
Abstract
Background
Intracerebral administration of amyloid-β (Aβ) peptides remains a widely used experimental approach for modeling key neurochemical features of Alzheimer's disease (AD), including neuroinflammation, oxidative stress, and synaptic dysfunction. Among these models, Aβ1−42 and Aβ1−40 are most frequently employed. However, their effects are often treated as interchangeable despite recognized differences in aggregation behavior and biological activity.
Objective
This systematic review aimed to synthesize evidence on how peptide selection between Aβ1−42 and Aβ1−40 shapes neurochemical outcomes in intracerebral rat models of AD.
Methods
The review was conducted in accordance with PRISMA guidelines and registered in PROSPERO. A structured search identified in vivo rat studies using Aβ1−42 or Aβ1−40. Given methodological heterogeneity, findings were synthesized using a qualitative framework. To address potential structural confounding, stratified analyses were performed according to injection route and aggregation state.
Results
Eighty-three studies met inclusion criteria (Aβ1−42 = 60; Aβ1−40 = 23). Across neurochemical domains, Aβ1−42 was more frequently associated with earlier and glial activation, increased pro-inflammatory mediators, oxidative imbalance, and synaptic alterations, often accompanied by behavioral deficits. In contrast, Aβ1−40 models showed a more variable and generally attenuated neurochemical response with greater dependence on experimental parameters. Structural analyses indicated clustering by injection route, whereas aggregation-state distribution was not statistically skewed between peptides.
Conclusions
The available evidence suggests that Aβ1−42 and Aβ1−40 should not be assumed interchangeable without explicit methodological consideration. Observed differences appear context-dependent and influenced by experimental design variables. Peptide selection with defined mechanistic objectives may enhance interpretability and translational relevance in preclinical AD research.
Keywords
Introduction
Alzheimer's disease (AD) is a progressive neurodegenerative disorder characterized by pathophysiology involving synaptic dysfunction, neuroinflammation, oxidative stress and ultimately neuronal loss.
Amyloid-β (Aβ) accumulation represents a central pathological feature of AD, while translation of amyloid-based mechanisms into effective disease-modifying therapies has remained limited. One important contributor to this translational gap is the persistent heterogeneity of experimental models used to study amyloid pathology, particularly in preclinical research, where substantial methodological variability exists between study designs. 1
Among available experimental approaches, intracerebral administration of Aβ peptides in rodents remains a widely used model for investigating specific pathogenic mechanisms and for evaluating candidate therapeutic strategies. Transgenic models, based on the expression of mutant human amyloid precursor protein and presenilin genes, have provided valuable insights into amyloid deposition and cognitive impairment. However, their complexity, cost, and limited predictive value for clinical trial outcomes have highlighted the continued relevance of intracerebral Aβ models for targeted mechanistic and pharmacological studies. 2
A fundamental limitation of intracerebral Aβ models is the implicit assumption that Aβ constitutes a single molecular entity. In practice, the two most used peptides, Aβ1−42 and Aβ1−40, differ substantially in their biophysical properties, aggregation kinetics, and biological effects. 3 Aβ1−42 exhibits a higher propensity for oligomer and fibril formation and is generally considered more neurotoxic, whereas Aβ1−40 is less aggregation-prone and has been more frequently associated with vascular amyloid deposition and chronic pathological processes. Despite these well-established biochemical differences, both peptides continue to be used across intracerebral models with limited consideration of how peptide selection may shape experimental outcomes. 4
Experimental findings indicate that Aβ1−42 and Aβ1−40 induce distinct neurochemical trajectories following intracerebral administration. Studies employing Aβ1−42 commonly report rapid activation of microglia and astrocytes, robust increases in pro-inflammatory cytokines, pronounced oxidative stress, and early disruption of synaptic integrity.5,6 In contrast, Aβ1−40 administration is more often associated with attenuated or delayed inflammatory responses, more subtle oxidative alterations, and relatively preserved neuronal and synaptic markers, with some studies emphasizing vascular-associated changes rather than overt neurotoxicity. These differences have important implications for the interpretation of behavioral outcomes and for the evaluation of candidate neuroprotective or anti-inflammatory interventions. 3
Despite the growing body of literature employing intracerebral Aβ injection models, the field remains fragmented. Studies vary widely in peptide preparation protocols, solvents, aggregation states, dosing regimens, injection sites, and timing of outcome assessment. Such methodological diversity not only limits reproducibility but also obscures peptide-specific effects, making it difficult to determine whether discrepant findings reflect true biological differences or methodological confounders. 7 Notably, few syntheses of the literature have systematically examined how Aβ peptide choice interacts with neurochemical endpoints across studies, and existing reviews often address amyloid models at a conceptual level without disentangling peptide-specific molecular profiles. 8
In this context, the present systematic review aims to synthesize current evidence on how Aβ peptide selection shapes neurochemical outcomes in intracerebral rodent models of AD. Rather than organizing the literature by experimental protocol, we structure this review around key neurochemical domains, including neuroinflammation, oxidative and mitochondrial dysfunction, synaptic integrity, and associated behavioral outcomes. By integrating findings across these domains, we seek to clarify consistent peptide-specific patterns, identify major sources of methodological variability, and discuss their implications for experimental design and translational interpretation in AD research.
Methods
Protocol registration
This systematic review was conducted in accordance with the PRISMA 2020 guidelines for systematic reviews of health research. The review protocol was prospectively registered in PROSPERO (registration number: CRD420251080137), where the review objectives, eligibility criteria and outcome domains were predefined.
