
Abstract
Campylobacter jejuni and Campylobacter coli are leading causes of foodborne gastroenteritis worldwide, with poultry representing the primary reservoir for human infection. Understanding species-specific differences in virulence-associated features, antimicrobial resistance genes, and population structure is essential for assessing public health risks and guiding surveillance strategies. A total of 26 Campylobacter isolates (22 C. coli and 4 C. jejuni) were recovered in 2023 from poultry caeca collected from farms in north-western, central-western, and south-western Romania. Species identification and detection of the hcp gene, a marker of the type VI secretion system (T6SS), were performed using multiplex PCR. Multilocus sequence typing (MLST) based on seven housekeeping genes was performed to evaluate genetic diversity and population structure. The hcp gene was detected exclusively in C. jejuni isolates, indicating that it was detected only in C. jejuni within this collection of the primary T6SS component. Both species shared a broad set of core virulence genes related to motility, colonization, quorum sensing, adhesion, and surface polysaccharide biosynthesis, while notable differences were observed in accessory virulence gene profiles. Genes associated with antibiotic resistance, including the tetracycline resistance gene tet(O) and multiple efflux-related determinants, were frequently detected in both species. MLST analysis revealed high genetic diversity, with most isolates not assignable to previously defined sequence types, consistent with an open pan-genome and substantial genomic plasticity. These findings feature clear differences between C. jejuni and C. coli in the distribution of virulence markers, while validating the widespread presence of antimicrobial resistance genes in poultry-associated isolates. This study highlights the adaptive potential of Campylobacter and underscores the need to advance molecular surveillance to monitor virulence and antimicrobial resistance risks relevant to animal and human health.
Introduction
Campylobacter species (spp.) are among the most important zoonotic foodborne pathogens, with poultry identified as the primary route of transmission to humans, although other sources also contribute (Balta et al., 2021; Wei et al., 2024). During 2024, surveillance data from 26 EU countries documented over 168,000 confirmed campylobacteriosis cases, giving an overall EU notification rate of 55.3 per 100,000 inhabitants, representing an ≈12% increase compared with 2023 (European Food Safety A, European Centre for Disease P and Control, 2025). Not all countries followed this upward tendency; the lowest notification rates in 2024 were observed in Bulgaria, Cyprus, Poland, and Romania (≤6.6 per 100,000) (European Food Safety A, European Centre for Disease P and Control, 2025). Campylobacter occupies ecological niches shared with a wide range of other bacterial species, particularly in animal gastrointestinal systems and environmental reservoirs. This close coexistence creates opportunities for the exchange of genetic material, enabling Campylobacter to acquire antimicrobial resistance determinants through horizontal gene transfer (Zhang et al., 2023).
In this study, we aimed to characterize Campylobacter isolates collected in Romania from three poultry farms located in the north-western, central-western, and south-western areas of the country, to compare species-associated virulence and antimicrobial resistance genes, and to contextualize the isolates within the population structure. We combined PCR detection of hcp (a type VI and type IV secretion system [T6SS and T4SS] marker) with multilocus sequence typing (MLST) and Roary genome-based profiling of virulence- and resistance-associated loci, and we assessed gene-content diversity using pan-genome analysis.
Materials and Methods
Campylobacter isolates
All experimental procedures were performed in the microbiology laboratories of the University of Life Sciences “King Michael I” of Timișoara. The study analyzed 26 Campylobacter isolates collected during May-July 2023, including 22 Campylobacter coli and 4 Campylobacter jejuni strains, recovered from 30 poultry cecal samples in three counties, as shown in Figure 1. The geographic map was created using QGIS software (version 3.44.7) (Team QD, 2025). Isolation of Campylobacter spp. was carried out using Campylobacter blood-free selective agar base (Modified CCDA-Preston, Oxoid, Basingstoke, UK), followed by incubation at 41.5°C for 48 h under microaerophilic conditions (5% O2, 85% N2, 10% CO2) given by CampyGen Compact sachets (Thermo Fisher Scientific Oxoid Ltd., Basingstoke, UK). Isolation and identification were performed according to the procedures indicated in the ISO 10272-1:2017 for Campylobacter spp. Detection followed by multiplex PCR as previously described (Balta et al., 2021). Genomic DNA was extracted using the PureLink Genomic DNA Mini Kit (Invitrogen, Thermo Fisher Scientific, Carlsbad, CA, USA) in accordance with the manufacturer’s protocol. DNA concentration and purity were measured with a NanoDrop 1000 spectrophotometer. Species identification was confirmed by multiplex PCR according to the protocol described by Wang et al. (2002), using the primers listed in included in Supplementary Table S1 (Wang et al., 2002).
