
Abstract
Knowledge of resident bacteria present within animal food manufacturing facilities, as well as the understanding of antimicrobial resistance genes (ARGs), can help inform cleaning and sanitation practices to reduce microbial risks for the animal and human food chains. The goal of this study was to characterize resident bacteria previously isolated in selected swine feed mills within the Midwest region of the United States at multiple seasonal timepoints. About 121 resident bacterial isolates were identified and classified into six genera, consisting of 68 Enterobacter, 34 Citrobacter, four Cronobacter, eight Klebsiella, three Proteus, and four Pseudomonas, in addition to 30 Escherichia coli and 33 Salmonella characterized previously. Among the 121 isolates discussed within the current work, the majority (48; 39.7%) came from environmental samples taken from either non-feed contact surfaces such as floors and feed contact surfaces (38; 31.4%) or transient surfaces such as brooms and workers’ shoes (20; 16.5%), and only 12 samples (9.9%) came directly from finished feed. A range of ARGs and metal tolerance genes (MTGs) were identified. This research provides an initial framework to understand the diversity of resident flora in feed mill facilities in the Midwest region and identify areas to focus on for housekeeping and sanitization within mills. This information is critical to developing feed safety strategies and preventing antimicrobial resistance spread in the farm-to-table continuum.
Introduction
Understanding the diversity of resident bacteria in the animal food processing environment during operation and after cleaning and sanitation is important to assess the food safety risks associated with animal food commodities. Several studies indicated that the survival of foodborne pathogens is heavily influenced by non-pathogenic resident bacteria (Giaouris et al, 2015). Resident bacteria are usually isolated from non-selective media and defined as the bacterial community that persists over time in production facilities (Møretrø and Langsrud, 2017). Enterobacterales are usually used as indicators for hygiene and sanitation (Marriott et al., 2018). A recent survey, which sampled 647 pet food and 378 animal feed samples, reported that this indicator group was widely present (Ge et al, 2020). These bacteria may also have a role in antimicrobial resistance (AMR) spread in animal food-related commodities, which is a growing and pressing concern for both human and animal health (World Health Organization (WHO), 2015). While characterization of antimicrobial resistance genes (ARGs) and metal tolerance genes (MTGs) has been done on specific bacterial pathogens present within feed mills and ingredients (Davies and Wales, 2010; Torres et al, 2011; Burns et al, 2015; Munoz et al, 2021; Magossi et al, 2020), little is known about the overarching bacterial presence and associated ARGs and MTGs within facilities. Therefore, in this study, we characterized resident bacteria (mostly Enterobacterales) isolated previously from feed mills located in the Midwest region of the United States with the goal of assessing the food safety risks associated with animal food commodities.
Material and Methods
Sample collection and isolation
This study is an extension of our previous survey conducted in six commercial swine feed mills located in the Midwest region (Iowa, Missouri, Kansas, and Oklahoma) of the United States during three different seasons: fall (October of 2018), late winter/early spring (March of 2019), and summer (June of 2019) (Magossi et al, 2020). The selection of the mills was based on location and the feed produced. Sampling sites were selected from areas of high pedestrian traffic, ingredient processing, and dust accumulation, following production flow (Magossi et al., 2019). A total of 405 samples were collected across a range of sampling sites, including finished feed, floors, brooms, and feed manufacturing equipment. Each site was analyzed for the presence of pathogens and commensals. From all the presumptive positive samples, at least two colonies were selected for further investigation (Schwan et al, 2023). Isolates were stored in Trypticase Soy Broth (TSB; Hardy Diagnostics, Santa Maria, CA, USA) containing glycerol at −80°C until analysis. On day 1 of experiments, isolates were retrieved from −80°C and grown on Trypticase Soy Agar (TSA, Hardy Diagnostics, Santa Maria, CA, USA) or Blood Agar (Hardy Diagnostics, Santa Maria, CA, USA) at 37°C overnight.
Bacterial speciation
The VITEK 2 Compact system (bioMérieux, Marcy l’Etoile, France) or matrix-assisted laser desorption/ionization time-of-flight (MALDI-TOF) mass spectrometry on the MALDI Biotyper (Bruker, Billerica, MA) was used for the identification of bacterial genera and species following the manufacturer’s instructions.
Antimicrobial susceptibility testing (AST)
AST against 14 antimicrobial agents was performed by broth microdilution (Schwan et al, 2023). The MICs were interpreted using the CLSI standards and NARMS breakpoints valid for Enterobacterales (Clinical and Laboratory Standards Institute, 2019; Food and Drug Administration, 2015). Multidrug resistance (MDR) was defined as resistance to ≥3 antimicrobial classes.