Literature search strategy
A comprehensive literature systematic search was performed in PubMed, Scopus, and Web of Science databases, covering publications from database inception through June 30, 2025. Only peer-reviewed articles published in English were considered. The search strategies combined controlled vocabulary and free-text terms related to amyloid pathology and experimental modeling, including “Alzheimer's disease”, “amyloid beta”, “Aβ1−42”, “Aβ1-40”, and “intracerebral injection”. Database-specific filters were applied to restrict results to animal studies. Reference lists of eligible articles were manually screened to identify additional relevant studies.
Eligibility criteria
Studies were eligible for inclusion if they fulfilled the following criteria:
original experimental research using a wild-type rat model; intracerebral administration of Aβ peptides with explicit specification of the peptide species (Aβ1−42 and/or Aβ1−40); inclusion of a control group; reporting of at least one relevant outcome domain, including neurochemical, molecular, histological, or behavioral outcomes relevant to AD–associated pathology.
Studies relying exclusively on transgenic models, in vitro systems, or peripheral amyloid administration were excluded. Conference abstracts, narrative commentaries, and articles lacking sufficient methodological detail were also excluded.
Study selection and data extraction
Study selection was performed in a stepwise manner. Titles and abstracts were screened for relevance, followed by full-text evaluation of potentially eligible articles. When uncertainties regarding eligibility arose, studies were discussed and inclusion was determined based on the clarity of experimental design and relevance of reported outcomes.
From each included study, information was extracted regarding animal characteristics, Aβ peptide type, peptide preparation and solvent, administered dose, injection paradigm and targeted brain region, timing of outcome assessment, and reported endpoints. Extracted outcomes were categorized into predefined thematic domains, including neuroinflammatory markers, oxidative and mitochondrial parameters, synaptic and neuronal integrity markers, and behavioral correlates. This classification guided the organization of the review and facilitated comparison of peptide-specific patterns across studies. Doses were extracted as reported and analyzed descriptively. Formal normalization across studies was not feasible due to variability in reporting formats, injection volumes, and concentration units.
Risk of bias assessment
Methodological quality was assessed using the SYRCLE risk-of-bias tool for animal studies. Two reviewers independently evaluated each study across predefined domains, including sequence generation, allocation concealment, blinding, incomplete outcome data, and selective reporting. Domains were rated as low risk only when explicitly described by the authors; otherwise, they were classified as unclear risk. Disagreements were resolved through discussion, and results were summarized descriptively for each bias domain. Overall strength of evidence was evaluated narratively based on indirectness, structural confounding, heterogeneity, and risk-of-bias distribution. Formal assessment of publication bias (e.g., funnel plot asymmetry or regression-based tests) was not feasible due to the absence of standardized effect sizes and the qualitative nature of the synthesis.
