
Abstract
Salmonella enterica
is a leading cause of foodborne illness worldwide, responsible for an estimated 93.8 million cases and approximately 155,000 deaths annually, according to the World Health Organization. This foodborne pathogen imposes a significant burden on public health and the global economy. A key factor contributing to the persistence and widespread impact of S. enterica is its potential to form biofilms, which may enhance its survival in clinical, industrial, and agricultural environments, making it a major and ongoing public health concern. Biofilms are structured microbial communities encapsulated in a self-produced extracellular matrix that protects against environmental stressors, disinfectants, and antimicrobial agents. This complex phenotype enables Salmonella to colonize food-contact surfaces, medical devices, and host tissues, hampering efforts to eliminate contamination and control transmission. The poultry industry, a key component of the global food supply, is particularly vulnerable to emerging Salmonella strains with increased virulence, stress tolerance, and disinfectant resistance, making biofilm control a top priority.
This review aims to provide an updated and comprehensive overview of the mechanisms involved in Salmonella biofilm formation, its implications for food safety, and recent advances in detection and control strategies. Emerging technologies such as CRISPR-Cas systems are receiving particular attention due to their potential as precise molecular tools for investigating genes implicated in biofilm formation.
By integrating current findings, this review underscores the urgent need for novel and effective strategies for biofilm control. It highlights the importance of a One Health approach that links human, animal, and environmental health to address the risks posed by Salmonella biofilms in the food production and public health sectors.
Introduction
Salmonella is a genus of Gram-negative bacteria within the Enterobacteriaceae family, comprising numerous medical and veterinary significant pathogens (Graziani et al., 2017). It is one of the leading global causes of foodborne illness and continues to pose a major public health challenge.
The bacterium was first isolated in 1884 from a pig’s intestine by the American bacteriologist D.E. Salmon, who initially named it Bacillus choleraesuis (Salmon and Smith, 1886). Later, in 1900, Lignières renamed it Salmonella choleraesuis, establishing the nomenclature used today (Euzéby, 1999). Advances in genomic and serological analyses have since refined its taxonomy, and the genus currently includes two species: Salmonella enterica and Salmonella bongori. Of these, S. enterica is the most clinically relevant, comprising six subspecies and more than 2600 identified serovars (Brenner and McWhorter-Murlin, 1998).
Based on pathogenicity and host range, S. enterica serovars are classified into typhoidal and nontyphoidal groups (Su and Chiu, 2007). Typhoidal serovars, such as S. Typhi and S. Paratyphi, are human-specific and cause typhoid fever, a systemic and potentially life-threatening disease (Dougan and Baker, 2014). In contrast, nontyphoidal serovars (NTS), including S. Typhimurium and S. Enteritidis, infect a wide range of hosts and are typically associated with self-limiting gastroenteritis, though they can occasionally progress to invasive disease (Lamichhane et al., 2024). Given their broad distribution, persistence in food environments, and public health impact, this review focuses on NTS and their role in biofilm formation as it relates to food safety.
Early diagnosis of Salmonella infection is crucial to reducing morbidity and preventing its spread (Kurtz et al., 2017). Recent advances in rapid and precise detection methods have enhanced bacterial identification and contributed to more effective control strategies (Montoro-Dasi et al., 2023). However, the growing problem of antimicrobial resistance (AMR) among certain serovars poses a major public health challenge, highlighting the importance of epidemiological surveillance and responsible antibiotic use (Crump et al., 2015).
The mechanisms underlying AMR in Salmonella are multifactorial, but biofilm formation is a particularly important contributor (Blair et al., 2015). Biofilms function as physical barriers that limit antibiotic penetration, while the sessile cells within them adopt a reduced metabolic state that further decreases susceptibility to antimicrobial agents (Steenackers et al., 2012). Moreover, biofilm-associated Salmonella can exchange resistance genes, exacerbating the issue (Gamazo et al., 2009). In food processing facilities, Salmonella readily forms biofilms on common contact surfaces such as stainless steel, plastic, and rubber (Tsai et al., 2020). This persistence increases the risk of recurrent outbreaks and contributes to significant economic losses (Merino et al., 2019). The ability of Salmonella to switch between planktonic (free-living) and biofilm-associated states further complicates eradication, underscoring the need for innovative strategies that go beyond conventional sanitation practices (Białucha et al., 2020; Cadena et al., 2019).
This review, therefore, examines the mechanisms underlying Salmonella biofilm formation, the virulence factors involved, and their implications for the food industry. It also highlights emerging strategies to mitigate biofilm-related risks, with a particular focus on the adaptive advantages conferred by biofilms and the development of effective control measures to safeguard food safety in an increasingly complex global supply chain.
Factors of virulence in Salmonella and their role in biofilm formation
Salmonella’s capacity to persist both within the host and in the external environment relies on a complex interplay of virulence factors that converge in the formation of biofilms. The process begins with adhesion to biotic and abiotic surfaces and colonization of host tissues, critical early events that determine the course of infection. Fimbriae, for instance, act as molecular anchors that preferentially recognize intestinal enterocytes, facilitating not only attachment but also translocation to Peyer’s patches, where the bacteria can evade innate immune surveillance (Bao et al., 2020; Edwards et al., 2000). This adhesive step is reinforced by the type I secretion system, which releases adhesins and toxins that strengthen epithelial attachment and lay the foundation for colonization (Holland et al., 2016). Lipopolysaccharide (LPS) (Fig. 1) contributes to biofilm development by mediating the initial adhesion of Salmonella cells to abiotic and biotic surfaces through electrostatic and hydrophobic interactions (Laekas-Hameder and Daigle, 2024; Rhee, 2014). In addition, LPS molecules play a structural role within the extracellular matrix (ECM), enhancing biofilm integrity and providing protection against environmental stressors and antimicrobial agents (Laekas-Hameder and Daigle, 2024). At the same time, the structural variability of LPS, particularly its O antigen of variable length, provides a protective shield that modulates host immune recognition (Krzyżewska-Dudek et al., 2022). While flagella (Fig. 1), beyond driving motility, also guide bacteria to favorable niches for aggregation, a crucial step in the maturation of biofilms (Smith and Selander, 1991).
