scale in the Nomogram model, suggesting that RPL23 may play a more significant role in disease prediction.
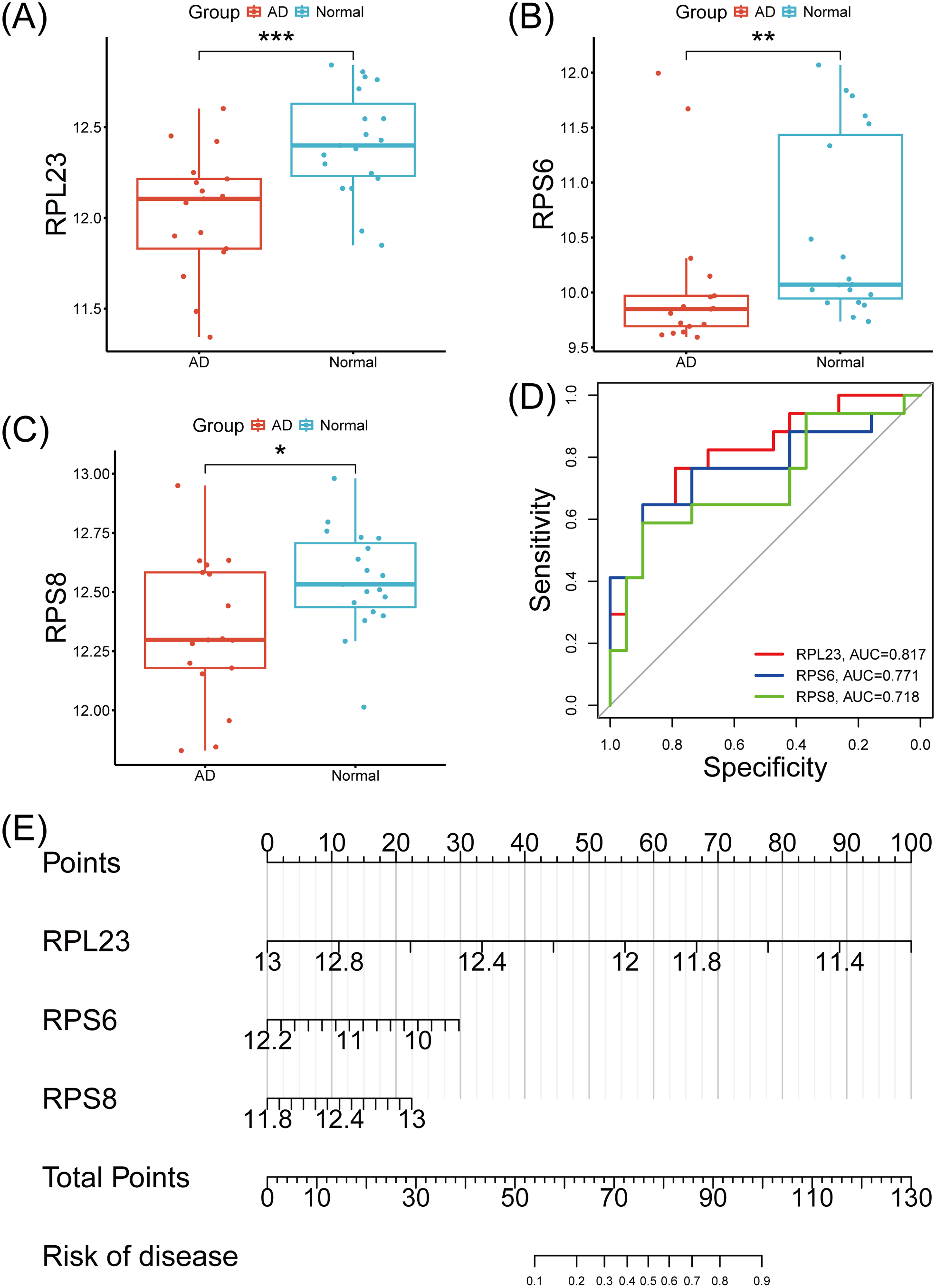
Diagnostic model based on external GEO datasets. (A–C) Expression levels of RPL23, RPS6 and RPS8 in Alzheimer's disease (AD) versus normal groups. (D) ROC curves for each core gene in AD. (E) Nomogram model for AD diagnosis based on core genes.