WGS characterization
DNA extraction, library preparation, sequencing, and in silico analysis were performed as previously described (Domesle et al, 2021) with some modifications. Genomic DNAs were extracted using the Qiagen DNeasy Blood & Tissue kit (Germantown, MD, USA). Nextera XT Library Preparation kit (Illumina, San Diego, CA, USA) was used for preparing paired-end libraries, and WGS was carried out on the MiSeq sequencer, using a 2 × 300-cycle MiSeq reagent V3 kit (Illumina). Trimming and de novo assemblies were obtained with Shovill version 9 (https://github.com/tseemann/shovill), available in the GalaxyTrakr pipeline (http://galaxytrakr.org/) with default parameters (Zhang et al., 2019).
Detection of ARGs and MTGs
ARGs and MTGs were identified using AMRFinderPlus v3.12.8 (https://www.ncbi.nlm.nih.gov/pathogens/antimicrobial-resistance/AMRFinder/) with default parameters (Feldgarden et al., 2021).
Data availability
The accession numbers and additional genomic information for the isolates can be found under the BioProject number PRJNA561052 at the NCBI, National Library of Medicine.
This study was approved by the KSU Institutional Review Board.
Results and Discussion
Bacterial species characterization
Out of 405 samples, a total of 212 isolates were recovered, which were classified into 16 genera by WGS. Of these 212 isolates, 28 isolates were not assembled as they belonged to uncommon Enterobacterales, including Kosakonia cowanii, Leclercia adecarboxylata, Lelliottia amnigena, Pantoea spp., and Rahnella aquatilis. Isolates belonging to E. coli (n = 30) and Salmonella (n = 33) were characterized in an earlier report (Schwan et al., 2023). In the present study, we further investigated the remaining 121 isolates, which consisted of 34 Citrobacter, 4 Cronobacter, 68 Enterobacter, 8 Klebsiella, 3 Proteus, and 4 Pseudomonas.
Isolate location profile within animal feed manufacturing facilities
The isolates obtained in this study (n = 121) were primarily found through environmental swabbing, with only 12 derived from finished feed (Tables 1–6). Most of the environmental samples were taken on hard, stationary surfaces, with slightly more isolates coming from non-feed contact surfaces like floors than those on feed contact surfaces (38/121), such as receiving grates, fat inlet valves, pellet mills, and load-out equipment within the different facilities. Twenty (20/121) isolates were obtained from transient surfaces like worker shoes and brooms (Tables 1–6).
Sample Metadata of the Six Bacterial Genus Isolated in This Study (n = 121)
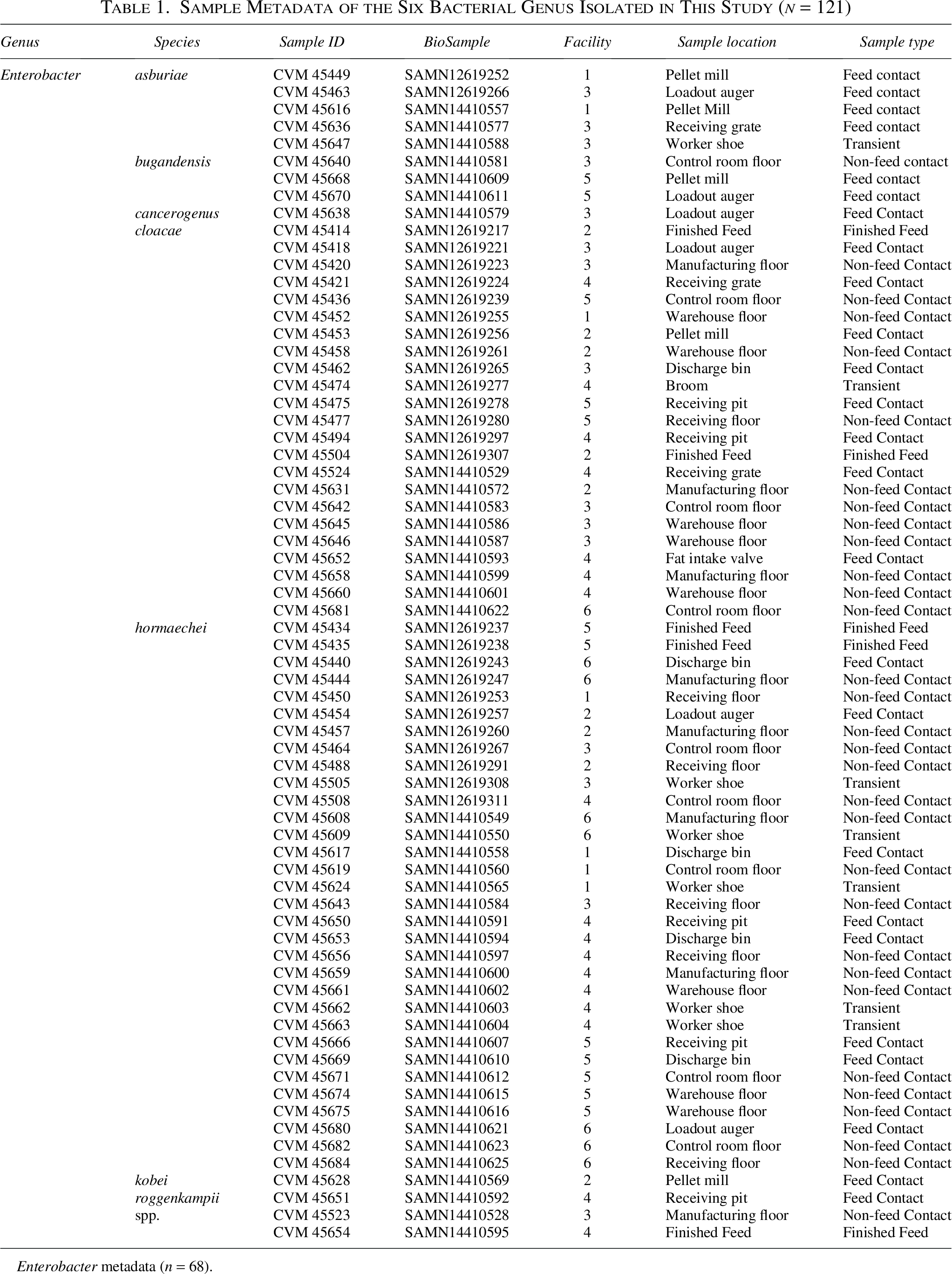
Enterobacter metadata (n = 68).
Sample Metadata of the Six Bacterial Genus Isolated in This Study (n = 121)
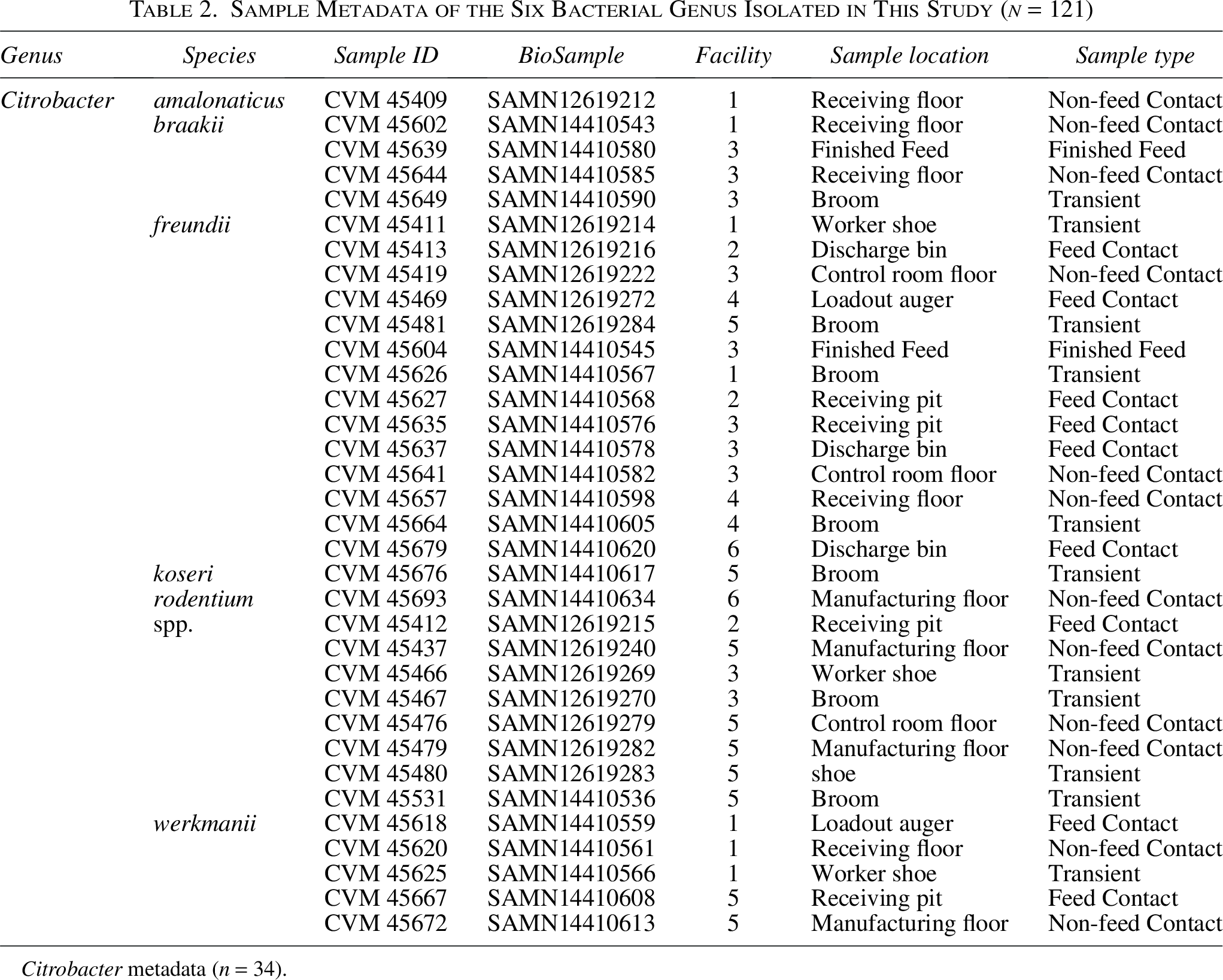
Citrobacter metadata (n = 34).
