
Abstract
Antipsychotics are commonly prescribed to treat several neuropsychiatric disorders, including schizophrenia, mania in bipolar disorder, autism spectrum disorder, delirium, and organic or secondary psychosis, for example, in dementias such as Alzheimer’s disease. There is evidence that typical antipsychotics such as haloperidol are more effective in reducing positive symptoms than negative symptoms and/or cognitive deficits. In contrast, atypical antipsychotic agents have gained popularity over typical antipsychotics, due to fewer extrapyramidal side effects and their theoretical efficacy in controlling both positive and negative symptoms. Although these therapies focus on neuron-based therapeutic schemes, glial cells have been recognized as important regulators of the pathophysiology of neuropsychiatric disorders, as well as targets to improve the efficacy of these drugs. Glial cells (astrocytes, oligodendrocytes, and microglia) are critical for the central nervous system in both physiological and pathological conditions. Astrocytes are the most abundant glial cells and play important roles in brain homeostasis, regulating neurotransmitter systems and gliotransmission, since they express a wide variety of functional receptors for different neurotransmitters. In addition, converging lines of evidence indicate that psychiatric disorders are commonly associated with the triad neuroinflammation, oxidative stress, and excitotoxicity, and that glial cells may contribute to the gliotoxicity process. Conversely, glioprotective molecules attenuate glial damage by generating specific responses that can protect glial cells themselves and/or neurons, resulting in improved central nervous system (CNS) functioning. In this regard, resveratrol is well-recognized as a glioprotective molecule, including in clinical studies of schizophrenia and autism. This review will provide a summary of the dual role of antipsychotics on neurochemical parameters associated with glial functions and will highlight the potential activity of glioprotective molecules to improve the action of antipsychotics.
Impact statement
Glial cells are crucial to maintain brain homeostasis, although they can also participate in the pathogenesis of neuropsychiatric disorders, such as schizophrenia. In addition, these cells are putative targets of antipsychotics. These medications are subdivided in typical and atypical, depending on their receptor interaction. Our group has published several studies investigating the effects of antipsychotics on glial functions, particularly glutamate metabolism, redox homeostasis, trophic factors release, and inflammatory response. In addition, in this review, we described glioprotective agents that could act as adjuvants to attenuate glial dysfunctions associated with neuropsychiatric disorders.
Introduction
In the 1950s, the discovery of the effects of antipsychotics changed paradigms in the treatment of several neuropsychiatric disorders, such as schizophrenia, mania in bipolar disorder, autism spectrum disorder, delirium, and organic or secondary psychosis, for example, in dementia.1,2 Typical antipsychotics were first developed and were used to treat schizophrenia. These drugs act on dopamine receptors (D1, D2, D3) in the brain, particularly blocking D2 receptor in the mesolimbic and mesocortical systems, which are respectively associated with reward/motivation and emotion/cognition. 3 In line with this, blockade of limbic D2 receptors is associated with the attenuation of the positive symptoms of schizophrenia, such as hallucinations and delusions. Classic examples of typical antipsychotics are chlorpromazine and haloperidol, and this last medication can induce sedative side effects and increased weight. 4 In psychiatric patients, chronic treatment with typical antipsychotics is limited by extrapyramidal side effects induced by dopamine blockade, such as parkinsonism, akathisia, dystonia, and tardive dyskinesia. 5
The second-generation antipsychotics, also known as atypical antipsychotics, include clozapine, risperidone, quetiapine, and olanzapine. These drugs reduce not only dopamine neurotransmission but also act on serotonin receptors, especially 5HT2A receptors, typically as antagonists.4,6 In addition, atypical antipsychotics act as adrenergic (α1, α2), cholinergic (M1), and histaminergic (H1) receptor antagonists.7,8 Therefore, these antipsychotics are more effective for treating negative and cognitive symptoms of schizophrenia, since they also contribute to improve mood and anxiety. 9 It is important to note that although there are some efficacy differences between antipsychotics, the most relevant are associated with differences in side effects. 10 However, this greater range of action brought about new side effects, especially weight gain, hyperglycemia, dyslipidemia, insulin resistance, and even type 2 diabetes mellitus, which affect almost half of patients. Weight gain effects are more pronounced with clozapine and olanzapine, attributed to their actions on serotonergic 5HT2C and histaminergic H1 receptors. Ziprasidone and aripiprazole act less on 5HT2C receptors and cause less weight gain. 11 Figure 1 depicts some mechanisms of antipsychotics.