Data synthesis
Given the substantial heterogeneity across experimental designs and outcome measures, a quantitative meta-analysis was not performed. Instead, findings were synthesized using a qualitative approach. Evidence was organized according to major neurochemical domains, neuroinflammation, oxidative and mitochondrial dysfunction, synaptic and neuronal integrity, and associated behavioral outcomes. This domain-based synthesis was selected to support mechanistic interpretation and to clarify how Aβ peptide selection shapes neurochemical outcomes in intracerebral models of AD.
To explore potential structural confounding, descriptive stratified analyses were conducted according to injection route (i.h.c. versus i.c.v.) and aggregation state. Frequency distributions were examined within each subgroup to assess clustering patterns between peptide types.
The feasibility of quantitative synthesis was evaluated by assessing consistency of outcome reporting, availability of variance measures, and methodological comparability within potential subgroups (route + aggregation state). Effect size extraction was not feasible in most studies due to inconsistent reporting formats and the absence of standardized outcome metrics.
Results
Study selection and general characteristics
The literature search identified a total of 83 experimental studies investigating intracerebral Aβ peptide administration in a rat model of AD and met the inclusion criteria. Of the selected papers, 23 employed Aβ1−40, whereas 60 studies used Aβ1−42. A flow diagram summarizing the study selection process is presented in Figure 1.
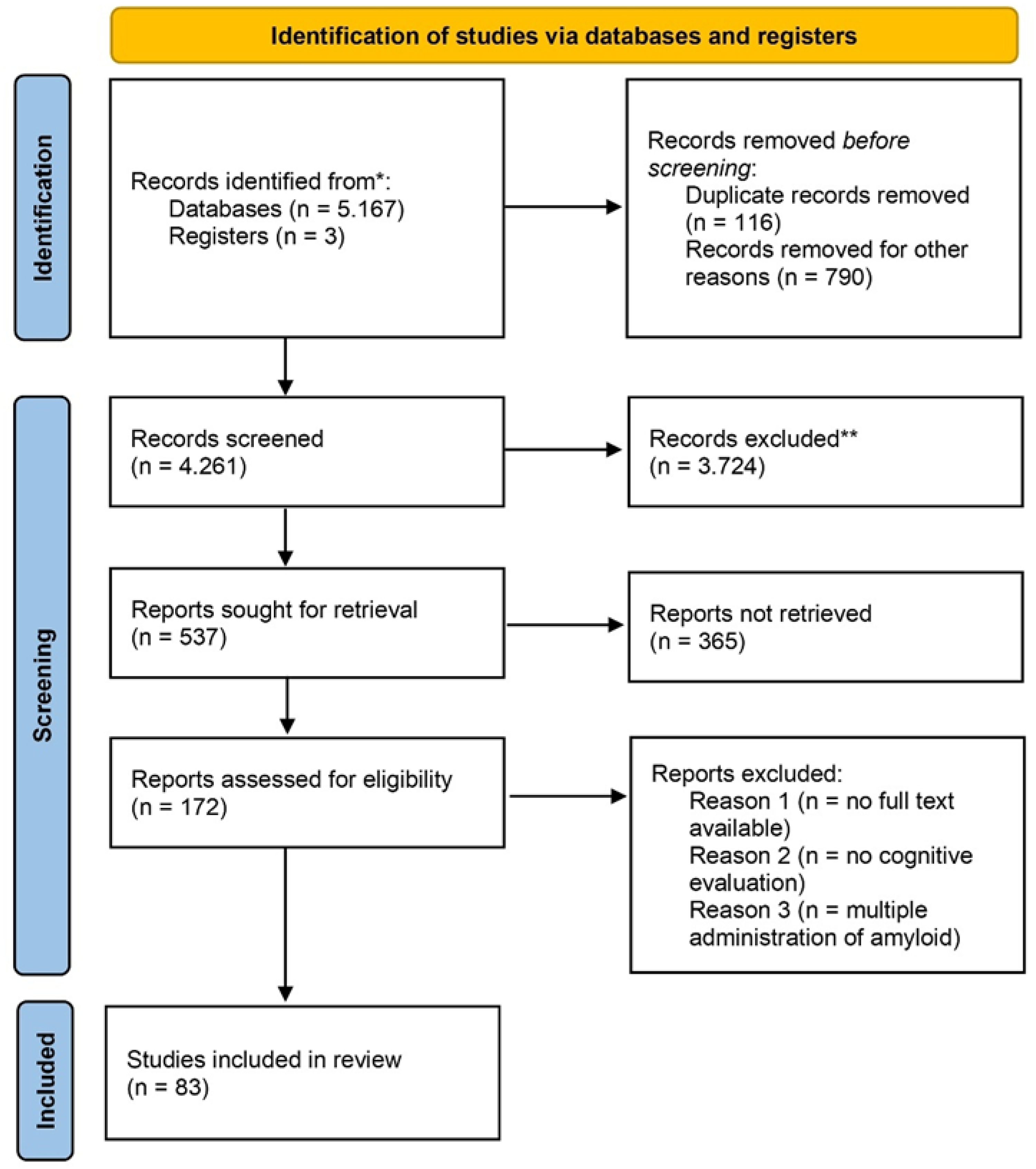
PRISMA 2020 flow diagram of study selection.
Across studies, group sizes were modest, with a median of 8.5 (AD) and 8 (control). Wistar rats were the most frequently used strain (n = 54), followed by Sprague–Dawley rats (n = 27), with isolated use of Charles-Foster and Holtzman strains. Sex reporting was markedly imbalanced, with most studies using male animals exclusively; only a small subset included female or mixed-sex cohorts.
Considerable heterogeneity was observed with respect to peptide formulation, aggregation state, solvent use, injection route (intracerebroventricular or intrahippocampal), laterality, administered dose, and timing of outcome assessment (Table 1).
Characteristics of the included intracerebral Aβ rat studies stratified by peptide species (Aβ1−40 and Aβ1−42), including injection route (i.h.c. or i.c.v.), aggregation state, dose, survival interval, and principal behavioral and neurochemical outcomes.
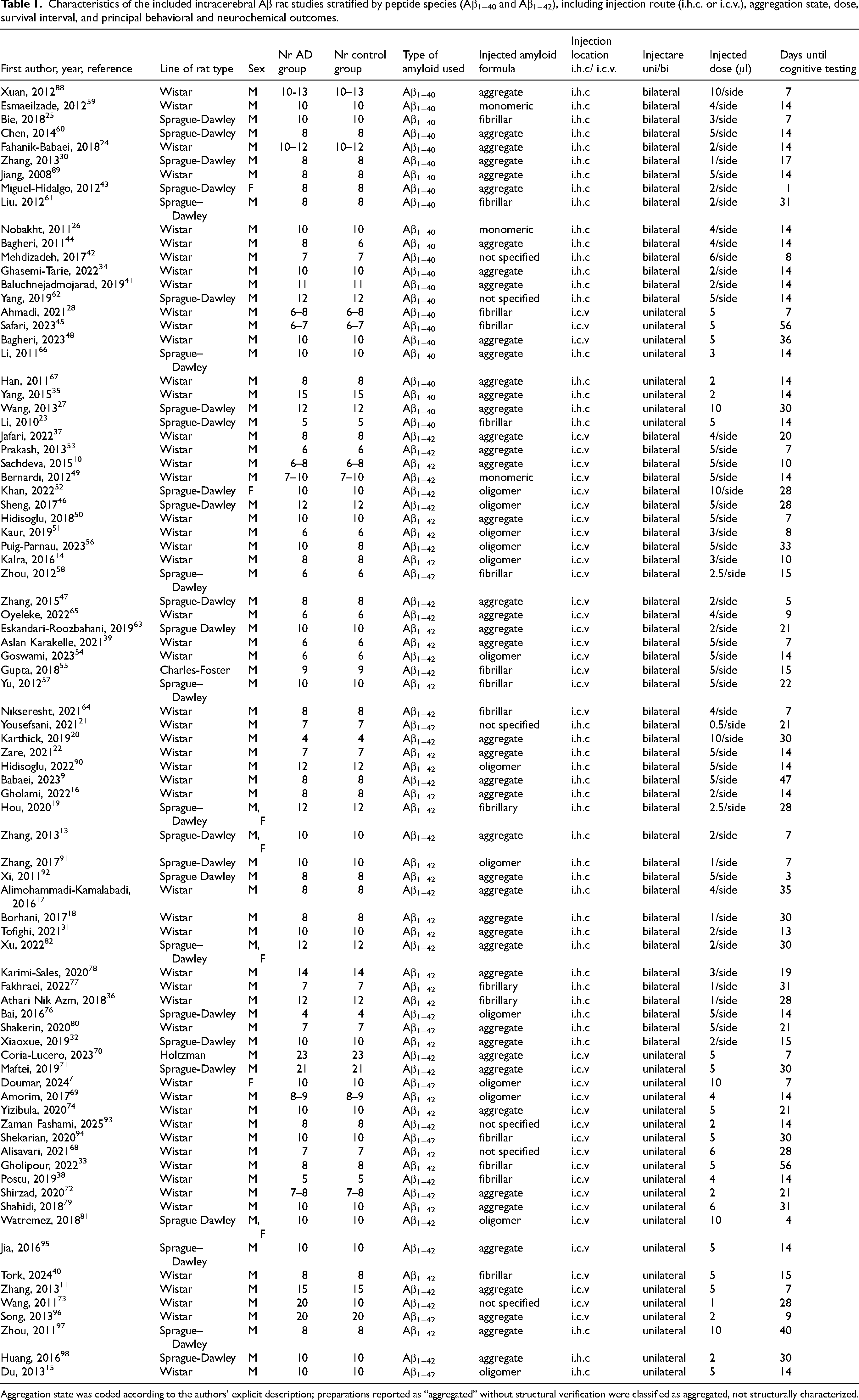
Aggregation state was coded according to the authors’ explicit description; preparations reported as “aggregated” without structural verification were classified as aggregated, not structurally characterized.
