
Abstract
Aims:
This study aims to investigate whether melatonin (MLT) exerts protective effects against cognitive impairment following exertional heat stroke (EHS) by modulating ferroportin (Fpn) to alleviate hippocampal ferroptosis and neuroinflammation.
Results:
Following EHS, genes such as Mt1, Mt2, and Trf were notably upregulated in the hippocampal tissue, whereas genes such as Slc40a1 (encoding Fpn 1) and Il33 were downregulated. Kyoto Encyclopedia of Genes and Genomes analysis implicated ferroptosis as a dominant. MLT significantly ameliorated learning and memory deficits observed in EHS mice. This treatment also modulated ferroptosis markers, such as Fpn, xCT, ferritin H, and glutathione peroxidase 4, reduced hippocampal iron overload, and decreased the secretion of proinflammatory cytokines interleukin (IL)-6 and tumor necrosis factor-α (TNF-α). Furthermore, MLT treatment reduced oxidative stress and lipid peroxidation and mitigated mitochondrial and neuronal damage in the hippocampal tissue. Strikingly, conditional Fpn knockout abolished MLT’s benefits: Fpn-cKO + MLT mice showed persistent iron accumulation, elevated IL-6 and TNF-α, and failed cognitive recovery.
Innovation:
Our study reveals that MLT prevents EHS-induced neurodegeneration by enhancing Fpn-dependent iron efflux, a mechanism that concurrently resolves hippocampal iron overload, suppresses ferroptosis, and dampens neuroinflammation.
Conclusion:
Our findings indicate that MLT mitigates EHS-related cognitive impairment by restoring hippocampal iron homeostasis and suppressing neuroinflammation, primarily through Fpn-dependent mechanisms. Antioxid. Redox Signal. 43, 509–527.
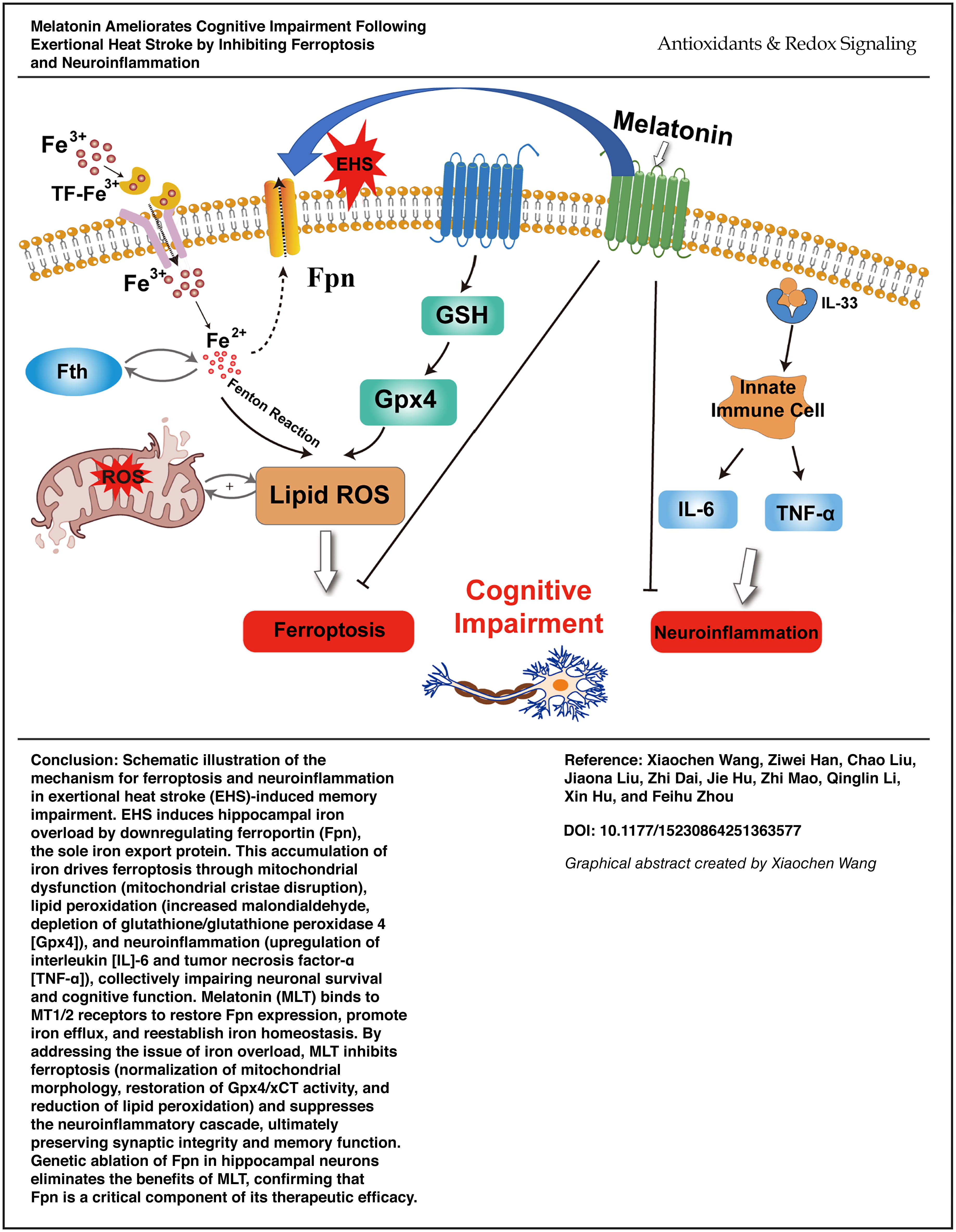
Introduction
Exertional heat stroke (EHS) is a life-threatening condition induced by intense activity in hot environments, which frequently leads to permanent neurological sequelae (Barletta et al., 2025). Up to 76% of survivors experience long-term cognitive dysfunction, including learning and memory deficits, imposing profound burdens on individuals and society (Leon and Bouchama, 2015; Bouchama et al., 2022). While systemic inflammation triggered by heat stress-induced endotoxemia is recognized as a key driver of central nervous system (CNS) damage (González et al., 2014), emerging evidence implicates ferroptosis—an iron-dependent cell death pathway marked by lipid peroxidation and glutathione peroxidase 4 (Gpx4) suppression—as a critical mediator of neuronal degeneration (Alim et al., 2019; Streit et al., 2025; Weiland et al., 2019). Notably, heat stress disrupts hippocampal iron homeostasis, causing iron overload that exacerbates oxidative damage and cognitive impairment (Liu et al., 2019), yet the molecular regulatory mechanisms linking iron dysmetabolism and ferroptosis to cognitive dysfunction resulting from EHS remain undefined.
Central to iron homeostasis is ferroportin (Fpn), the sole known cellular iron exporter encoded by Slc40a1. By mediating iron efflux, Fpn prevents cytotoxic iron accumulation and the subsequent ferroptosis (Zhang et al., 2018). In neurodegenerative contexts, Fpn deficiency correlates with hippocampal iron overload, mitochondrial dysfunction, and neuroinflammation (Helgudottir et al., 2019; Junceda et al., 2024), suggesting its pivotal role in neuroprotection. However, whether Fpn participates in EHS-related cognitive decline, particularly through cross talk between iron metabolism and inflammatory cascades, is unexplored.
Melatonin (MLT), a pleiotropic hormone with potent anti-inflammatory and antioxidant properties, shows promise in mitigating CNS disorders by modulating MT1/2 receptor signaling (Xu et al., 2019; Jilg et al., 2019). Recent studies have intriguingly demonstrated that MLT regulates iron metabolism by upregulating Fpn, which helps alleviate iron overload in specific diseases. (Park et al., 2022; Zhao et al., 2022). This dual ability to suppress both oxidative stress and neuroinflammation positions MLT as a potential therapeutic for EHS-induced brain injury. Nevertheless, no studies have investigated whether MLT rescues cognitive deficits in EHS via Fpn-dependent mechanisms or how this intersects with ferroptosis inhibition.
In this study, we hypothesize that MLT ameliorates EHS-induced cognitive impairment by enhancing Fpn-mediated iron efflux, thereby concurrently attenuating hippocampal ferroptosis and neuroinflammation. Our study combines transcriptomic, behavioral, and molecular approaches to unravel this mechanism, offering novel insights into therapeutic strategies for EHS-related neurodegeneration.
Innovation
This study utilized transcriptomic methods to identify differentially expressed genes in the hippocampus following EHS and then performed functional enrichment analysis. The analysis suggested that ferroptosis could play a role in CNS damage after EHS. Subsequently, this study confirmed that ferroptosis and inflammatory responses occurred in the CNS of mice after EHS. The MT1/2 agonist MLT was administered to mice post-EHS, revealing that MLT mitigated EHS-induced cognitive impairment by suppressing ferroptosis and reducing CNS inflammation. These findings suggest that MLT, a commonly used clinical drug, may ameliorate EHS-related cognitive deficits. This study provides a preliminary basis for further investigating the specific mechanisms through which MLT alleviates CNS damage and cognitive dysfunction following EHS.