Sample Metadata of the Six Bacterial Genus Isolated in This Study (n = 121)
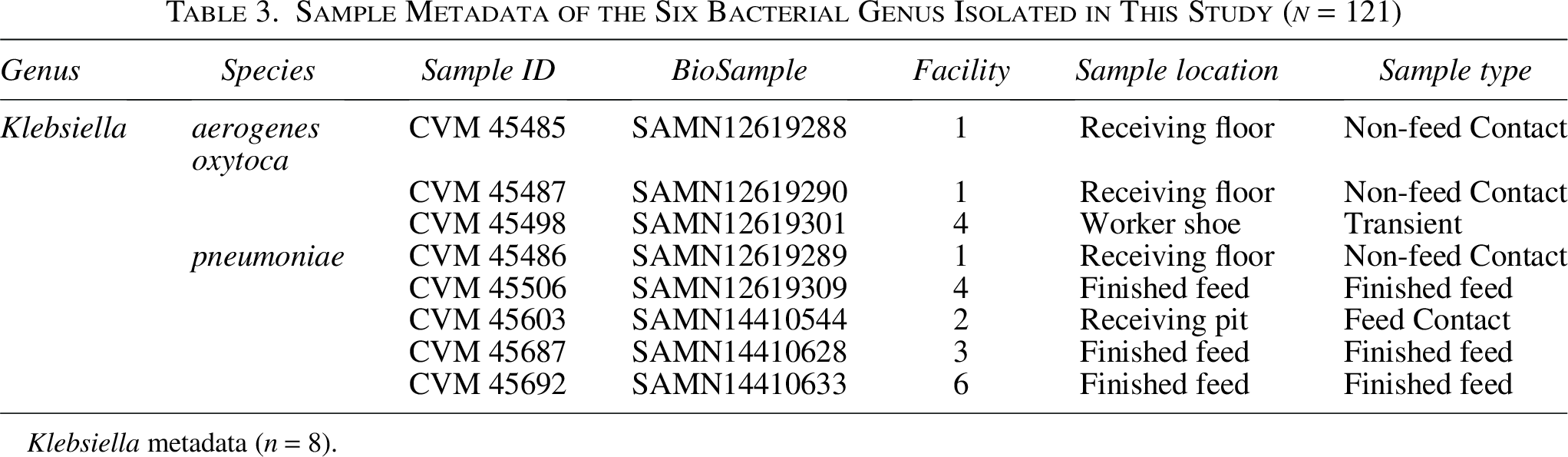
Klebsiella metadata (n = 8).
Sample Metadata of the Six Bacterial Genus Isolated in This Study (n = 121)

Crobobacter metadata (n = 4).
Sample Metadata of the Six Bacterial Genus Isolated in This Study (n = 121)
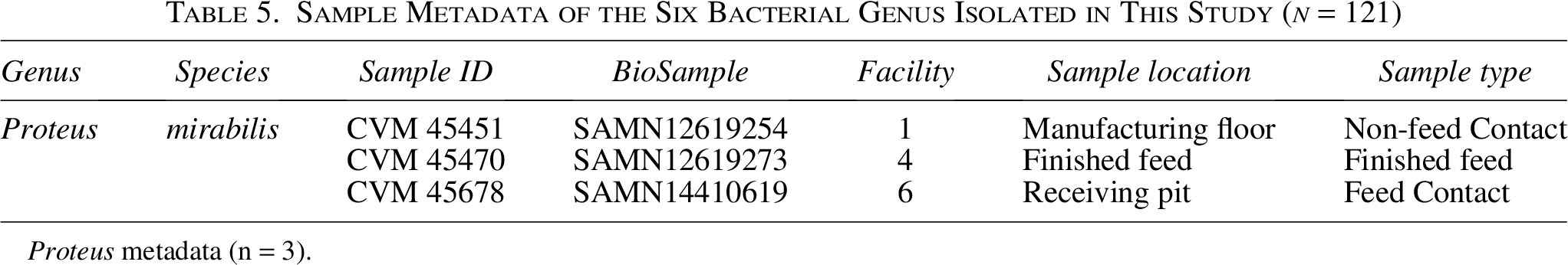
Proteus metadata (n = 3).