According to SYRCLE criteria, explicit random sequence generation was reported in 77 of 83 studies (92.8%), whereas allocation concealment was documented in only 1 study (1.2%). Blinding of outcome assessment was reported in 30 studies (36.1%), with the remaining studies rated as unclear risk due to incomplete reporting.
Descriptive analysis revealed clear methodological clustering by peptide species. Studies employing Aβ1−42 more frequently used intracerebroventricular administration (61.7%), whereas Aβ1−40 studies predominantly relied on intrahippocampal injection paradigms (86.9%) (Table 2). Bilateral administration was more commonly reported in Aβ1−42 studies, whereas unilateral paradigms were more frequent among Aβ1−40 models.
Distribution of injection routes across intracerebral amyloid-β rat models, stratified by peptide species.
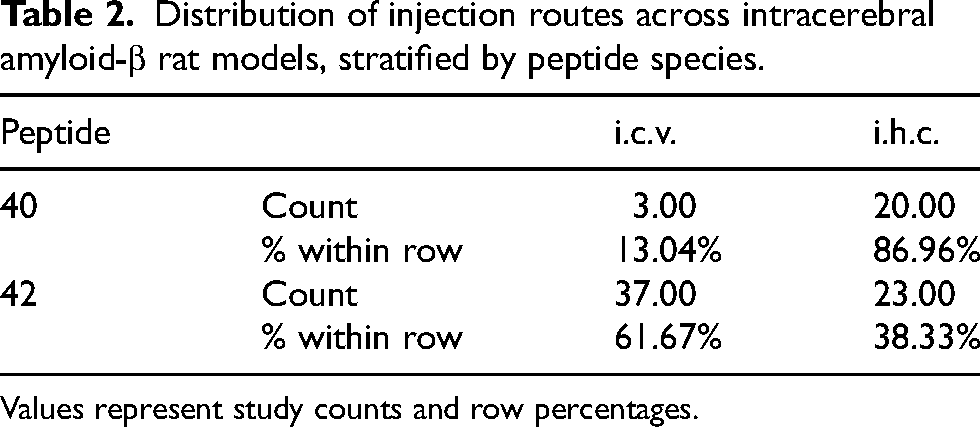
Values represent study counts and row percentages.
Route-stratified descriptive synthesis and analysis
To examine the potential influence of the injection route of the analyzed amyloid types, a route-stratified descriptive synthesis was performed.
The intrahippocampal route (i.h.c.) was comparably represented (Aβ1−42: n = 23; Aβ1−40: n = 20), allowing a balanced cross-peptide comparison. Within this subgroup, we examined the distribution of aggregation states (Supplemental Table 1). In both peptide groups, most preparations were described as aggregated without structural characterization (65.2% for Aβ1−42 versus 65.0% for Aβ1−40), with similar proportions of fibrillar forms (13.0% versus 15%, respectively). Oligomer-enriched preparations were limited to Aβ1−42 studies (17.4%), whereas monomeric preparations were reported only in Aβ1−40 models (10%). Overall, aggregation-state distribution within the i.h.c. subgroup was not markedly skewed toward one peptide species.
Within intrahippocampal paradigms, dose ranges did not significantly differ between peptides (p = 0.587). Survival intervals showed moderate differences in central tendency (p = 0.010), although ranges substantially overlapped. Together, these findings indicate partial methodological comparability within the intrahippocampal subgroup.
The intracerebroventricular route (i.c.v.) demonstrated greater structural diversity among Aβ1−42 studies, with 24.3% oligomer-enriched, 21.6% fibrillar and the majority aggregated but not structurally characterized (43.2%). Given the substantial numerical imbalance in this subgroup (Aβ1−42 = 37; Aβ1−40 = 3), subgroup analysis remains limited and precludes definitive route-controlled inference regarding peptide-specific differences. Detailed frequency tables for the i.c.v. route distribution are provided in Supplemental Table 2.
Peptide species were significantly associated with injection route (χ2(1) = 15.74, p < 0.001, Cramér's V = 0.436), indicating structural clustering between Aβ1−42 and intracerebroventricular paradigms (Supplemental Table 3). Aggregation-state distribution was not significantly associated with peptide species either globally (χ2(4) = 7.71, p = 0.103) or when stratified by injection route (i.c.v.: p = 0.498; i.h.c.: p = 0.178) (Table 3; Supplemental Table 4).
Structural clustering analyses between peptide species, injection route, and aggregation state.

Chi-square tests were performed to evaluate associations between peptide species (Aβ1−40 versus Aβ1−42) and methodological variables. Effect size is reported as Cramér's V. Stratified analyses were conducted separately for intracerebroventricular (i.c.v.) and intrahippocampal (i.h.c.) paradigms.