Results
EHS induces cognitive impairment
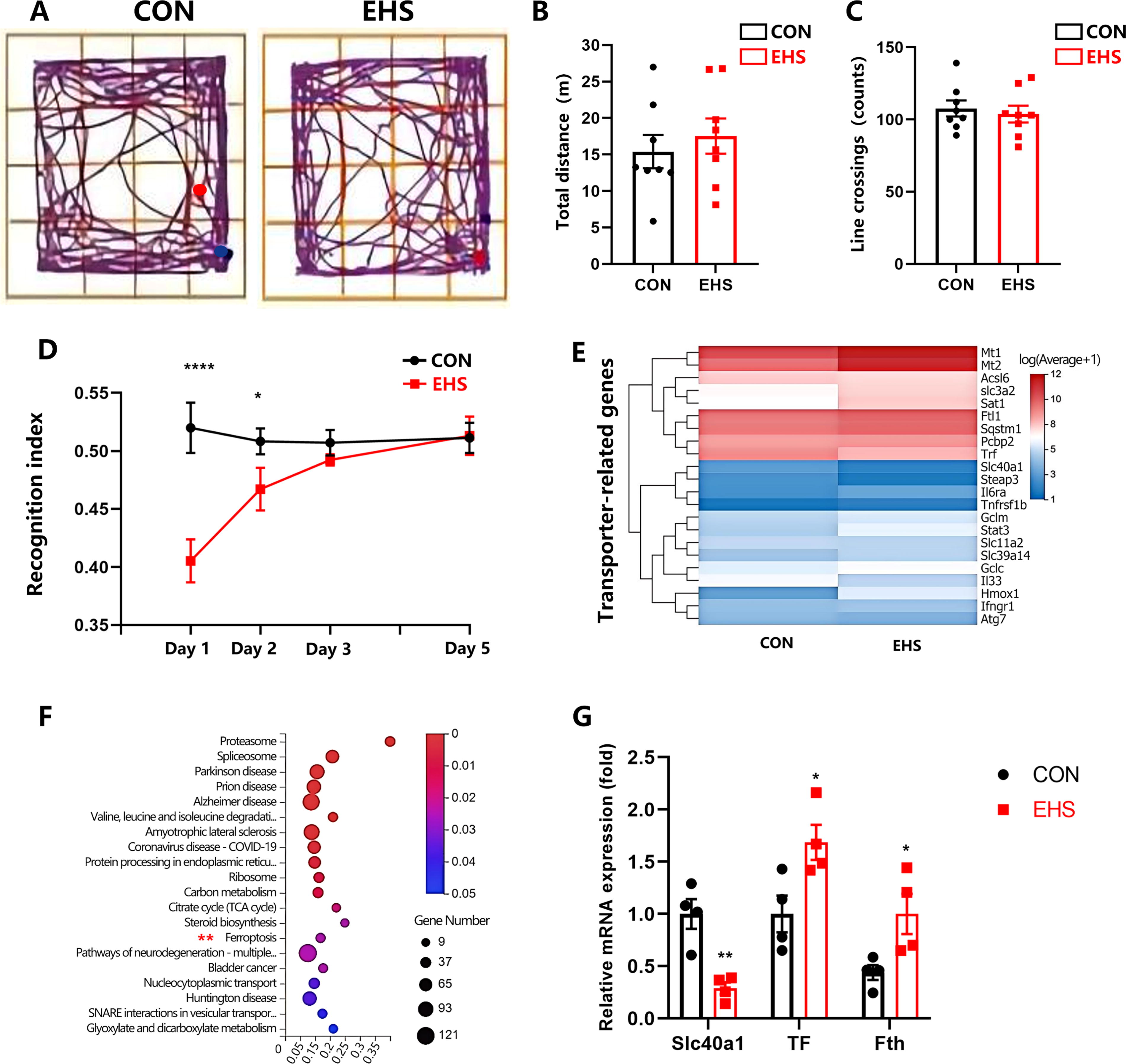
The study found no significant differences in open-field (OF) test results between the two groups of mice 24 h post-EHS exposure, as indicated by the total distance and number of line crossings (representative trajectories shown in Fig. 1A; total distance and number of crossed lines, p = 0.54 and 0.64, respectively; Fig. 1B, C). These results suggest that EHS does not affect spontaneous activity, ruling it out as a contributing factor to subsequent behavioral changes. The novel object recognition (NOR) test revealed a significant decline in recognition memory 24 h postexposure (p < 0.001; Fig. 1D). Transcriptome analysis showed upregulation of Mt1 and Mt2 genes and downregulation of the Fpn gene, potentially linked to cognitive deficits post-EHS. Inflammatory genes such as Il 6ra, Tnfrsf 1b, and Il 33 were also implicated (Fig. 1E). Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis highlighted ferroptosis in the hippocampus after EHS (Fig. 1F). qRT-PCR confirmed decreased Fpn mRNA and increased time spent exploring familiar objects (TF) and ferritin H (Fth) mRNA levels (Fig. 1G), indicating the involvement of the ferroptosis pathway in EHS-induced cognitive impairment.
MLT alleviated EHS-induced neuronal damage in the hippocampus
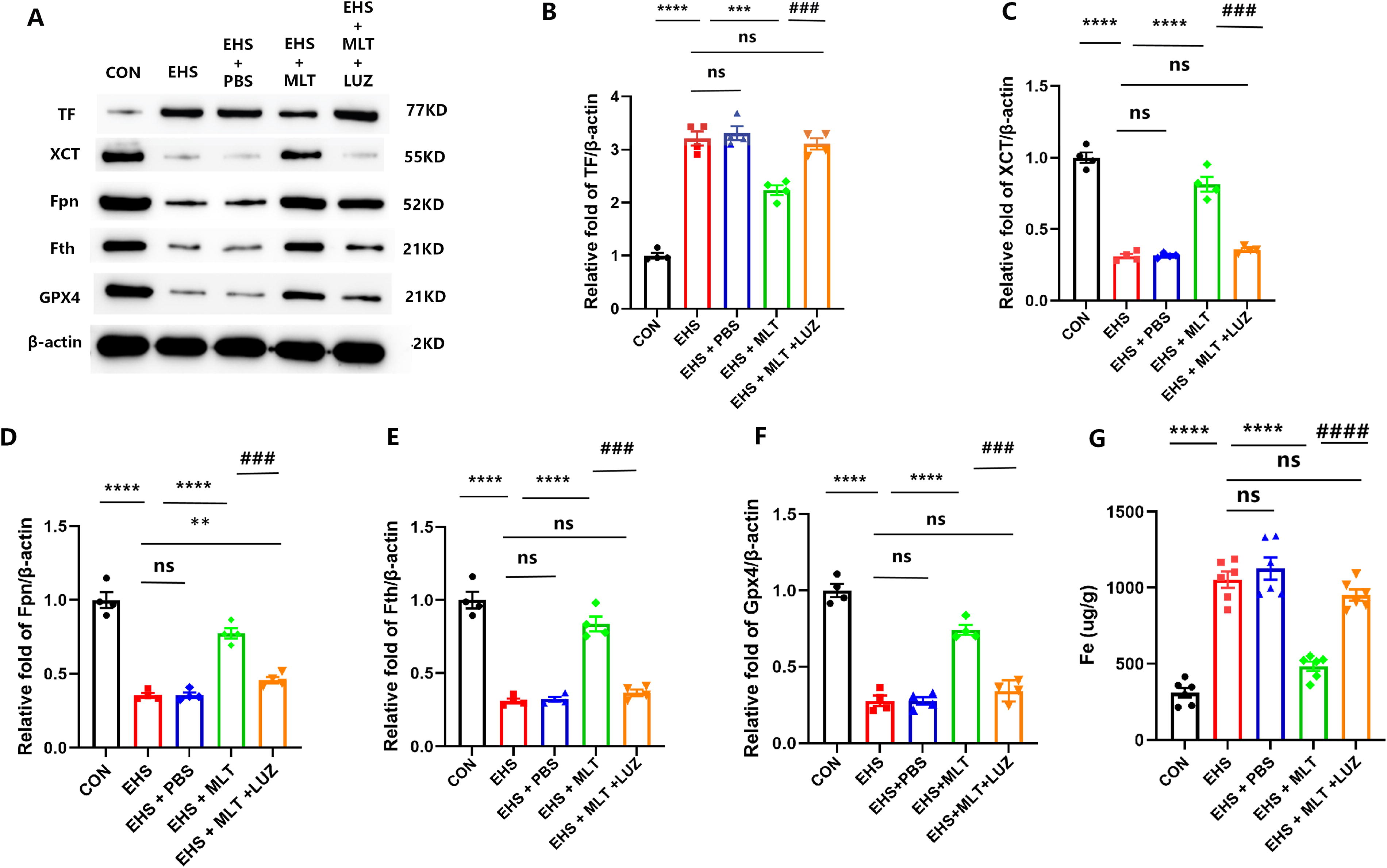
Nissl staining is commonly used to visualize Nissl bodies within neurons, observe cell structures, and assess neuronal damage. As shown in Figure 2A, the cytoplasm of damaged neurons shrunk. The EHS and EHS + phosphate-buffered saline (PBS) group exhibited a significant reduction in the number of Nissl bodies compared with the control (CON) group (p < 0.05; Fig. 2C), whereas the EHS + MLT group showed an increase in Nissl bodies compared with the EHS and EHS + PBS group (p < 0.05; Fig. 2C). This protection exhibited receptor-dependency, as coadministration of MLT receptor antagonist LUZ significantly attenuated the neuroprotective effects of MLT (p < 0.05; Fig. 2C). A nonsignificant difference was observed between the EHS and EHS + PBS groups (p > 0.05, Fig. 2C). TUNEL staining (Fig. 2B) illustrates neuronal death in the hippocampus. Compared with the CON group, the TUNEL-positive cells were significantly higher in the EHS and EHS + PBS groups (p < 0.0001, Fig. 2D); however, in the EHS and EHS + PBS groups, the number of TUNEL-positive cells was significantly lower in the EHS + MLT group (p < 0.0001, Fig. 2D). A nonsignificant difference was observed between the EHS and EHS + PBS groups (p > 0.05, Fig. 2D). LUZ appeared to counteract the neuroprotective effects of MLT. TUNEL staining showed that the apoptotic cell count in EHS + MLT + LUZ was similar to EHS (p = 0.64) but higher than EHS + MLT (p < 0.001, Fig. 2D).