Geographic distribution of the poultry farms located in north-western (47.0465°N, 21.9189°E), central-western (45.9203°N, 22.8761°E), and south-western (44.4919°N, 26.2813°E) Romania. Black dots indicate the locations.
The speciation of the isolates was based on the sizes of distinct bands corresponding to amplified DNA fragments from the isolate collection. According to Wang et al., a 650 bp band indicates the presence of the 23S rRNA gene, a marker of the genus Campylobacter. A band at 323 bp signifies the presence of the hipO gene, which is associated with C. jejuni, while a band at 126 bp corresponds to the glyA gene, associated with C. coli (Wang et al., 2002). Based on this analysis, 22 isolates were identified as C. coli (126 bp band), whereas 4 isolates were identified as C. jejuni (323 bp band). No other species has been identified.
Detection of hcp gene and of resistance genes
The hcp gene was detected by multiplex PCR analysis as previously described by Harrison et al. (2014), using gltA as a control gene (Harrison et al., 2014). The hcp gene was amplified with the help of hcpF (caagcggtgcatctactgaa) and hcpR primers, while for gltA amplification, primers gltAR (gcgctttggggtcatgcaca) and gltAF (gcccaaagcccatcaagcgga) were applied. Table 1 displays the PCR protocol followed in the investigation. The reference strain Campylobacter NCTC12502 was included as a positive control for hcp amplification, and the PCR conditions are summarized in Table 2. PCR amplification was performed under the following conditions: an initial denaturation at 94°C for 1 minute, followed by 30 cycles of denaturation at 94°C for 15 seconds and annealing at 60°C for 15 seconds. A final extension step was conducted at 72°C for 5 minutes, after which the reactions were held at 4°C.
Virulence Profiles of Campylobacter jejuni and Campylobacter coli Isolates
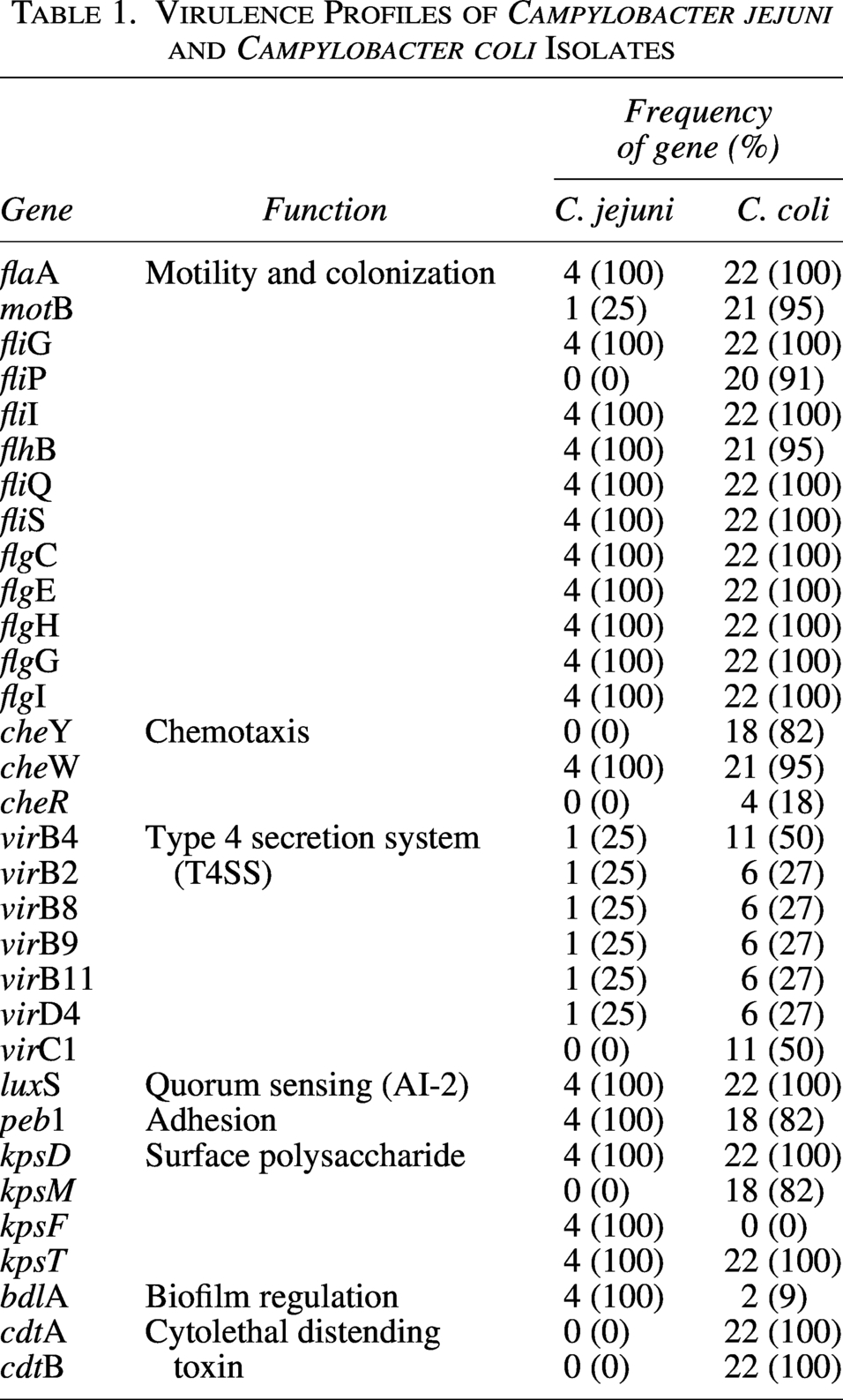
PCR Protocol
