
Abstract
Background
Chronic inflammation contributes to neurodegeneration and cerebrovascular injury, yet it remains unclear whether inflammatory biomarkers influence white matter hyperintensity (WMH) volume and cognition independent of Alzheimer's disease (AD) plasma biomarkers.
Objective
This study examined whether inflammatory biomarkers were associated with WMH volume and multidomain cognitive performance independent of AD plasma biomarkers, and whether WMH volume mediated associations between IL-6 and cognition.
Methods
Using data from 1806 participants in the Health and Aging Brain Study–Health Disparities (HABS-HD), we examined associations between log transformed plasma inflammatory biomarkers (IL-10, TNF-α, IL-6, and IL-5) and WMH volume and cognitive performance (memory, executive function, processing speed, language). Multivariable linear regression models adjusted for demographic and vascular risk factors were repeated with additional adjustment for AD biomarkers (p-Tau181, NfL, total tau, Aβ42/40). Mediation analyses were included to test whether WMH meditated the association between IL-6 and cognitive domains.
Results
Higher IL-6 levels were consistently associated with increased WMH volume in base and AD-adjusted regression models. Increased levels of IL-6 were also associated with poorer performance in all cognitive domains, but these associations did not survive multiple comparisons correction. WMH partially mediated the association between IL-6 and memory (23%) and processing speed (19%).
Conclusions
Elevated IL-6 is independently linked to cerebrovascular injury and poorer cognition beyond classical AD biomarkers, supporting a vascular–inflammatory pathway distinct from amyloid and tau pathology. These findings underscore the importance of integrating inflammatory and neurodegenerative markers to elucidate heterogeneous mechanisms of brain aging across diverse populations.
Keywords
Introduction
Chronic systemic inflammation plays a pivotal role in the pathogenesis of neurodegenerative diseases, including Alzheimer's disease (AD). 1 Inflammatory processes have been implicated in neuronal injury, synaptic dysfunction, and cerebrovascular dysfunction which are also known to be underlying pathological mechanisms influencing cognitive decline and dementia. 2 Studies have demonstrated that a persistent immune response in the brain exacerbates both amyloid-β (Aβ) and neurofibrillary tangles (NFT) pathologies, suggesting that the inflammatory response may provide a link between the initial Aβ pathology and the later development of NFT.3,4 Given the difficulty of directly measuring neuroinflammation in vivo, identifying reliable peripheral biomarkers for not only systemic inflammation but also associated brain changes and symptomology is crucial for the clinical assessment of AD neuropathology.
Among peripheral inflammatory markers, interleukin-6 (IL-6) and high-sensitivity C-reactive protein (hs-CRP) have been most consistently linked to adverse brain outcomes. Elevated IL-6 levels have been associated with increased risk of dementia onset, increased white matter hyperintensity (WMH) volumes, reduced gray matter volume, and impaired performance across multiple cognitive domains including executive function and memory.5–7 Meta-analytic and longitudinal findings confirm IL-6 as a significant predictor of accelerated cognitive decline and incident dementia, independent of traditional vascular risk factors.8,9 However, IL-6 has not been included in larger frameworks for neurodegenerative diagnoses and disease tracking along their respective continuums like AD.
The National Institute on Aging–Alzheimer's Association (NIA-AA) have reframed AD using core biological markers of amyloid (A), tau (T), and neurodegeneration (N), the AT(N) framework. 10 There has been a large push to develop and evaluate plasma-based biomarkers for their reduced cost and to increase accessibility for probable AD prior to advising individuals to pursue more invasive measurements like positive emission topography and lumbar punctures for cerebral spinal fluid assessment. AT(N) plasma-based biomarkers exist for Aβ42/40 (A), phosphorylated tau (p-tau181 and p-tau217) (A and T), neurofilament light chain (NfL) (N), and glial fibrillary acidic protein (GFAP) (I-inflammation). These biomarkers have demonstrated strong prognostic utility, predicting cognitive decline and disease progression in both cognitively normal and mildly impaired individuals.11–13 In addition, several studies have also linked these plasma markers to WMH volume, with NfL and p-Tau217 showing consistent associations with white matter damage and cognitive deterioration.14,15 Although differentially abundant proteins regulating neuroinflammation and immune signaling have been associated with these AD and related dementias plasma biomarkers, 16 the operationalization of the role of inflammatory biomarkers such as IL-6 in neurodegenerative pathological processes remains vacant. Importantly, there is a limitation in the understanding of whether systemic peripheral plasma inflammatory markers such as IL-6 is independently associated with WMH volume and cognition independent of the AT(N) framework.
As stated earlier, increased IL-6 has been previously found to be associated with increased WMH volume and cognitive decline, most notably in global cognition. However, prior studies have three key limitations: 1) not accounting for plasma-based AT(N) biomarkers, 2) smaller sample sizes, and 3) conducted in primarily non-Hispanic White and European populations. To date, no study has evaluated whether associations between peripheral inflammatory markers and WMH volume or cognition remain significant independent of AT(N) biomarkers, nor have such associations been explored across distinct cognitive domains.
The present study aimed to address these critical gaps through two primary objectives: (1) to examine associations between plasma inflammatory markers (IL-10, tumor necrosis factor-alpha (TNF-α), IL-6, and IL-5) and WMH volume and four cognitive domains, and (2) to evaluate whether these associations remain significant after adjusting for plasma AT(N) biomarkers. We hypothesized that elevated levels of inflammatory biomarkers would be associated with greater WMH volume and poorer performance across all cognitive domains, even after controlling for AT(N) markers, suggesting an independent vascular–inflammatory pathway. Given the observed associations between IL-6, WMH volume, and multidomain cognition, we conducted mediation analyses to determine whether WMH volume accounted for the association between IL-6 and cognitive performance. This study utilizes cross-sectional data from the Health and Aging Brain Study—Health Disparities cohort, which includes Non-Hispanic White (NHW), Non-Hispanic Black (NHB), and Hispanic participants, thus addressing key limitations of prior research by incorporating a larger and more racially and ethnically representative population of the United States.
Methods
Study design
The Health and Aging Brain Study–Health Disparities (HABS-HD) is a prospective, community-based cohort established at the Institute for Translational Research, University of North Texas Health Science Center (Fort Worth, TX).17,18 The research study recruits older adults who self-identify as NHW, NHB, or Hispanic from the greater Dallas–Fort Worth region through community-engagement activities. All procedures are reviewed and approved by the North Texas Regional Institutional Review Board and comply with the ethical guidelines of the 1975 Declaration of Helsinki. All participants provide written informed consent for study participants and then they complete standardized demographic and health questionnaires, medical and cognitive examinations, and venipuncture for laboratory assays. Neuroimaging includes structural magnetic resonance imaging (MRI) and positron-emission tomography (PET) to quantify β-amyloid and neurofibrillary tau burden. Comprehensive details of the HABS-HD protocol have been published elsewhere.17,18
Inclusion criteria
Participants were included in the analytic cohort using a stepwise complete-case approach across predefined variable domains. Starting from the full received dataset (N = 3833), individuals were first required to have complete demographic data (age, sex, education), resulting in the exclusion of 16 participants. Next, complete data were required for vascular and lifestyle covariates (hypertension, diabetes, dyslipidemia, obesity, tobacco use, and race and ethnicity), excluding an additional 25 participants. Participants were then required to have available cognitive diagnosis and apolipoprotein (APOE) genotype, resulting in the removal of 491 individuals. Complete plasma inflammatory biomarker data (IL-10, TNF-α, IL-6, IL-5) were subsequently required, which reduced the sample by 1408 participants. Availability of neurodegenerative biomarkers (p-tau181, NfL, total tau, and Aβ42/40 ratio) further excluded 77 individuals. Finally, participants were required to have complete cognitive domain scores (memory, executive function, processing speed, and language), resulting in the exclusion of 10 additional participants. The final analytic sample comprised 1806 individuals resulting in a complete-case dataset suitable for statistical analyses to extrapolate the findings across the selective sample.