Sample Metadata of the Six Bacterial Genus Isolated in This Study (n = 121)

Pseudomonas metadata (n = 4).
Among the 68 Enterobacter isolates, five came from finished feed samples, with the remaining from environmental swabs. Most of the isolates were found on either non-feed contact surfaces (n = 3) or direct feed contact (n = 24), with only 7 from transient surfaces such as shoes or brooms. Six different Enterobacter species were identified as E. asburiae (5), E. bugandensis (3), E. cancerogenus (1), E. cloacae (22), E. hormaechei (32), E. kobei (1), and E. roggenkampii (1), with two isolates unable to be speciated. Fifty-nine isolates identified were grouped into the Enterobacter cloacae complex (ECC), a group of six species able to produce human infections (Mezzatesta et al, 2012).
Among the 34 Citrobacter isolates, most were from environmental swabs, including non-feed contact surfaces (n = 12), transient surfaces (n = 11), and feed contact surfaces (n = 9). Two isolates were obtained from finished feed samples. There were 6 different species identified C. amalonaticus (1), C. braakii (4), Citrobacter freundii (14), C. koseri (1), C. rodentium (1), and C. werkmanii (5), with an additional 8 isolates that could not be speciated. A total of 25 isolates belong to the C. freundii complex (Brenner et al., 1993). Citrobacter spp. are found in soil, water, and intestinal tracts of animals and humans, and some species have been documented to cause infections in humans (Ranjan and Ranjan, 2013).
Four Cronobacter species were identified within the samples taken for this study, one coming from finished feed, with the other three isolated from environmental samples taken within the receiving area of two different facilities. Of the environmental samples, two (2/4) were from the same facility, but were identified at different sampling timepoints and sites (ingredient receiving pit vs receiving area floor). Cronobacter spp. can be found naturally within a variety of environments and cause severe human illnesses (Healy et al, 2010).
Three species of Klebsiella (n = 8) were identified, which were K. pneumoniae (5), K. oxytoca (2), and K. aerogenes (1). Three of the K. pneumoniae isolates were obtained from finished feed (3/5), with the other two (2/5) found on surfaces within raw ingredient receiving areas (1 feed contact, 1 non-feed contact). Two K. oxytoca (2/3) isolates were found on workers’ shoes; the remaining one was found in a non-feed contact surface sample. One K. aerogenes was obtained from a non-feed contact surface within an ingredient receiving area. Klebsiella spp. are commonly found in animal and human intestinal tracts, as well as plants, soil, and water (Bagley, 1985), and have been previously documented within feed samples (Zadoks et al, 2011).
All three Proteus isolates belonged to P. mirabilis and were identified in separate facilities, with one isolate coming from finished feed (1), one from a feed contact surface (1), and one from a non-feed contact surface (1/3). This bacterium is considered commensal within the gastrointestinal tracts, but it can also be found in water and soil. Foodborne illness due to P. mirabilis consumption in contaminated meat and seafood has been documented (Gong et al, 2019; Ma et al, 2022).
Of the four Pseudomonas isolates, three were P. aeruginosa, and one was P. putida. Three of the four isolates were identified from environmental swabs on non-feed contact surfaces, while one P. aeruginosa was identified from a feed contact surface. Pseudomonas spp. are commonly found in soil and water and can also cause infection in humans and plants (Raposo et al., 2017).
Phenotypic AST and presence of MDR
None of the 121 isolates were resistant to GEN (aminoglycosides), MER (cefoxitin; carbapenems), AXO (cefriaxone; cephems), COT (cotrimoxazole; polymyxin), ciprofloxacin (CIP), or NAL (nalidixic acid; quinolones). As shown in Table 7, resistance was observed to FOX (cefoxitin; n = 85), ampicillin (AMP; n = 81), amoxicillin-clavulanic acid (AMC; n = 75), tetracycline (TET; n = 40), streptomycin (STR; n = 17), sulfisoxazole (FIS; n = 5), azithromycin (AZI; n = 5), and chloramphenicol (CHL; n = 4).
Resistance Profiles of Bacteria from Swine Feed Production Environments

X indicates resistance to antimicrobial. NA means no MIC breakpoints for Pseudomonas to that antimicrobial.
