
Abstract
Pan, Cuiping, Qunyuan Wang, Minglei Zhang, Ruizhe Li, Yining Liu, and Zongbin Li. Role of Mitochondrial tRNALeu 12320 A>G Mutation on Heart Rate Variability and Blood Pressure at Low Altitude and in a High-Altitude Hypoxic Environment. High Alt Med Biol. 00:00–00, 2025.
Introduction:
Mutations—gene mutations—can exert dual, environment-dependent effects on cardiovascular disease.
Materials and Methods:
We analyzed clinical data and mitochondrial DNA from 84 young Han Chinese men ascending from low-altitude to high-altitude regions (>4,000 m).
Results:
Nine subjects harbored the mitochondrial tRNA A12320G mutation. Notably, none of these 9 mutation carriers developed acute mountain sickness (AMS), whereas 34 (40.5%) of the 84 volunteers in the cohort were diagnosed with AMS, indicating a significant protective association (p = 0.009). In the low-altitude environment, time domain parameters of heart rate variability (HRV) in the mutant volunteers were significantly lower than those in the unmutated volunteers (SDNN: 141.76 ± 30.75 ms vs.169.08 ± 32.83 ms, p = 0.020; RMSSD: 29.70 ms vs. 41.35 ms, p = 0.022; HRV triangular index: 41.8% vs. 52.4%, p = 0.024; SDSD: 37.4 ms vs. 50.5 ms, p = 0.016; PNN50: 3.1% vs 7.6%, p = 0.008); 24-hour mean systolic blood pressure (118.63 ± 9.54 vs. 113.39 ± 6.30 mmHg, p = 0.029), nocturnal mean diastolic blood pressure (67.37 ± 7.91 vs. 61.11 ± 7.08 mmHg, p = 0.015) were also higher; HRV and blood pressure were not statistically different between the mutant and nonmutant groups in the high-altitude environment. Hematological analysis revealed mutants at high altitude had significantly lower plateletcrit (0.23% vs. 0.26%, p = 0.019) and total protein (72.78 ± 3.84 g/L vs.75 .65 ± 3.0 g/L, p = 0.008).
Conclusion:
The A12320G mutation acts as a cardiovascular risk factor (reduced HRV, increased blood pressure) at low altitude, whereas at high altitude it was associated with a reduced incidence of AMS, possibly through metabolic adaptive mechanisms, reflecting the environmentally dependent dual roles of this gene effect.
Introduction
Many people arrive at high altitude for recreation, work, or military activities, and as the altitude increases, the partial pressure of oxygen decreases (Gatterer et al., 2024). After one to five days following an altitude increase to ≥2,500 m, individuals are at risk for acute mountain sickness (AMS). Its typical symptoms include headache, dyspnea, nausea, and sleep disturbances, etc. Most of its symptoms is self-limiting, and patients will gradually adapt to the hypoxic environment after resting for a few days (Luks et al., 2017). In severe cases it can progress to high-altitude pulmonary edema or high-altitude cerebral edema, which is a serious threat to life (Mehta et al., 2008). In recent years, the prevention of AMS and prediction of individual susceptibility have become the focus of clinical and scientific research. Genetic studies have revealed the complex pathogenesis of AMS, involving polymorphisms in hypoxia-adapted genes (e.g., EPAS1, EGLN1) (Zhang et al., 2020; Buroker et al., 2012), oxidative stress pathways (e.g., SOD2, HMGB1), and genes related to autonomic regulation (Pham et al., 2022; Bao et al., 2002; MacInnis et al., 2011; Domingos et al., 2020). However, the role of mitochondrial gene variants in AMS and their environmental dependence remain under-elaborated.
Mutations in mitochondrial DNA (mtDNA) of mitochondria, which serve as the energy factories of the cell, may directly lead to impaired energy metabolism by interfering with the oxidative phosphorylation process, which in turn affects the function of energy-consuming tissues, such as heart and muscle (Lawless et al., 2020). Among them, tRNA genes are hotspot regions for mtDNA mutations, accounting for more than 50% of reported pathogenic mutations (Taylor and Turnbull, 2005). For example, the A12320G mutation in the mitochondrial leucine tRNA (MT-TL2) gene has been shown to be associated with mitochondrial myopathy (Clark et al., 1997). Notably, this mutation is located in the TΨC environmentally guarded region of the tRNA and may affect mitochondrial translational efficiency by disrupting the secondary structure of the tRNA, ultimately leading to defective function of respiratory chain complexes such as cytochrome c oxidase (CCO) (Suzuki et al., 2011). However, whether this mutation affects physiological regulation in high-altitude environments through autonomic imbalance or metabolic adaptation mechanisms has not been systematically investigated.