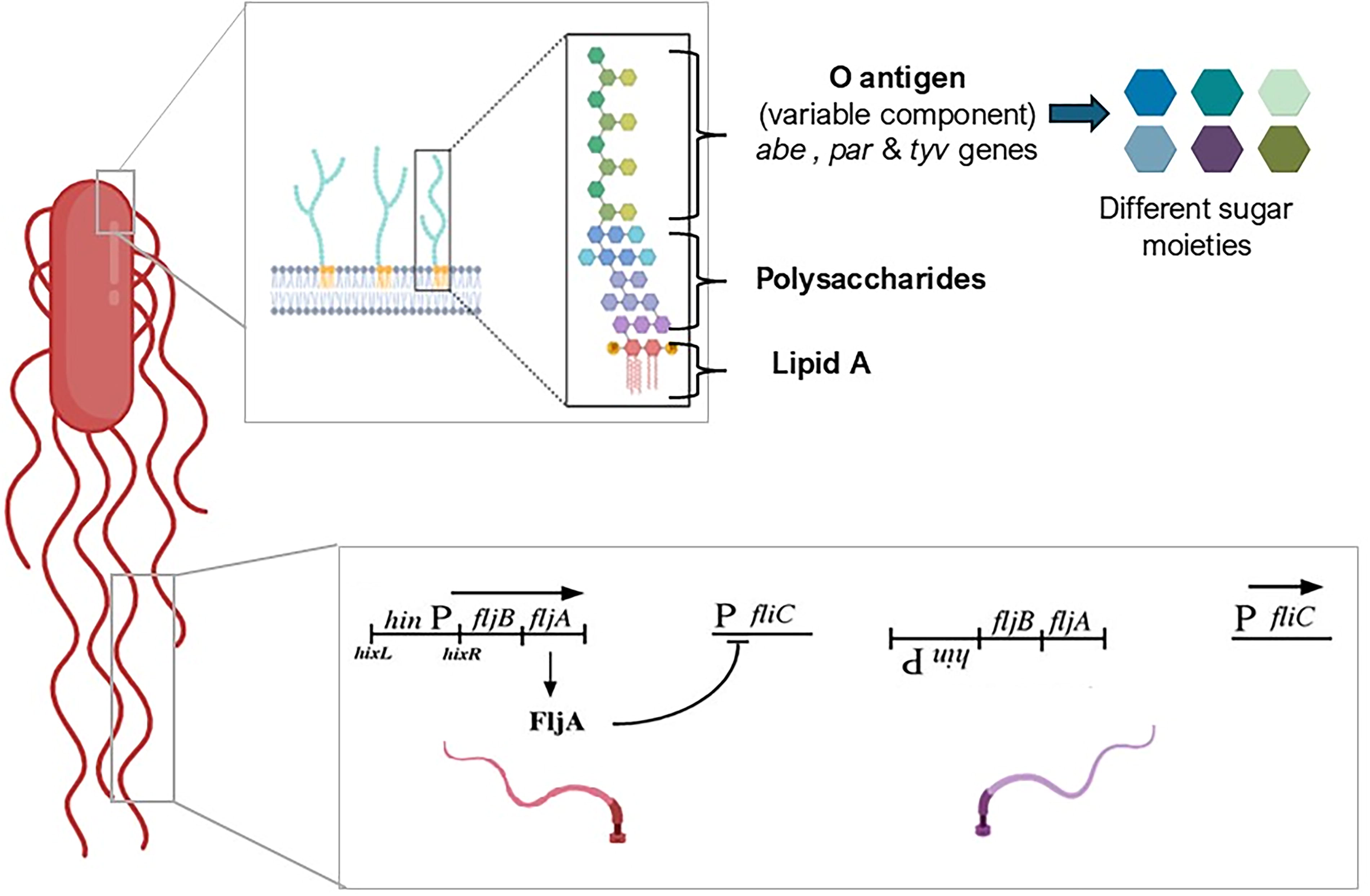
Serotype conversion. The complete LPS molecule consists of lipid A, a core oligosaccharide, and the variable O antigen (OAg). The rfb locus regulates OAg biosynthesis, while genes (abe, par, and tyv) add serovar-specific modifications. Additional genes (oafA, gtrC1, gtrC2, and gtrC3) further modify OAg, contributing to antigenic diversity (Gasperini et al., 2024). Meanwhile, Salmonella alternates between flagellar H antigens (phases 1 and 2) through an 800 bp DNA inversion (hinP), controlling promoter activation. When the promoter is off, fliC is expressed, producing one flagellum. When activated, fljB is expressed instead, along with fljA, which represses fliC, ensuring mutually exclusive flagellar expression. This mechanism allows Salmonella to be biphasic, monophasic, or nonmotile (McQuiston et al., 2004). (This figure was created with Biorender. Available from: https://app.biorender.com accessed on May 7, 2024). LPS, lipopolysaccharides.