Demographic questionnaire
HABS-HD participants self-report their race and ethnicity as NHW, NHB, or Hispanic. For each participant, their age at the visit and the numbers of years of education and whether they completed high school or not was documented at the baseline visit.
Covariates
As previously described,19–21 the HABS-HD study characterizes vascular-metabolic risk according to established clinical thresholds and structured interview data:
Hypertension—classified when any of the following were present: (i) self-reported physician diagnosis, (ii) use of antihypertensive medication, or (iii) mean systolic/diastolic blood-pressure from two seated measurements ≥ 140/90 mmHg. Diabetes—identified by self-reported diagnosis, current insulin/oral hypoglycemic therapy, or glycated hemoglobin (HbA1c) > 6.5%. Dyslipidemia—defined by self-report of high cholesterol/triglycerides, use of lipid-lowering agents, total cholesterol ≥ 200 mg/dL, or triglycerides ≥ 150 mg/dL. Obesity—body-mass index (BMI) ≥ 30 kg/m2. Tobacco dependence—ascertained from questionnaires capturing current and past use of any tobacco product.
Apolipoprotein (APOE) genotyping
DNA was isolated from whole blood and genotyped for the two single-nucleotide variants that determine APOE isoforms (rs429358 and rs7412) using TaqMan® assays (Thermo Fisher Scientific) on an Applied Biosystems 7500 real-time PCR system with GTXpress chemistry. Fluorescence clustering yielded the six possible diplotypes: ε2/ε2, ε2/ε3, ε2/ε4, ε3/ε3, ε3/ε4, and ε4/ε4. For the present analysis, participants were grouped as ε4 carriers (≥ one ε4 allele) versus non-carriers.
Plasma biomarkers
Fasting plasma samples were collected and processed in accordance with international guidelines for Alzheimer's disease biomarker studies. 22 Assay preparation was performed using a custom automated StarPlus liquid-handling system (Hamilton Robotics, Reno, NV). Plasma concentrations of biomarkers were measured with single molecule array (Simoa) technology on the HD-X platform (Quanterix, Billerica, MA). Detailed analytic protocols have been described previously.23–25
MRI acquisition and white-matter lesion quantification
Brain MRI was acquired for every participant on a 3T Siemens scanner using an identical imaging protocol across the cohort. Structural anatomy was captured with a high-resolution T1-weighted MPRAGE sequence (TR = 2300 ms, TE = 2.93 ms, field-of-view = 270 mm, 256 × 256 matrix, 1.2 mm slices, isotropic voxels ≈ 1.1 × 1.1 × 1.2 mm). WMH volumes were delineated from co-registered T2-weighted fluid-attenuated inversion-recovery (FLAIR) scans acquired sagittally (TR = 4800 ms, TE = 441 ms, TI = 1650 ms, FOV = 256 mm, voxel size ≈ 1.0 × 1.0 × 1.2 mm). MPRAGE and FLAIR volumes were processed with the Lesion Growth Algorithm implemented in the Lesion Segmentation Toolbox for SPM (www.statisticalmodelling.de/lst.html) to derive total WMH volume. Each lesion mask underwent manual quality control by trained raters who confirmed anatomical plausibility and absence of artefacts; masks with extreme outlier volumes were re-evaluated, and scans exhibiting probable non-vascular pathology were excluded. Less than 5% of segmentations failed this quality check. The full neuroimaging workflow for the HABS-HD study has been detailed previously.17,18
WMH
WMH volume was quantified using raw volumetric measures in mL. The raw WMH distribution was right skewed; therefore, we applied a log-transformed using a small constant (
Clinical cognitive status
Cognitive classification (cognitively unimpaired, mild cognitive impairment [MCI], or dementia) was determined through a standardized multidisciplinary consensus procedure implemented in the HABS-HD cohort.26,27 Diagnostic determinations were based on reported changes in daily functioning from participants and informants, along with performance across a comprehensive neuropsychological test battery. Neuroimaging findings were not incorporated into the diagnostic adjudication process.
Harmonized cognitive domain scores
Following previous methodology,28,29 the raw scores from individual neuropsychological tests were standardized using z-score transformations to facilitate comparability across measures with differing scales. Cognitive domain scores were then harmonized by averaging relevant z-scores for each domain. Memory was calculated as the mean of z-scored scores from the Spanish English Verbal Learning Test immediate and delayed recall, and the Logical Memory I and II tests. Executive function was derived from the average of z-scores from the Digit Span total score and the Digit Symbol Substitution test. Processing speed was calculated as the average of z-scores from Trails A and Trails B completion times; because higher values on timed tasks reflect slower performance, the composite was inverted. Language performance was computed as the mean of z-scores from the Animal Naming and FAS verbal fluency tests. These four harmonized domain scores (memory, executive function, processing speed, and language) served as primary cognitive outcomes in all regression analyses. All domain scores retained approximately normal distributions following transformation.
Statistical analyses
Descriptive and baseline comparisons
Baseline characteristics were summarized by cognitive status using a descriptive table that included demographics, vascular risk factors, APOE ε4, inflammatory biomarkers, neurodegenerative biomarkers, WMH/ICV metrics, and cognitive scores. For continuous variables, omnibus between-group differences were tested using one-way ANOVA, and Tukey HSD post hoc pairwise comparisons were performed when the omnibus ANOVA p-value was significant. For categorical variables, contingency-table analyses were performed using Pearson chi-square tests; Fisher's exact tests were used when expected cell counts were small.
Visualization of biomarker-outcome associations
To visualize cross-sectional relationships, scatterplots were generated for each biomarker-outcome pair, with one biomarker per row and outcomes (WMH volume and four cognitive domains) as columns. Points, fitted linear trends, and confidence bands were displayed separately by cognitive status group to show potential heterogeneity by diagnosis.
Correlation analyses
Correlation analyses were conducted for the full cohort and separately within cognitive-status strata. A mixed-method correlation framework was used: Spearman correlations were applied when either variable in a pair was a biomarker variable, and Pearson correlations were applied otherwise. Correlation matrices were computed using pairwise complete observations and visualized with lower-triangle heatmaps containing numeric correlation coefficients. Supplemental correlation tables reporting coefficient and p-value pairs were also generated for overall and stratified samples, with additional Spearman-based supplemental output for robustness.
Multivariable regression models
For each outcome, two linear regression specifications were estimated. The base model included age, sex, education, vascular risk factors (hypertension, diabetes, dyslipidemia, obesity, tobacco), race and ethnic group, APOE ε4, and inflammatory biomarkers (IL-10, TNF-α, IL-6, and IL-5). The biomarker-adjusted model added neurodegenerative biomarkers (p-Tau181, NfL, GFAP, and Aβ42/40). These model sets were fit for normalized WMH and for each cognitive domain (memory, executive function, processing speed, and language). Coefficients, confidence intervals (CI), and p-values were reported. Forest plots for each outcome, biomarker beta estimates and 95% CI were plotted with standardized visual formatting and per-term labels that included Holm-corrected p-values were used for visualization.