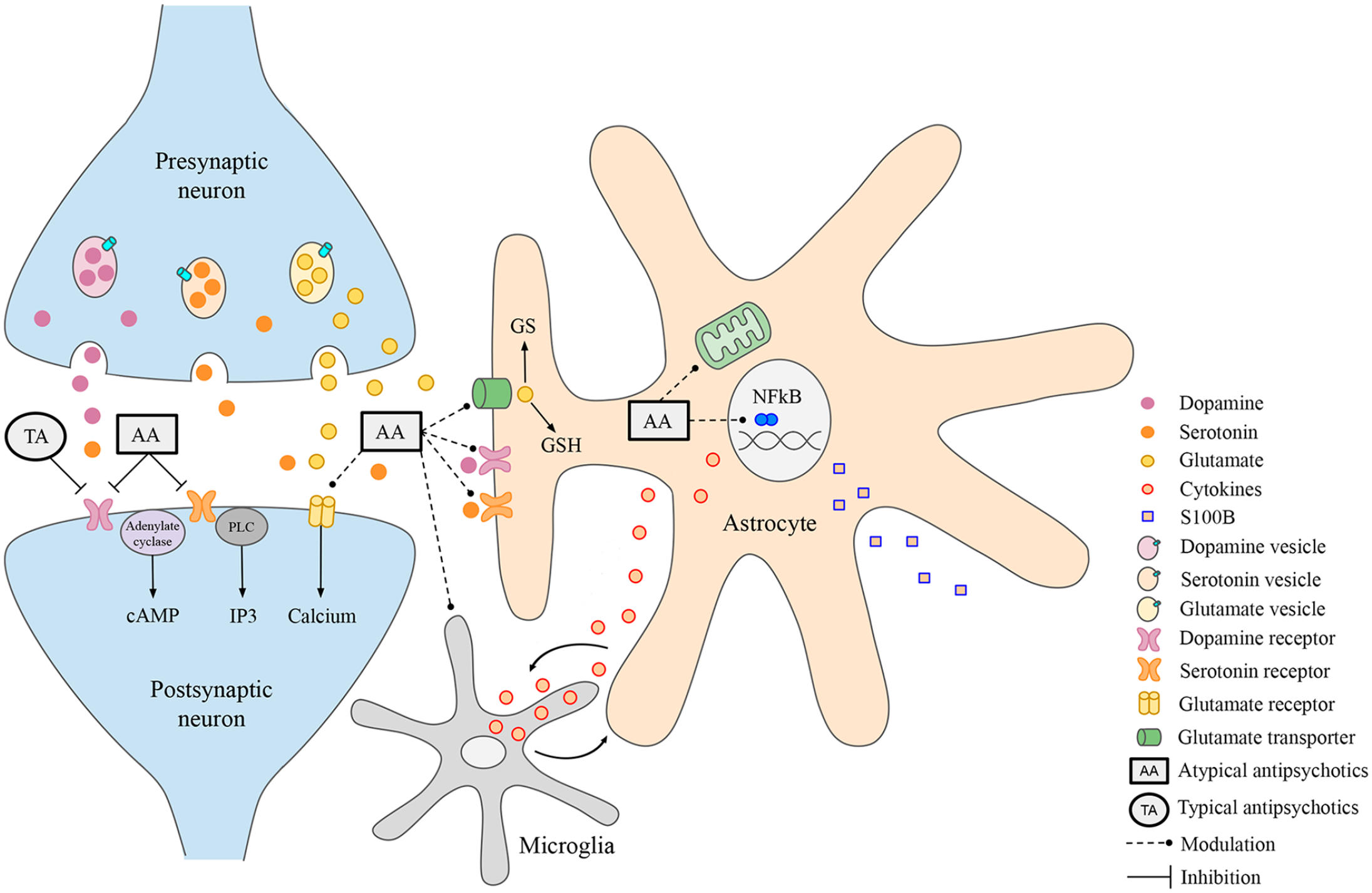
The tripartite synapse as a target of antipsychotics. Presynaptic neurons release neurotransmitters (dopamine, serotonin, glutamate), accordingly to their specialization and brain region, from synaptic vesicles within the synaptic cleft. Neurotransmitters interact with their specific ionotropic and/or metabotropic receptors at postsynaptic neurons. Astrocytes also express dopamine and serotonin receptors, as well as glutamate transporters, therefore modulating synaptic transmission. Typical and atypical antipsychotics, TA and AA, respectively, can act blocking mainly neuronal dopamine and/or serotonin receptors. In addition, AA can modulate glutamate receptors and transporters (dashed lines). Therefore, astrocytes can also be putative targets of antipsychotic medications, as discussed in the text. Astrocytes actively participate in NFκB-coordinated inflammatory, mitochondrial responses, and glutamate metabolism via GS activity and GSH synthesis, all of which are involved in the pathogenesis of neuropsychiatric disorders and may be also modulated by antipsychotics. Finally, there is a close relationship between astrocytes and microglia, particularly related to inflammatory response.
Finally, receptor pleiotropism (D2/5HT2) and the rapid D2 receptor dissociation must be considered to differentiate typical from atypical antipsychotics, as well as the better tolerability and improved effectiveness for negative, affective, and cognitive symptoms.12,13 Moreover, although antipsychotics are available for treating neuropsychiatric disorders, multidisciplinary treatments are needed, such as modern therapy and psychological support, to improve not only the symptoms of these conditions but also behavioral aspects and social interactions.14,15
Considering that our research group has studied the effects of typical and atypical antipsychotics on glial cells, this review will provide a summary of the dual role of these drugs on cellular, molecular, and pharmacological mechanisms associated with glial functions. In addition, we will highlight the potential activity of glioprotective molecules to improve the action of antipsychotics on neurochemical parameters, particularly related to glial cells, reducing side effects of these drugs and consequently improving life quality during the aging process, since these medications are chronically used.