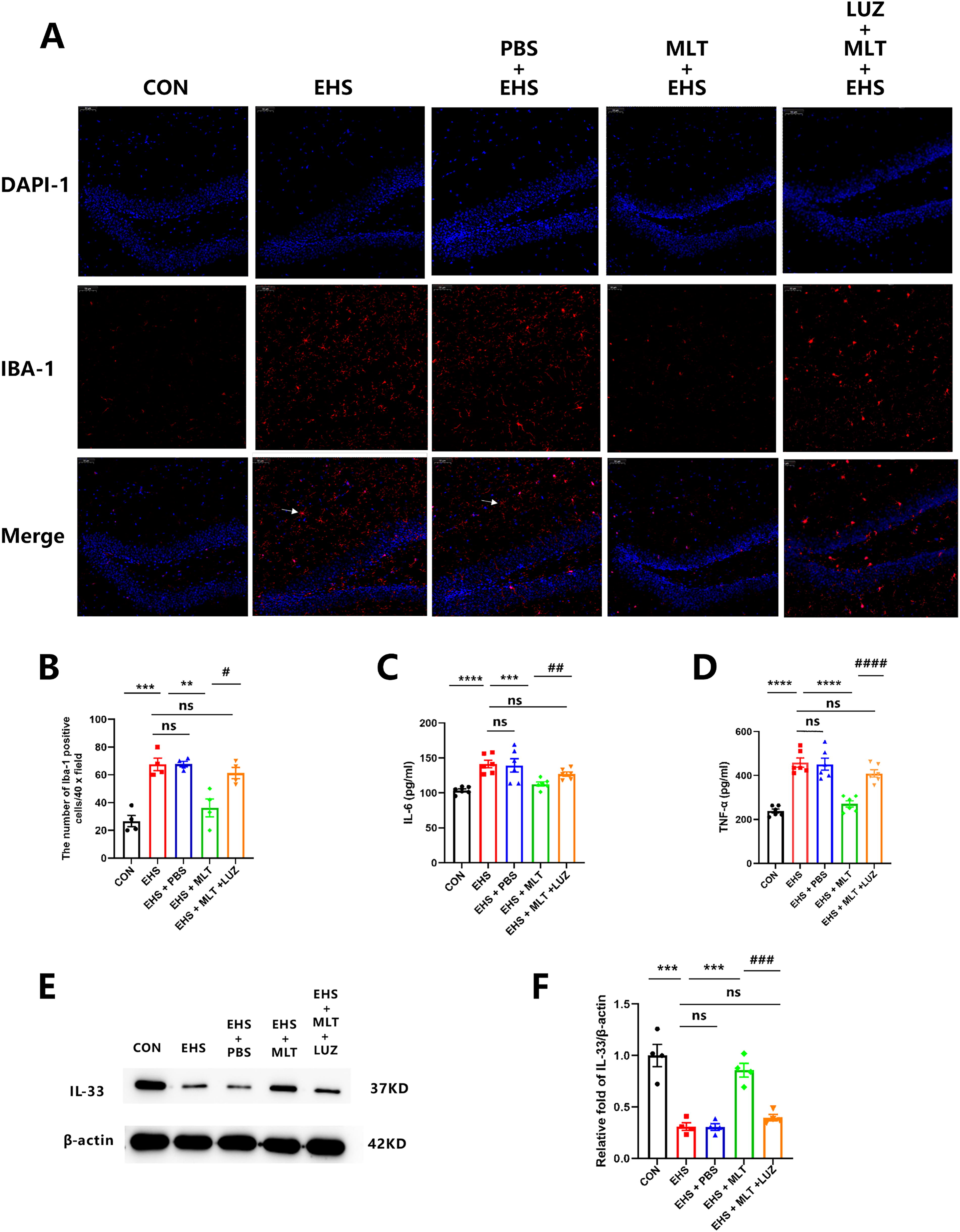
MLT inhibited microglial activation and mitigated the EHS-triggered inflammatory response
We examined the concentrations of various inflammatory cytokines and proteins (such as IL-6, TNF-α, IL-33, and ionized calcium-binding adapter molecule 1 [Iba-1]) in the hippocampus. This brain region is primarily affected by neuroinflammation following an injury or inflammation.
Immunofluorescence revealed a significant increase in microglial activation (Fig. 3A). The number of activated microglia was notably higher in the EHS and EHS + PBS group than in the CON group. However, the number of activated microglia was not significantly different between the EHS and EHS + PBS groups. Conversely, microglial activation was significantly reduced in the EHS + MLT group compared with the EHS and EHS + PBS group (p < 0.001; Fig. 3B). LUZ cotreatment counteracted this suppression, resulting in significantly higher microglial activation compared with MLT monotherapy (p < 0.05; Fig. 3B). Moreover, enzyme-linked immunosorbent assay (ELISA) and Western blot (WB) demonstrated a marked increase in IL-6 (p < 0.001; Fig. 3C) and TNF-α (p < 0.001; Fig. 3D) levels and reduced expression of anti-inflammatory cytokine IL-33 (p < 0.001; Fig. 3E) in the hippocampus of the EHS and EHS + PBS group compared with the CON group. MLT treatment significantly mitigated the increases in IL-6 (p < 0.05; Fig. 3C) and TNF-α (p < 0.01; Fig. 3D) and markedly increased IL-33 levels (p < 0.01; Fig. 3E) in the EHS + MLT group. Importantly, LUZ restored the inflammatory profile to levels comparable with the EHS baseline (IL-6: p = 0.08; TNF-α: p = 0.10) and significantly decreased IL-33 expression (p < 0.001; Fig. 3C–E compared with the MLT treatment group). There were no significant differences in the levels of IL-6, TNF-α, and IL-33 among the EHS, EHS + PBS, and EHS + MLT + LUZ groups.
MLT attenuated oxidative stress and lipid peroxidation and improved mitochondrial injury in hippocampal neurons
Lipid peroxide accumulation and lipid oxidative stress are the fundamental stages of ferroptosis induction (Yang et al., 2016; Stockwell et al., 2017). Our findings revealed a significant increase in malondialdehyde (MDA) (p < 0.001; Fig. 4C) levels in the EHS and EHS + PBS group compared with the CON group, while the levels of superoxide dismutase (SOD) and glutathione (GSH) (p < 0.0001 and p < 0.01, respectively; Fig. 4D, E) were notably decreased. Also, a nonsignificant difference was observed between the EHS and EHS + PBS group. This indicated a higher level of oxidative stress in the EHS and EHS + PBS group. However, treatment with MLT counteracted these changes, suggesting its potential therapeutic effect in mitigating oxidative stress-related damage to the hippocampus.
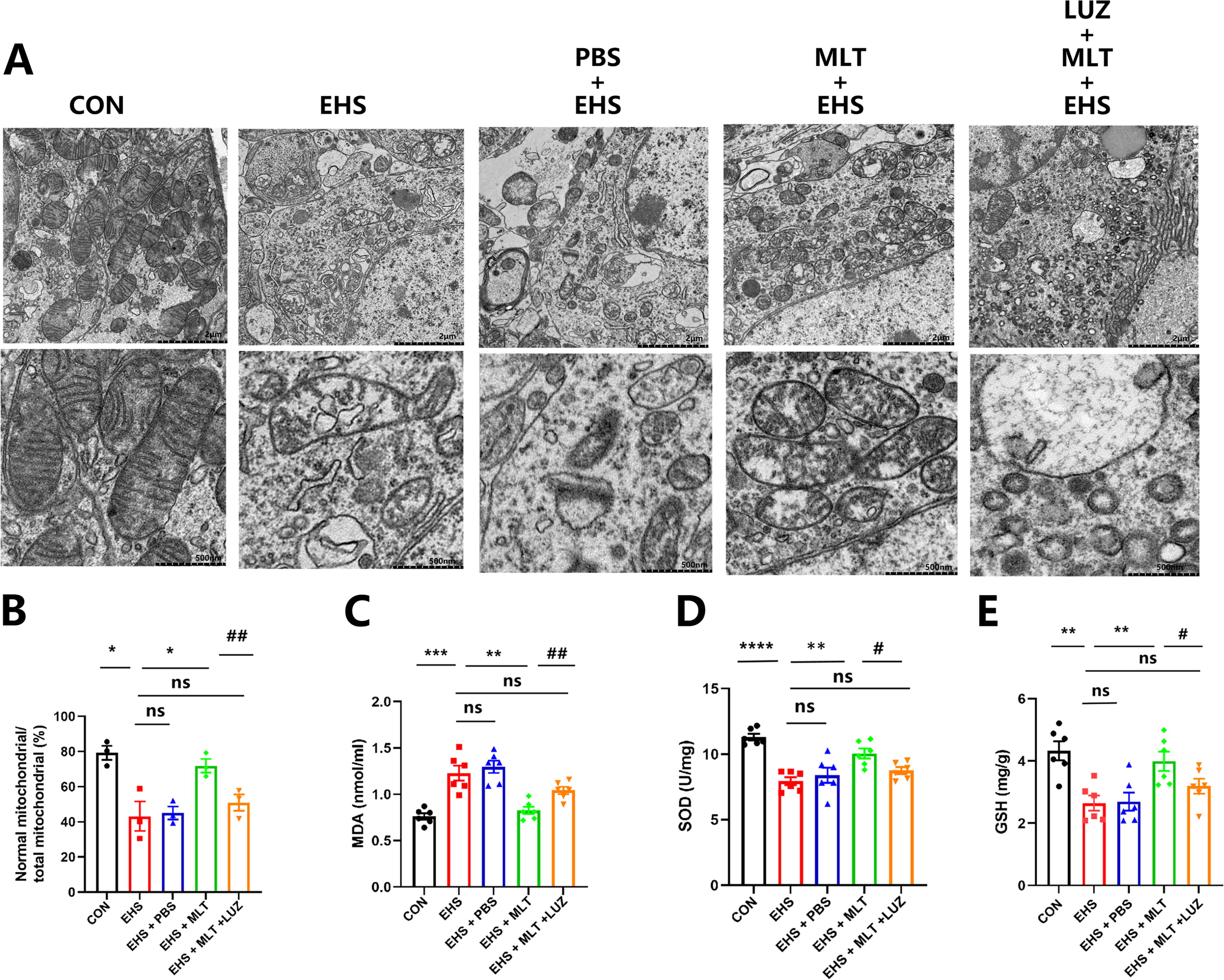
Figure 4A illustrates notable alterations in the mitochondria associated with ferroptosis in the hippocampus. Conversely, mitochondria in the MLT-treated EHS group appeared relatively unaffected, with clear cristae. As depicted in Figure 4B, the proportion of healthy mitochondria was substantially lower in the EHS and EHS + PBS group than in the CON group (p < 0.01; Fig. 4B). Treatment with MLT led to a significant increase in the number of healthy mitochondria (p < 0.01; Fig. 4E). LUZ cotreatment significantly attenuated MLT’s protective effects, with MDA levels returning to those observed in the EHS group (p < 0.01 vs. MLT; Fig. 4B). SOD and GSH concentrations in LUZ-treated group showed no significant difference compared with EHS controls (p > 0.05; Fig. 4C, D). Mitochondrial integrity was markedly compromised in LUZ cotreatment group versus MLT monotherapy (p < 0.01; Fig. 4A, B).