Multiplicity control
Because multiple biomarker-outcome tests were conducted, multiplicity was addressed using Holm family-wise error correction with global adjustment across pre-specified test families which was used as the primary corrected inference metric. For base models, the primary family included 4 inflammatory biomarkers across 5 outcomes (20 tests). For biomarker-adjusted models, the primary family included 8 biomarkers across 5 outcomes (40 tests). Holm-corrected p-values were prioritized for confirmatory reporting.
Collinearity diagnostics
Multicollinearity was evaluated for all base and biomarker-adjusted regression models using variance inflation diagnostics. For models containing multi-degree categorical terms, generalized VIF metrics were converted to `GVIF^(1/(2*Df))` for comparability to standard VIF scales. Diagnostic summaries included the maximum VIF metric per model and counts of terms exceeding common thresholds (>=5 and >=10), with interpretive flags for low, moderate, and high collinearity. There was not any collinearity in the models, so these findings were not reported.
Stratified and sensitivity analyses
Stratified biomarker-adjusted models were estimated separately within unimpaired, MCI, and dementia groups to assess potential effect heterogeneity by cognitive status. A sensitivity analysis added cognitive-status indicator terms to full-sample cognitive outcome models to evaluate whether biomarker associations were independent of diagnostic group membership. Results from these analyses were compared qualitatively with primary models to assess robustness of direction and magnitude of effects.
Mediation analyses
Causal mediation analyses were performed using nonparametric bootstrap procedures (5000) with IL-6 as exposure, normalized WMH volume as mediator, and each cognitive domain (memory, executive dysfunction, processing speed, and language) as outcome in separate models. For each outcome, three models were fit: a mediator model (`M ∼ A + covariates`), an outcome model including mediator (`Y ∼ A + M + covariates`), and a total-effect model excluding mediator (`Y ∼ A + covariates`). Covariates included age, sex, education, vascular risk factors, race and ethnic group, APOE ε4, other inflammatory biomarkers, and neurodegenerative biomarkers. Reported mediation quantities included ACME (indirect effect), ADE (direct effect), total effect, and proportion mediated/ mediation percentages, each with 95% CI and p-values.
Statistical software and general approach
All analyses were conducted in R v4.2.3. Cognitive status was modeled as a three-level factor (unimpaired, MCI, dementia). WMH was analyzed as an intracranial volume-normalized residual score, and cognitive outcomes (memory, executive function, processing speed, and language) were analyzed as z-scores. In regression and mediation models, log-transformed biomarker variables were used as specified in the model formulas.
Results
Participant characteristics by cognitive status
A total of 1806 participants were included: 1387 unimpaired (76.8%), 310 MCI (17.2%), and 109 dementia (6.0%) (Table 1). Mean age increased across groups (unimpaired 66.0 years, MCI 66.5 years, dementia 69.8 years). Post hoc tests showed dementia participants were older than both unimpaired (mean difference 3.80 years, 95% CI 1.78 to 5.82, p < 0.001) and MCI participants (mean difference 3.28 years, 95% CI 1.01 to 5.54, p = 0.002) (Supplemental Table 1). MCI and unimpaired participants did not differ (difference 0.52 years, p = 0.607). Cognitive groups differed in sex (χ2(2) = 18.30, p < 0.001), education (χ2(2) = 18.36, p < 0.001), hypertension (χ2(2) = 14.97, p < 0.001), diabetes (χ2(2) = 12.78, p < 0.01, race (χ2(4) = 46.77, p < 0.001), and APOE ε4 status (χ2(2) = 22.29, p < 0.001). Inflammatory biomarkers did not differ significantly by cognitive group (IL-10 p = 0.646, TNF-α p = 0.168, IL-6 p = 0.584, IL-5 p = 0.782). In contrast, neurodegenerative biomarkers showed clear differences for p-tau181 (F(2,1803) = 51.48, p < 0.001) and NfL (F(2,1803) = 25.95, p < 0.001), Aβ42/40 ratio differed overall (F(2,1803) = 4.38, p = 0.01). For ICV-adjusted WMH residuals, participants with dementia had higher WMH residuals than unimpaired (difference +1.15, 95%: CI 0.77 to 1.54, p < 0.0001) and MCI (difference +0.91, 95% CI: 0.48 to 1.35, p < 0.001). Progressively worse cognitive performance was apparent from unimpaired to MCI to dementia participants across all tested cognitive domains (p < 0.001). The associations between plasma biomarkers with WMH and cognitive domains by cognitive status are shown in Figure 1.
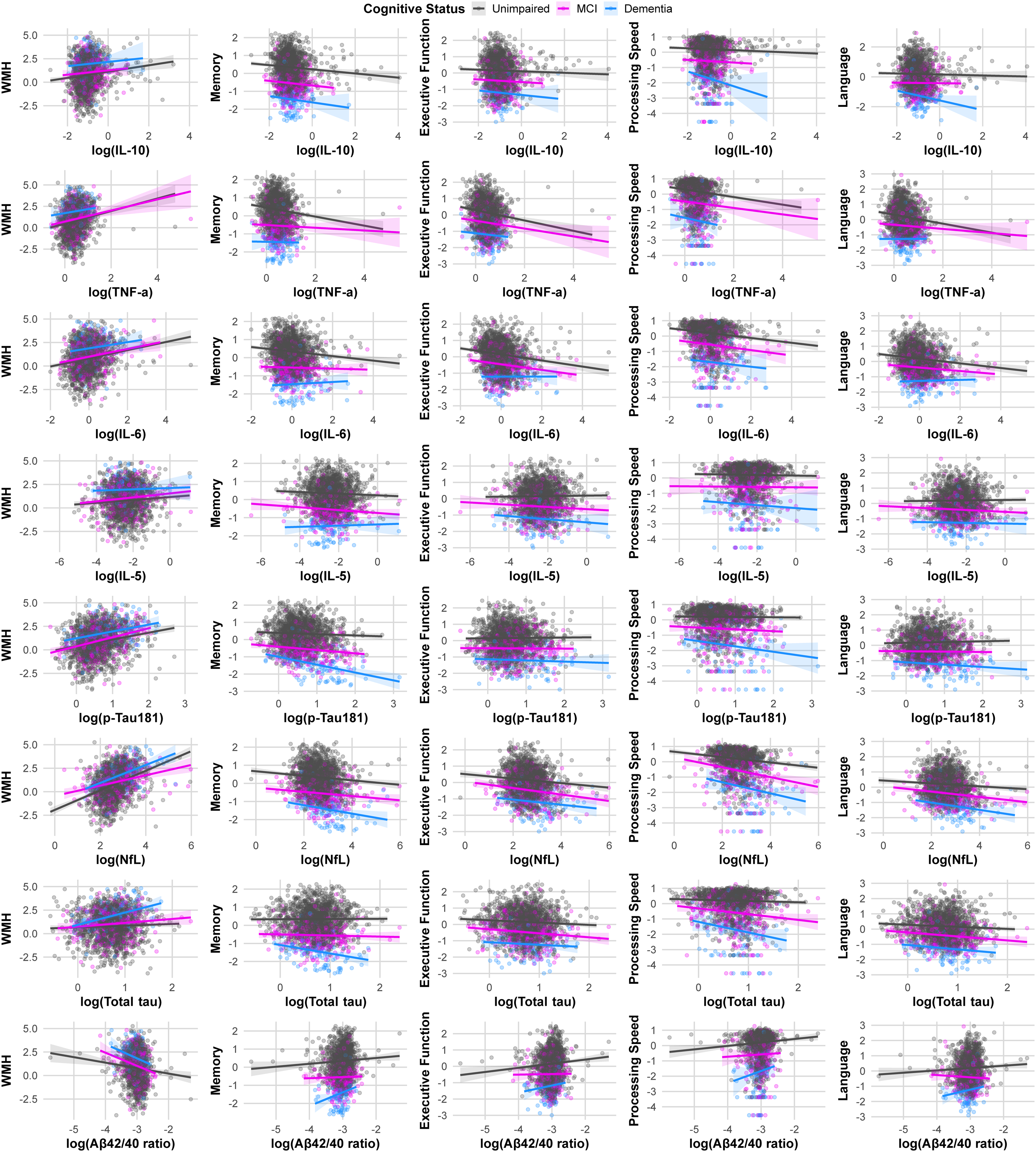
Trajectories between plasma biomarkers with WMH and cognitive domains by cognitive status. Each panel shows a scatterplot with linear fit (solid line) and 95% confidence band for the relationship between one biomarker (x-axis) and one outcome (y-axis). Rows represent biomarkers (IL-6, IL-5, p-tau181, NfL, total tau, and Aβ42/40 ratio), and columns represent outcomes (WMH, memory, executive function, processing speed, and language). Points are individual participants, colored by cognitive group: Unimpaired (gray), MCI (magenta), and Dementia (blue). Biomarker values were natural log-transformed before plotting; non-positive biomarker values were set to missing (Color figure available online).