Analysis of molecular docking results
We had previously demonstrated that RPL23 had good diagnostic value for AD. To further validate the interactions of 3 DDTs with RPL23, molecular docking was performed. As shown in Figure 5, DDT exhibited the strongest binding affinity to RPL23 (−7.2 kcal/mol), forming key interactions with residues Asp127, Ala12, Glu124, Lys91, Arg90 and Phe14 in the 2D structure. Similarly, DDE bound to RPL23 (−6.5 kcal/mol) via residues Lys91, Ala12, Leu128, Asp127, Glu124, Phe14, and Lys123 in the 2D structure. DDD also formed stable complexes with RPL23 (−7.1 kcal/mol) through interactions with residues Arg90, Asp92, Lys123, Glu124, Asp127, Ala12, Phe14, and Lys91 in the 2D structure. These computational predictions suggest a stable and secure interaction of DDTs with RPL23, underscoring RPL23's capacity to effectively bind these compounds, which may elucidate their biological effects.

Molecular docking results of the lowest binding energy in 3 DDTs with the RPL23.
Discussion
Similar to most other organochlorine insecticides, DDT exhibits relatively low acute toxicity, an advantage that was once a favorable factor for its widespread use. 28 However, due to the hydrolytic stability of its carbon-chlorine bonds, the compound exhibits high resistance to biodegradation and photolysis, with a half-life in soil ranging from 2 to 15 years and up to approximately 150 years in aquatic environments. Its large-scale, uncontrolled, and intensive use has led to global pollution. 29 Although most countries have banned the use of DDT, some developing countries still use it for vector control for public health reasons. 6 However, a more serious problem lies in the fact that environmental pollutants (such as microplastics) exhibit extremely strong enrichment capabilities for residual DDT. 30 This originates from the properties of microplastics, including small particle size, large surface area, and high hydrophobicity. 30 Furthermore, microplastics act as vectors, enhancing the transport and accumulation of small molecular contaminants in the environment and thereby exacerbating the severity of pollution. 30 Previous study collected polyethylene pellet samples from 30 beaches across 17 countries, with results showing DDT and DDE detection in most samples. 31 Importantly, given that microplastics readily penetrate various tissues throughout the body, particularly accumulating in highly perfused organs such as the liver, kidneys, and small intestine due to their preferential circulatory transport, the co-exposure scenario becomes increasingly concerning. 32 Notably, the brain also shows considerable microplastics deposition, exceeding levels in the heart and lungs, with a predominance of sub-10-μm particles likely penetrating the BBB via circulation and oxidative stress-induced endothelial permeability. 32 This neurotropic accumulation pattern, combined with the strong affinity of microplastics for DDTs, suggests potential synergistic neurotoxic effects that may contribute to the pathogenesis of neurodegenerative diseases such as AD. In addition, DDTs have been shown to accumulate in all biological tissues, particularly in adipose tissue, where even long-term exposure to low concentrations can lead to significant accumulation. 28 Previous research has revealed DDT residues in all birds and fish, including species inhabiting desert areas or deep-sea environments. 28
To explore the causal relationship between DDTs and AD, we chose the DDT and its two key metabolites, DDE and DDD for investigation. Predictions from toxicity prediction websites revealed that DDTs exhibited significant neurotoxicity. Additionally, various studies have shown an association between DDT and its metabolites exposure and AD, with this effect being particularly pronounced in individuals with specific genetic predispositions. 10 For example, Richardson et al. 10 demonstrated that serum DDE concentrations were markedly elevated in AD patients relative to healthy controls, yielding an odds ratio of 4.18 for the highest exposure tertile, an association that was especially robust among carriers of the APOE ε4 allele. In addition, previous studies have indicated a significant dose-response relationship between DDT exposure levels and the progression of AD. Kalia et al. 33 found that DDT at a 3 μM concentration can significantly increase tau protein aggregation and impair mitochondrial respiratory function. Similarly, in in vitro cell experiments, treatment with 10 μM DDT not only significantly increased Aβ40 and Aβ42 levels but also impaired the functions of ATP-binding cassette transporter A1 and insulin-degrading enzyme. 34 More importantly, after 3xTG-AD mice underwent long-term low-dose exposure (3 mg/kg, administered every 3 days for 90 days), a significant increase in Aβ deposition and neuropathological changes was also observed in their brains, 13 demonstrating chronic environmental exposome factors can contribute to neurodegenerative pathology. Furthermore, preclinical studies reveal that exposure to DDT during the critical prenatal and neonatal phases of neurodevelopment precipitates behavioral and neurochemical alterations observable in adulthood. 35 Despite the prohibition of DDT use in most nations since the 1970s, the long half-life of DDT and its metabolite DDE lead to their bioaccumulation and persistent exposure, 36 which may increase the risk of AD. This cumulative toxicity suggests that even chronic exposure equivalent to environmentally relevant concentrations (within the 0.1–1 μM range) may progressively promote the development of AD-related pathological processes by interfering with voltage-gated sodium channels and associated cellular signaling pathways. 13 Data from the National Health and Nutrition Examination Survey (NHANES), conducted by the Centers for Disease Control and Prevention, show that DDE was still present in human serum samples as of 2016. 37 Therefore, our research on the impact of DDTs on AD is well-supported by existing theoretical foundation and background. These findings underscore the cumulative influence of lifelong exposure to DDT, DDE, and DDD on the risk of disease.