AMC, amoxicillin-clavulanic acid; AMP, ampicillin; AZI, azithromycin; CHL, chloramphenicol; CIP, ciprofloxacin; FIS, sulfisoxazole; STR, streptomycin; TET, tetracycline.
Because of the production of AmpC β-lactamase, Enterobacter are intrinsically resistant to a variety of β-lactams (Bouza and Cercenado, 2002), which was mirrored within the phenotypic susceptibility results of this study. Of the 68 Enterobacter isolates, only five were not resistant to any of the three antibiotics listed previously. The most common resistance indicated outside of AMC, AMP, and FOX for Enterobacter isolates was TET. Four E. cloacae isolates were found to exhibit MDR, resistant to aminoglycosides, β-lactams, cephems, penicillin, and tetracycline. Only one Enterobacter isolated was resistant to macrolides.
Among the Citrobacter isolates, 21 were FOX resistant, 17 AMP resistant, eight TET resistant, and seven CIP resistant. Additionally, five isolates were STR-resistant, two FIS-resistant, and one resistant to AMC. Almost all the C. freundii isolates (11/14) were AMC-resistant. None of the Citrobacter isolates were found to have MDR. Citrobacter spp. are considered intrinsically resistant to cephalosporins (Liu et al, 2017; Pepperell et al, 2002) and ampicillin (Wanger et al, 2017). Specific species, such as C. freundii, are intrinsically resistant to ampicillin-sulbactam and cephalosporins (Wanger et al, 2017).
Cronobacter spp. (n = 4) have been found to have intrinsic resistance to antibiotics such as AMP and FOX (Donnenberg, 2015). Other studies have shown a high level of resistance to AMC (Pakbin et al., 2022). Three of the isolates from this study were resistant to both AMC and FOX; two of those isolates were also resistant to AMP. Additionally, CIP and TET resistance were found in one separate isolate, respectively. None of the Cronobacter spp. Isolates were MDR, and one was resistant to all antimicrobials tested.
Klebsiella spp. are intrinsically resistant to penicillin (Bouza and Cercenado, 2002). Of the nine Klebsiella isolates from this study, six were AMP-resistant. Two isolates were resistant to STR and TET, and resistance to AMC, CIP, FIS, and FOX was found in one individual isolate. Two isolates (K. oxytoca and K. pneumoniae) were found to be MDR. The K. oxytoca isolate showed resistance to CIP, STR, and TET, while K. pneumoniae was resistant to STR, FIS, and TET.
All Proteus mirabilis isolates in this study were resistant to TET, which is considered intrinsic resistance (Danilo de Oliveira et al, 2021; Mbehang Nguema et al, 2021). Additionally, all isolates showed resistance to AZI, and one isolate was also resistant to STR. None of the three isolates exhibited MDR.
Pseudomonas is well regarded as having significant amounts of intrinsic resistance to a variety of antimicrobials and is generally adept at acquiring resistance (Breidenstein et al, 2011; Pang et al, 2019). All 4 Pseudomonas isolates in this study were intrinsically resistant to AMC, AMP, AXO, AZI, CHL, CIP, COT, NAL, and STR; additionally, three were resistant to TET, two to FIS, and one to MER.
Presence of ARGs
Overall, a wide range of ARGs were identified among the isolates collected in this study (Fig. 1). Phenotypic antimicrobial resistance testing showed overall good agreement with genotypic predictions (e.g., identifications of corresponding ARGs among isolates demonstrating phenotypic resistance), though a few discrepancies were noted (i.e., one Citrobacter and one Enterobacter to AMP, all five Citrobacter, Klebsiella, and Pseudomonas isolates to AZI and four Pseudomonas isolates to CHL, two Pseudomonas isolates to FIS, three Citrobacter isolates to STR, and one Enterobacter to TET), potentially due to previously uncharacterized ARGs, overexpression of efflux pumps, or membrane permeability barriers.