Participant characteristics stratified by clinical cognitive status.
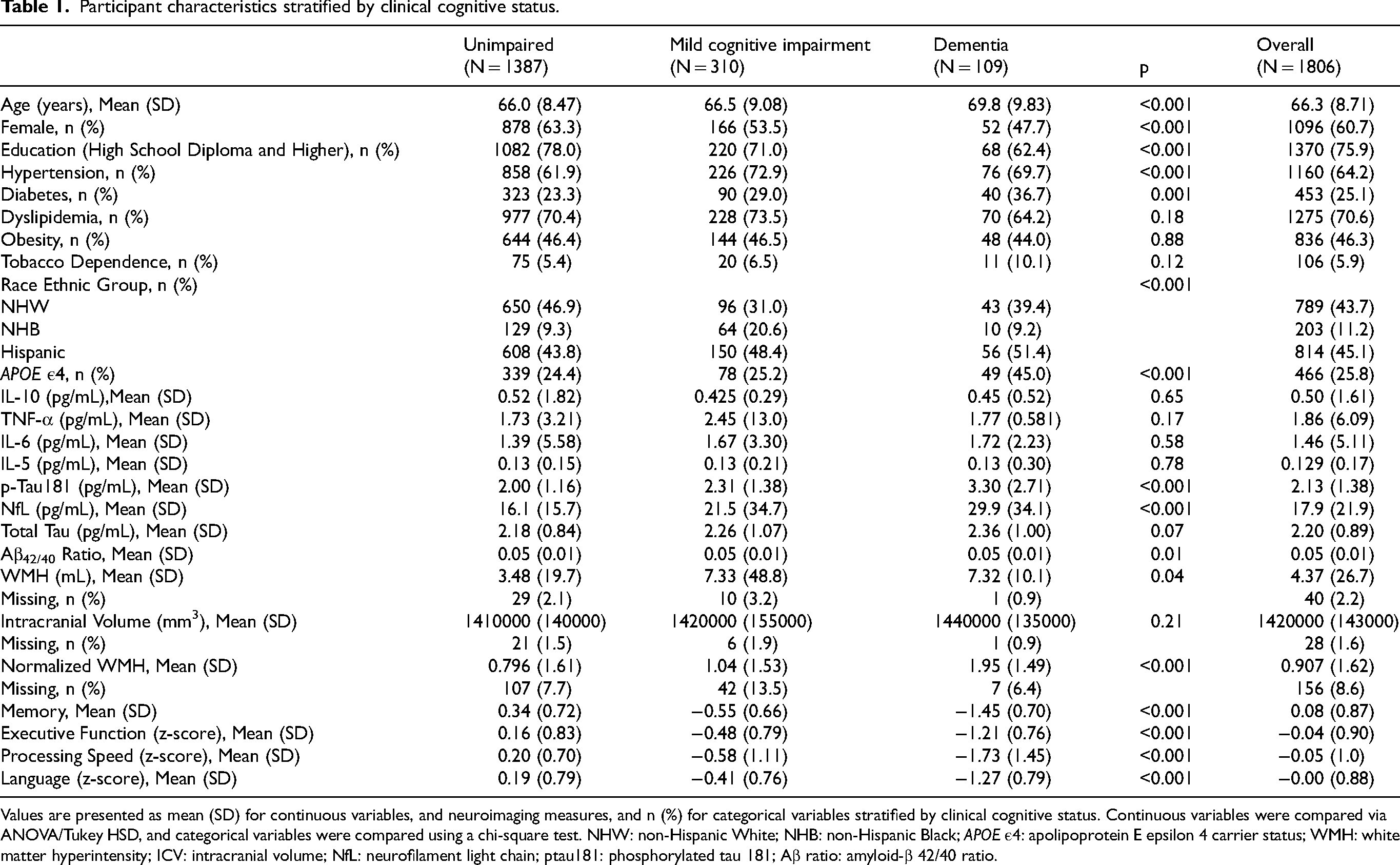
Values are presented as mean (SD) for continuous variables, and neuroimaging measures, and n (%) for categorical variables stratified by clinical cognitive status. Continuous variables were compared via ANOVA/Tukey HSD, and categorical variables were compared using a chi-square test. NHW: non-Hispanic White; NHB: non-Hispanic Black; APOE ε4: apolipoprotein E epsilon 4 carrier status; WMH: white matter hyperintensity; ICV: intracranial volume; NfL: neurofilament light chain; ptau181: phosphorylated tau 181; Aβ ratio: amyloid-β 42/40 ratio.
Correlation structure among inflammatory, neurodegenerative, and cognitive measures
Correlation analyses between the variables of the study in the overall cohort are presented in Figure 2. In the overall cohort, older age was strongly associated with greater WMH volume (r = 0.52, p < 0.001), higher p-Tau181 (r = 0.44, p < 0.001), and higher NfL (r = 0.57, p < 0.001), and was modestly associated with higher TNF-α (r = 0.21, p < 0.001), IL-10 (r = 0.14, p < 0.001), IL-6 (r = 0.09, p < 0.001), and IL-5 (r = 0.18, p < 0.001). WMH volume was inversely associated with memory (r = −0.20, p < 0.001), executive function (r = −0.22, p < 0.001), processing speed (r = −0.25, p < 0.001), and language (r = −0.20, p < 0.001), and positively correlated with p-Tau181 (r = 0.27, p < 0.001) and NfL (r = 0.40, p < 0.001). Inflammatory biomarkers showed consistent but modest associations with structural and cognitive measures as shown in Figure 3. IL-6 was positively correlated with WMH (r = 0.19, p < 0.001) and inversely correlated with all cognitive domains. TNF-α demonstrated similar patterns while other inflammatory biomarkers patterns varied. The neurodegenerative biomarkers were strongly associated with cognitive performance. Overall, age, WMH volume, and neurodegenerative biomarkers demonstrated the strongest correlations with multidomain cognitive performance, while inflammatory markers showed consistent but smaller magnitude associations. Correlations were similar when stratified by participants that were unimpaired (Supplemental Figure 1), had MCI (Supplemental Figure 2), and had dementia (Supplemental Figure 3) with the r and p values provided in Supplemental Materials 1.