Our study provided substantial evidence that DDTs were a risk factor for the development of AD. Based on network toxicology studies, DDTs may promote the development of AD by regulating multiple key proteins and regulatory factors in AD, primarily through the apoptosis and p53, TNF and IL-17 signaling pathways. Apoptosis is a tightly regulated process that plays a crucial role in maintaining tissue homeostasis and eliminating damaged or unnecessary cells. In the context of neurodegenerative diseases, the dysregulation of apoptosis contributes to the primary factor to the reduction of neurons in the brains of individuals with AD.38,39 p53, TNF-α, and IL-17 drive pathological progression in AD through different mechanisms. p53 may induce tau phosphorylation and facilitate neurofibrillary tangle formation. 40 Tau is a key neuronal microtubule-associated protein (MAP) crucial for stabilizing microtubules, regulating axonal transport, and maintaining cytoskeletal integrity. 41 However, hyperphosphorylation of tau can reduce its ability to bind microtubules, leading to cytoskeletal disruption, and promotes tau misfolding into neurofibrillary tangles, sequesters normal tau and other MAPs, impairs axonal transport, and causes neuronal degeneration and inhibits tau degradation, resulting in toxic aggregate accumulation. 41 In AD, hyperphosphorylated tau inhibits p53 activation by reducing its phosphorylation at Ser33 via CDK-5 and GSK-3β, thereby suppressing p53-mediated apoptosis and promoting chronic neuronal degeneration. 42 Conversely, activated p53 enhances GSK-3β activity, forming a positive feedback loop that further drives tau hyperphosphorylation and Aβ accumulation, exacerbating synaptic dysfunction and cognitive decline. 41 In addition, inflammation plays a crucial role in the initiation and/or deterioration of AD neurodegeneration, and increasing evidence suggests that TNF-α and IL-17 contribute to neuroinflammation that aggravates both Aβ and tau pathology. TNF-α, a pro-inflammatory cytokine upregulated in AD, which can exacerbate neurodegeneration, BBB disruption, and overall disease progression. 43 IL-17, primarily produced by immune cells like γδ T cells, accumulates in the brain and meninges during early AD stages, triggering synaptic and cognitive deficits that precede significant Aβ or tau pathology. 44
To further validate whether the core proteins targeted by DDTs have a causal relationship with AD, we performed PPI network analysis and identified seven hub genes playing a central role in DDTs-induced AD, in which five of these targets belong to the ribosome family. Ribosomes are specialized complexes composed of nucleic acids and proteins that are responsible for mediating all protein synthesis. 45 Various studies have demonstrated that ribosomal dysfunction is critically involved in AD. Ding et al. 46 demonstrated significant ribosomal impairment within parietal lobule and the superior and middle temporal gyri of mild cognitive impairment and AD patients. This compromise is evidenced by diminished protein synthesis kinetics, reduced rRNA/tRNA abundance, and elevated RNA oxidation. Notably, such alterations were absent in the cerebellum. Furthermore, when ribosomes undergo dysfunction, exemplified by ribosomal stalling during the translation of the APP-C99 fragment, it precipitates a failure in ribosome-associated protein quality control. 47 Such ribosomal stalling is amplified by mutant or hyperphosphorylated tau, which binds more strongly to ribosomes and disrupts their function by decreasing the biogenesis of the 60S ribosomal subunit, primarily through the N-terminal projection domain of tau that interferes with ribosomal protein assembly and translation efficiency. 48 This failure results in the aberrant addition of alanine residues to the C-terminus of C99, yielding aberrantly modified APP-C99 proteins. These modified C99 fragments exhibit a propensity for aggregation, generating insoluble aggregates that perturb cellular proteostasis, impair lysosomal function and autophagic pathways, and thereby facilitate the accumulation of Aβ. 47 Moreover, the early accumulation of APP-CTFs in AD results in aberrant mitochondrial morphology and defective mitophagy, augmenting the generation of reactive oxygen species (ROS). Excessive ROS inflict damage upon mRNA, compromising its base-pairing integrity, which induce ribosomal stalling and exacerbates neuronal degeneration. 49 Contributing to this vicious cycle, hyperphosphorylated tau further promotes ROS accumulation, and the elevated ROS in turn directly hastens the excessive phosphorylation of tau protein. 50 This positive feedback loop may lead to persistent ribosomal dysfunction, mitochondrial defects, and escalating oxidative stress, thereby perpetuating and worsening the processes of mRNA damage and neuronal loss. 51 To further find core targets pivotal to DDT-induced AD, we developed a diagnostic model based on AD-related datasets for subsequent analysis. The results showed that RPL23, RPS6, and RPS8 play a key role in DDTs-induced AD, with RPL23 being particularly influential in disease prediction. Each of RPL23, RPS6, and RPS8 has specific functions in the ribosome's 60S and 40S subunits. Previous research has suggested that early anomalies in ribosomal proteins such as RPL23, RPS6, and RPS8 could serve as potential biomarkers for AD. Moreover, a diminished expression of these proteins is observed in the tissues derived from AD patients,52,53 which also find in our results. Therefore, we suggest a significant correlation between ribosomes, particularly RPL23, and DDTs in the progression of AD.