Sex distribution descriptive analysis
Sex distribution was markedly unbalanced across studies. In intrahippocampal paradigms, 90.7% of experiments were conducted exclusively in male animals, 2.3% in females, and 7% in mixed cohorts. A similar predominance of male animals was observed in intracerebroventricular models (92.5% male-only). No study directly compared Aβ1−40 and Aβ1−42 under identical experimental conditions in female cohorts (Supplemental Table 5).
Neuroinflammatory responses following intracerebral amyloid-β administration
Across the included literature, neuroinflammatory activation emerged as the most consistently reported neurochemical response following intracerebral administration of Aβ. However, the magnitude and temporal profile of this response differed substantially depending on peptide species.
Studies employing Aβ1–42 commonly reported early and robust inflammatory response characterized by microglial activation, astrogliosis, evidenced by increased expression of markers such as Iba1 and GFAP, together with elevated levels of pro-inflammatory cytokines, including interleukin-1β, tumor necrosis factor-α, and interleukin-6.9–11 These inflammatory changes were frequently detected within days to weeks after injection and were often localized to regions adjacent to the injection site, although more widespread inflammatory signaling was reported in some paradigms.10,12–22
In contrast, studies using Aβ1−40 generally reported a more attenuated inflammatory profile. Glial and astrocytic activation was less pronounced and, in several cases, emerged only at later time points. Cytokine alterations were either attenuated or absent compared with those observed following Aβ1−42 administration.23,24 When present, inflammatory changes associated with Aβ1–40, they appeared to be more sensitive to experimental parameters such as peptide dose, aggregation state, and survival interval.24–28
Comparative studies directly assessing both peptides within the same experimental framework consistently supported these observations. Aβ1−42 induced stronger and more sustained activation of glial cells and inflammatory mediators, whereas Aβ1−40 produced subtler responses that were often contingent on higher doses or extended survival intervals. These findings indicate that peptide choice is a critical determinant of the inflammatory phenotype elicited by intracerebral Aβ models.3,29,30
Oxidative stress and mitochondrial alterations
Markers of oxidative stress and mitochondrial dysfunction were frequently assessed alongside inflammatory outcomes, revealing distinct peptide-associated patterns. Following Aβ1−42 administration, numerous studies reported increased lipid peroxidation, elevated reactive oxygen species production, and reductions in endogenous antioxidant defenses, including superoxide dismutase, catalase, and glutathione levels.14,21,24 Mitochondrial impairments, such as altered membrane potential and reduced activity of respiratory chain complexes, were also described, particularly at early post-injection time points.31–40
By comparison, oxidative alterations associated with Aβ1−40 were less consistent and generally less severe. Some studies reported mild increases in oxidative stress markers, whereas others observed no significant changes relative to controls.41,42 When mitochondrial parameters were examined, Aβ1−40 tended to induce modest functional alterations without the pronounced disruption commonly attributed to Aβ1−42. 43 These differences suggest that oxidative and mitochondrial pathways are preferentially engaged by Aβ1−42, in line with its higher aggregation propensity and neurotoxic potential.24,28,30,41,42,44,45
Synaptic and neuronal integrity markers
Alterations in synaptic and neuronal integrity were reported across a substantial proportion of studies following intracerebral Aβ administration. Experiments employing Aβ1−42 described reductions in presynaptic and postsynaptic markers, including synaptophysin and postsynaptic density protein 95 (PSD-95), often accompanied by evidence of neuronal stress or loss reflected by changes in NeuN immunoreactivity or neuronal density within hippocampal and cortical regions.46,47
In contrast, studies utilizing Aβ1−40 more often reported preserved synaptic marker expression at early time points, with reductions observed only after prolonged survival intervals or higher peptide doses. Evidence of overt neuronal loss was less consistently reported following Aβ1−40 administration, suggesting a comparatively lower impact on neuronal integrity under otherwise similar experimental conditions.43,45,48
Behavioral correlations with neurochemical alterations
Behavioral impairments were assessed in a large subset of studies and generally mirrored the neurochemical profiles induced by Aβ administration. Experiments employing Aβ1−42 commonly reported deficits in spatial learning, memory performance, and recognition tasks, with impairments often detectable within weeks of injection. These behavioral changes were frequently interpreted in the context of concurrent inflammatory activation, oxidative stress, and synaptic disruption.49–58
By contrast, behavioral outcomes following Aβ1−40 administration were more heterogeneous. Some studies reported subtle or delayed cognitive deficits, whereas others observed no significant behavioral impairment despite detectable molecular or histological alterations. In several paradigms, behavioral deficits emerged only at later time points, suggesting a dissociation between early neurochemical alterations and functional outcomes.59–62
Peptide-specific temporal trajectories
Timing of outcome assessment emerged as a critical factor influencing reported findings. Aβ1−42 administration was consistently associated with early-onset neurochemical alterations, including rapid glial activation, oxidative stress, and synaptic disruption, which often preceded or coincided with measurable behavioral deficits.63–65
In contrast, Aβ1−40 induced changes generally followed a more protracted course. Neurochemical alterations, when present, developed gradually and were highly sensitive to experimental parameters such as dose, aggregation state, and duration of survival. 45 Behavioral deficits associated with Aβ1−40 were typically delayed and less pronounced, reinforcing the notion that this peptide models a distinct and slower-evolving pathological process.59,66,67
Timepoints were examined as continuous variables across studies. Within intrahippocampal paradigms, substantial overlap was observed between peptides. Notably, Aβ1−42 studies were, on average, assessed at later timepoints compared to Aβ1−40 studies, indicating that the earlier neuroinflammatory and synaptic alterations associated with Aβ1−42 cannot be attributed to earlier outcome measurement (Supplemental Table 6).