MLT alleviates ferroptosis in the mouse hippocampus after EHS
Abnormal iron metabolism and lipid peroxidation are key processes in ferroptosis (Du et al., 2019). The expression levels of ferroptosis-related proteins were analyzed using WB (Fig. 5A). MLT treatment significantly attenuated the decrease in the expression levels of Fpn and Fth and the increase in the expression levels of TF observed after EHS induction in the mouse hippocampus. In addition, MLT lowered the levels of other key ferroptosis-related molecules, such as xCT and Gpx4, in the EHS and EHS + PBS group compared with the CON group (p < 0.05 for TF, xCT, Fth, Fpn, and Gpx4; Fig. 5B–F). In addition, a significant increase in the total iron content was observed in the EHS and EHS + PBS group, which was alleviated by MLT (p < 0.05; Fig. 5G). Ferroptosis-related protein analysis revealed significantly lower (p < 0.05; Fig. 5B–F) expression levels in LUZ-treated group compared with MLT-treated animals, while total iron content reached values indistinguishable from EHS baseline (Fig. 5G). However, the expression levels of Fpn, Fth, xCT, and Gpx4 and the total iron content were not significantly different between EHS, EHS + PBS, and EHS + MLT + LUZ groups.
MLT relieves EHS-induced cognitive dysfunction in mice
The OF test confirmed comparable locomotor activity across groups, as evidenced by similar total distance traveled and line crossings 24 h post-EHS (representative OF trajectories shown in Supplementary Fig. S1A; p = 0.95 and 0.61 for total distance and line crossings, respectively; Supplementary Fig. S1B, C).
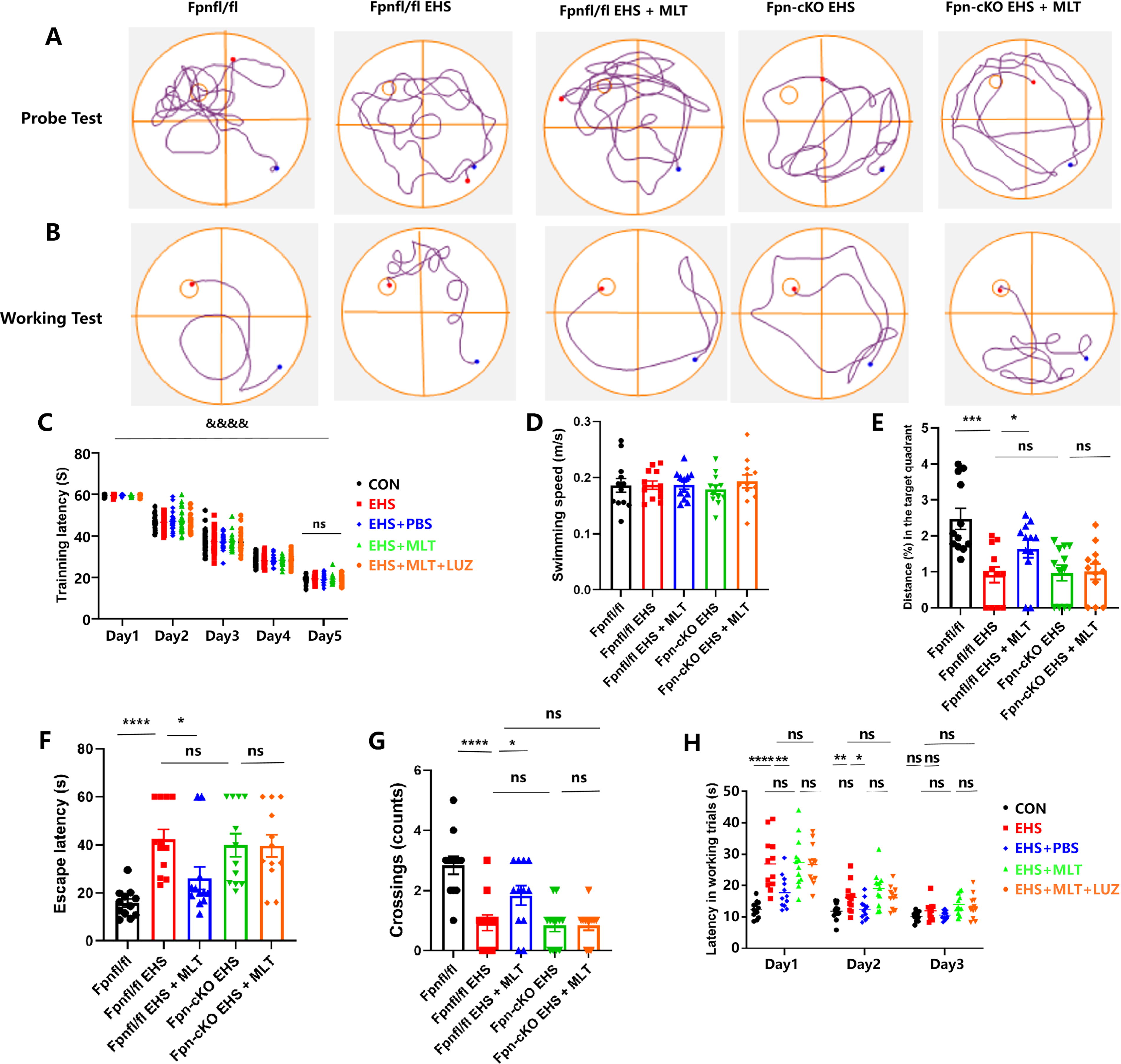
Morris water maze (MWM) assessments revealed EHS-associated cognitive impairment. There were no significant differences in platform latency among the different groups of mice on a daily basis (p > 0.05; Fig. 6C). The differences in swimming speed among the five groups were not statistically significant, indicating that performance differences were not caused by differences in spontaneous activity or motor ability (p > 0.05, Fig. 6D). During probe testing, EHS and EHS + PBS groups exhibited prolonged escape latency (p < 0.001 vs. CON; Fig. 6E), reduced target quadrant occupancy (p < 0.01; Fig. 6F), and decreased platform crossings (p < 0.001; Fig. 6G). MLT treatment significantly shortened escape latency (p < 0.01 vs. EHS; Fig. 6E) and increased platform crossings (p < 0.05; Fig. 6G), although target quadrant occupancy showed no improvement (p > 0.05; Fig. 6F).
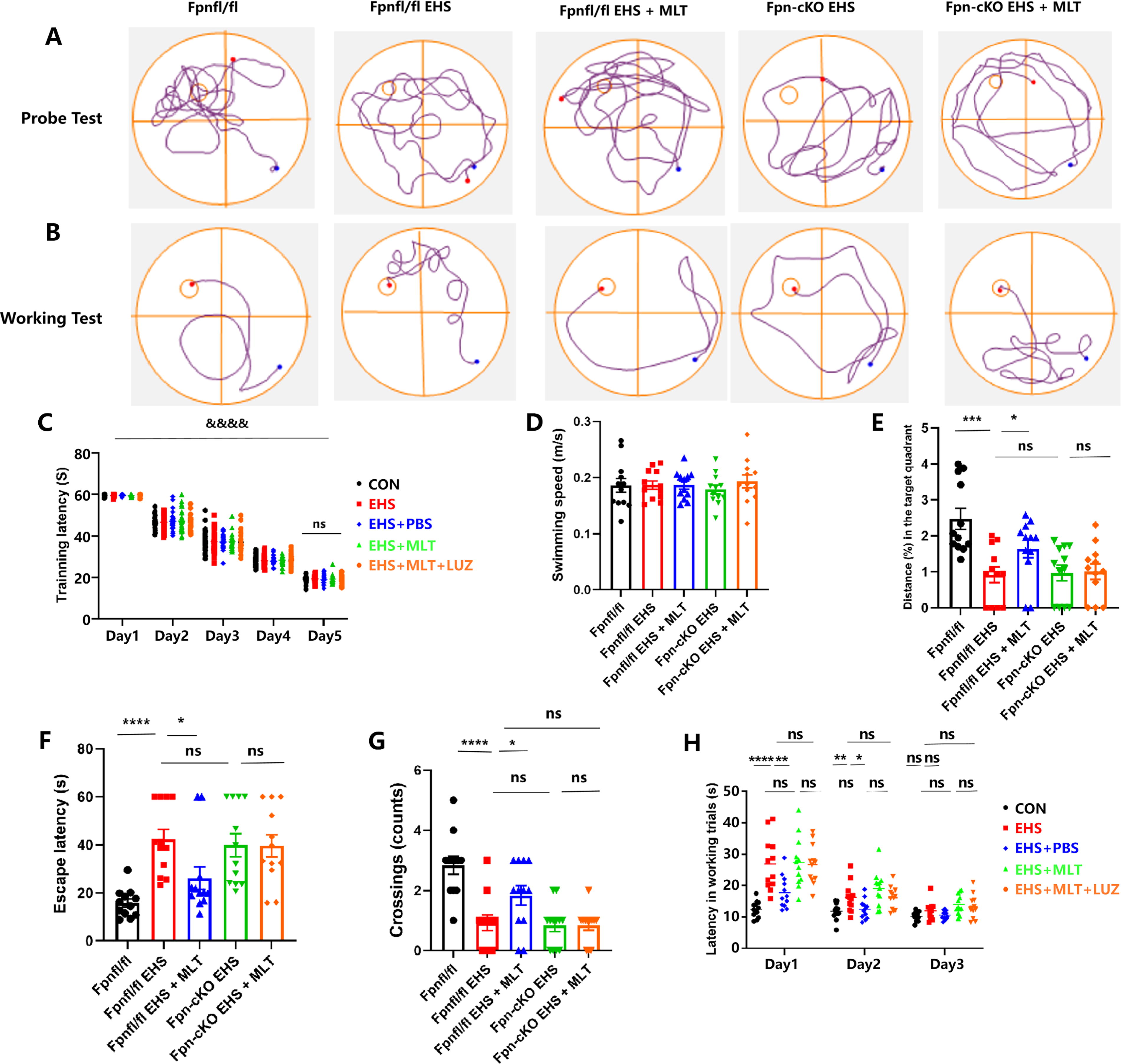
Working memory evaluation demonstrated extended escape latencies in the EHS/EHS + PBS groups versus CON on days 1–2 (p < 0.001; Fig. 6H), which were mitigated by MLT administration (p < 0.001 vs. EHS; Fig. 6H). No intergroup differences were observed in swimming speed (p > 0.05; Fig. 6D) or day 3 performance (p > 0.05).