Once in contact with host tissues, invasion mechanisms become essential. The pathogenicity islands of Salmonella (SPIs) (Hacker et al., 1997; Park et al., 2018) are genetic reservoirs of virulence (Table 1). Five SPIs (SPI-1 to SPI-5) are conserved across all Salmonella serotypes (Jennings et al., 2017), while others, such as SPIs 19–23, are restricted to specific serovars, including S. enterica Dublin, Gallinarum, and Derby (Duran Gonzalez et al., 2021; Lerminiaux et al., 2020). These islands encode the sophisticated type III secretion systems that deliver effector proteins directly into host cells (Leung and Finlay, 1991). These effectors hijack signaling pathways, remodel the actin cytoskeleton, and induce membrane ruffling, thereby promoting bacterial uptake (Garcia-del Portillo et al., 1993). Invasion is not merely a gateway to the intracellular space; by establishing protected niches, Salmonella ensures replication and creates microenvironments that indirectly favor the persistence and stability of biofilm-associated bacterial populations on mucosal surfaces (Finlay et al., 1991).
Summary of the Main Genes of SPIs
T1SS, type I secretion system; T3SS, type III secretion system.
Equally critical are the strategies that guarantee survival and dissemination under hostile conditions. Siderophores, with their high affinity for iron (Leclerc et al., 2017), secure this essential but limited nutrient and simultaneously contribute to the stabilization of the biofilm matrix (Khasheii et al., 2021). In parallel, stress response systems, including antioxidant enzymes, heat shock proteins, and plasmid-acquired resistance genes (Vilas Boas et al., 2024), provide broad protection against oxidative stress, extreme pH, desiccation, and antimicrobial agents (Arnold, 2024). These adaptive traits are particularly advantageous in extra-host environments such as soil, water, and food-processing facilities, where biofilms act as resilient reservoirs that sustain recurrent contamination (Hall-Stoodley et al., 2004). Within their ECM, composed of curli fimbriae, cellulose, extracellular DNA, and polysaccharides, bacterial cells are shielded from environmental challenges and immune defenses, ensuring long-term survival and transmission (Mirghani et al., 2022). Table 2 summarizes these virulence factors and their molecular mechanisms, showing how adhesion, invasion, and resistance strategies converge to establish biofilm formation as a central pillar of Salmonella pathogenesis and persistence.
Factors of Virulence in Salmonella and Their Role in Biofilm Formation
The importance of Salmonella biofilms in persistence and environmental survival
Salmonella enterica is a highly adaptable pathogen capable of surviving in a wide range of environments, including host tissues, water systems, food processing surfaces, and agricultural settings. One of its most critical survival strategies is the formation of biofilms, which are structured microbial communities embedded in a self-produced ECM (Zogaj et al., 2001). This matrix is primarily composed of curli fimbriae, cellulose, exDNA, and other polysaccharides that provide both physical protection and biochemical resilience against hostile conditions, such as desiccation, disinfectants, antibiotics, and immune responses (Svarcova et al., 2021).
The regulatory network controlling biofilm development is complex (Fig. 2), with csgD acting as the master regulator. This gene promotes the production of curli fimbriae and cellulose, essential ECM components that facilitate surface adherence and intercellular cohesion (Adcox et al., 2016; Atiencia-Carrera et al., 2022; Białucha et al., 2020; Tsai et al., 2020). ECM composition also adapts to environmental conditions, enhancing biofilm resilience by modulating properties such as porosity, mechanical stability, and resistance to desiccation (Bridier et al., 2011; Stewart and William Costerton, 2001).

Schematics of the different stages of biofilm formation.
Biofilm formation allows Salmonella to transition from a planktonic lifestyle to a sessile, community-based lifestyle confers multiple advantages (Giacomodonato et al., 2022). Biofilm-associated cells exhibit increased resistance to antimicrobial agents and enhanced survival on abiotic surfaces, such as stainless steel, plastic, and rubber, all materials commonly found in food processing environments (Silva et al., 2014). Consequently, Salmonella biofilms act as long-term reservoirs for contamination, contributing to recurrent outbreaks even after standard cleaning procedures (Ehuwa et al., 2021).
A major concern is the ability of Salmonella to participate in multispecies biofilms alongside other genera such as Escherichia, Listeria, Pseudomonas, and Staphylococcus (Cufaoglu et al., 2021; Gkana et al., 2017; Milho et al., 2019). In these consortia, interspecies interactions often promote synergistic survival mechanisms. For example, metabolic cooperation and shared matrix production can enhance the overall biofilm biomass, stability, and resistance to environmental stressors (Gkana et al., 2017). Moreover, certain species within the biofilm may degrade disinfectants or sequester nutrients, indirectly benefiting Salmonella. For example, Pseudomonas species are known to produce biosurfactants and matrix components that confer resistance to chemical agents, indirectly shielding Salmonella within the shared matrix (Pang et al., 2017). In turn, Salmonella may benefit from enhanced nutrient retention or pH buffering capacities provided by the consortia (Pang et al., 2023). Conversely, antagonistic interactions are also observed, where microbial competitors secrete bacteriocins, quorum-sensing inhibitors, or resource-sequestering molecules that suppress Salmonella proliferation. Such dynamics have been reported on food contact surfaces and in livestock environments, where the balance between competitive exclusion and mutualistic protection significantly influences Salmonella’s persistence (Visvalingam et al., 2019).
Polymicrobial biofilms also act as hotspots for horizontal gene transfer, driven by high cell density, extracellular DNA presence, and close interspecies contact. These conditions promote the dissemination of AMR genes and virulence factors among Salmonella and its microbial neighbors, compounding the challenge of eradication in clinical and food production settings (Aleksandrowicz et al., 2023).
Salmonella persistence in environmental reservoirs is further linked to its ability to transition into a viable but nonculturable state within biofilms (Vestby et al., 2009). In this dormant phenotype, Salmonella cells can drastically reduce metabolic activity and undergo profound gene expression shifts, making them undetectable by conventional culture-based diagnostics, yet retaining full pathogenic potential (Harrell et al., 2021). A range of environmental stressors, including pH fluctuations, oxidative stress, temperature shifts, and nutrient scarcity, can trigger viable but nonculturable state (dormant) cells. Importantly, once environmental conditions become favorable again, these dormant cells can resuscitate, regaining full virulence and potentially initiating new outbreaks or contaminations. This capacity for phenotypic switching significantly complicates microbial detection, surveillance, and risk assessment in food safety systems (Mah and O’Toole, 2001).