Glial cells as targets of antipsychotics
Although neurons have been assumed to be the major mediators of antipsychotic pharmacological effects, glial cells have emerged as important cellular targets for these drugs. Glial cells (astrocytes, oligodendrocytes, and microglia) are critical for the central nervous system (CNS) in both physiological and pathological conditions. Astrocytes are the most abundant glial cells and play important roles in brain homeostasis, being responsible for trophic support, maintenance of the blood-brain barrier (BBB), energy metabolism, antioxidant defenses, inflammatory responses, gliotransmission, among others.16,17 These cells express a wide variety of functional receptors for different neurotransmitters, such as glutamate, GABA, as well as adrenergic, purinergic, dopaminergic, and serotonergic receptors.18,19 However, the effects of antipsychotics in these cells are potentially relevant and appear not to be only directly related to these receptors; thus, typical and atypical antipsychotics can differentially modulate glial-related processes. With regard to the tripartite synapse, in particular, the communication between neurons and astrocytes reinforces the versatility of the glial cells in the regulation of neural functions from development to mature brain. 16 Of note, alterations in neuron–glia communication are critical for understanding the pathomechanisms of psychiatric disorders due to commitment of neurotransmitter systems, excitatory-inhibitory balance, neurotrophic state, and inflammatory response.20,21 In terms of astrocytes and dopamine-glutamate neurotransmitter systems, D2 receptors are highly expressed in astrocytes22,23 and, therefore, glial cells may participate in the pathophysiology of neuropsychiatric disorders, and may consequently be involved in the mechanism of action of antipsychotic drugs.24,25
Excessive activation of dopamine receptors can lead to a decrease in the activity of the glutamatergic system, which contributes to psychotic symptoms, commonly observed in neuropsychiatric disorders. By blocking dopamine receptors, antipsychotics can increase the activity of the glutamatergic system, helping to reduce psychotic symptoms.26,27 Glutamate is the main excitatory neurotransmitter in the CNS and plays a key role in regulating synaptic plasticity and memory formation. 28 However, high glutamate levels in the extracellular space are potentially neurotoxic. 29 Most of the synaptically released glutamate is taken up by astrocytes, the main cells that express glutamate transporters. 26 Thus, these cells optimize neuronal functions and prevent glutamate excitotoxicity. 30 Moreover, astrocytes convert glutamate into glutamine via the enzyme glutamine synthetase (GS), and glutamine is then released back to the neurons where glutamate is regenerated by glutaminase. 31
Schizophrenia is among the most disabling mental health conditions and affects approximately 24 million people worldwide. 6 With the reformulation of the hyperdopaminergic hypothesis and its integration with the glutamatergic hypofunction hypothesis, the role of astrocytes in schizophrenia has been further valued, since glutamate uptake and glutamine synthesis are specific functions of these cells.27,32 Furthermore, as the relevance of astrocytes has grown in synaptic communication, they have a role in underpinning other neuropsychiatric disorders, including bipolar and autism spectrum disorders, that use atypical antipsychotics as therapeutic strategies.17,33–36 In addition, atypical antipsychotics have been prescribed for delirium, for example, postsurgery, alcohol withdrawal, and organic or secondary psychosis, for example, dementias including Alzheimer’s disease.13,37–39
Evidence suggests abnormalities in astroglial glutamate transporter functions and perisynaptic glutamate buffering in schizophrenia,
40
as well as in neurodegenerative conditions.
41
It has been shown that the atypical antipsychotic, clozapine, can reduce glutamate transporter 1 (GLT-1) expression and function in astrocytes, which can be driven by nuclear factor kappa B (NFκB), protein kinase B (Akt), and extracellular signal-regulated kinase (ERK) pathways,42,43 and is associated with changes in the expression of the synaptic vesicle protein synaptophysin.
43
The release of glutamate by astrocytes can also be enhanced by clozapine, quetiapine, and brexpiprazole through activated hemichannels, which can enhance tripartite synaptic glutamatergic transmission.
44
In addition, clozapine activates glial exocytosis of D-serine, an endogenous N-methyl-
With particular regard to risperidone and haloperidol, over the last two decades, our group has published several studies that have investigated the effects of these antipsychotics on glial functionality (Table 1). In line with this, in C6 astroglial cells, a comparative study between typical and atypical antipsychotics showed that risperidone, but not haloperidol, is able to increase glutamate uptake and GS activity.47,48 In addition, the chronic blockade of D2 receptors with haloperidol elevates extracellular glutamate levels in the striatum.49,50 Taken together, these findings indicate that the modulation of astrocyte functions related to the maintenance of glutamate neurotransmission is relevant for the therapeutic effects of antipsychotics. In this context, while specific antipsychotics, such as clozapine and haloperidol, ameliorate schizophrenia-related glutamate hypofunction, their long-term use can be associated with glutamate imbalance toward excitotoxicity and the emergence or progression of neurodegeneration. On the contrary, other antipsychotics, such as risperidone, may exert neuroprotective effects against brain disorders associated with excitotoxicity, including autism.51–53
Effect of typical and atypical antipsychotics in glial cells.