LUZ cotreatment significantly attenuated MLT-mediated cognitive protection, with EHS + MLT + LUZ group demonstrating prolonged escape latency (p < 0.01 vs. MLT; Fig. 6E) and reduced platform crossings (p < 0.01; Fig. 6G) compared with MLT-treated animals. No parameter exhibited significant differences between EHS and EHS + PBS groups (p > 0.05; Fig. 6D–H).
Fpn ablation abolishes MLT-mediated neuroprotection in EHS
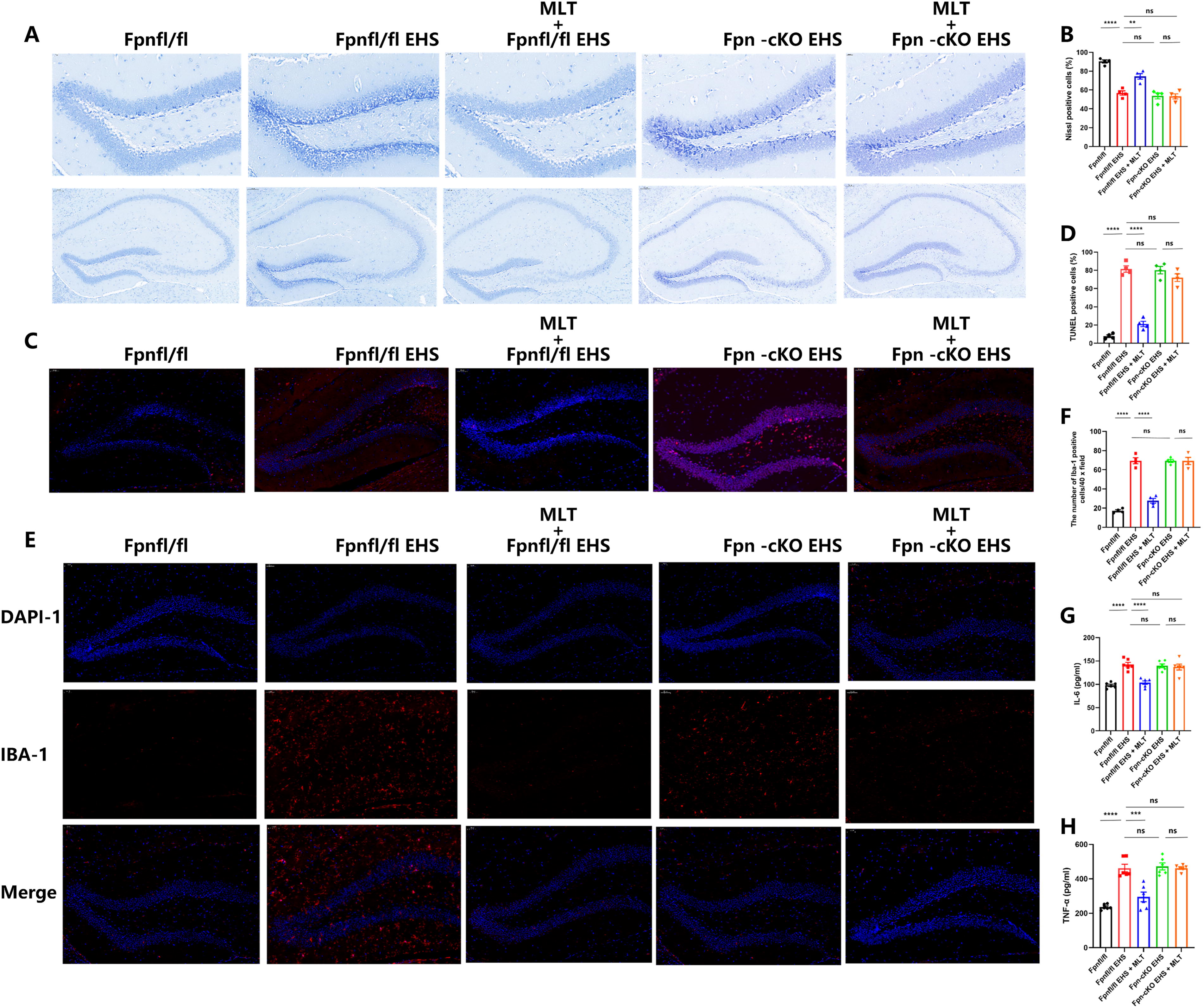
Before phenotypic characterization, Fpn ablation was confirmed through WB analysis of four randomly selected Fpn-cKO mice. The hippocampal tissues exhibited complete depletion of Fpn protein, while the peripheral organs (kidney, liver, heart, and lung) maintained normal Fpn expression levels (Supplementary Fig. S2). Under baseline conditions, Fpn-cKO mice demonstrated significant iron accumulation in the hippocampus (p < 0.0001 vs. Fpnfl/fl; Supplementary Fig. S3E), accompanied by compensatory iron regulatory adaptations: elevated transferrin and reduced ferritin (Supplementary Fig. S3A–D). Lipid peroxidation-related proteins (GPX4, xCT, MD) showed no statistically significant alterations (p > 0.05; Supplementary Fig. S3F–H). Nissl staining combined with inflammatory cytokine analysis (IL-6, TNF-α) revealed that Fpn ablation did not induce central neuroinflammation in the hippocampus, as evidenced by neuronal architecture and unaltered proinflammatory mediator levels (p > 0.05; Supplementary Fig. S4). During probe testing, there were no significant differences among the three groups of mice (p > 0.05; Supplementary Fig. S5).
To assess the impact of Fpn deficiency in the hippocampus on the neuroprotective effects of MLT against EHS-induced neuronal damage and neuroinflammation, analyses including Nissl staining (Fig. 7A, B), TUNEL staining (Fig. 7C, D), and evaluation of inflammatory cytokine (Fig. 7E–H) expression were conducted. The Fpn-cKO EHS + MLT group did not exhibit a higher number of Nissl bodies compared with the Fpn-cKO EHS group (p > 0.05; Fig. 7A, B) and microglial activation (p > 0.05) comparable with untreated Fpn-cKO + EHS controls.
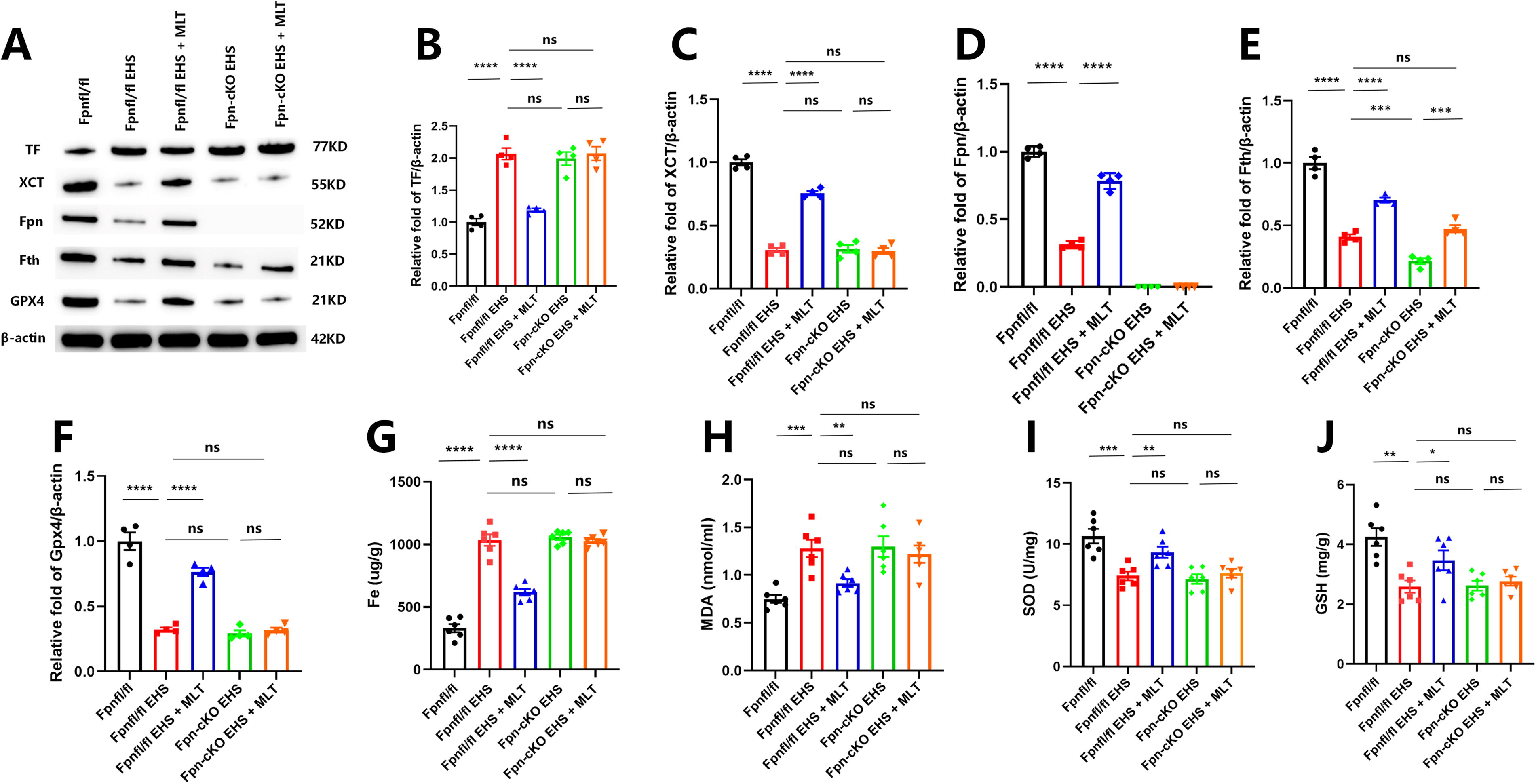
To investigate the impact of neuronal Fpn deficiency on MLT’s modulation of ferroptosis post-EHS, we analyzed potential biomarkers linked to ferroptosis and oxidative stress. In Fpnfl/fl mice, MLT treatment significantly decreased lipid peroxidation levels (MDA: p < 0.05; GSH: p < 0.05 compared with Fpn-cKO + EHS) and reduced expression of GPX4 and xCT (p < 0.05 compared with MLT-treated Fpnfl/fl mice). The MLT-mediated restoration of iron homeostasis (hippocampal iron: p < 0.05; Fth: p < 0.05) and preservation of mitochondrial integrity observed in Fpnfl/fl mice were absent in Fpn-KO mice (Fig. 8A–E).