Temperature also plays a pivotal role in modulating both Salmonella survival and biofilm dynamics, making it a critical factor in the context of climate change (Wu et al., 2016). Elevated ambient temperatures associated with global warming can promote active proliferation of S. enterica and accelerate biofilm maturation (Vice et al., 2025). Under such conditions, the bacteria form robust, three-dimensional ECMs that provide structural integrity and enhanced protection against environmental stress and routine sanitization. Conversely, lower temperatures, such as those used in cold-chain logistics or meat processing environments, may reduce metabolic rates but do not eliminate the risk. In fact, these temperatures can still allow initial surface adhesion and biofilm establishment, while simultaneously limiting the efficacy of cleaning agents and delaying bacterial detection (Vice et al., 2025).
Mature Salmonella biofilms formed under thermal conditions optimal for growth can withstand mechanical stress and chemical disinfection more effectively than immature or monolayered communities (Gkana et al., 2017). Furthermore, once established, these biofilms can harbor dormant cells that remain viable even in refrigerated settings, posing a continuous threat of cross-contamination in food production environments (Dixon et al., 2024). As global temperatures rise and extreme weather patterns become more frequent, the ecological stability and distribution of Salmonella are likely to shift, expanding its environmental range and prolonging its survival outside of hosts (Saleem et al., 2024). This prospect may have significant implications for food safety, particularly in the meat, poultry, and produce sectors, where biofilm-associated contamination is already a major concern (Balta et al., 2024). Furthermore, biofilm formation significantly influences host-pathogen interactions. Within the host, biofilm-like structures can develop on surfaces such as gallstones, the intestinal mucosa, and even within macrophages, where they protect bacteria from clearance by the immune system and antimicrobial treatments (Harrell et al., 2021). These protected niches allow bacteria to persist and replicate, turning host cells and tissues into reservoirs and disseminators of infection. Such persistence is particularly relevant in chronic carriage and may facilitate vertical transmission in birds, including transovarial infection (Merino et al., 2019). Consequently, intracellular survival enables infected animals, especially poultry, to act as asymptomatic carriers and disseminators of Salmonella, while biofilm-associated colonization further contributes to vertical transmission routes that pose a significant risk for infection (Merino et al., 2019). In veterinary and agricultural contexts, biofilms on equipment, water lines, or animal hides facilitate transmission both among livestock and between animals and humans (Balta et al., 2024; Chia et al., 2009; Giaouris and Nychas, 2006; Joseph et al., 2001; Kim and Wei, 2009). Although standard sanitation protocols aim to reduce microbial loads, biofilm-embedded Salmonella cells display markedly greater resistance to disinfectants and antimicrobials than planktonic cells, often undermining decontamination efforts (Corcoran et al., 2014; Galié et al., 2018; Joseph et al., 2001; Sheffield et al., 2009). Further compounding this issue, Salmonella can also establish biofilms directly on food products, including raw meats, poultry, fresh produce, and even low-moisture items such as cereals, highlighting its adaptability and reinforcing the view that biofilm formation is a key strategy for persistence outside the host organism (Harrell et al., 2021; Lamas et al., 2018; Podolak et al., 2010; Yaron and Römling, 2014).
Given the intricate connections between environmental, animal, and human reservoirs of Salmonella, it is increasingly evident that an isolated approach to control is insufficient. The persistence of biofilms across ecological compartments underscores the need for an integrated, transdisciplinary strategy. This brings into focus the One Health perspective, which recognizes the interdependence of human, animal, and environmental health (Balta et al., 2024; Locke et al., 2025; Soltan Dallal et al., 2024). By addressing biofilm-associated persistence within this holistic framework, we can better anticipate and mitigate the risks posed by Salmonella across the entire food chain and public health landscape (Vestby et al., 2009).
Impact of Salmonella in Public Health: A One Health Perspective
Salmonella is one of the leading causes of diarrheal diseases worldwide (World Health Organization, 2018). In the United States, it is responsible for around 1.2 million foodborne illnesses, 23,000 hospitalizations, and 450 deaths annually, with contaminated food (especially poultry) being the main source. However, the actual incidence is likely underestimated, as it is estimated that up to 30 cases go unreported for every confirmed one (Steenackers et al., 2012).
The persistence of Salmonella in food processing environments, particularly poultry facilities, has been well-documented (Musa et al., 2024; Obe et al., 2021). Genetically identical strains have been repeatedly isolated from specific locations, even after cleaning and disinfection (Giaouris et al., 2012; Pang et al., 2017). These findings emphasize the role of biofilms as long-term reservoirs that enable reintroduction of the pathogen into the food chain.
As previously mentioned, biofilms have been detected on stainless steel surfaces, conveyor belts, scalding tanks, and drains. Dantas et al. (2018) observed persistent contamination linked to biofilm formation on conveyor belts. Biyashev et al. (2025) reported Salmonella survival through multiple production cycles despite sanitation efforts (Fig. 3). Similarly, Milanov et al. (2017) described repeated isolations from the same points within poultry plants, further supporting biofilm-driven colonization (Biyashev et al., 2025; Dantas et al., 2018; Milanov et al., 2017).