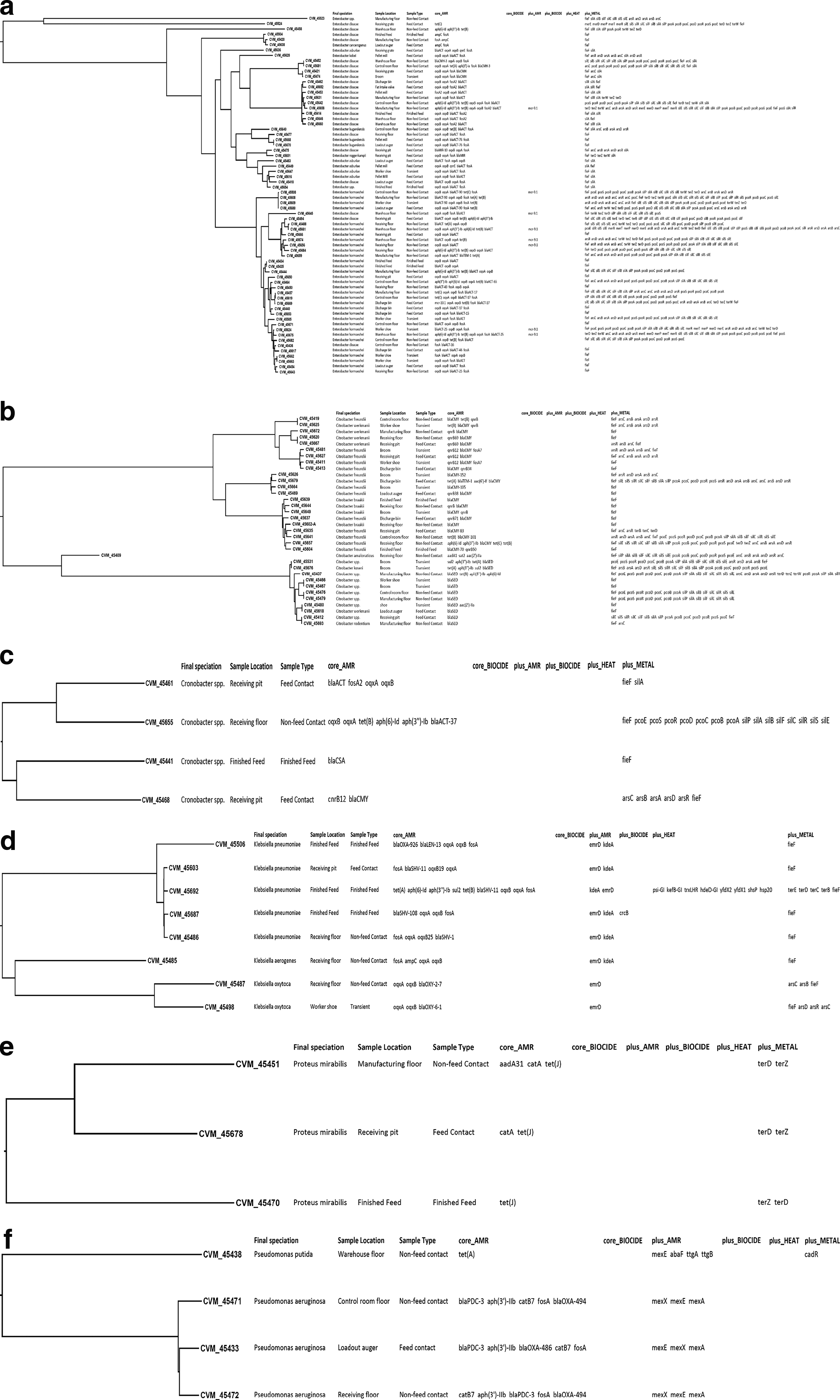
Phylogenetic tree by genus showing isolate ID, Final speciation, sample location and type, core AMR, and Metal:
Almost all Enterobacter spp. isolates contained at least one β-lactamase gene (64/68), and 51 contained at least one fosfomycin resistance gene. Tetracycline resistance was found in 22 isolates, and aminoglycoside resistance in 10. All Citrobacter spp. isolates (34) were found to have at least one ARG, and 32 contained a gene indicative of β-lactam resistance. The second most common gene identified was qnrB, with 14 indicating resistance. Additionally, 7 isolates were found to have aminoglycoside resistance, and 8 to tetracycline.
All Cronobacter spp contained a gene indicative of β-lactam resistance, and three isolates contained additional resistance genes: two for phenicol/quinolone resistance, and one having aminoglycoside, fosfomycin, quinolone, and tetracycline resistance genes. The nine different Klebsiella spp. isolates contained β-lactamase genes and expressed also oqxA and/or oqxB, indicating phenicol and quinolone resistance (Lam et al, 2021). Fosfomycin resistance from fosA was found in all but one K. pneumoniae, and one sample also expressed aminoclycoside, sulfonamide, and tetracycline resistance.
All Pseudomonas aeruginosa isolates contained three separate genes indicative of increased efflux pump activity (mexA, mexE, mexX) and resistance to phenicol (catB7), fosfomycin (fosA), aminoglycosides [aph(3′)-Iib)], and multiple genes for β-lactams (genes within the blaOXA family, as well as blaPDC-3). Only the Pseudomonas putida isolate contained the tet(A) gene, indicative of tetracycline resistance.
All three Proteus mirabilis isolates showed intrinsic tetracycline resistance (Danilo de Oliveira et al, 2021). Two isolates also contained catA, indicating phenicol resistance, with one also containing aadA31, indicative of resistance to aminoglycosides.