Multilocus sequence typing
Partial nucleotide sequences from seven housekeeping loci (aspA, glnA, gltA, glyA, pgm, tkt, and uncA) were analyzed following the MLST scheme described by Dingle et al. (Dingle et al., 2001). Allelic profiles, corresponding sequence types (STs), and clonal complex assignments were subsequently determined by comparison of the obtained sequences with entries in the Campylobacter MLST database. MLST was determined using PubMLST (https://github.com/tseemann/mlst). Genome annotation was provided by using Prokka. To read the genomes and assess the presence of specific genes, Artemis and ACT software were used. Pan-genome analysis was performed using Roary and visualized using Phandango. Phylogenetic analysis was performed using the alignments generated by Roary, followed by ClustalX, and visualized using FigTree (http://tree.bio.ed.ac.uk/software/figtree/).
Results
Detection of hcp gene (T6SS marker) and other virulence-associated gene profiles
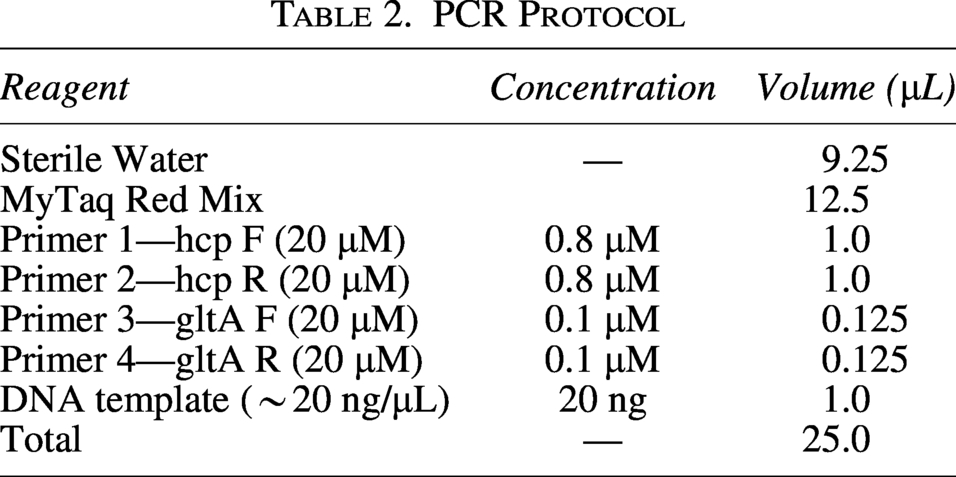
As shown in Figure 2, multiplex PCR detected hcp in 4/4 (100%) C. jejuni isolates, whereas 0/22 (0%) C. coli isolates were hcp-positive. Virulence-associated loci showed both shared and species-differentiated detection patterns (Table 1). Motility/flagellar genes, including flaA, fliG, fliI, fliQ, fliS, flgC, flgE, flgH, flgG, and flgI, were detected in 4/4 (100%) C. jejuni and 22/22 (100%) C. coli. In contrast, fliP was detected in 0/4 (0%) C. jejuni and 20/22 (91%) C. coli, while flhB was detected in 4/4 (100%) C. jejuni and 21/22 (95%) C. coli. motB was detected in 1/4 (25%) C. jejuni and 21/22 (95%) C. coli. Chemotaxis-associated loci differed between species. For example, cheW was detected in 4/4 (100%) C. jejuni and 21/22 (95%) C. coli, whereas cheY was absent in C. jejuni but frequently present in C. coli 18/22 (82%), while cheR was detected only in 4/22 (18%) C. coli.