MWM testing revealed MLT-mediated improvements in Fpnfl/fl mice (escape latency: p < 0.01; platform crossings: p < 0.001), which were absent in Fpn-cKO + EHS + MLT group (p > 0.05 vs. Fpn-KO + EHS). No intergroup differences in locomotor activity (p > 0.05; Supplementary Fig. S3) confirmed cognitive-specific effects (Fig. 9A–H).
Discussion
CNS dysfunction is a frequent complication of EHS, potentially leading to a high incidence of lasting neurological aftereffects, such as reduced learning and memory capabilities (Argaud et al., 2007). However, the mechanism by which EHS induces cognitive impairment remains unclear. Through the animal experiments in this study, we established that EHS can lead to cognitive impairment, which peaks at 24 h post-EHS exposure. To elucidate the underlying mechanisms that cause cognitive impairment, we conducted a transcriptome analysis of the hippocampus at 24 h when the most severe cognitive damage was observed. Bioinformatic analysis revealed differences in the expression of Mt1 and Mt2, ferroptosis-related genes Slc40a1 and Ftl, and inflammation-related genes such as Il6ra, Tnfrsf1b, and Il33. KEGG enrichment analysis of the altered genes confirmed that ferroptosis induction plays a crucial role in EHS-related cognitive impairment.
Among markers of the ferroptosis pathway, the gene Slc40a1, which encodes Fpn, displayed the most notable downregulation with a log2 fold change of −1.05 (|log2FC| > 1), indicating a substantial decrease in iron efflux capacity under EHS-induced stress. Conversely, the gene Ftl, encoding the ferritin light chain, exhibited minimal change (log2FC = 0.43), while Tf, which encodes transferrin, showed a moderate downregulation (log2FC = −0.79) that did not reach statistical significance (|log2FC| < 1). The significant suppression of Fpn, known as the primary cellular iron exporter (Zhang et al., 2018), renders hippocampal neurons vulnerable to lethal iron accumulation, suggesting that targeting Fpn could be a strategically important approach to investigate the therapeutic potential of MLT. In conclusion, this study establishes that MLT effectively mitigates cognitive impairments in EHS mice by suppressing iron-induced cell death and central inflammation via the enhancement of Fpn expression.
Based on the results of transcriptome sequencing and prior research, we hypothesized that MLT could potentially mitigate cognitive function impairments induced by EHS by inhibiting ferroptosis in the hippocampus of EHS mice. Consequently, MLT was administered immediately following EHS, with dosage selection primarily informed by existing studies on MLT dosages in various animal models (Tian et al., 2013; Gao et al., 2023; Zhu et al., 2024; Wang et al., 2021a). Furthermore, considering that MLT is a significant biologically active compound capable of crossing the blood–brain barrier through multiple mechanisms (Akgun-Unal et al., 2023; Chavarria et al., 2023; Zhu et al., 2024; Cakir Cetin et al., 2021; Kong et al., 2021), we opted for intraperitoneal injection as the method of administration. Ultimately, this study demonstrates that MLT can alleviate cognitive impairment in EHS mice, while also reducing oxidative stress, inflammatory responses, iron accumulation, and mitochondrial damage.
The previous studies have demonstrated that exposure to environmental heat can lead to cognitive deficits (Laxe et al., 2013; Zhang et al., 2016). MWM and NOR tests assess learning and memory in rodents, showing that environmental heat exposure causes cognitive deficits. The OF test at 24 h post-EHS showed no significant movement variance, leading to the decision to conduct NOR and MWM tests then, which indicated a decline in cognitive abilities, particularly at 24 h. This emphasizes the immediate impact of EHS on cognition and the need for early intervention.
Previous studies confirmed MLT’s diverse biological functions, including antiapoptotic, antioxidant, and anti-inflammatory properties (Ding et al., 2014; Luo et al., 2019), and its role in ameliorating organ damage from various diseases (Onk et al., 2016; Gao et al., 2021; Zhao et al., 2022; Cheng et al., 2023; Gao et al., 2023). Researchers increasingly recognize MLT’s significance in neuroprotection for neurodegenerative diseases and brain injuries (Fang et al., 2022; Watanabe and Hattori, 2025; Wang et al., 2021a). However, the role and mechanisms of MLT in cognitive dysfunction from EHS. This study aims to show MLT’s effectiveness in alleviating EHS-induced impairments in learning and memory and explore its specific mechanisms.
Transcriptome sequencing showed that inflammation-related genes, such as Il6ra, Tnfrsf1b, and Il33, are differentially expressed. MLT acts via G-protein-coupled receptors Mt1 and Mt2 enhancing nuclear factor erythroid 2-related factor 2 (Nrf2) accumulation, increasing anti-inflammatory cytokines (IL-10, IL-4), and reducing proinflammatory cytokines (IL-1β, TNF-α) in serum and brain (Wang et al., 2019). Inhibitors of MT1/2 lower anti-inflammatory cytokines (IL-33, IL-13, IL-10) and raise proinflammatory cytokines (TNF-α, IL-6) (Liu et al., 2020b; Gao et al., 2023), underscoring the importance of MT1/2 in anti-inflammatory responses. Moreover, studies have demonstrated that heat shock protein (HSP) regulates central inflammation and inflammatory factors (Calabrese et al., 2006). MLT inhibits IL-6 production by upregulating HSP expression, reducing serum IL-6, and increasing HSP70 (Wang et al., 2021b). Our findings indicate that EHS causes nerve cell damage and increased proinflammatory cytokines in mice, while MLT treatment repairs damage and reduces inflammation, laying the groundwork for future research on MLT’s mechanisms (Ashraf et al., 2020; Du et al., 2019; Yao et al., 2023). Our study extends this paradigm to EHS, where KEGG pathway analysis identified ferroptosis as a dominant contributor to hippocampal damage. EHS-triggered distinct ferroptotic hallmarks as follows: iron overload, mitochondrial cristae disruption, and elevated lipid peroxidation markers. Critically, the downregulation of Slc40a1 and compensatory upregulation of iron-storage protein Fth suggest a failed adaptive response to iron dyshomeostasis. These findings align with clinical observations of hippocampal iron accumulation postheat stroke (Weiland et al., 2019) and establish Fpn as a central regulator in this pathology.
Fpn, as the sole iron export protein in mammals, is indispensable for neuronal iron efflux. Previous studies have linked the deficiency of Fpn with iron retention, oxidative stress, and cognitive deficits in Alzheimer’s disease models (Bao et al., 2021). Our work confirms the following: Hippocampus-specific deletion of Fpn leads to baseline iron accumulation, reduced exploratory behavior, and increased sensitivity to EHS-induced ferroptosis. Notably, MLT restored iron homeostasis and cognitive function in Fpnfl/fl mice but failed to do so in the Fpn-KO model, clearly indicating that the benefits of MLT depend on Fpn-mediated iron export. This mechanism is consistent with the known role of MLT in regulating iron regulatory genes (Zhao et al., 2024) but provides the first evidence of its dependence on Fpn in the CNS.
Recent studies show that MLT affects iron metabolism by upregulating hepcidin in hepatocytes, regulating ferritin expression for iron storage, and influencing Fpn gene expression (Huang et al., 2022; Zhao et al., 2024). Previous research has indicated that the levels of iron in the hippocampus increase postheat stroke (Weiland et al., 2019).
Lipid peroxidation in ferroptosis occurs when oxygen reacts with lipids, making the brain vulnerable to oxidative stress due to high reactive oxygen species (ROS) levels. SOD scavenges ROS, while MDA is a by-product of lipid peroxidation (Cenini et al., 2019; Liu et al., 2020a; Bao et al., 2021; Cui et al., 2021). Increased lipid peroxidation may trigger ferroptosis, a process regulated by GSH, xCT, and Gpx4 (Imai et al., 2017; Lei et al., 2019). MLT reduces ferroptosis by modulating lipid metabolism, upregulating antioxidant enzymes such as GPX4, and enhancing resistance to oxidative stress via the Nrf2 pathway (Ren et al., 2023; Ma et al., 2023). In this study, MLT mitigated EHS-induced ferroptosis through a dual mechanism as follows: (1) Restoring Fpn expression to resolve iron overload and (2) directly quenching lipid peroxidation via GPX4/xCT upregulation and MDA/GSH normalization. The reversal of mitochondrial damage further highlights MLT’s role in preserving redox equilibrium. Crucially, Fpn ablation abolished these effects—MLT-treated Fpn-KO mice exhibited persistent iron retention, unmitigated lipid peroxidation, and microglial activation. This underscores that Fpn is not merely incidental but essential for MLT’s antiferroptotic action.