Methodological heterogeneity of reported outcomes
Across all outcome domains, substantial methodological heterogeneity was observed. Differences in peptide preparation protocols, solvent use, aggregation state, administered dose, injection site, laterality, and timing of outcome assessment contributed to variability in reported neurochemical and behavioral findings. This heterogeneity appeared to disproportionately affect studies employing Aβ1−40 in which peptide-specific effects were more easily obscured by experimental design factors.58,68–74 These observations highlight the need to consider methodological context when comparing outcomes across studies and reinforce the importance of peptide-specific framing in experimental design.
Marked differences in peptide preparation were observed across studies and are illustrated in Figure 2. Aβ1−42 models more frequently employed aggregated and oligomeric peptide preparations, whereas Aβ1−40 studies showed a higher proportion of monomeric and fibrillar forms. These differences highlight systematic methodological clustering by peptide species. The differences were observed across both intracerebroventricular and intrahippocampal paradigms.
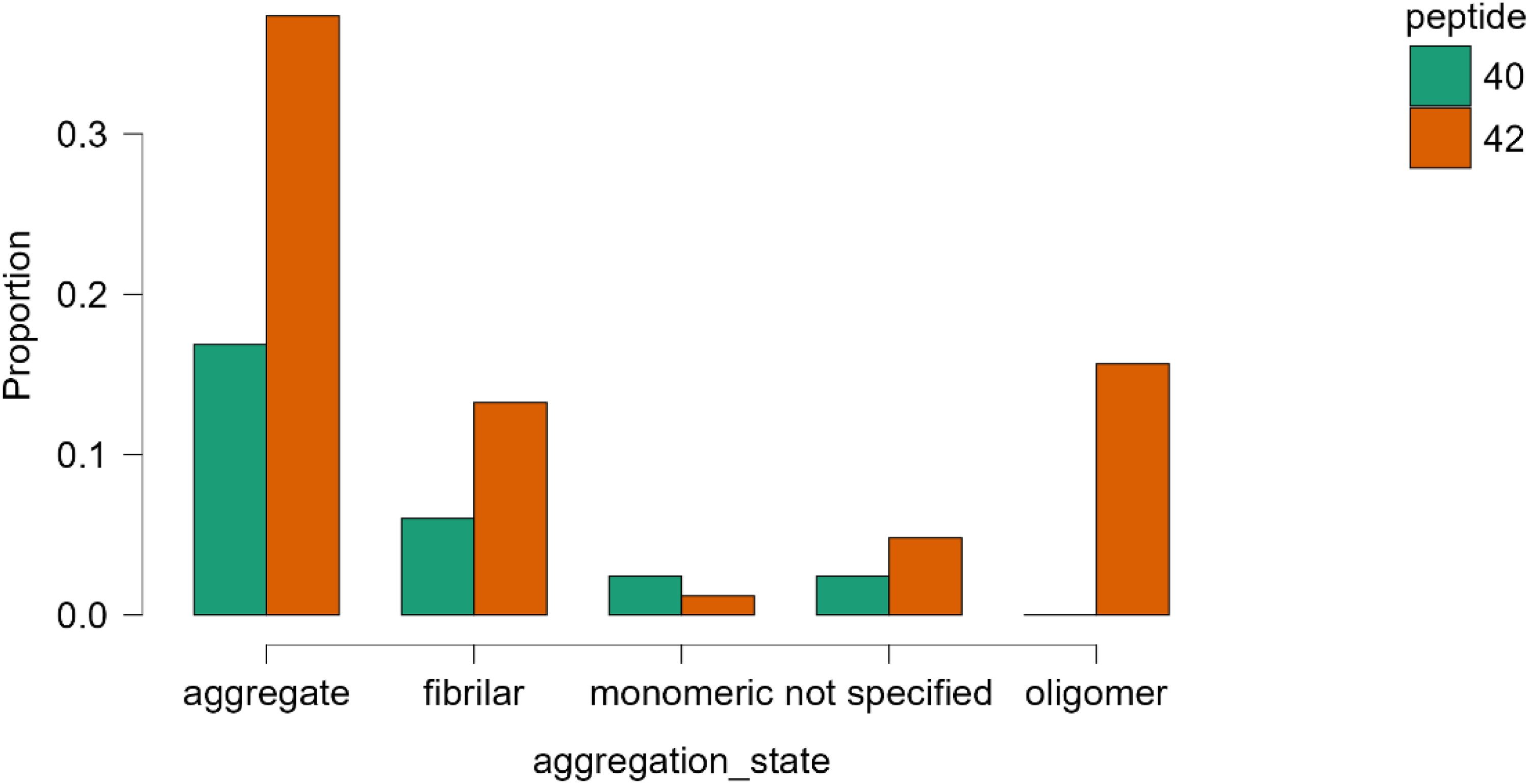
Distribution of amyloid-β aggregation states across intracerebral rat models, stratified by peptide species. Bars represent conditional proportions within each peptide group. Preparations described as “aggregated” or incubated under aggregation-promoting conditions without structural verification were classified as aggregated, not structurally characterized.
Although subgroup homogeneity was explored (route + aggregation state), quantitative pooling was not feasible due to limited overlap in outcome measures, inconsistent variance reporting, and substantial methodological heterogeneity.