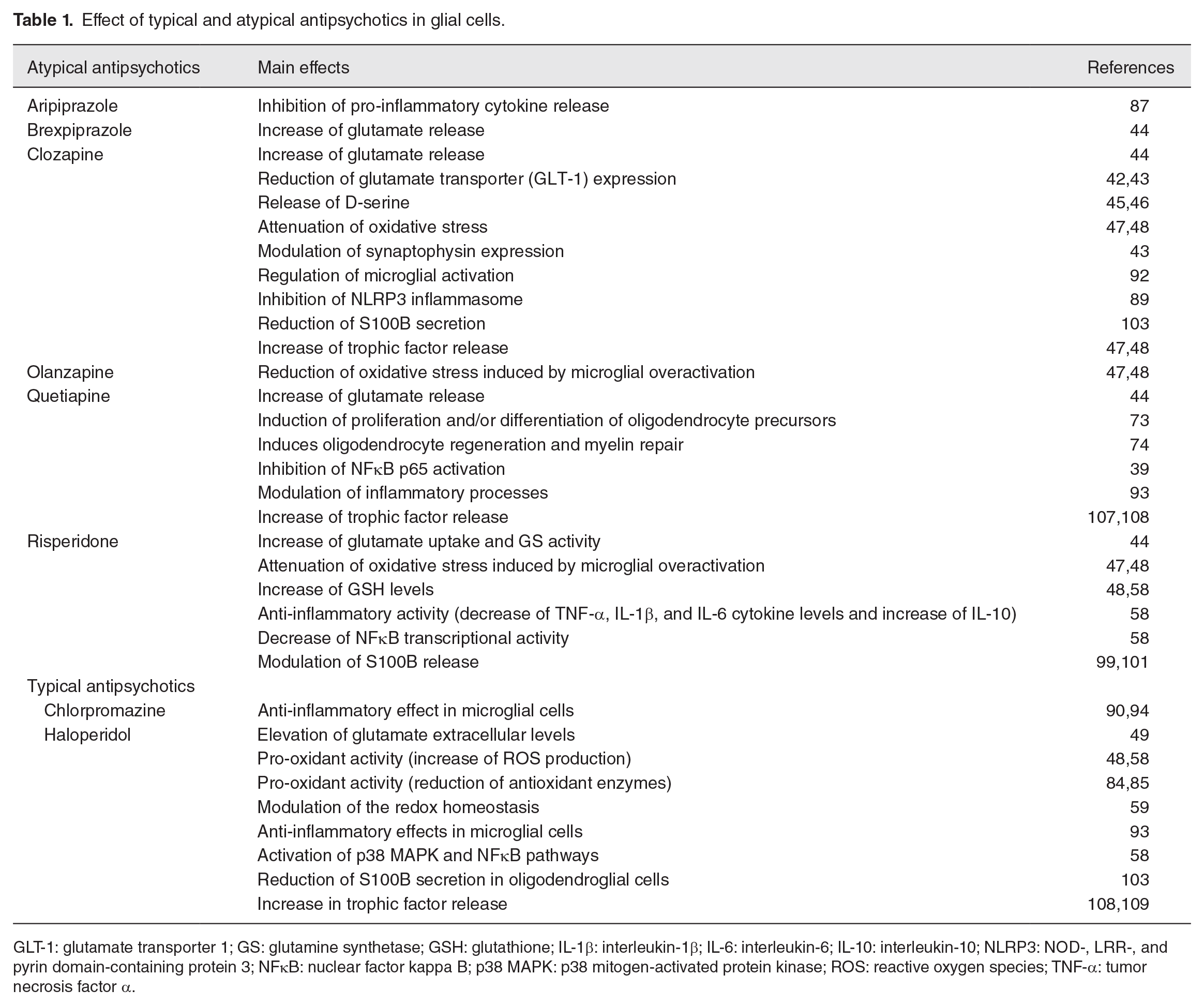
GLT-1: glutamate transporter 1; GS: glutamine synthetase; GSH: glutathione; IL-1β: interleukin-1β; IL-6: interleukin-6; IL-10: interleukin-10; NLRP3: NOD-, LRR-, and pyrin domain-containing protein 3; NFκB: nuclear factor kappa B; p38 MAPK: p38 mitogen-activated protein kinase; ROS: reactive oxygen species; TNF-α: tumor necrosis factor α.