In order to find the optimal binding site of RPL23 and DDTs, we performed molecular docking to better explore the interaction between toxic molecules and protein macromolecules. The strong binding affinity was observed between DDT, DDE, DDD, and RPL23. Specifically, through comprehensive bioinformatics analysis, we suggest that increased exposure to DDTs may enhance their binding to RPL23, thereby affecting RPL23 expression. This alteration might trigger apoptosis and inflammatory signaling pathways, potentially exacerbating the progression of AD. Evans et al. 48 reported decreased synthesis of RPL23 in a tau transgenic mouse model, and this finding was also observed in the tissues of frontotemporal dementia patients in their study. Additionally, Willett et al. 54 study confirmed abnormal expression of the RPL23 gene in the brain tissues of AD patients, particularly in excitatory and inhibitory neurons, further suggesting that RPL23 may contribute to AD pathology through its role in ribosomal function. Therefore, RPL23 may represent a key pathway in the effects of DDTs on AD progression.
In the present study, we identified RPL23 as an important target associated with DDTs-induced AD. Consequently, it was plausible to hypothesize that AD patients exposed to DDTs might exhibit decreased RPL23 expression levels compared to healthy individuals. Furthermore, this finding could also enhance public health strategies. For instance, the identified core targets offer novel perspectives for advancing research into targeted therapeutic drugs. Moreover, our finding may support the creation of pharmaceuticals designed to prevent or treat AD and similar conditions in affected populations. By deepening our understanding of how various pollutants initiate disease, this research establishes a foundation for vaccine innovation, thereby strengthening public health measures and disease prevention approaches.
In summary, our study identifies potential targets of DDTs that may contribute to the development of AD. Using network toxicology and molecular docking techniques, we found that DDTs affect proteins RPL23, RPS6, and RPS8, which are associated with AD. This might lead to the dysregulation of apoptosis and the p53, TNF, and IL-17 signaling pathways. Furthermore, we explored the causal relationship between these core proteins and DDT-induced AD, identifying RPL23 as a risk factor. Molecular docking confirmed the binding stability between RPL23 and DDTs. Our research highlights the potential impact of the environmental pollutant DDTs on the onset and progression of AD, providing new insights for DDT usage and regulatory guidelines. However, the clinical metadata in the dataset do not include age at symptom onset or age at diagnosis, which prevents reliable classification of cases as early-onset or late-onset AD. As a result, the applicability of our findings to specific onset subtypes cannot be determined from the current data. To address this limitation, future research will prioritize recruiting individuals with well-documented onset ages, designing targeted experiments, and validating the results in independent cohorts to assess generalizability.
Footnotes
Acknowledgements
We would like to express our sincere gratitude to the network toxicology research platforms for their invaluable contributions to this study.
Ethical considerations
Not applicable
Consent to participate
Not applicable
Consent for publication
Not applicable
Author contribution(s)
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the project of Hubei Provincial Administration of Traditional Chinese Medicine (grant number: ZY2025M012) and the project of The Affiliated Hospital of Wuhan Sports University (grant number: WTFY202304).
Data availability statement
The data that support the findings of this study are available from the corresponding author, Shuna Jin, upon reasonable request.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.