Discussion
The present systematic review indicates that intracerebral Aβ injection models do not represent a uniform experimental paradigm but rather encompass peptide-specific neurochemical trajectories with distinct inflammatory, oxidative, synaptic, and behavioral profiles. Across the literature, Aβ1−42 and Aβ1−40 consistently diverge not only in the magnitude of their effects but also in their temporal evolution and mechanistic emphasis. 3 These differences have important implications for experimental design, data interpretation, and translational relevance in preclinical AD research.75–82
The distinct aggregation kinetics of Aβ1−42 and Aβ1−40 are rooted in their biophysical differences at the C-terminus. The additional isoleucine and alanine residues in Aβ1−42 increase hydrophobicity and promote β-sheet stabilization, facilitating faster nucleation and oligomer formation. 83 These structural features enhance the stability of soluble oligomeric assemblies, which are widely implicated in synaptic dysfunction and microglial activation.84,85 In contrast, Aβ1−40 exhibits slower aggregation kinetics and a reduced propensity to form stable oligomeric intermediates. These intrinsic biophysical differences may provide a theoretical mechanistic framework for the earlier and more pronounced neuroinflammatory and synaptic alterations frequently observed in Aβ1−42 models.86,87
One of the most robust and reproducible distinctions emerging from this synthesis concerns the neuroinflammatory response. Intracerebral administration of Aβ1−42 is consistently associated with an early and pronounced activation of glial cells, accompanied by increased expression of pro-inflammatory mediators and downstream oxidative stress.10,12–22 This inflammatory milieu appears to set the stage for subsequent synaptic dysfunction and behavioral impairment. In contrast, Aβ1−40 elicits a more variable and generally attenuated inflammatory response, often dependent on dose, aggregation state, or extended survival intervals. 88 These observations suggest that Aβ1−42 may engage innate immune signaling pathways quicker, whereas Aβ1−40 may model subtler or alternative pathological processes.24–28,30
Closely linked to inflammatory activation are alterations in oxidative stress and mitochondrial function, which also display peptide-specific patterns. Aβ1−42 is frequently associated with increased reactive oxygen species production, lipid peroxidation, and impairment of endogenous antioxidant defenses, together with mitochondrial dysfunction. These changes are often detected at an early time point and may amplify inflammatory signaling and neuronal vulnerability.46,47 By comparison, oxidative and mitochondrial alterations following Aβ1−40 administration are less consistent and generally less severe, reinforcing the notion that oxidative stress pathways are preferentially engaged by Aβ1−42 in intracerebral models.3,48,66
Differences in synaptic and neuronal integrity further support the existence of divergent peptide-driven pathological cascades. Reductions in synaptic markers and evidence of neuronal compromise are commonly reported following Aβ1−42 administration, often in parallel with inflammatory and oxidative changes. These findings align with the view that synaptic dysfunction represents an early and sensitive consequence of Aβ1−42 induced neurochemical stress. In studies employing Aβ1−40, synaptic alterations are more heterogeneous and frequently delayed, suggesting that synaptic vulnerability may arise as a secondary or cumulative effect rather than as a primary response to peptide exposure.
Behavioral outcomes, while frequently reported, should be interpreted cautiously in isolation. Across studies, cognitive impairments tend to emerge in the context of converging neurochemical disturbances rather than as standalone effects of peptide administration.49–58 The more consistent behavioral deficits observed following Aβ1−42 injection align with its robust molecular impact, whereas the variable behavioral findings associated with Aβ1−40 reinforce the notion that functional impairment depends on both peptide-specific properties and experimental context.59–62,66 Behavioral variability observed in Aβ1−40 models may partially reflect heterogeneity in pre-injection preparation. A minority of studies employed explicitly monomeric preparations, whereas others utilized fibrillar or aggregated forms. However, variability in behavioral outcomes was not restricted to monomeric preparations and was also observed in fibrillar models, suggesting that aggregation state alone does not fully account for the heterogeneous behavioral profile associated with Aβ1−40.
A key contribution of this review is the identification of distinct temporal trajectories associated with each peptide. Aβ1−42 is characterized by a relatively compressed timeline from molecular disturbance to functional impairment, whereas Aβ1−40 follows a more protracted and context-dependent course. These temporal differences suggest that the two peptides do not merely differ in toxicity but instead model partially distinct aspects of AD–related pathology. Consequently, peptide selection should be aligned with the specific research question under investigation, particularly when targeting early inflammatory mechanisms versus longer-term synaptic or vascular-related changes.
Timepoints were not pre-categorized into acute, subacute, or chronic phases due to the heterogeneity of experimental designs and reporting across studies. Distributional analyses demonstrated substantial overlap between peptides within comparable paradigms. Importantly, Aβ1−42 models were not evaluated at earlier timepoints; therefore, the observed earlier pathological signatures are unlikely to reflect measurement bias related to timing. These temporal patterns are presented as synthesis-level observations rather than rigid phase-defined contrasts.
Dose variability represents a potential confounder in cross-study synthesis. However, within intrahippocampal paradigms, where peptide representation was balanced, dose ranges were comparable between Aβ1−40 and Aβ1−42. These findings suggest that the stronger inflammatory and synaptic signatures observed in Aβ1−42 models are unlikely to be explained solely by systematic dose differences. Nevertheless, residual dose heterogeneity cannot be fully excluded as a contributing factor.
Methodological heterogeneity remains a major challenge in this field. Variability in peptide preparation, solvent use, aggregation protocols, dosing regimens, injection sites, and timing of assessments contributes substantially to inconsistent findings, particularly in studies employing Aβ1−40.61,67 While such heterogeneity complicates cross-study comparisons, it also highlights the sensitivity of intracerebral amyloid models to experimental parameters. Importantly, the present synthesis suggests that some apparent discrepancies in the literature may reflect differences in peptide selection rather than true contradictions in biological effect.58,63–65,68–74
In this context, disentangling peptide-intrinsic effects from structural clustering becomes essential. Although aggregation state may contribute to divergent neurochemical trajectories, given the higher inflammatory and synaptotoxic potential of aggregated and oligomeric species, our stratified analyses indicate that peptide-associated differences do not appear to be fully explained by structural clustering. It should be noted, however, that oligomer-enriched preparations were reported exclusively in Aβ1−42 studies. Although this asymmetry did not reach statistical significance, the relatively smaller Aβ1−40 subgroup may limit statistical power to detect structural skewing. Accordingly, the absence of statistical association should not be interpreted as definitive equivalence in aggregation-state distribution.