Glutamate can also be used for the synthesis of the antioxidant molecule, glutathione (GSH), particularly in astrocytes. Notably, oxidative stress may be another common mechanism in the pathogenesis of psychiatric and neurodegenerative disorders.54,55 In this regard, both glutamate and dopamine neurotransmitters are redox active molecules and can be involved in the generation of reactive oxygen species (ROS) even during normal neurotransmission. Following alterations in glutamatergic and dopaminergic activity, ROS production may be exacerbated and contribute to an increased brain vulnerability to oxidative damage, such as glutamate transporter oxidation.56,57 Concomitantly with the increase in glutamate uptake and GS activity, risperidone increases GSH levels in C6 astroglial cells. However, haloperidol did not change this antioxidant defense and increased the production of ROS.48,58 Accordingly, chronic treatment with haloperidol has been associated with oxidative stress,59,60 at least in part due to an impairment in GSH levels,61–64 and therefore with the pathogenesis of extrapyramidal side effects. 59 The cytotoxicity induced by the typical antipsychotics, haloperidol, and chlorpromazine in astrocytes depends on increased intracellular calcium levels,63,64 which is also a mechanism underlying oxidative stress. Therefore, although typical antipsychotics such as haloperidol can solve neurotransmitter dysfunctions, they can aggravate oxidative stress. In contrast, the ability of risperidone to modulate the endogenous antioxidant, GSH, reinforces its neuroprotective properties rather than neurotoxic ones. Corroborating this idea, it has been suggested that N-acetylcysteine, a precursor of cysteine for GSH synthesis, may be an adjunctive strategy in the treatment of psychiatric disorders.65,66
It is important to note that risperidone has been used to treat behavior problems such as aggression, self-injury, and sudden mood changes in children who have autism, a condition that causes difficulty in interacting with others, and problems with communication. 67 In this context, risperidone offers a valuable emerging option for the treatment of social behavior associated with autistic disorder in children and adolescents,68,69 and our research group contributes to demonstrate the neuroprotective effects of this antipsychotic drug against brain illness, at least partially via modulation of astrocyte functions.
In addition to astrocytes, oligodendrocytes and microglia are crucial to the maintenance of brain homeostasis. 30 Oligodendrocytes form myelin around axons in the CNS to ensure fast and efficient electrical conduction along myelinated axons. 70 Decreased numbers of oligodendrocytes and/or abnormalities involving myelin-related structures may be involved in the pathogenesis of neuropsychiatric disorders, while protection of oligodendrocytes/myelin may improve symptoms associated with schizophrenia.71,72 The atypical antipsychotics, quetiapine and olanzapine, promote the proliferation and/or differentiation of precursor oligodendrocyte cells in vitro, 73 and quetiapine also enhances oligodendrocyte regeneration and myelin repair in experimental models of cuprizone-induced demyelination 74 and global cerebral ischemia. 75 These neuroprotective effects are also related to the anti-inflammatory properties of atypical antipsychotics,76–78 which are possibly associated with their H1 receptor antagonism. 77 In contrast, the typical antipsychotic haloperidol was not effective in promoting differentiation and maturation of oligodendrocyte precursor cells exposed to cuprizone, reinforcing the differential effects of antipsychotics in the development of oligodendrocytes. 79
Microglia are responsible for innate brain immune response. These cells may initiate the formation of intracellular multiprotein complexes called inflammasomes, as well as cytokine release and production, and consequently, neuroinflammation. 80 In this context, they can trigger neurodegenerative processes, which can result in cognitive dysfunction, being a potential aggravating factor of psychiatric disorders. 81 It is important to note that, in neuropsychiatric disorders, such as schizophrenia, microglia can induce astrocyte reactivity, neuroinflammation, and oxidative stress, which can cause neuronal death.17,82,83 Table 1 also shows that treatment with antipsychotics such as risperidone, clozapine, and olanzapine reduce oxidative stress induced by microglial overactivation by increasing GSH, the main antioxidant of the CNS.47,48 Interestingly, first-generation antipsychotics can induce oxidative stress by reducing antioxidant enzymes, as observed with haloperidol treatment, which can contribute to the development of side effects.84,85 In addition, several in vitro studies in microglial cells have demonstrated anti-inflammatory effects particularly related to the atypical antipsychotics quetiapine, clozapine, risperidone, olanzapine, and aripiprazole. In general, these molecules are able to inhibit the release of a wide range of pro-inflammatory cytokines, including tumor necrosis factor α (TNF-α), interleukin-1β (IL-1β), interleukin-2 (IL-2), monocyte chemoattractant protein-1 (MCP-1), and interferon-γ (IFN-γ), in addition to decreasing nitric oxide (NO) generation and nicotinamide adenine dinucleotide phosphate (NADPH) oxidase activity.86–89 Moreover, atypical antipsychotics can increase the anti-inflammatory cytokine interleukin-10 (IL-10).89,90 The anti-inflammatory effects of these drugs may occur both at basal conditions and upon inflammatory stimuli and may involve different mechanisms. Of note, clozapine can inhibit the NLR family pyrin domain containing 3 (NLRP3) inflammasome, 89 while quetiapine inhibits the activation of the NFκB p65 pathway, 39 a master regulator of inflammation. Inhibition of microglial proton currents is also involved in the anti-inflammatory effect of clozapine and does not involve serotonin receptors. 91 Moreover, in vivo studies showed that clozapine can regulate microglial activation in a model of chronic experimental autoimmune encephalomyelitis, 92 while quetiapine modulates brain inflammatory processes in adult rats prenatally exposed to lipopolysaccharide (LPS), a model of schizophrenia-like disturbances, 93 and in a transgenic mice model of Alzheimer’s disease. 39 With regard to typical antipsychotics, chlorpromazine and haloperidol have been demonstrated to exert some anti-inflammatory effects in microglial cells,90,94 also by decreasing the release of pro-inflammatory cytokines. However, it is important to note that more significant anti-inflammatory activities have been associated with atypical drugs in microglial cells.