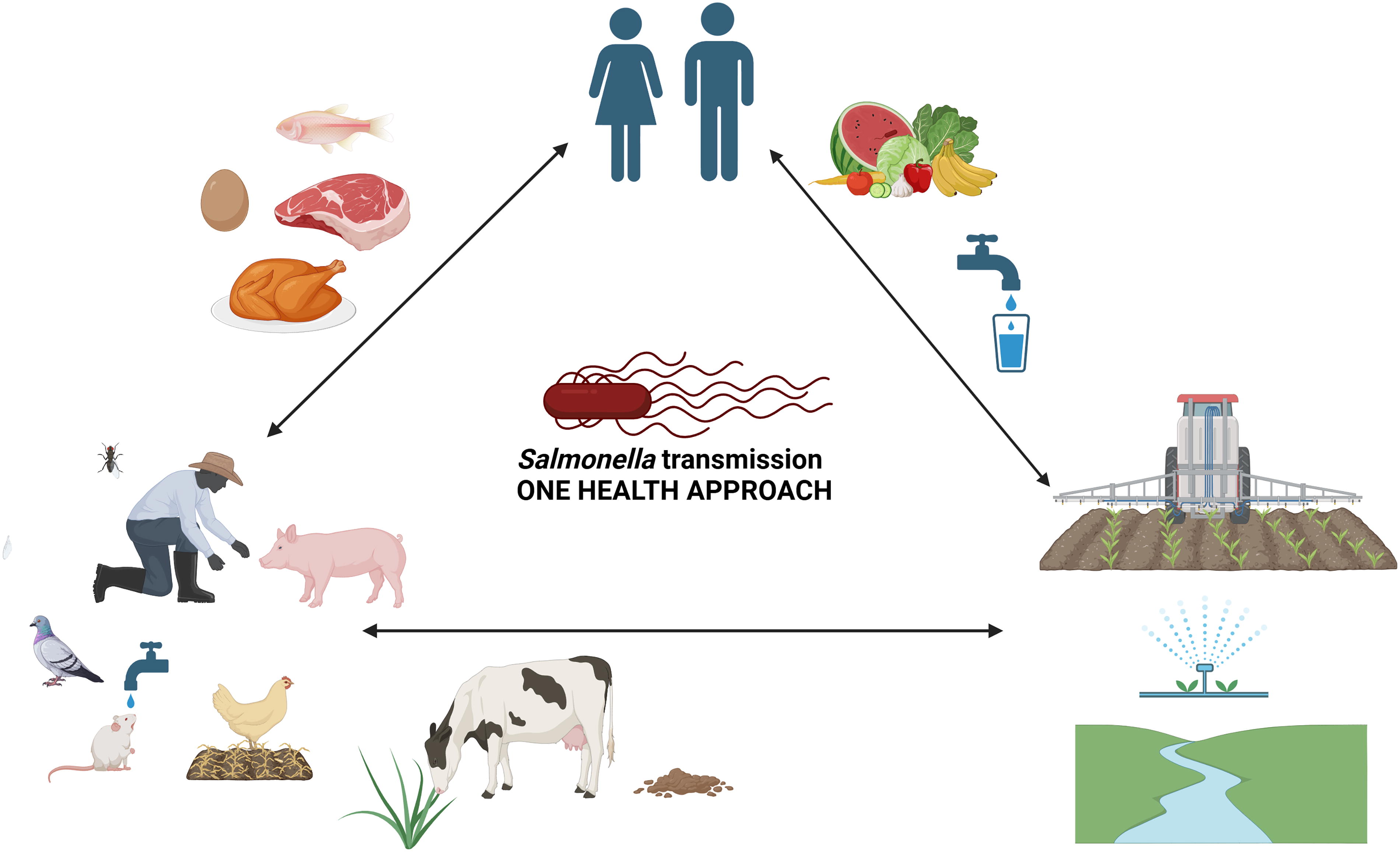
Salmonella ongoing cycle of transmission. Salmonella infection persists through a continuous cycle involving humans, animals, and the environment. It spreads via contaminated food, water, or contact with infected individuals. Wastewater and fecal contamination are major dissemination routes, with water sources acting as key points for reintroducing Salmonella into the food chain. Wild animals may also contribute to its transmission in both natural and agricultural settings. (This figure was created with Biorender. Available from: https://app.biorender.com accessed on June 10, 2024).
In dairy environments, Salmonella has been shown to persist for over 6 months due to biofilm formation on processing equipment (Giaouris et al., 2012). Although current monitoring systems are not designed to directly detect biofilms, the recurrence of strains at critical points and their genetic similarity with clinical isolates support their relevance to public health (Yaron and Römling, 2014).
Table 3 summarizes major Salmonella outbreaks linked to diverse food sources. While poultry remains a primary vehicle (Mejia et al., 2021), contamination can also occur via fecal-oral transmission, water, and vectors such as insects and rodents (Kumar et al., 2019). Vertical transmission can happen in hens during egg formation, contaminating the yolk or shell (Rincón Acero et al., 2011), often without symptoms in birds (Lublin and Sela, 2008).
Overview of Salmonella Outbreaks in Different Food Matrices
CA, Canada; CH, China; CHI, Chile; EU, European Union; FR, France; GR, Greece; RTE, ready to eat; SP, Spain; USA, United States of America; UK, United Kingdom.
Salmonella’s adaptability is driven by its genetic diversity. For example, S. Derby and S. Enteritidis infect multiple hosts (Yang et al., 2019), whereas S. Typhi is human-specific and capable of forming biofilms on gallstones, serving as chronic reservoirs (Chowdhury et al., 2021). Infections may manifest as acute, producing rapid symptoms that can resolve or progress (Chowdhury et al., 2021), or as chronic, frequently asymptomatic and associated with biofilms that shield the pathogen from immune clearance (Atiencia-Carrera et al., 2022; Cabezas-Mera et al., 2023; Machado and Cerca, 2015). Biofilms create protected niches, such as granulomas or intracellular compartments, enabling persistence under reduced metabolic activity (Giacomodonato et al., 2022; Thakur et al., 2019), even in the presence of host defenses (Gunn et al., 2014).