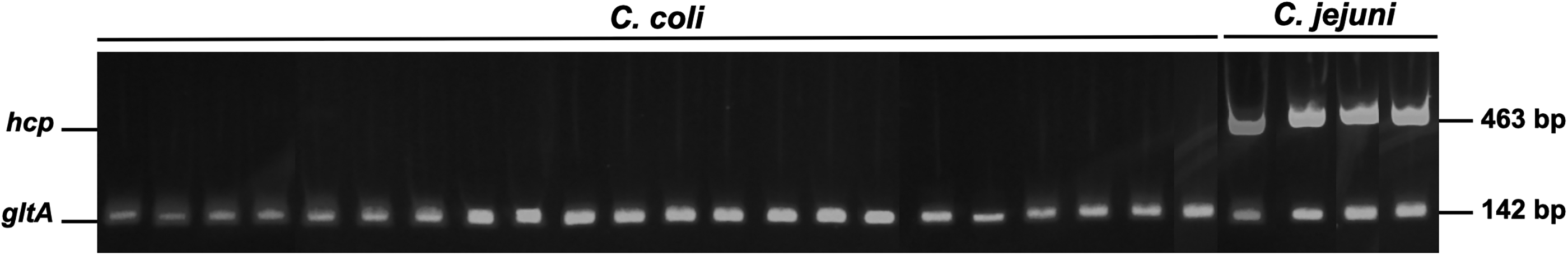
PCR detection of hcp gene in Campylobacter coli (n = 0) and Campylobacter jejuni (n = 4) isolates. Both positive and negative isolates are shown.
Loci associated with the T4SS (virB2, virB4, virB8, virB9, virB11, virD4, and virC1) were detected at higher frequencies in C. coli than in C. jejuni in this dataset. Likewise, virB4 was detected in 1/4 (25%) C. jejuni isolates and in 11/22 (50%) C. coli isolates. virB2, virB8, virB9, virB11, and virD4 were each detected in 1/4 (25%) C. jejuni and 6/22 (27%) C. coli. virC1 was detected only in C. coli 11/22 (50%). The QS locus luxS was detected in 4/4 (100%) C. jejuni and 22/22 (100%) C. coli. Adhesion- and surface polysaccharide-linked loci also showed additional differences, whereas peb1 was detected in 4/4 (100%) C. jejuni and 18/22 (82%) C. coli; kpsD and kpsT were each detected in 4/4 (100%) C. jejuni and 22/22 (100%) C. coli; kpsM was detected only in C. coli (82%); and kpsF was detected in 4/4 (100%) C. jejuni and 0/22 (0%) C. coli. The biofilm-associated locus bdlA was detected in 4/4 (100%) C. jejuni and 2/22 (9%) C. coli. Cytotoxin loci cdtA and cdtB were detected in C. coli 22/22 (100%). In addition, several adhesion and invasion genes, such as cadF, jlpA, peb1A, ciaB, and iamA, were detected in both species, although detection frequencies varied. Genes implicated in toxin production, including cdtA, cdtB, and cdtC, were consistently present in all C. jejuni isolates and most C. coli isolates. These findings highlight the diverse repertoire of virulence factors within the studied Campylobacter populations, contributing to their pathogenic potential.