Converging lines of evidence indicate that psychiatric disorders are commonly associated with the triad neuroinflammation, oxidative stress, and excitotoxicity.52,95,96 However, information about the effects of antipsychotics on inflammatory response in astrocytes is still scarce, but typical and atypical drugs seem to have differential roles. While risperidone decreased TNF-α, IL-1β, and IL-6 and increased IL-10 in astroglial cells, haloperidol had the opposite effect. The anti-inflammatory response induced by risperidone also involved a decrease in transcriptional activity of NFκB, in contrast to an activation of the p38 mitogen-activated protein kinase (p38 MAPK)/NFκB pathway by haloperidol. 58
In addition to pro-inflammatory cytokines, activated astrocytes can release elevated levels of S100B, a glia-derived protein that has been shown to be increased in psychiatric disorders, including schizophrenia.17,97,98 S100B release is increased by pro-inflammatory cytokines, and both the antipsychotics, haloperidol, and risperidone can decrease S100B release stimulated by IL-6 in astroglial cells,99,100 while risperidone positively modulates S100B extracellular levels at basal conditions. 101 These drugs also inhibited the decrease in S100B secretion induced by dopamine agonist apomorphine. 102 Furthermore, both haloperidol and clozapine reduce the release of S100B from oligodendroglial cells. 103 Therefore, although differential effects are observed, dopamine receptors seem to regulate S100B release by astrocytes, and this protein may represent a specific astroglial target of antipsychotics. Indeed, S100B can have dual functions, acting either as a pro-inflammatory signal or exerting trophic roles.104–106 In line with a role in modulating trophic signaling, quetiapine, clozapine, and haloperidol increased the release of trophic factors, including brain-derived neurotrophic factor (BDNF) and glial cell line-derived neurotrophic factor (GDNF) from cultured astroglial cells.107–109
Ultimately, there is a close relationship between the triad neuroinflammation, oxidative stress, and excitotoxicity with the release of trophic factors, which can be modulated by transcription factors including NFκB and nuclear factor erythroid 2-related factor 2 (Nrf2). 30 Furthermore, the AMP-activated protein kinase (AMPK) signaling pathway regulates energy balance, mitochondrial function, detoxification, and antioxidant defenses, promoting cellular homeostasis. 110 This pathway crosstalks with Nrf2 and is increased by quetiapine in cultured astrocytes from the hypothalamus, a brain region that is crucial for the control of metabolism, indicating a potential link between antipsychotics and metabolic dysfunction and antipsychotic-induced obesity.30,111 Thus, changes in cellular responses and mechanisms related to glial cells may be associated with the pathogenesis of neuropsychiatric disorders and side effects caused by these medications.
Antipsychotics and gliotherapy
The role of glial cells in the development and progression of CNS diseases, commonly associated with gliotoxicity, as well as specific responses of glial cells by which they can protect themselves or neurons (a process named as glioprotection) is critical for brain homeostasis. 30 In this regard, glial cells may represent putative cellular targets to treat neuropsychiatric disorders, as well as to improve preventive and therapeutic strategies to reduce the side effects associated with these medications. It is important to note that although the pathophysiology behind the psychiatric disorders, such as schizophrenia and mood disorders, is not fully understood, several studies indicate alterations in neuron–glia interaction as potential mechanisms for these conditions. 20
Table 1 displays specific effects of typical and atypical antipsychotics on glial parameters, particularly related to astrocyte functions. As demonstrated, antipsychotics can induce glial responses, including glial damage. In this sense, glial cells are key players in schizophrenia pathology since a plethora of studies have shown structural and functional abnormalities in all three types of glial cells in schizophrenia patients. In this context, both the progression of schizophrenia as well as the effects of the prolonged use of these medications can be associated with changes in glial cells. Moreover, alterations in glutamate neurotransmission are common in hippocampus of schizophrenia patients, 40 and cognition problems may be less common in people with late onset schizophrenia, 112 while people who receive a diagnosis of very late onset schizophrenia (over 60 years old) have a significant risk of dementia.