Previous studies have largely focused on the pathological effects of gene mutations in a single environment, overlooking the dynamic modulation of gene expression and phenotype by environmental stressors-such as the contrast between low- and high-altitude conditions. Our previous research has shown that young Han Chinese males who did not experience acute altitude sickness carry the mitochondrial A12320G mutation, while volunteers who experienced acute altitude sickness did not carry the mutation, and its cardiovascular effects (e.g., on HRV and blood pressure) have not been explored (Li et al., 2022). In addition, although HRV, an important indicator for assessing autonomic function, has been shown to be closely associated with high-altitude response adaptive capacity (Karinen et al., 2012), how mitochondrial gene mutations affect cardiovascular vascular homeostasis through HRV changes remains unclear, and this research gap limits the possibility of parsing an individual’s high-altitude adaptive capacity from a gene-environment interaction perspective.
Materials and Methods
Participants
This project included 84 Chinese male adults between the ages of 18 and 35 years, none of whom had experienced high-altitude exposure. Other exclusion criteria included any history of primary headache, brain tumor, heart failure, or chronic obstructive pulmonary disease. The study was approved by the Ethics Committee of the General Hospital of the Chinese People’s Liberation Army. The ethics committee approval number was “S2025-801-01”. Individuals in this article obtained written informed consent and verbal explanation of the study before participation.
Study design
Participants’ demographic data, including age, height, weight, and body mass index (BMI), were first collected. Ambulatory electrocardiograms and ambulatory blood pressure were also performed at sea level (Beijing) before departure, and after collecting this baseline data, participants traveled by train for 30 hours to reach Golmud City (2,800 m) in Qinghai Province. After waiting for several hours in Golmud, these participants arrived at their destination (4,000 meters) by bus within 3 hours. Recordings of ambulatory electrocardiography and ambulatory blood pressure monitoring were initiated at 2,800 m above sea level and terminated at the destination, in addition to measurements of ambulatory electrocardiography and ambulatory blood pressure in a hypobaric oxygen chamber environment.
Mutational analysis
Genomic DNA was isolated from participants’ whole blood using Genomic DNA Isolation Kits (Thermo Fisher Scientific, Minneapolis, Massachusetts, MA).The mitochondrial genome was enriched and purified by long polymerase chain reaction (PCR) using primers specific for the human mitochondrial genome. Amplification products were segmented by sonomechanical methods to 100–500 bp. Sequencing libraries were prepared according to the protocol of the SureSelect QXT Library Prep Kit (Agilent, California, CA). Exon captures were prepared according to the protocol of Agilent SureSelectQXT Target Enrichment for Illumina Multiplexed Sequencing version E0. Prior to hybridization, the quantity and quality of library DNA were assessed using an Agilent 2200 TapeStation. DNA libraries were mixed with capture probes for the target region using the SureSelect Human All Exon V6 kit. Hybridization was performed at 65°C for 1 hour to ensure adequate binding of the target region to the capture probe. The probe containing the target region is captured using streptavidin beads. The enriched library on the beads is then amplified by PCR. The enriched libraries were cleaned using Ampure XP beads (Agencourt, Boston, MA, USA) according to the SureSelect QXT protocol, and then analyzed by Agilent 2200 TapeStation and qPCR for quality control. Cleaned libraries were denatured and diluted to optimal concentrations and applied to the cluster generation step. Paired-end 2 × 150 bp sequencing was performed on an Illumina NovaSeq 6000 system using the Illumina NovaSeq 6000 S2 kit (300 cycles).
Observation indicators
AMS was assessed using the 2018 Lake Louise Questionnaire Score, which has been the most commonly used scale for diagnosing AMS since its release in 1991 (Ahluwalia and Underwood, 2023). Patients with AMS were categorized into mild, moderate, and severe grades based on four symptoms: headache, gastrointestinal symptoms, fatigue, and dizziness. The diagnostic criterion for AMS was a total score of 3 or more, including headache symptoms.