Extensive inflammation is associated with the regulation of ferroptosis, although the underlying mechanisms have not been fully elucidated. Inflammatory mediators such as IL-6 promote the production of hepcidin, leading to the degradation of Fpn and iron accumulation, thereby facilitating the onset of neurodegenerative diseases (Ikeda et al., 2020; Haschka et al., 2021; Zhang et al., 2018). In the case of subarachnoid hemorrhage, liproxstatin-1 can inhibit the activation of microglia and reduce inflammatory markers (Lei et al., 2019). These findings underscore the interplay between ferroptosis and inflammation, suggesting that ferroptosis may play a significant role in triggering inflammatory responses. IL-33 possesses antioxidant properties and inhibits lipid peroxidation, while MLT enhances the expression of key proteins through the IL-33 pathway (Wang et al., 2020; Xie et al., 2022). Our research indicates that MLT can alleviate cognitive impairment induced by exercise-induced heat stress, reduce hippocampal neuronal damage, mitigate inflammatory responses, and suppress ferroptosis, providing new intervention targets for cognitive dysfunction associated with EHS. Although our study established Fpn as a key mediator of the neuroprotective effects of MLT in EHS-induced cognitive impairment, several limitations warrant cautious interpretation and further investigation. First, while we demonstrated that MLT rescues hippocampal iron homeostasis and inhibits ferroptosis/inflammation by upregulating Fpn, the precise molecular mechanisms linking MLT receptor (MT1/2) activation to Fpn expression remain unresolved. Second, while hippocampal-specific Fpn knockout (Fpn-cKO) confirmed the necessity of Fpn for MLT’s effects, complementary gain-of-function studies (e.g., Fpn overexpression) are needed to confirm its sufficiency in reversing EHS pathology. Third, this study only assessed changes in the relevant indicators 24 h after EHS, which is the final time point for observing cognitive impairment. The chronological order of EHS occurrence and the neuroprotective effects of MLT remain unclear. Moreover, it is unclear if other cell death forms such as neutrophil inflammatory death and pyroptosis contribute to EHS-related cognitive impairment; the interplay with ferroptosis needs more study. Fpn-cKO mice show iron buildup and cognitive issues, but lack of human hippocampal validation limits insights. Future research with heat stroke patient brain samples or human neuronal models would improve clinical relevance.
Conclusion
Our research findings illustrate a causal chain. EHS-induced downregulation of Fpn drives iron overload in the hippocampus, leading to ferroptosis and neuroinflammation. MLT interrupts this cascade by enhancing Fpn-dependent iron export, thereby protecting neuronal integrity and cognitive function. The benefits of MLT in Fpn-KO mice are completely reversed, confirming that Fpn is a critical component of the mechanism. These insights suggest that MLT holds promise as a potential therapy for EHS-related neurodegenerative diseases and highlight the broad potential of Fpn-targeting strategies in ferroptosis-related CNS disorders.
Materials and Methods
Animals
Male wild-type C57BL/6J mice (8 weeks old, weighing 25–30 g) were obtained from Beijing SPF Animal Technology Company (Beijing, China; license number SCXK [Beijing] 2023-0010). Hippocampal-restricted deletion of Slc40a1 (encoding Fpn) was achieved using CRISPR-Cas9-mediated genome editing in collaboration with GemPharmatech Co., Ltd. (Nanjing, China). Briefly, two sgRNAs flanking Slc40a1 exon 3 were designed using the MIT CRISPR Design Tool (http://crispr.mit.edu/) and cloned into the pGK1.1 vector. A donor vector containing loxP sites and homologous arms was constructed for homology-directed repair. Cas9 mRNA and sgRNAs were synthesized via in vitro transcription. A mixture of Cas9 mRNA, sgRNAs, and donor DNA was microinjected into C57BL/6 zygotes using a TE2000U micromanipulator. Founder (F0) mice were genotyped by PCR and Sanger sequencing of tail DNA. Germ-line-transmissible F1 heterozygotes (Slc40a1fl/wt) were crossed with Scnn1a-P2A-iCre mice (hippocampal neuron-specific Cre, GemPharmatech Strain T037471) to generate Slc40a1fl/fl; Scnn1a-P2A-iCreki/wt mice. Littermates lacking Cre (Fpnfl/fl) served as controls. Hippocampal Fpn knockout (Fpn-cKO) was validated by WB. The mice were provided ad libitum access to food and water and were housed under controlled conditions (22°C–25°C temperature, 40%–55% relative humidity, and a 12-h light/dark cycle). All animal procedures were conducted in accordance with guidelines established by the Animal Welfare Ethics Committee of the Chinese People’s Liberation Army General Hospital (2023-X19-15).
Mouse EHS model
Before initiating the heating and exercise protocols, all mice in each group were acclimated to the equipment and conditions used in the study. This included exposure to an artificial climate chamber (Model LTH-575N-01; Shanghai Longyue, Shanghai, China), a small animal running stage (Model ZS-PT-III; Beijing Zhongshi, Beijing, China), and a rectal thermonuclear probe (BW-TH1101; Billion, Shanghai, China). The adaptive training was conducted at an environmental temperature of 27°C ± 0.5°C, with a relative humidity of 55% ± 5%. The training protocol required the mice to run on a treadmill for 2 h each day at a speed of 10 m/s over a period of 2 weeks, followed by a 2-day rest period before the commencement of the main experiment. During this period, mice were given ad libitum access to food and water, and their weights were recorded 1 h before the experiment.
Based on previous research, a labor-induced thermal fire model was established. To induce EHS, the mice were placed in the climate chamber under the conditions of high temperature and relative humidity (39.5°C, 65%). Mice were then subjected to a forced running protocol at a speed of 10 m/s. The behavior of the mice was closely monitored, and their rectal temperatures were measured at 10-min intervals. The core body temperature (Tc) was measured using a thermal probe (BW-TH1101, Billion). The establishment of EHS was considered successful when two criteria were met simultaneously: the Tc reached 42.7°C, and the mice exhibited no reaction after three consecutive painful stimuli (He et al., 2020; Song et al., 2022).
Drug administration
MLT, also known as N-acetyl-5-methoxytryptamine, was obtained from Sigma-Aldrich (St. Louis, MO) and dissolved in anhydrous 0.1 M PBS to achieve a final concentration of 1 g/L. Immediately following EHS, a single intraperitoneal injection of MLT at a dosage of 10 mg/kg was administered. Luzindole (Luz) is a nonselective antagonist of MLT receptors (MT1 and MT2) solved in ethanol/normal saline at a concentration of 2.5% (v/v). The LUZ group was dosed at a dose of 30 mg/kg antagonist pretreatment 30 min before MLT administration to assess receptor specificity. The dosage and administration schedule were selected based on previous studies (Tian et al., 2013; Gao et al., 2023).
Experimental groups
In Experiment 1, we investigated the recognition memory characteristics of mice following EHS and the genes associated with EHS-induced recognition impairment. Sixty mice were randomly assigned to two groups, the CON group and the EHS group, with 30 mice in each group. All mice underwent adaptive training, whereas the mice in the EHS group were subjected to EHS. Subsequently, 12 mice from each group were randomly chosen for the OF and NOR tests. In addition, RNA was extracted from the remaining hippocampal tissues of mice for transcriptome sequencing.
In Experiment 2, the potential mechanism by which MLT attenuated EHS-induced cognitive impairment was explored. Three hundred and fifty mice were randomly divided into four groups (n = 70 per group) as follows: CON, EHS, EHS + PBS, EHS + MLT, and EHS + MLT + LUZ. All mice were given adaptive training. In the EHS + PBS group, PBS was injected intraperitoneally immediately after EHS induction. Mice in the EHS + MLT group were administered MLT intraperitoneally immediately after EHS induction. Following EHS, 12 mice from each group were randomly selected for OF and MWM tests. In addition, six mice from each group underwent hippocampal ELISA, while the whole brains of four mice per group were incubated in 4% paraformaldehyde for at least 48 h. Subsequently, the brains were protected by freezing in 30% sucrose at 4°C for 48 h, after which Iba-1 immunofluorescence staining, TUNEL staining, and Nissl staining were performed. The hippocampi of three mice in each group were fixed with 2.5% glutaraldehyde overnight at 4°C for mitochondrial transmission electron microscopy. Furthermore, the remaining hippocampal tissues were stored at −80°C for immunoblotting, real-time qPCR, and iron content detection.
In the experiment, 300 mice were divided into five groups to investigate the impact of genetic ablation of Fpn on the therapeutic effects of MLT in preventing hippocampal ferroptosis and neuroinflammation. Cognitive function was assessed using MWM and NOR tests. Hippocampal cytokines, iron levels, neuronal survival, microglial activation, apoptosis, and mitochondrial ultrastructure were evaluated. Protein analysis was conducted through WB and qPCR.
OF test
The OF test is commonly used to evaluate rodent activity, exploratory behavior, and the presence of anxiety and depression. Furthermore, it is often used to assess whether alterations in behavioral performance are linked to changes in motor activity. The mouse was positioned in one corner of the test box, which had dimensions of 60 × 60 × 40 units, with the base divided into 16 equal areas. The mice were allowed to explore freely for 10 min. Various behaviors of the mice were recorded, such as the distance traveled in both the OF and the central area, the number of entries made, and the time spent in the central area.
NOR test
The NOR task was divided into three key stages as follows: familiarization, instruction, and investigation. This is a standard method used to assess an animal’s capacity to interact with a new item based on the rat’s innate inclination to identify new items in its surroundings (Antunes and Biala, 2012). Recognition of novel objects was operationally defined as sniffing, touching with the forepaws, or guiding with the nose at a distance of ≤2 cm. The TF and time spent exploring novel objects (TN) were recorded. The recognition index = time spent exploring the new object/(time spent exploring the old object + time spent exploring the new object).
MWM test
The MWM test was used to evaluate spatial learning and memory (Vorhees and Williams, 2006). A circular swimming pool filled (diameter = 122 cm) with water (depth = 30 cm) was used with an escape platform (10 cm2) submerged below the water surface. The pool was housed in a visually cued room divided into four separate quadrants. The mouse movements were recorded using video cameras.
Spatial learning ability (platform location) test
To test spatial learning capabilities (platform position), the mice were placed in the water facing the pool wall from any one of the four quadrants. In each trial, the mice were required to swim until they successfully reached the platform within 60 s. In the event of failure, the mouse was assisted to the platform and given 15 s to remember its location, after which it was dried off and returned to its cage, and the subsequent mouse underwent the same trial. This pattern continued until all the mice passed through the first trial. Subsequent trials 2, 3, and 4 were conducted until four trials were completed per day over 5 consecutive days. If any mice failed to reach the platform within the specified time limit over the course of these 5 days, they were eliminated from the study as outliers. Additional data on swimming speed and distance traveled around the platform were also recorded. On day 6, an EHS mouse model was established.