Our study showed a high prevalence of β-lactamase genes in Pseudomonas, Klebsiella spp., Citrobacter, and Cronobacter spp. Similarly to previous studies (Davies and Wales, 2019). Several of our observations highlight the potential of ARG transmission through horizontal transfer, posing significant risks for livestock and human health (Frost et al., 2005; Van Hoek et al., 2011), such as the detection of aminoglycoside resistance genes, typically associated with clinical or environmental Salmonella and E. coli strains (Cabal et al., 2022), and the presence of fosA2, commonly linked to Cronobacter in clinical settings (Sapkota et al., 2011). Furthermore, the Enterobacter cloacae complex identified in our study has been well documented in causing infections in both swine and humans, underscoring the clinical relevance of ARG reservoirs in feed mill environments (Annavajhala et al., 2019; Mezzatesta et al., 2012).
Presence of MTGs
The presence of at least one MTG was identified within each isolate analyzed in this study, excluding Pseudomonas isolates (Fig. 1). Overall, 59 isolates contained genes indicative of tolerance to copper/silver; individually, genes responsible for copper tolerance were found in 37 isolates and silver in 38 isolates. Arsenic tolerance genes were found in 43 isolates. Research has indicated that the presence of MTGs can be indicative of selective pressures being applied to bacteria from the environment, which may lead to co-selection of genes that confer AMR (Nguyen et al, 2023).
Enterobacter spp. Isolates displayed joint copper/silver tolerance within 47 isolates. Standalone tolerance to silver was indicated within 27 isolates, as well as copper in 26 isolates. Arsenic tolerance was indicated in 24 isolates. Genes indicating tolerance to tellurium were found in 19 isolates, and mercury tolerance was detected in 4 isolates. The most prevalent MTGs are present within the Citrobacter spp. Isolates were isolated to arsenic, with 15 isolates containing at least one gene. Copper/silver tolerance genes were present in 10 isolates, with the same isolates also having genes indicative of standalone silver tolerance and nine copper tolerance. A small portion of the isolates (3) contained genes indicative of tellurium tolerance. Of the four Cronobacter spp. Two contained at least one gene indicative of copper/silver tolerance, with one also containing genes for standalone copper and silver tolerance. One isolate contained an arsenic tolerance gene. All Klebsiella oxytoca isolates were tolerant to arsenic, and one Klebsiella pneumoniae contained a gene indicative of tellurium tolerance. All three Proteus mirabilis contained terD and terZ genes for tellurium tolerance. While the widespread presence of copper, silver, and arsenic tolerance genes aligns with documented selective pressures from metal use in animal settings (Nguyen et al., 2023; Yu et al., 2017), the detection of tellurium tolerance genes (terD, terZ) in P. mirabilis, K. pneumoniae, and Enterobacter spp. isolated in our study represents a novel finding, as such genes have been infrequently reported in feed-associated isolates (Toptchieva et al., 2003). A study on selected MDR clinical isolates sourced within a hospital observed the presence of the fieF gene, an efflux modulator, within Enterobacterales (Yaikhan et al, 2024), as we did in our samples. Furthermore, the co-occurrence of MTGs and ARGs raises concerns about co-selection, whereby exposure to metals may promote the persistence and transfer of ARGs within bacterial communities able to use horizontal gene and transposons to exchange genetic materials (Frost et al., 2005; Van Hoek et al., 2011) and highlighting the potential role of feed mill environments as reservoirs for resistance that could impact both livestock and human health.
Conclusion
This study proposes a novel and comprehensive map of microbial distribution and resistance within a swine feed mill, offering actionable insight into feed safety, such as the risk of cross-contamination (shoes and brooms), the importance of workers' training, and the need for longitudinal studies to monitor the persistence and evolution of AMR and MTG within feed production facilities. Nevertheless, the small sample size and the regional scope of the sampling constrain the extent to which these findings can be generalized, pointing to the importance of future studies with more diverse and representative populations. Still, the current study provides an important first step in documenting the diversity of resident flora in feed mill facilities in the Midwest region and identifies areas to focus on for housekeeping and sanitization within mills, offering insights that can guide and refine broader investigations.
Authors’ Contributions
C.J. and V.T. planned the experimental design and analyzed data. B.G., S.Y., K.D., and C.-H.H. were instrumental in data curation and analysis, susceptibility testing, whole-genome sequencing, and bioinformatic analysis. S.C.S. conducted experiments and drafted an article. All the authors contributed to the writing.
Footnotes
Acknowledgments
The authors want to thank Crystal Rice-Trujillo and Cong Li at the U.S. Food and Drug Administration for laboratory assistance and sequence submission.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.