While the injection route was significantly associated with peptide species, the aggregation-state distribution was not statistically skewed either globally or within route-specific subgroups. Accordingly, the present review does not argue for the superiority of one peptide over the other but rather underscores their contextual justification as experimental tools.
Even though no study directly compared Aβ1−40 and Aβ1−42 under identical experimental conditions, intrahippocampal paradigms provided a partially matched methodological context. Dose ranges were comparable; however, Aβ1−42 studies tended to assess outcomes at later time points. This temporal asymmetry represents a potential structural confounder and limits causal inference. Accordingly, conclusions are framed as synthesis-level patterns rather than definitive peptide-specific effects established under controlled head-to-head designs.
Several limitations of the present review should be acknowledged. Despite adherence to PRISMA guidelines and prospective protocol registration, the synthesis is constrained by the quality and reporting standards of the included studies. The lack of uniform outcome measures and the predominance of male-only animal cohorts limit the generalizability of findings. In addition, the heterogeneity of experimental designs precluded quantitative meta-analysis, necessitating a qualitative and thematic approach. Although random sequence generation was frequently reported, allocation concealment was rarely documented and blinding of outcome assessment was inconsistently described. Most domains were therefore rated as unclear rather than explicitly high risk. These reporting limitations warrant cautious interpretation of cross-study comparisons and reinforce that the present conclusions reflect synthesis-level patterns rather than effect-size–weighted causal inferences.
The overall certainty of evidence supporting strict non-interchangeability is best characterized as low to moderate. This rating reflects indirect cross-study comparisons, structural clustering by injection route, residual heterogeneity in dose and timing, and incomplete reporting in key risk-of-bias domains. At the same time, consistency of peptide-associated patterns within the more methodologically comparable intrahippocampal subgroup supports the interpretation that Aβ1−40 and Aβ1−42 should not be assumed interchangeable without explicit contextual justification.
Publication bias and small-study effects cannot be formally excluded. Given the predominance of small preclinical experiments and the absence of standardized effect-size reporting, statistical evaluation of funnel plot asymmetry was not feasible. The possibility that studies reporting stronger peptide-associated effects are overrepresented must therefore be acknowledged. Accordingly, conclusions are framed as synthesis-level trends rather than quantitative magnitude estimates.
The pronounced male predominance across studies represents an important limitation. Given the known modulatory role of estrogen on amyloid-induced neuroinflammatory responses and microglial activation, the absence of controlled female comparisons restricts conclusions regarding potential sex-dependent peptide effects. Future studies incorporating sex-balanced and head-to-head comparative designs will be necessary to clarify whether peptide-associated trajectories differ across biological sex.
In conclusion, the evidence synthesized in this review cautions against treating Aβ1−42 and Aβ1−40 as interchangeable tools in intracerebral amyloid models. Instead, each peptide appears to model distinct aspects of AD–related pathology, with Aβ1−42 appearing to model more consistently rapid inflammatory and synaptotoxic processes and Aβ1−40 reflecting more gradual or context-dependent alterations. Careful alignment of peptide selection with specific experimental objectives is therefore essential for improving interpretability and translational relevance. At the same time, distinguishing peptide-intrinsic biology from structural methodological clustering remains a central interpretative challenge. Formal analyses confirmed significant clustering by injection route, whereas aggregation-state distribution was not statistically skewed between peptides. Within intrahippocampal paradigms, where representation was balanced, dose ranges were comparable, although survival intervals differed moderately. Taken together, these findings suggest that the observed differences are unlikely to be explained solely by systematic methodological asymmetry, while acknowledging that residual confounding cannot be entirely excluded. Conclusions are therefore presented proportionally to the strength and limitations of the available evidence.
Conclusion
Intracerebral Aβ injection models remain a valuable experimental approach for investigating neurochemical mechanisms relevant to AD. The evidence synthesized in this systematic review suggests that these models should not be assumed interchangeable without explicit methodological consideration, as peptide selection appears to influence molecular, cellular, and functional readouts. Across studies, Aβ1−42 is more frequently associated with earlier and more pronounced neuroinflammatory activation, oxidative imbalance, synaptic alterations, and behavioral impairment, whereas Aβ1−40 is generally linked to more variable and context-dependent neurochemical profiles.
Supplemental Material
sj-docx-1-alz-10.1177_13872877261443555 - Supplemental material for How amyloid-β peptide choice shapes neurochemical outcomes in intracerebral rat models of Alzheimer's disease: A systematic review
Supplemental material, sj-docx-1-alz-10.1177_13872877261443555 for How amyloid-β peptide choice shapes neurochemical outcomes in intracerebral rat models of Alzheimer's disease: A systematic review by Loredana Mariana Agavriloaei, Ivona Costachescu, Andrei Szilagyi, Maria-Raluca Gogu, Bogdan-Florin Iliescu and Mihaela Dana Turliuc in Journal of Alzheimer's Disease
Footnotes
Acknowledgements
The authors have no acknowledgments to report.
Ethical considerations
Not applicable
Consent to participate
Not applicable
Consent for publication
Not applicable
Author contribution(s)
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was funded by Grigore T. Popa University of Medicine and Pharmacy in Iasi, grant number 10060/15.05.2025 and doctoral program.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data availability statement
The datasets analyzed during the current study are not publicly available, as this work represents a qualitative synthesis of previously published data, but are available from the corresponding author upon reasonable request.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.