Memory test (probe trial)
After 24 h, the platform was removed from the pool, and each mouse underwent a probe assessment lasting 60 s for memory comparison. The analyses included swimming speed, number of site crossings, percentage of time and distance in the target quadrant, and escape latency.
Working-dependent learning and memory of the mice were assessed from days 1 to 3 following the establishment of the EHS model. To evaluate working- or trial-dependent learning and memory, the platform and mice were moved to different positions daily. Two trials were performed daily, with trial 1 serving as the sample task. During trial 1, the mice were required to grasp the new spot of the platform. Trial 2, the matching task, aimed at gauging memory retention from trial 1. The second test trial began 15 s later. A shorter swimming distance in the test trial indicated better recall from the training. Given the daily relocation of the platform, learning from the previous day was not carried over, thus isolating the assessment of temporary memory from working memory. The record of escape latency during the test trial provided insight into temporary memory. If a mouse failed to find the platform within 60 s of the test trial, the escape latency was set at 60 s. MLT was administered 2 h before the daily behavioral assessment.
Tissue sampling
After inducing severe cognitive impairment in mice 24 h after EHS, hippocampal tissues were collected at this time point for molecular biology analyses in alignment with changes in behavioral performance. Following the successful modeling of EHS, the mice were anesthetized with pentobarbital and perfused with saline to clarify and uniformly whiten the liver. Brain samples were fixed in paraformaldehyde for immunofluorescence staining or cryoprotected in a sucrose solution for Nissl staining. In addition, hippocampal tissues were fixed with glutaraldehyde for electron microscopy and stored for immunoblotting. Commercial assay kits and iron content assays were conducted using hippocampal tissues from additional mice.
Transcriptome sequencing
RNA was extracted from the CON and EHS groups, followed by differential expression analysis using RNA-Seq by Expectation Maximization (RSEM) (v1.3.1; Li and Dewey, 2011) and DESeq2 (v1.4.5), with a significance threshold of Q value ≤0.05 or false discovery rate ≤0.001. Subsequently, Gene Ontology and KEGG enrichment analyses of the differentially expressed genes were conducted, with significant levels of terms and pathways corrected using a stringent Q-value threshold of ≤0.05 (Storey et al., 2021).
Quantitative RT-PCR
Real-time fluorescence quantitative PCR (qRT-PCR) detects the mRNA levels of SLC40A1 (the gene encoding FPN), as well as the mRNA levels of TF and FTH. Total RNA was extracted using the TRIzol reagent (Vazyme, Nanjing, China), following the manufacturer’s instructions. The RNA concentration was determined using the NANO-300 (A Ozo, Hangzhou, China). Subsequently, cDNA was synthesized using Hifair™ lll 1st Strand cDNA Synthesis SuperMix for qPCR (Yeasen, Shanghai, China). qRT-PCR was performed using the CFX-Connect 96 (Bio-Rad, USA) with SYBR Green Master Mix (Yeasen) to conduct real-time PCR on all samples. Differences in gene expression levels normalized to the arithmetic mean of β-actin were determined using the ΔΔCt method. The primers were purchased from Tianyihuayu Biological Technology (Wuhan China), and the sequences are listed in Supplementary Table S1.
Immunofluorescence staining
Brain tissues were paraffin embedded and meticulously cut into 6-μm sections to ensure precision. Next, the sections underwent a 30-min incubation in a 0.1% hydrogen peroxide solution to block endogenous peroxidase activity. This step was followed by another 30-min incubation with 1.5% normal goat serum to prevent nonspecific binding. To specifically detect microglial cells, the sections were incubated overnight at 4°C with an anti-Iba-1 antibody (Santa Cruz Biotechnology, Dallas, TX). The following day, the sections were thoroughly washed and then incubated for 1 h at 22°C with a peroxidase-coupled secondary antibody (Dako, Glostrup, Denmark). After incubation, the sections were washed and observed under a microscope. The activity of glial cells was assessed by observing an increase in cell count and specific morphological changes, such as rounding of cell bodies and thickening of cellular processes. This change in morphology, coupled with an elevated level of Iba-1 labeling, indicated heightened glial cell reactivity. The level of microglial cell reactivity was further quantified by measuring the integrated intensity per pixel area of Iba1 staining. The increased intensity was interpreted as a sign of elevated microglial reactivity. The number of microglia showing positive staining was quantified using a microscope at 200× magnification. Imaging was performed using an Olympus BX5 imaging system Olympus, Tokyo, Japan), and the acquired images were analyzed quantitatively using Image-Pro Plus 6.0.
TUNEL staining
TUNEL staining was performed to measure cell death in the hippocampus 24 h after EHS according to the manufacturer’s instructions, with a cell death assay kit (In Situ Cell Death Detection Kit, TMR red). Hippocampal tissues were fixed in 4% PFA for 24 h, and then embedded in paraffin or cryosectioned. Coronal sections were mounted on poly-l-lysine slides. Deparaffinized sections underwent antigen retrieval with proteinase K, followed by permeabilization with Triton X-100/sodium citrate. The In Situ Cell Death Detection Kit was applied, incubating with TdT enzyme mixture for 1 h. Nuclei were counterstained with DAPI, and TUNEL-positive cells in the hippocampal were counted using a fluorescence microscope at a 20× objective.
Nissl staining
Twenty-four hours after EHS, Nissl staining was used to observe morphological changes in hippocampal neurons. Nissl staining was performed according to the manufacturer’s instructions (Nickel Staining Kit, G1036, Servicebio, Wuhan, China). Briefly, slices of the hippocampal region were flushed with 0.01 M PBS and dipped in dimethylbenzene I, Ⅱ, III at concentrations of 90%, 80%, and 70%, respectively, for 5 min each. The slices were then covered with Nissl staining solution for 10 min at 37°C in the dark. After incubation, the slices were immersed in 95% alcohol for color separation. The total number of Nissl-positive neurons in the hippocampal region under a light microscope (BX5; Olympus, Tokyo, Japan).
Transmission electron microscopy
One cubic millimeter of tissue from the dentate gyrus area of the mouse hippocampus was collected after establishing the EHS model. The brain tissue was made into a 50-μm-thick tissue block by fixation, rinsing, dehydration, infiltration, and embedding. Then, it was double-stained with uranium-lead at 4°C for 20 min and observed under an HT7700 projection electron microscope (120 kV; Hitachi, Tokyo, Japan).
Enzyme-linked immunosorbent assay
IL-6 and TNF-α levels were measured using ELISA kits (R&D Systems, Minneapolis, MN). Hippocampal tissues were homogenized using radioimmunoprecipitation lysis buffer (Applygen, China), and the protein concentration of the supernatant was determined by centrifugation at 12,000 g for 15 min using a BCA protein assay kit (Thermo Fisher Scientific, Waltham, MA).
Assay of SOD activity and concentrations of MDA and GSH
The levels of MDA, GSH, and SOD were measured 24 h after EHS was induced. Measurements were performed using a commercial assay kit (Dojindo, Kumamoto, Japan).
Detection of iron levels
The iron concentration in the hippocampus was measured following the guidelines provided in the Iron Content Assay Kit (Dojindo). In summary, hippocampal tissue was weighed and homogenized, and the resulting supernatant was collected. The supernatant was combined with a solution containing iron-detection reagents. Subsequently, absorbance was measured at 450 nm using an enzyme marker on a SpectraMax i3× instrument (Molecular Devices, San Jose, CA).
Western blot
WB was performed to analyze the expression of key factors in the hippocampal tissues of mice from each experimental group. Hippocampal tissue proteins were extracted and separated using gel electrophoresis based on their molecular weights. These proteins were subsequently transferred onto a polyvinylidene fluoride (PVDF) membrane and incubated with specific primary antibodies overnight at 4°C. The primary antibodies were as follows: anti-Gpx4 (1:1000, ab125066; Abcam, Cambridge, UK), anti-Fth (1:1000; Cell Signaling Technology, Danvers, MA), anti-SLC7A11/xCT (1:1000; Cell Signaling Technology), anti-TF (1:1000, DF13383; Affinity Biosciences, Cincinnati, OH), anti-SLC40A1/FPN1 (1:1000; Abcam), anti-IL-33 (1:200; R&D Systems, AF3626), and anti-β-actin (1:1000; Cell Signaling Technology). Subsequently, the PVDF membrane was washed three times with TBST and incubated with a secondary antibody at room temperature for 1.5 h. The protein concentration was detected using an Odyssey scanner (LI-COR, Lincoln, NE) and analyzed using ImageJ analysis software (National Institutes of Health, Bethesda, MD), with normalization to glyceraldehyde 3-phosphate dehydrogenase.
Statistical analysis
GraphPad Prism (version 8.0; GraphPad Software, San Diego, CA) was used for statistical analysis. Differences among the four experimental groups were compared using a two-way analysis of variance followed by Tukey’s post hoc test. Statistical significance was set at p < 0.05, and the results are presented as mean ± standard error.
An electronic laboratory notebook was not used.
Footnotes
Acknowledgments
The authors thank Nan Kai University, the First Medical Centre, the Chinese PLA General Hospital, and the National Clinical Research Center for Kidney Diseases for their support.
Authors’ Contributions
F.Z.: Conceptualization, formal analysis, and writing—review and editing. X.W.: Conducted experiments, data curation, investigation, formal analysis, and writing—original draft, and review and editing. Z.H.: Conducted experiments, data curation, formal analysis, investigation, and review and editing. C.L.: Formal analysis and writing—review and editing. J.L.: Data curation, investigation, and writing—review. Z.D.: Data curation, investigation, and formal analysis. J.H., Z.M., Q.L., and X.H.: Writing—review and editing.
Data Availability
Data are available from the corresponding author upon reasonable request.
Author Disclosure Statement
The authors declare no conflicts of interest.
Funding Information
No funding was received for this article.
Supplementary Material
Supplementary Data
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.