Psychosis is a common symptom in schizophrenia, as well as in Alzheimer’s disease, and few safe and effective treatments are available for this later condition. In addition, psychotic symptoms are associated with hospitalization or institutionalization, and cognitive and functional impairment may accelerate cognitive decline and mortality in these patients. 113 It is noteworthy that around 40% of people with Alzheimer’s disease reported psychosis, and atypical antipsychotics are used to treat psychosis in dementia. These medications can also modulate the inflammatory profile, which is an important contributor the pathogenesis/progression of dementia.114,115
Although atypical antipsychotics present relevant efficacy for treating schizophrenia and autism spectrum disorder, they demonstrate modest efficacy for dementia with substantial adverse effect burden and mortality risk. Overall, meta-analytical evidence best supports aripiprazole and risperidone, compared to olanzapine and quetiapine. 113 However, the risk of each medication must be evaluated in the choice of the treatment. Thus, pharmacokinetic–pharmacodynamic studies have explored nuanced dosing approaches to better optimize the balance between efficacy and safety, and astrocytes emerge as potential targets, because they are also responsible for BBB maintenance and integrity. It is important to note that neuropsychiatric disorders, such as Alzheimer’s disease, present disrupted BBB;116,117 thus, astrocytes may be key cells to control the entry of drugs within the CNS, and consequently, the dose of medications needed to ensure a successful treatment. In line with this, many studies strive to develop specific therapies for neuropsychiatric disorders, but they are not focused on glial cells. With gliotherapy approaches, the development of new drugs or the repurposing of existing drugs could contribute to improve the treatment of neurodegeneration associated with antipsychotic use. The long-term effects of antipsychotics on glial cells in different brain regions should be properly considered, as these may present innovative perspectives for pharmacotherapy. 17
Could glioprotective molecules attenuate neuropsychiatric disorders? Focus on glioprotection/glial pharmacology
Increased evidence shows glial impairment in in vitro and in vivo experimental models of autism, a neurodevelopmental disorder. In addition, atypical antipsychotics used to treat this condition can induce glial alterations and the long-term treatment with these medications can contribute to the progression of neurodegenerative diseases; thus, new drugs for autistic patients need to be investigated. Glioprotective strategies may attenuate these abnormalities, particularly because mood disorders are glial disorders resveratrol, a well-tested glioprotective molecule, presents efficacy, tolerability, and safety, as well as anti-inflammatory, antioxidant, and neuroprotective properties, emerging as a promising therapeutic strategy for large-scale studies in autism spectrum disorder, as well as in schizophrenia. 118
Resveratrol can also protect neurons in brain-implantable electrodes, such as those used in deep brain stimulation. 119 In this regard, glioprotective molecules can result in an overall improvement in CNS functioning, including during glial damage associated with long-term use of antipsychotics in neuropsychiatric disorders. Our group has demonstrated that resveratrol and other molecules can regulate several astrocyte functions, including antioxidant defenses, inflammatory response, trophic factor release, and glutamate homeostasis, both at basal conditions and against harmful stimuli. 30 In addition, resveratrol is able to prevent age-related functional alterations of astrocytes. These effects are associated with different signaling pathways, including Nrf2, heme oxygenase 1 (HO-1), SIRT1, PI3K/Akt, AMPK, and NFκB. 30 In this context, selective overexpression of HO-1 in the astrocytes of mice induced subcortical oxidative stress and mitochondrial damage and resulted in a neurophenotype that bears resemblances to human schizophrenia, suggesting a role of HO-1 in the development of monoaminergic circuitry.120,121 Interestingly, we recently demonstrated that resveratrol was able to decrease Nrf2 and HO-1, indicating that these pathways may not be the main target of resveratrol in hypothalamic astrocytes. 122 Furthermore, evidence suggests a possible link between hypothalamic gliogenesis and weight gain associated with olanzapine treatment in mice. 123 In addition, olanzapine seems to increase the expression of astrocyte markers (glial fibrillary acidic protein—GFAP and S100B) and hypothalamic activation of the toll-like receptor 4 (TLR4) signaling pathway, which may be associated with olanzapine-induced obesity. 124 Another study from our group showed that resveratrol decreased the expressions of GFAP, S100B, TLR4, and NFκB after long-term treatment in hypothalamic astrocyte cultures from aged rats. 125 These data corroborate the potential multitarget feature of resveratrol and its potential role for avoiding antipsychotic side effects involving the hypothalamus, which is also critically affected in neuropsychiatric disorders.
Moreover, resveratrol exhibits glioprotective effects on microglial and oligodendroglial cells. 30 Haloperidol and olanzapine significantly decreased the number of astrocytes, but not of oligodendrocytes, in the brain of antipsychotic-exposed monkeys. 126 Thus, resveratrol could attenuate this astrocyte damage, presenting a significant glial pharmacological potential.
Finally, specific interleukins, such as IL-10, IL-12, IL-2, interferon-α (INF)-α, TNF-α, and transforming growth factor β (TGF-β), are signaling mediators that normally increase during the early stages of schizophrenia but are normalized by antipsychotic treatment. Resveratrol can modulate cytokine release in several experimental models and could contribute to regulate this early inflammatory response and, consequently, the onset and/or progression of neurodegeneration. Besides resveratrol, other naturally occurring molecules of plant origin can promote glioprotection, such as curcumin, isoflavones, and sulforaphane, as well as endogenous mammalian compounds including lipoic acid and guanosine (a guanine-based purine). Therefore, these potential glioprotective agents could be investigated as adjuvants to attenuate glial disorders, because neuropsychiatric disorders are also indeed glial disorders.
Concluding remarks
In summary, due to the critical role of glial cells in the pathogenesis and progression of neuropsychiatric disorders, as well as their potential role as cellular targets to treat these conditions, we highlight in this review how glioprotective molecules can attenuate the long-term effects associated with typical and atypical antipsychotics (Figure 2). In addition, glial cells, particularly astrocytes, may be relevant in the repurposing of existing antipsychotics, as these cells are the most versatile in the brain. With this in mind, glioprotective molecules can form part of adjuvant treatments, as proposed in clinical studies that indicated resveratrol as adjuvant therapy for negative symptoms in patients with schizophrenia. By understanding how antipsychotics act in glial cells, glial-based preventive/therapeutic strategies might emerge to delay and to prevent the development of undesirable consequences associated with these medications. Therefore, antipsychotics have two faces (Janus face), because they are critical to treat neuropsychiatric disorders, but they can induce side effects and contribute to glial reactivity; however, glioprotective molecules could attenuate these contrasting aspects.