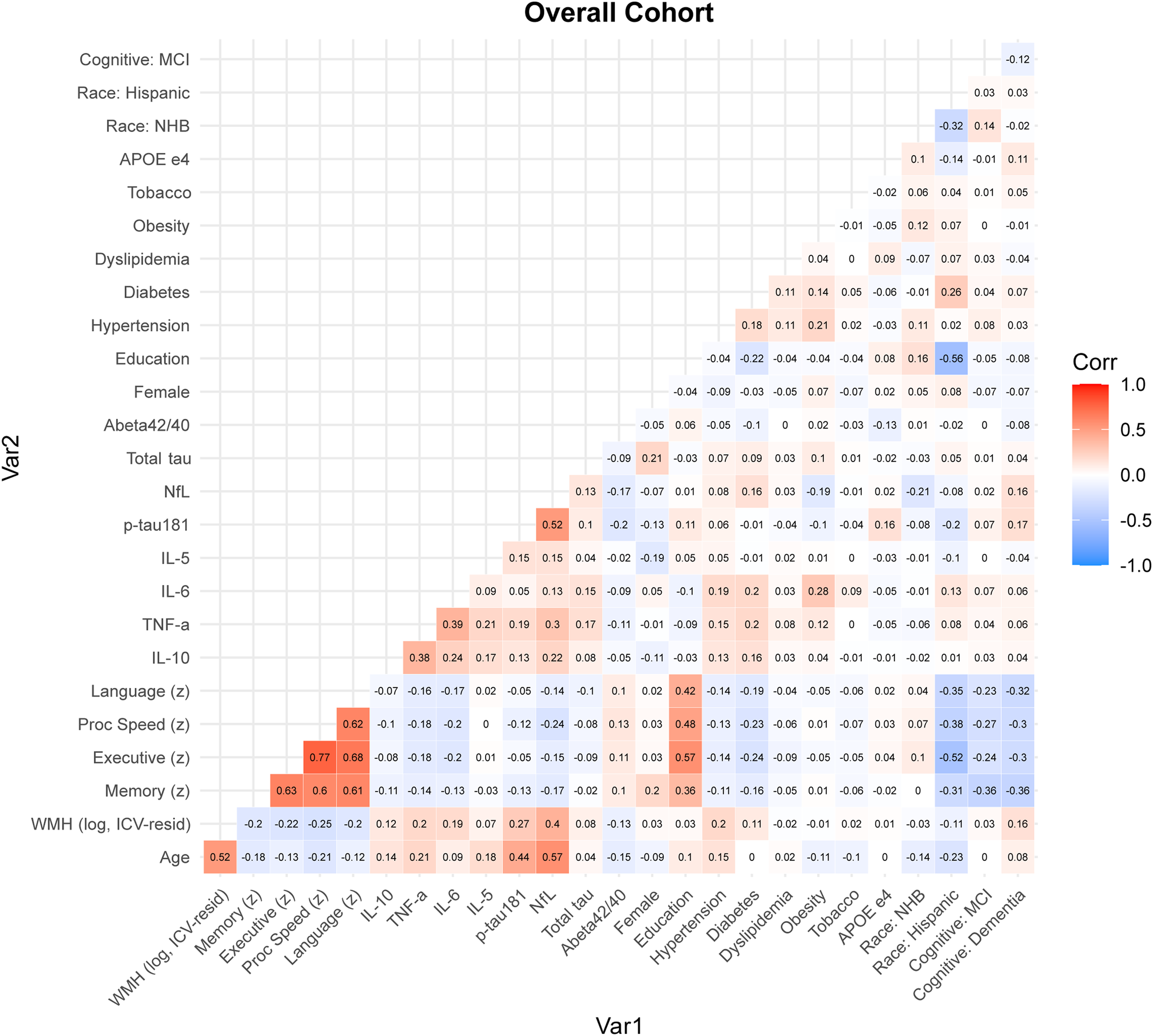
Correlation matrix in the overall cohort. Lower-triangle correlation matrix for the full analytic cohort. Cell values represent pairwise correlation coefficients (r). Correlations were estimated using pairwise complete observations with a mixed approach: Pearson correlation was used when both variables were continuous Pearson-type measures (Age, WMH, memory, executive function, processing speed, and language), and Spearman correlation was used for all other variable pairs. Cognitive status was dummy coded (MCI and dementia, with unimpaired as the reference) and included only in the overall matrix. Race and ethnicity was dummy coded (NHB and Hispanic indicators). Color scale: blue, negative correlations; white, near zero; red, positive correlations (Color figure available online).

Associations of inflammatory biomarkers with WMH and cognitive outcomes from multivariable linear regression models. Forest plots show beta coefficients and 95% confidence intervals for associations of IL-10, TNF-α, IL-6, and IL-5 (log-transformed plasma measures) with (A) WMH volume, (B) memory, (C) executive function, (D) processing speed, and (E) language. For each outcome, estimates were obtained from base linear regression models adjusted for age, sex, education, hypertension, diabetes, dyslipidemia, obesity, tobacco use, race and ethnicity, APOE ε4 status, and the other inflammatory biomarkers (mutually adjusted). Points indicate beta estimates, horizontal bars indicate 95% confidence intervals, and the vertical dashed line indicates the null (beta = 0). Labels report beta (95% CI) and Holm-adjusted p-values; Holm correction was applied globally across all biomarker-outcome tests shown.
Associations of inflammatory biomarkers with WMH and cognitive domains
In the base models adjusted for age, sex, education, vascular risk factors (hypertension, diabetes, dyslipidemia, obesity, tobacco), race and ethnic group, APOE ε4, higher IL-6 was independently associated with greater WMH (β = 0.22; 95% CI: 0.11 to 0.33; unadjusted p < 0.001; Holm-adjusted p = 0.002). No other inflammatory markers were associated with WMH (all Holm-adjusted p = 1.00). For memory, higher IL-6 was associated with lower performance (β = −0.07; 95% CI: −0.13 to −0.01; unadjusted p = 0.015; Holm-adjusted p = 0.276), though this did not survive multiple comparison correction. For executive function, IL-6 was similarly associated with poorer performance (β = −0.06; 95% CI: −0.12 to −0.01; unadjusted p = 0.016; Holm-adjusted p = 0.280), but this association did not remain significant after adjustment. For processing speed, higher IL-6 was associated with worse performance (β = −0.10; 95% CI: −0.16 to −0.03; unadjusted p = 0.003; Holm-adjusted p = 0.048), and this association remained significant after global Holm correction. IL-6 showed a borderline association with language performance (β = −0.06; 95% CI: −0.12 to 0.00; unadjusted p = 0.054). No significant associations were observed for IL-10, TNF-α, or IL-5 with WMH or any cognitive domain (all Holm-adjusted p ≥ 0.31).