Despite strong evidence implicating biofilms in outbreaks, directly linking them to specific contamination events remains challenging due to technical limitations in detecting sessile cells during routine surveillance. Control of Salmonella is particularly difficult because of its ability to persist and spread through multiple pathways (Gunn et al., 2014). Consequently, effective detection is critical in the food industry to prevent outbreaks. Many countries enforce zero-tolerance policies for Salmonella in food products, underscoring the urgent need for rapid and reliable testing methods (Gunn et al., 2014).
Although microbial culture remains the gold standard because of its low cost and proven reliability, it is a slow process, often taking up to five days, which limits its practicality for perishable products and making it impractical for perishable items (Ehuwa et al., 2021). To address this, a variety of rapid detection technologies have been developed, including immunoassays, polymerase chain reaction (PCR), mass spectrometry, optical and electrochemical biosensors, DNA aptamers, and bacteriophage-based tools (Lin et al., 2020). However, many of these emerging approaches, such as mass spectrometry and whole-genome sequencing, are costly and require further validation due to their complexity, which limits their widespread use in routine surveillance programs (Awang et al., 2021; Bergamo and Gandra, 2021; Wang et al., 2018).
Figure 5 and Table 4 summarize these methods, outlining their advantages and limitations. Enhancing detection capacity is key to reducing the global burden of Salmonella-related illnesses and improving food safety.
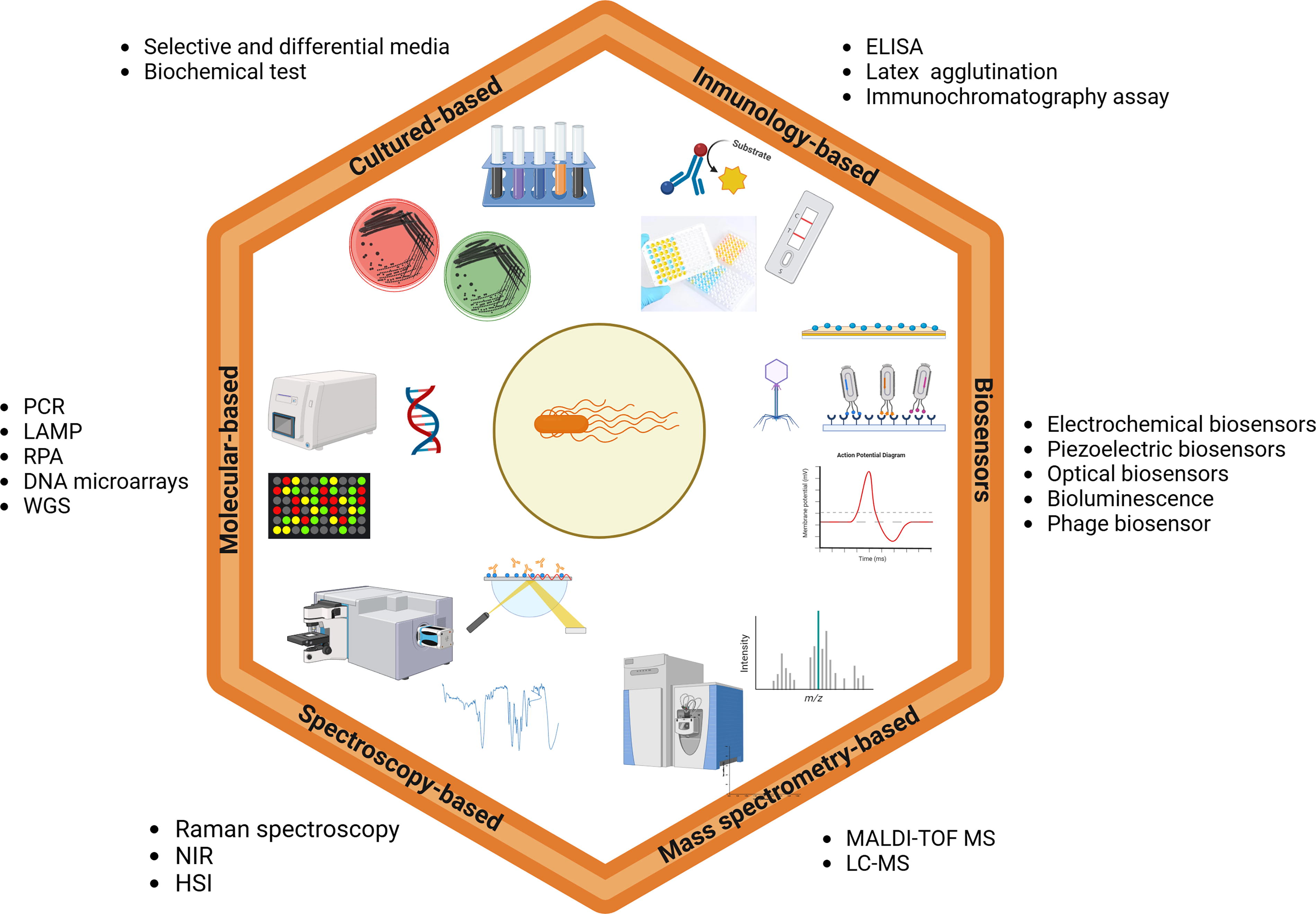
Overview of the principal detection methods for Salmonella. ELISA, enzyme-linked immunosorbent assay; HIS, hyperspectral imaging; LAMP: loop-mediated isothermal amplification; LC-MS, liquid chromatography-mass spectrometry; MALDI-TOF MS: matrix-assisted laser desorption ionization-time-of-flight mass spectrometry; NIR, near-infrared spectroscopy; PCR, polymerase chain reaction; RPA, recombinase polymerase amplification; WGS: whole genome sequencing. (Available from: https://app.biorender.com accessed on June 11, 2024).