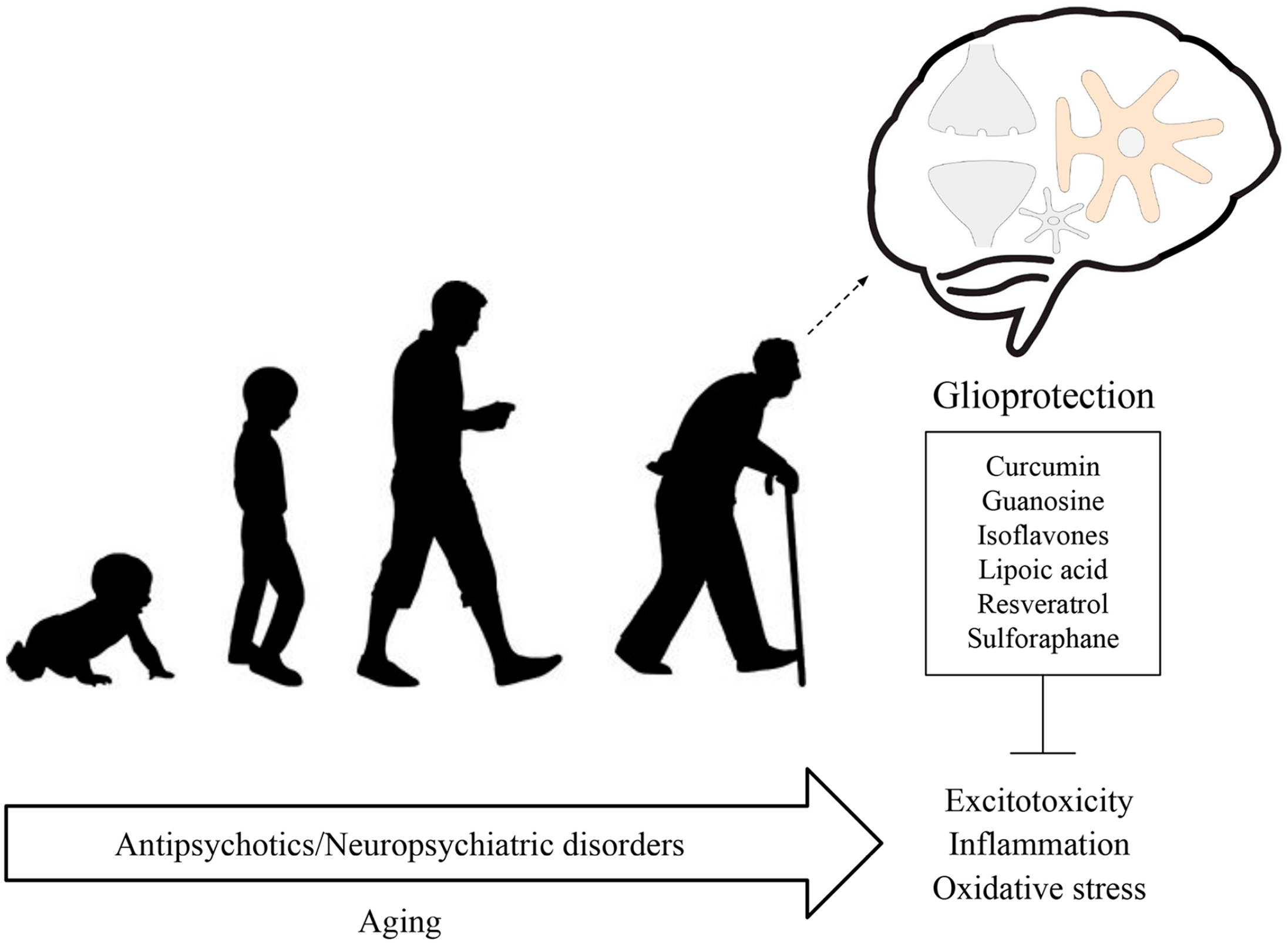
The Janus face of antipsychotics. Glial cells display key roles in the pathogenesis and progression of neuropsychiatric disorders. Accumulating evidence has shown that antipsychotics can affect glial functions. The long-term use of these medications, in association with age, may lead to changes in inflammatory response, oxidative stress, and glutamate homeostasis imbalance/excitotoxicity. Of note, all these processes are also closely related to neuropsychiatric disorders. Therefore, glioprotection (glial-based preventive/therapeutic strategies) might emerge to avoid such detrimental effects.
Footnotes
Authors’ Contributions
IS, AdS, LDB, C-AG, JS, and AQ-S conceptualized the review, written the original draft of the manuscript, revised, edited, and approved the manuscript.
Declaration Of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The authors are supported by the Brazilian funding agencies Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq); Coordenação de Aperfeiçoamento Pessoal de Nível Superior (CAPES); Fundação de Amparo à Pesquisa do Estado do Rio Grande do Sul (FAPERGS); Instituto Nacional de Ciência e Tecnologia para Excitotoxicidade e Neuroproteção (INCTEN/CNPq).