Associations of inflammatory biomarkers with WMH and cognitive domains adjusted for neurodegenerative biomarkers
In fully adjusted models further included neurodegenerative biomarkers, higher IL-6 was associated with greater WMH (β = 0.21; 95% CI: 0.10 to 0.32; unadjusted p < 0.001; Holm-adjusted p = 0.005) (Figure 4). Similarly, higher NfL was associated with greater WMH (β = 0.32; 95% CI: 0.18 to 0.46; unadjusted p < 0.001; Holm-adjusted p < 0.001). IL-6 was modestly associated with lower memory (β = −0.07; 95% CI: −0.13 to −0.01; unadjusted p = 0.024; Holm-adjusted p = 0.715), but this did not survive correction. IL-6 showed a small negative association (β = −0.06; 95% CI: −0.11 to −0.01; unadjusted p = 0.030) that did not remain significant after correction with executive function. IL-6 was also associated with lower processing speed (β = −0.09; 95% CI: −0.15 to −0.03; unadjusted p = 0.005; Holm-adjusted p = 0.157). Higher levels of NfL and p-Tau 181 were also associated with lower cognitive performance although not always significant after the Holm correction.
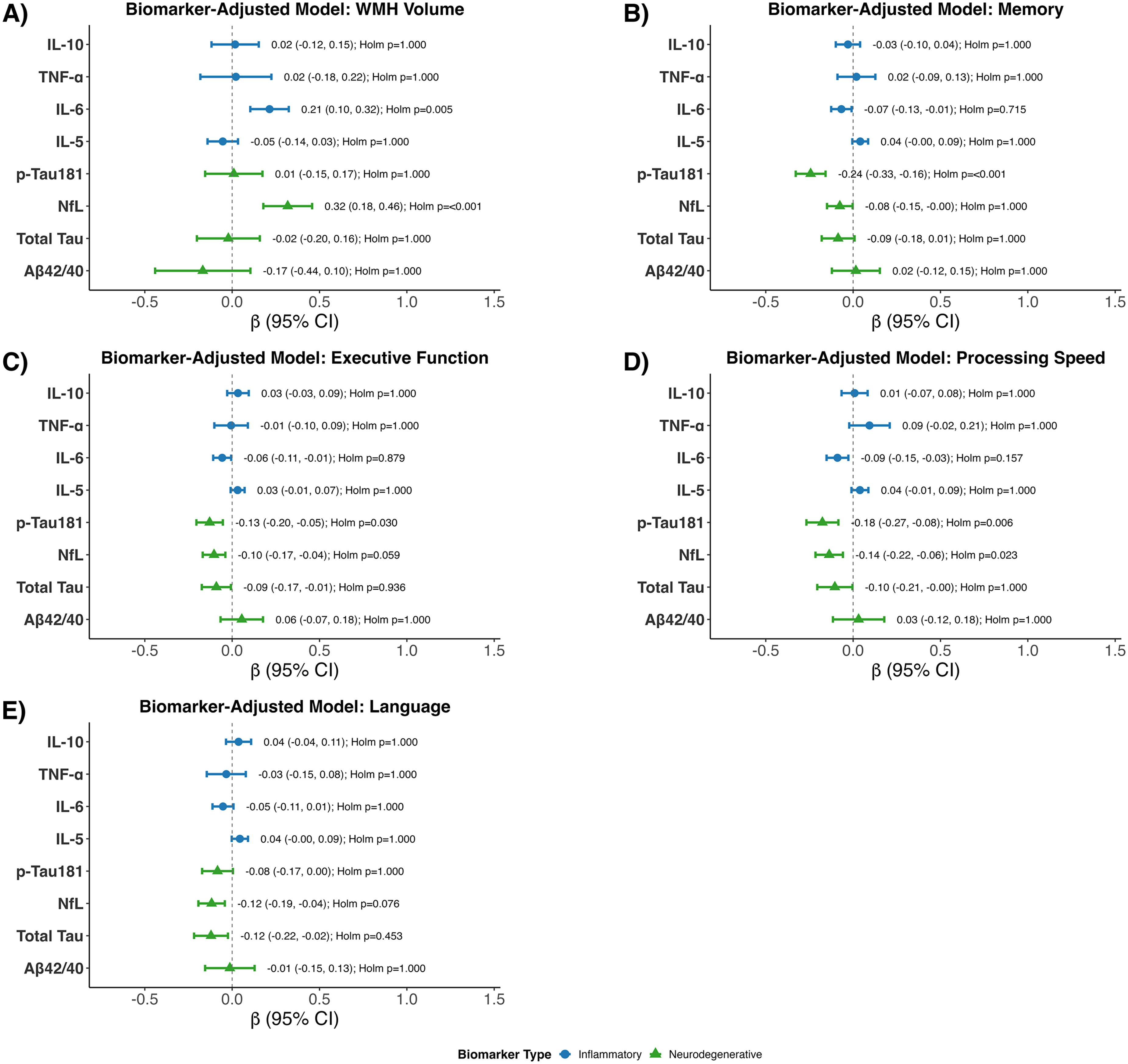
Associations of plasma biomarkers with WMH and cognitive outcomes in fully adjusted multivariable linear regression models. Forest plots show beta coefficients and 95% confidence intervals for associations of plasma biomarkers with (A) WMH volume, (B) memory, (C) executive function, (D) processing speed, and € language. Full linear regression models were adjusted for age, sex, education, hypertension, diabetes, dyslipidemia, obesity, tobacco use, race and ethnic group, and APOE ε4, and included all log transformed biomarkers simultaneously: IL-10, TNF-α, IL-6, IL-5, p-tau181, NfL, total tau, and Aβ42/40 ratio. Points indicate beta estimates, horizontal bars indicate 95% confidence intervals, and the vertical dashed line indicates the null (beta = 0). Labels report beta (95% CI) and Holm-adjusted p values, with Holm correction applied globally across all biomarker-outcome tests shown.
Associations of inflammatory biomarkers with WMH and cognitive domains adjusted for neurodegenerative biomarkers stratified by cognitive status
Among cognitively unimpaired individuals, higher IL-6 was independently associated with greater WMH volume (β = 0.20; 95% CI: 0.07 to 0.33; p < 0.01), whereas IL-6 was not associated with any cognitive domains nor were any of the neurodegenerative biomarkers (Supplemental Table 2). These findings were mostly consistent in only participants with MCI (Supplemental Table 3) or those with dementia (Supplemental Table 4).
Associations of inflammatory biomarkers with WMH and cognitive domains adjusted for neurodegenerative biomarkers controlling for cognitive status
In the overall cohort, when adjusting for clinical cognitive status, the previously observed biomarker associations with cognition were no longer significant (Supplemental Table 5).