Resistance-linked gene detection
The tetracycline resistance gene tet(O) was detected in both C. jejuni (25%) and C. coli (27%). Efflux-associated genes showed species-differentiated detection patterns in this dataset. mdtC and mdtI were detected in all isolates (4/4 C. jejuni, 22/22 C. coli), while mdtJ was detected in 4/4 C. jejuni and 21/22 (95%) C. coli. mdtE was not detected in C. jejuni and was detected in C. coli (22/22).
Although mdtC, mdtE, mdtI, and mdtJ were widely distributed among the analyzed isolates and are annotated as putative efflux transporters, their functional contribution in Campylobacter remains to be experimentally validated. mdtC was detected in all C. jejuni and all C. coli genomes. Similarly, both mdtI and mdtJ were present in all C. jejuni isolates and exhibited very high prevalence in C. coli (100% and 95%, respectively). In contrast, mdtE was not detected in any of the C. jejuni genomes, but was identified in 100% of C. coli. In this study, blaOXA-133 gene was found in 27% of the C. coli isolates and was absent from all C. jejuni genomes. Moreover, no C. jejuni isolates harbored yafp, whereas 100% of C. coli isolates were positive for this gene.
MLST results
MLST was performed using internal fragments of seven housekeeping genes (aspA, glnA, gltA, glyA, pgm, tkt, and uncA) (Table 3). MLST assignments are summarized in Table 3. Allele numbers identified for each locus were combined to generate allelic profiles, which were used to assign STs or to identify novel allelic combinations not previously recorded in the MLST database. Recent genotyping studies that apply conventional MLST to identify a broad diversity of Campylobacter have shown STs circulating across human and animal reservoirs, including poultry. For instance, in this study, the MLST results (Table 4) showed that all C. jejuni isolates could not be assigned to previously defined MLSTs, whereas C. coli isolates were predominantly assigned to ST10044, with a smaller number belonging to ST1628 or remaining unassigned.
Genetic Diversity of MLST Loci in Campylobacter Isolates
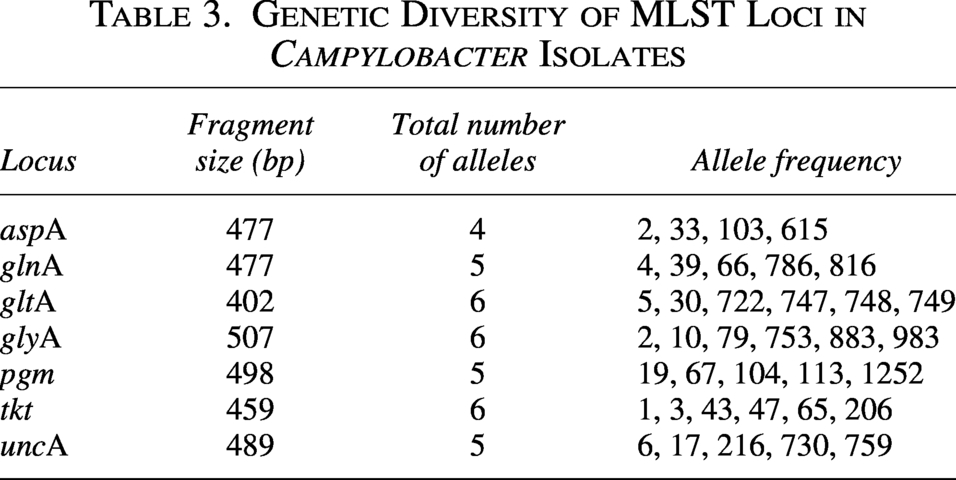
Association Between Multilocus Sequence Typing Lineages and hcp Gene Detection in Campylobacter Isolates

The hcp gene was detected exclusively in C. jejuni isolates, while all C. coli isolates were negative, despite the larger number of C. coli samples analyzed. MLST analysis indicated that most isolates could not be assigned to known STs; all C. jejuni isolates and the majority of C. coli isolates remained unassigned, and only a few C. coli isolates matched previously defined STs (ST10044 and ST1628). Across the seven MLST loci, all loci were polymorphic, with 4–6 alleles per locus identified among the 26 Campylobacter isolates is presented in Table 3. The aspA locus showed four alleles; gltA, glyA, and tkt each showed six alleles; and glnA, pgm, and uncA each showed five alleles. The aspA locus showed the lowest number of alleles (four), whereas gltA, glyA, and tkt each displayed six distinct alleles. The remaining loci unveiled five alleles each. Overall, variation was detected at all seven MLST loci across the analyzed isolates.