Comparative Overview of Salmonella Detection Methods, Summarizing Their Principles, Advantages, Limitations, and Applications Across Food, Environmental, and Clinical Samples
ELISA, enzyme-linked immunosorbent assay; HSI, hyperspectral imagi; hilA: hyperinvasive locus A gene; InvA, invasion gene; LAMP, loop-mediated isothermal amplification; LC-MS, liquid chromatography-mass spectrometry; MALDI-TOF MS, Matrix-assisted laser desorption ionization-time-of-flight mass spectrometry; NIR, Near-Infrared; OmpF, outer membrane porin F gene; PCR, polymerase chain reaction; RPA, recombinase polymerase amplification; ttr, tetrathionate reductase gene; spvC, virulence plasmid gene; WGS, whole genome sequencing.
Standard and Alternative Procedures to Fight Salmonella Colonization
Controlling Salmonella biofilms presents both challenges and opportunities, requiring advances in diagnostics, prevention, and treatment through integrated approaches. Preventive measures primarily aim to reduce the likelihood of Salmonella attachment and biofilm initiation, whereas therapeutic approaches target the disruption or eradication of biofilms once established. Faster and more accurate detection methods, such as biosensors and real-time monitoring, are crucial for early intervention, though affordability remains a limitation. Improving diagnostic sensitivity is particularly important for detecting Salmonella at low concentrations or within mature biofilms (Esteban Florez et al., 2016; Hemdan et al., 2023; Kırmusaoğlu, 2019; Sanchez et al., 2013).
Preventive measures
Preventive strategies aim to limit Salmonella persistence and reduce the risk of contamination in food production and clinical settings. Developing antimicrobial or anti-adhesive coatings for food processing environments represents one promising approach to minimizing bacterial colonization and biofilm formation (Davies and Breslin, 2003). Similarly, routine environmental monitoring, improved sanitation practices, and the use of biosensors for early detection can significantly reduce the likelihood of recurrent contamination. Integrating these preventive tools within a One Health framework, which links human, animal, and environmental health, further strengthens surveillance and control of Salmonella transmission (Andino and Hanning, 2015; Jones et al., 2021; Li et al., 2023; Sáenz et al., 2022).
Therapeutic interventions
In parallel, therapeutic approaches are being developed to target established biofilms and antibiotic-resistant strains. Enzymes capable of degrading the ECM, used in combination with bacteriophage-based therapies, represent innovative alternatives to conventional biocides (Dadkhah et al., 2025; Pérez-Lavalle et al., 2025). The growing challenge of antibiotic resistance also underscores the need for novel interventions, such as combining antibiotics with phytochemicals to enhance antimicrobial efficacy and delay the development of resistance. Several natural compounds, including essential oils (e.g., thymol, carvacrol, eugenol) and plant-derived polyphenols (e.g., catechins, quercetin, resveratrol), have demonstrated synergistic effects when used in combination with conventional antibiotics. These molecules act through diverse mechanisms, such as disrupting bacterial membranes, interfering with quorum-sensing pathways, inhibiting efflux pumps, and generating oxidative stress. By targeting multiple bacterial processes simultaneously, phytochemicals not only potentiate antibiotic activity but also reduce the likelihood of resistance emerging, making them promising candidates for integrated antimicrobial strategies (Cabezas‐Mera et al., 2024; Cabezas-Mera et al., 2023; Fernandez-Soto et al., 2023; Jara-Medina et al., 2024; Machado et al., 2023).
Gene-editing tools, particularly CRISPR/Cas systems, offer a powerful therapeutic avenue for modulating virulence and disrupting biofilm integrity (Sharma et al., 2022; Sheng et al., 2023). By designing sequence-specific guide RNAs, Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR) platforms can silence or eliminate regulators such as spvB, bcsA, or quorum-sensing genes, thereby impairing ECM structure and resilience (Basit et al., 2022). CRISPR interference (CRISPRi) provides an additional advantage by enabling reversible gene repression without inducing double-strand DNA breaks, offering a safer alternative for microbial population control (Senevirathne et al., 2021). Efforts to improve delivery, using bacteriophage vectors or nanoparticle carriers, aim to overcome the protective barriers of biofilm-embedded cells (El-Demerdash et al., 2023).
Nevertheless, the practical implementation of CRISPR- and phage-based therapies faces considerable challenges. Efficient delivery systems, production logistics, and cold-chain requirements remain major barriers in low-resource settings (Mayorga-Ramos et al., 2023). Furthermore, the absence of harmonized regulatory frameworks, along with societal concerns about advanced genetic technologies in food and agriculture, continues to delay broader approval and acceptance (Boubakri, 2023). Addressing these barriers requires not only technological innovation but also coordinated policy development, intersectoral collaboration, and investment in local infrastructure to ensure equitable access to these emerging tools.
Looking forward
Global collaboration will be essential to achieve equitable implementation of biofilm control strategies, particularly in low-resource regions. Strengthening public policy, epidemiological surveillance, and cross-sector coordination will be key to managing Salmonella as a persistent public health threat. Finally, climate change may influence biofilm formation by altering environmental conditions that favor bacterial persistence, highlighting the need for ongoing monitoring and adaptable control measures. Meeting these challenges through innovation, regulation, and education will be central to effectively managing Salmonella biofilms in the future (Kang et al., 2023; Mishra et al., 2020; Roy et al., 2018).