WMH mediate the association between IL-6 and cognitive domains
Causal mediation analyses were conducted using nonparametric bootstrapping with 5000 simulations among 1650 complete cases to evaluate whether WMH mediated the association between IL-6 and cognitive performance (Figure 5). For memory, IL-6 was significantly associated with greater WMH (Path a: β = 0.21; 95% CI: 0.10 to 0.32; p < 0.001), and greater WMH was associated with worse memory (Path b: β = −0.07; 95% CI: −0.10 to −0.05; p < 0.001). The total effect of IL-6 on memory was significant (Path c: β = −0.07; 95% CI: −0.13 to −0.01; p = 0.031). The indirect effect was significant (ACME: β = −0.02; 95% CI: −0.03 to −0.01; p < 0.001), with approximately 22.9% of the total effect mediated by WMH (95% CI: 0.07 to 1.14; p = 0.031). For executive function, the indirect effect between IL-6 and executive function was significant (ACME: β = −0.02; 95% CI: −0.03 to −0.01; p < 0.001), although the proportion mediated was not statistically significant (37.9%; 95% CI: −1.05 to 2.23; p = 0.070). The indirect effect between IL-6 and processing speed was significant (ACME: β = −0.02; 95% CI: −0.03 to −0.01; p < 0.001), with 19.2% of the association mediated by WMH (95% CI: 0.08 to 0.63; p = 0.004). For the language domain, the indirect effect was significant (ACME: β = −0.02; 95% CI: −0.03 to −0.01; p < 0.001), though the proportion mediated was not statistically significant (27.3%; 95% CI: −1.15 to 2.31; p = 0.109).
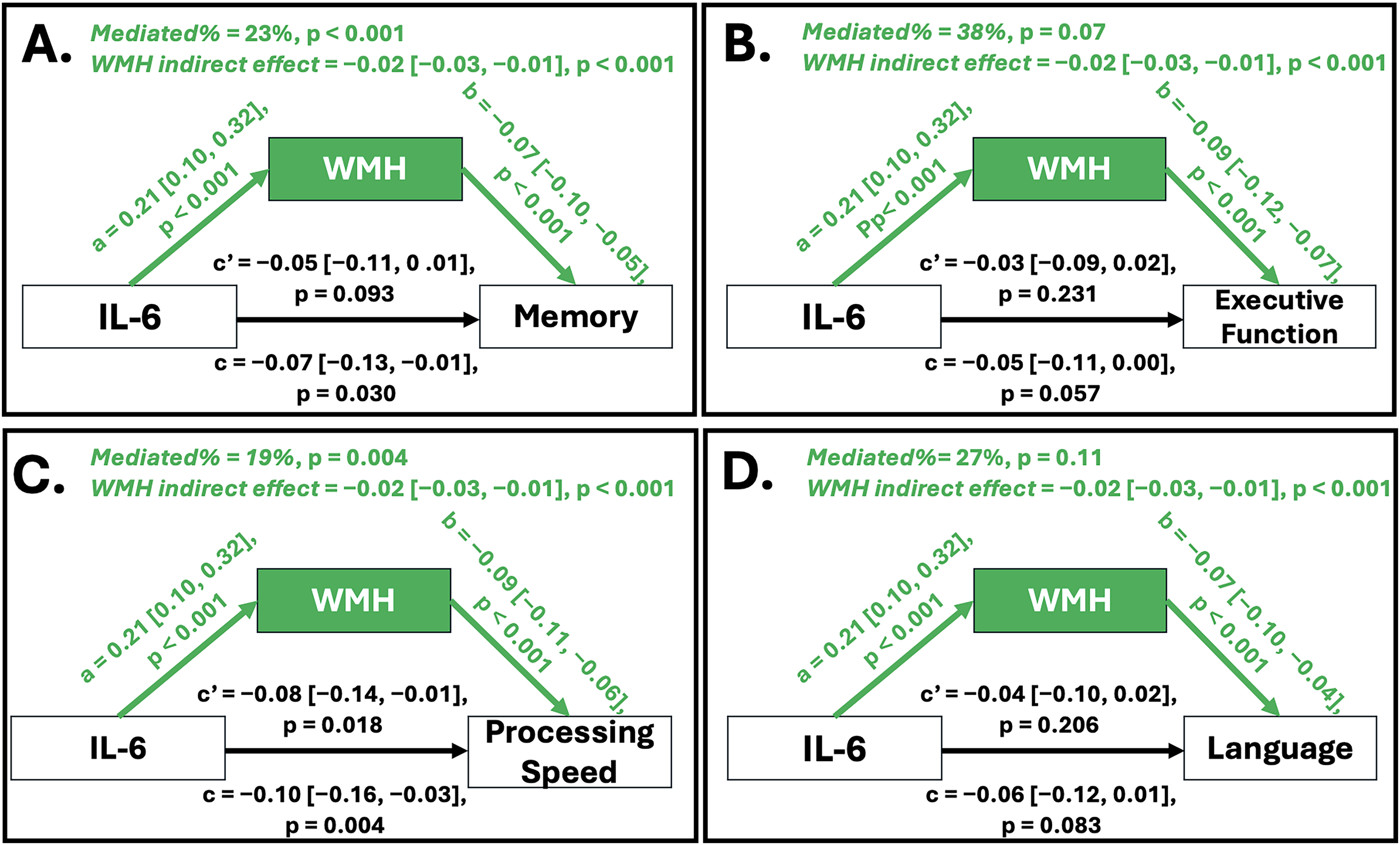
Mediation of the association between IL-6 and cognitive performance by WMH volume. Panels A–D display causal mediation models evaluating whether ICV-adjusted white matter hyperintensity (WMH) volume mediates the association between plasma IL-6 and cognitive performance across domains: (A) Memory, (B) Executive Function, (C) Processing Speed, and (D) Language. Path a represents the association between IL-6 and WMH; Path b represents the association between WMH and cognition; Path c represents the total effect of IL-6 on cognition; and Path c′ represents the direct effect after accounting for WMH. Indirect effects (ACME) and proportion mediated are shown at the top of each panel. Estimates are unstandardized β coefficients with 95% confidence intervals and p-values derived from 5000 bootstrap simulations.
Discussion
This study aimed to evaluate whether peripheral inflammatory biomarkers are associated with WMH volume and cognition independent of AT(N) plasma biomarkers. In a large, racially and ethnically diverse cohort of older adults, we found that higher plasma IL-6 levels were consistently associated with greater WMH volume and poorer performance in memory, executive function, and processing speed. Importantly, these associations remained significant even after adjusting for AD biomarkers (p-Tau181, NfL, total tau, and Aβ42/40) suggesting that IL-6 contributes to cerebrovascular and cognitive impairment via pathways that are at least partially independent of classical AD pathology. NfL also emerged as a robust predictor of WMH volume and cognitive deficits, particularly in executive function, processing speed, and language, reinforcing its role as a sensitive marker of neuroaxonal damage. In contrast, other inflammatory markers (IL-10, TNF-α, IL-5) and most AT(N) biomarkers showed no significant associations with WMH volume or cognitive outcomes after covariate adjustment in this selective subsample of HABS-HD participants.
Our results align with a large body of evidence implicating IL-6 in cerebral small vessel disease and neurovascular injury. For example, in a longitudinal study by Nadkarni and colleagues, higher sustained IL-6 exposure was significantly associated with increased WMH volume, independent of APOE ε4 status. 5 Similarly, community-based multiethnic cohorts have shown that elevated IL-6 and other inflammatory markers are associated with cerebrovascular disease burden, including WMH, infarcts, and microbleeds.6,30 At the mechanistic levels, these findings suggest IL-6 may reflect changes in endothelial dysfunction, increased blood-brain barrier permeability, and exacerbate microglial activation, all of which contribute to small vessel disease and subsequent cognitive decline.