Pan-genome summary
The overall structure of the Campylobacter pan-genome (Roary-generated), comprising a conserved core genome and a variable accessory component, is shown in Figure 3A. Gene categories reflect the level of conservation across the dataset and provide an overview of genomic diversity among the analyzed isolates. Most genes are shell genes (3051), which indicates high genetic variability among the strains. The core genome is small (134 genes), meaning that only a small set of genes is universally conserved. This suggests an open pangenome, in which new genes continue to be added as more strains are sequenced. The relatively high number of cloud genes (1219) points to genomic plasticity and a high potential for adaptation and evolution, while 118 genes (soft-core genes) are missing in only one or two strains. Figure 3B is described as comprising 4522 gene clusters across 26 isolates. Phylogenetic relationships inferred from the presence–absence patterns of 4522 gene clusters across 26 isolates, highlighting clear separation between C. jejuni and C. coli and revealing substantial intra-species diversity within the C. coli-dominated subset. Branch lengths reflect gene-content dissimilarity, and tip colors distinguish species, illustrating that closely related isolates can nevertheless differ in accessory gene repertoires. Together, these patterns underscore the accessory genome's contribution to Campylobacter population structure and adaptation in poultry reservoirs.
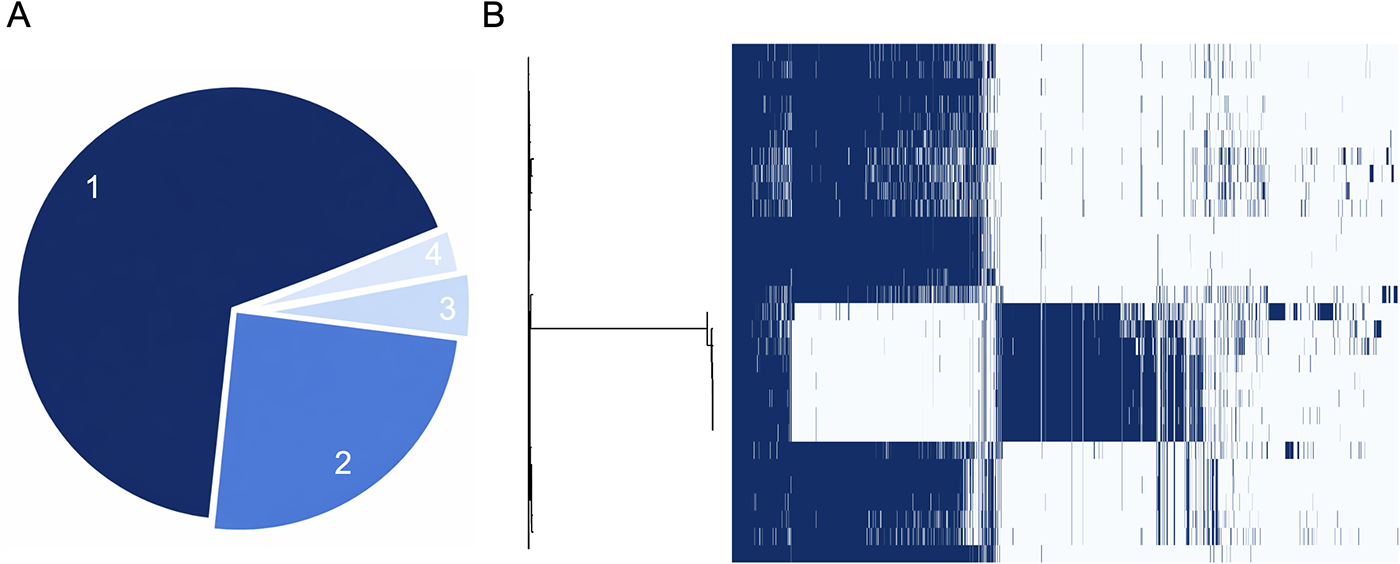
Roary pan-genome composition and gene content diversity of Campylobacter isolates.