Future Perspectives and Final Remarks
Animal production systems, particularly poultry, represent a significant reservoir for Salmonella, but they are not the only source (Milanov et al., 2017). The bacterium’s ability to persist in a wide range of hosts and environments poses a multifaceted challenge to food safety (Silva and López-Moreno, 2012). Emerging strains with enhanced virulence, stress tolerance, and resistance to disinfectants are increasingly detected across different livestock species, including swine and cattle (Cadena et al., 2019). Biosecurity and hygiene remain fundamental for mitigating contamination risks across the entire animal production sector. Although current vaccines (especially in poultry) provide partial protection, novel formulations, such as trivalent and recombinant vaccines, are under development with a focus on prevalent serovars (Dórea et al., 2010; Huberman et al., 2022). Beyond livestock, the agricultural use of untreated or poorly composted animal manure can introduce Salmonella into the environment and onto crops, particularly leafy greens and other fresh produce, which have frequently been implicated in outbreaks. The bacterium’s capacity to survive for extended periods in soil, water, and on inanimate surfaces facilitates cross-contamination along the food chain (Ehuwa et al., 2021). Processing environments, equipment, and improper food handling further exacerbate transmission risks. Epidemiological data confirm that Salmonella outbreaks have been linked to a diverse range of food matrices, including eggs, meat, dairy products, spices, fruits, vegetables, and processed foods, as summarized in Table 3.
Controlling Salmonella remains a complex challenge that requires an integrative approach rooted in the understanding of its physiology, biofilm-forming capacity, AMR, and environmental persistence. Current strategies have improved detection sensitivity and pathogen surveillance, particularly with the advent of molecular and genomic tools (Bergamo and Gandra, 2021; Paniel and Noguer, 2019). However, several limitations persist that hinder effective control across the food chain and clinical settings.
One of the most critical barriers is the limited ability to detect and eliminate Salmonella in its biofilm-associated state (Merino et al., 2019). These structured communities confer significant protection against environmental stressors and disinfectants, allowing Salmonella to persist in food processing environments and natural ecosystems. While the bacterium’s planktonic physiology is well-characterized, its behavior within biofilms, especially under environmental stress or sublethal antimicrobial exposure, remains underexplored and requires further investigation (Paniel and Noguer, 2019).
The growing prevalence of multidrug-resistant Salmonella strains exacerbates the public health burden, demanding urgent action to reduce antibiotic misuse and promote the development of alternative interventions (Almuzaini, 2023). CRISPR-Cas technologies represent a promising frontier in this context (Charpentier and Marraffini, 2014; Greene, 2018; Mayorga-Ramos et al., 2023). Their potential for targeted disruption of biofilm-related genes offers an innovative approach to weaken Salmonella persistence mechanisms. However, their practical application is still in the early stages and faces challenges related to delivery systems, biosafety, and regulation (Boubakri, 2023).
Future research should focus on the development of biofilm-specific detection systems, scalable antibiofilm agents, and combined strategies that integrate natural antimicrobials, bacteriophage therapy, and precision genome editing (Boyd and Brüssow, 2002; Levine et al., 1975; Pottker et al., 2024). The implementation of these tools must be guided by the One Health framework, which acknowledges the interconnectedness of human, animal, and environmental health. Achieving meaningful control of Salmonella will require coordinated surveillance, investment in infrastructure for real-time pathogen monitoring, and education campaigns to promote responsible antibiotic use and hygiene across sectors.
Conclusions
Salmonella virulence factors, such as its ability to form biofilms and establish reservoirs in diverse environments, are key to its persistence and spread. These biofilms confer significant protection against physical, chemical, and immune-mediated stressors, rendering the bacteria markedly more resistant to conventional antimicrobial treatments and sanitation measures. This resilience poses a major barrier to both the effective diagnosis and control of Salmonella infections. Detecting Salmonella within biofilm structures remains especially challenging due to the low metabolic activity and limited abundance of the embedded cells, which often fall below conventional detection thresholds.
Future efforts must prioritize the integration of innovative biotechnologies, such as CRISPR/Cas9, which could offer new strategies to intervene in biofilm formation mechanisms and improve diagnostic accuracy, as well as enhance the control of Salmonella. The development of gene-editing-based approaches promises to be a key tool in the fight against this bacterium, enabling more targeted and effective solutions for its eradication and prevention.
Acknowledgements
Special recognition deserves all colleagues of the Biofilm Research Group (BRG) at the Microbiology Institute of Universidad San Francisco de Quito (IM-USFQ), COCIBA, and the Research Office of USFQ for their support in this study.
Footnotes
Authors’ Contributions
Conceptualization: J.C.C.V., S.Z.-M., and A.M. Methodology: V.P.E.E., M.G.V.C., J.C.C.V., S.Z.M., and A.M. Validation: S.Z.-M. and A.M. Formal analysis and investigation: V.P.E.-E., M.G.V.-C., J.C.C.-V., S.Z.-M., and A.M. Resources: J.C.C.-V., S.Z.-M., and A.M. Data curation: V.P.E.-E., S.Z.-M., and A.M. Writing—original draft preparation: V.P.E.-E. and M.G.V.-C. Writing—review and editing: S.Z.-M. and A.M. Supervision: S.Z.-M. and A.M. Project administration and funding: J.C.C.-V., S.Z.-M., and A.M. All authors contributed to the article and approved the submitted version.
Data Availability Statement
The authors confirm that the data supporting the findings of the present review are available within the cited articles.
Disclosure Statement
The authors declare that they have no financial or personal relationships that could be perceived as potential conflicts of interest. No interests to disclose.
Funding Information
This work was supported by COCIBA Research Grants under the following projects: Project ID: 17357, titled “Alternative Approaches for Eliminating Biofilms”, and Project ID: 16801, titled “Characterization of Single and Mixed Biofilms”, both awarded to António Machado; Project ID: 17827, titled “Fagoterapia como Alternativa para el Control de Salmonella enterica en Superficies Vivas e Inertes para Utilización en la Producción Avícola en Ecuador”, awarded to Sonia Zapata; and Project ID: 23248, titled “Desarrollo y Caracterización de Sistemas CRISPR/Cas para la Inhibición de Formación de Biopelículas en Microorganismos Patógenos”, awarded to Juan Carlos Collantes Vela.