Notably, our study extends these findings by demonstrating that IL-6 remains independently associated with WMH volume and cognition even after accounting for AD-related plasma biomarkers, but these findings did not survive multiple comparisons corrections. Regardless, our findings support the hypothesis of a vascular–inflammatory pathway distinct from amyloid and tau pathology that contribute to cognitive impairment which has been highlighted in other pathological phenotypes using autopsied data.31,32 By comparison, other studies have reported weaker or null associations between IL-6 and WMH progression within the Alzheimer's Disease Neuroimaging Initiative cohort. 33 These discrepancies may reflect differences in sample size, population diversity, or inflammatory dynamics over time, underscoring the value of large, community-based, multiethnic cohorts like the HABS-HD.
Our findings that higher IL-6 levels are associated with worse performance across multiple cognitive domains align with previous studies implicating systemic inflammation in cognitive impairment. Higher IL-6 has been linked to lower Mini-Mental State Examination Scores, 34 faster cognitive decline 8 and greater dementia risk.7,9 In parallel, our observation that NfL was significantly associated with WMH volume and cognition is consistent with its role as a sensitive marker of axonal injury and neurodegeneration. Prior work in the Alzheimer's Disease Neuroimaging Initiative has shown that NfL predicts cognitive decline years before dementia onset,13,35 and is elevated even among cognitively normal individuals with abnormal amyloid profiles. 12 Our results extend this literature by showing that NfL associations persist even when adjusting for IL-6, highlighting the distinct yet complementary contributions of vascular-inflammatory and neuroaxonal injury pathways to cognitive aging.
Our mediation analyses suggest that WMH volume represents an important vascular pathway linking systemic inflammation to cognitive performance. Higher IL-6 levels were consistently associated with greater WMH volume, supporting the hypothesis that chronic inflammatory signaling may contribute to cerebral small vessel injury through mechanisms such as endothelial dysfunction, blood-brain barrier disruption, and microvascular remodeling. 36 In turn, greater WMH volume was associated with worse performance across cognitive domains,37–39 underscoring the structural relevance of small vessel disease to brain function. The indirect effects observed across domains indicate that part of the association between IL-6 and cognition operates through white matter injury and possible other adjacent structural brain regions. 40 This pathway was most robust for memory and processing speed suggesting that inflammatory contributions to cognitive decline may be particularly pronounced in white matter–dependent and frontal-subcortical processes. 41 Although mediation effects were also present for executive function and language, the weaker total effects in these domains imply that additional neurobiological pathways potentially including neurodegenerative or cortical processes may also contribute. The likely underlying mechanisms between IL-6, WMH, and cognition included but are not limited to blood-brain barrier dysfunction, 42 hippocampal atrophy, 43 deposition of amyloid and tau, 44 and molecular pathways such as suppressor of cytokine signaling 3 SOC3. 45 Collectively, these findings support a vascular–inflammatory model in which systemic inflammation contributes to small vessel disease, which in turn adversely affects multidomain cognitive performance, highlighting the potential importance of targeting both inflammatory and cerebrovascular mechanisms in efforts to preserve cognitive health.
From a statistical perspective, the associations between IL-6 and cognitive domains were attenuated after multiple comparison correction; however, there are several factors support the relevance of these findings. First, the direction and magnitude of associations were consistent across domains, suggesting a coherent underlying relationship. Second, IL-6 remained robustly associated with WMH, and mediation analyses demonstrated that WMH partially accounted for the association between IL-6 and cognition, supporting a vascular–inflammatory mechanism. Third, the use of Holm correction, while appropriate for controlling family-wise error, is conservative in the context of correlated outcomes such as cognitive domains, potentially reducing statistical power to detect modest but meaningful effects. Taken together, these findings suggest that IL-6 contributes to cognitive impairment through cerebrovascular pathways, even when associations do not meet stringent corrected thresholds.
Although our study overcomes several gaps in the literature, there are some limitations including this being a cross-sectional study. Longitudinal studies are needed to determine whether IL-6 predicts future WMH accumulation or cognitive decline. Second, while we adjusted for most vascular and demographic covariates, there could be a residual confounding effect of unmeasured lifestyle risk factors (such as diet, physical activity, etc.). Third, we only used AT(N) plasma biomarkers in our analyses, which provide indirect measures of central nervous system pathology. Integrating cerebral spinal fluid-based or positron emission tomography-based A neuroimaging biomarkers may yield complementary insights.
In summary, this study provides compelling evidence that IL-6 is independently associated with WMH and cognitive performance across multiple domains, even after accounting for AT(N) plasma biomarkers. These findings support a vascular–inflammatory pathway in cognitive aging that is distinct from classical AD pathology. Leveraging a large, racially and ethnically diverse cohort, our results underscore the importance of considering inflammatory processes alongside the ATN(N) framework of plasma biomarkers in understanding the heterogeneity of dementia risk. Future longitudinal and mechanistic studies are warranted to clarify how IL-6 and related pathways contribute to vascular brain injury and cognitive decline, and whether they represent viable targets for intervention. Ultimately, these findings are the first to confirm that IL-6 retains significance across multiple age-related outcomes even after controlling for AT(N) biomarkers suggests a vascular-inflammatory pathway distinct from AD pathology. From a mechanistic perspective, our findings suggest that WMH partially mediate the relationship between IL-6 and select cognitive domains, highlighting the potential value of combinatorial therapeutic approaches that target both systemic inflammation and cerebrovascular pathology.
Supplemental Material
sj-xlsx-1-alz-10.1177_13872877261458367 - Supplemental material for Interleukin-6 is associated with white matter hyperintensities and cognition independent of Alzheimer's disease plasma biomarkers—HABS-HD
Supplemental material, sj-xlsx-1-alz-10.1177_13872877261458367 for Interleukin-6 is associated with white matter hyperintensities and cognition independent of Alzheimer's disease plasma biomarkers—HABS-HD by Anhiti Dharmapuri, Soumilee Chaudhuri, Joey Annette Contreras, Cellas A. Hayes and in Journal of Alzheimer's Disease
Supplemental Material
sj-docx-2-alz-10.1177_13872877261458367 - Supplemental material for Interleukin-6 is associated with white matter hyperintensities and cognition independent of Alzheimer's disease plasma biomarkers—HABS-HD
Supplemental material, sj-docx-2-alz-10.1177_13872877261458367 for Interleukin-6 is associated with white matter hyperintensities and cognition independent of Alzheimer's disease plasma biomarkers—HABS-HD by Anhiti Dharmapuri, Soumilee Chaudhuri, Joey Annette Contreras, Cellas A. Hayes and in Journal of Alzheimer's Disease
Footnotes
Acknowledgements
We are thankful to the participants of the HABS-HD study for sharing their time and contributing to the advancement of this research.
Ethical considerations
All procedures are reviewed and approved by the North Texas Regional Institutional Review Board and comply with the ethical guidelines of the 1975 Declaration of Helsinki.
Consent to participate
All HABS-HD participants provided informed consent to participate in the study.
Consent for publication
Not applicable
Author contribution(s)
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Research reported in this publication was supported by the National Institute on Aging of the National Institutes of Health under Award Numbers R01AG054073 and R01AG058533, P41EB015922 and U19AG078109. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health. CAH is funded by Burroughs Wellcome Fund Postdoctoral Enrichment Program (PDEP) 1267001, and the HABS-HD Health Enhancement Scientific Program (HESP) U19AG078109.
Declaration of conflicting interests
The authors declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: Cellas A. Hayes is an Editorial Board Member of this journal but was not involved in the peer-review process of this article nor had access to any information regarding its peer-review.
The remaining authors declare no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data availability statement
Supplemental material
Supplemental material for this article is available online.