Discussion
Core component genes, such as hcp and other T6SS structural loci, are routinely used to identify T6SS+ isolates in genomic datasets and experimental studies (Gupta et al., 2021; 2022; 2024; Robinson et al., 2021). Functional studies show that C. jejuni relies on T6SS for contact-dependent elimination of competitors such as E. coli, influencing microbial community structure in the gut and experimentally in chickens (Gupta et al., 2022; 2024). More broadly, ecological syntheses highlight T6SS as a major component of competition in polymicrobial communities and host-associated microbiota (Gallegos-Monterrosa and Coulthurst, 2021; Rudzite et al., 2023; Singh and Kumari, 2023). In some C. jejuni lineages, complete T6SS loci can be encoded on large plasmids (pT6SS), forming a distinct plasmid group with characteristic gene content and comparative plasmidomics across bacteria further supports the occurrence of T6SS modules on megaplasmids, emphasizing potential for horizontal transfer and lineage-specific stability or loss (Harrison et al., 2025; Morgado and Vicente, 2022; Robinson et al., 2021; 2022).
In contrast, the cytolethal distending toxin (CDT) genes (cdtA and cdtB) were detected only in C. coli, while the C. jejuni subset was negative for targeted cdt genes, and cdtC was not detected. Campylobacter CDT biology is being actively re-examined, with recent work challenging assumptions about holotoxin organization and emphasizing that the precise structural and functional relationships among cdtA, cdtB, and cdtC, as well as the minimal subunit requirements for virulence, remain not entirely settled (Chen et al., 2023; Lara-Tejero and Galán, 2001; Méndez-Olvera et al., 2016; Pons et al., 2019). Nonetheless, converging epidemiological, in vitro, and in vivo data indicate that CDT is a major virulence and colonization factor for Campylobacter, contributing to substantial infection, mucosal inflammation, and, in some models, development of tumors (Asakura et al., 2007; Ge et al., 2008; Lai et al., 2016; Le et al., 2024). The detection of virB genes in a subset of isolates further supports heterogeneity among mobile elements. virB-associated T4SS loci are typically plasmid-linked (pVir-like elements) and have been variably associated with invasion phenotypes; however, reported clinical associations with severe outcomes are inconsistent across studies and should not be overstated in a poultry-only dataset (Garcia-Fernandez et al., 2024; He et al., 2025). Here, virB movement is most appropriately interpreted as an indicator of circulating plasmid diversity and potential horizontal gene transfer within the broiler reservoir, consistent with broader accessory-genome variability.
Limitations should be considered when interpreting these findings. The collection is modest in size and species-imbalanced (C. jejuni n = 4), which limits the interpretation of species-level distributions of accessory loci. Despite these limitations, the data provide a useful baseline for Romanian poultry reservoirs and motivate integrated WGS-phenotype surveillance to support risk-based control. Future work combining cgMLST/SNP phylogenies with phenotyping (MIC testing, colonization fitness, and competition assays) will be particularly valuable for resolving lineage-module associations in Romanian poultry reservoirs.
Conclusion
This study provides a molecular characterization of Campylobacter jejuni and C. coli isolates obtained from poultry caeca in Romania, focusing on virulence-associated traits, antimicrobial resistance determinants, and genomic diversity. Clear differences were observed between the two species in the distribution of virulence genes, including detection of the hcp gene only in C. jejuni isolates, suggesting a species-specific association with carriage of the T6SS in the analyzed samples. Both species harbored multiple resistance-related genes, including efflux-associated and tetracycline resistance genes. Pan-genome analysis further indicated significant genomic variability, consistent with an open pan-genome and ongoing gene gain and loss. Together, these findings reinforce the genomic complexity of Campylobacter in poultry and support the value of integrated molecular approaches for monitoring virulence and antimicrobial resistance in food-animal reservoirs.
Authors’ Contributions
I.A.B.: Conceptualization, investigation, methodology, project administration, and writing—original draft; L.S.: Project administration, resources, supervision, and writing—review and editing; O.G.: Formal analysis, methodology, software, and data curation; N.C.: Conceptualization, data curation, project administration, formal analysis, funding acquisition, resources, supervision, validation, visualization, and writing—review and editing; I.B.: Conceptualization, supervision, validation, visualization, and writing—review and editing.
Footnotes
Acknowledgments
The authors would like to thank the Doctoral School “Engineering of Vegetable and Animal Resources”, University of Life Sciences “King Mihai I” from Timişoara (Calea Aradului 119, 300645 Timişoara, Romania), for the academic support and resources provided throughout the development of this research.
Funding Information
This work was funded by the
Disclosure Statement
The authors declare no conflicts of interest.
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.