Statistical methods
Continuous variables are expressed as mean ± SD, quantitative data with nonnormal distribution are expressed as median and quartiles (p 25, p 75), and comparisons of categorical variables were performed using χ2 or Fisher’s exact test where appropriate. Normality was determined using the Kolmogorov-Smirnov test. For normally distributed continuous data, the t-test was used for comparison between groups. For nonnormally distributed continuous data, the Mann–Whitney U test was used for between-group comparisons. All analyses were performed using SPSS (version 22.0), and all tests were performed using the two-tailed method, with a two-tailed p-value of <0.05 considered significant.
Results
The development of AMS
Genetic analysis revealed that 9 out of 84 volunteers (10.7%) carried the A12320G mutation. According to the revised LLS criteria, 34 volunteers(40.5%)were diagnosed with AMS. A striking finding was that all 9 mutation carriers belonged to the AMS-negative group (n = 50). Conversely, the mutation was entirely absent (0%) in the volunteers who developed AMS. This distribution shows a statistically significant protective association of the A12320G mutation against AMS (Table 1).
Distribution of the A12320G Mutation Between AMS+ and AMS− Volunteers

P value was calculated using a two-tailed Fisher’s exact test, due to the small expected count in one cell. The A12320G mutation was present only in the AMS-group, indicating a significant negative association with AMS susceptibility (p = 0.009).
Baseline analysis for enrollment
A12320G is a mutation site present in AMS groups. Among the 84 enrolled volunteers, 9 individuals harbored A12320G gene mutations, accounting for 10.7% of the total.
There was no statistically significant difference in age, height, weight, and BMI between the two groups of volunteers upon enrollment (Table 2).
Basic Information of Two Groups of Volunteers
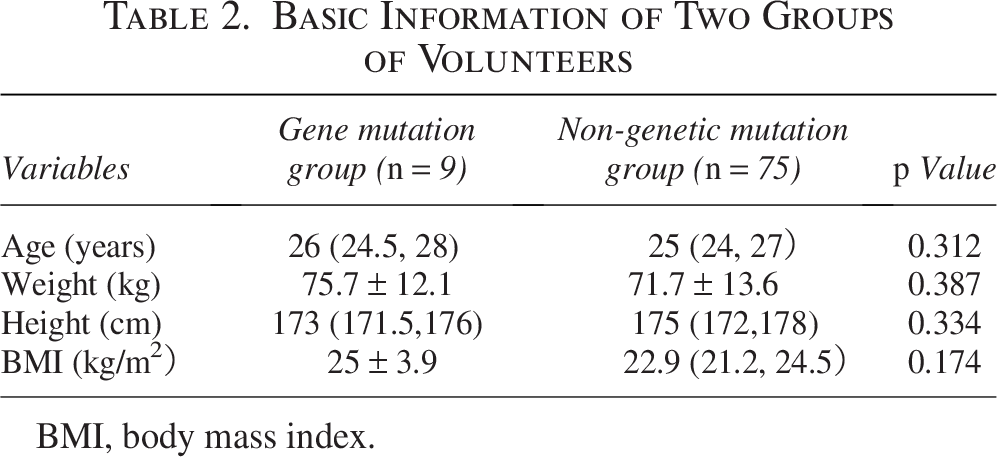
BMI, body mass index.
Analysis of results of Holter monitoring
When in a low-altitude environment, the time-domain parameters of HRV in mutant volunteers, SDNN (141.76 ± 30.75 ms vs. 169.08 ± 32.83 ms, p = 0.020), RMSSD (29.7 ms vs. 41.35 ms, p = 0.022), HRV trigonometric function (41.8% vs. 52.4%, p = 0.024), SDSD (37.4 ms vs. 50.5 ms, p = 0.016), PNN50 (3.1% vs. 7.6%, p = 0.008), SDANN (128.42 ± 31.64 ms vs. 155.36 ± 33.43 ms, p = 0.025), and minimum heart rate (54 bpm vs. 47 bpm, p = 0.03) were statistically significant and were all smaller than in the unmutated individuals (Table 3). After acute high exposure, the above indicators showed no statistical significance in the comparison between the two groups (Table 3).
Comparison of Holter Monitoring Parameters between Low and High Altitude Environments
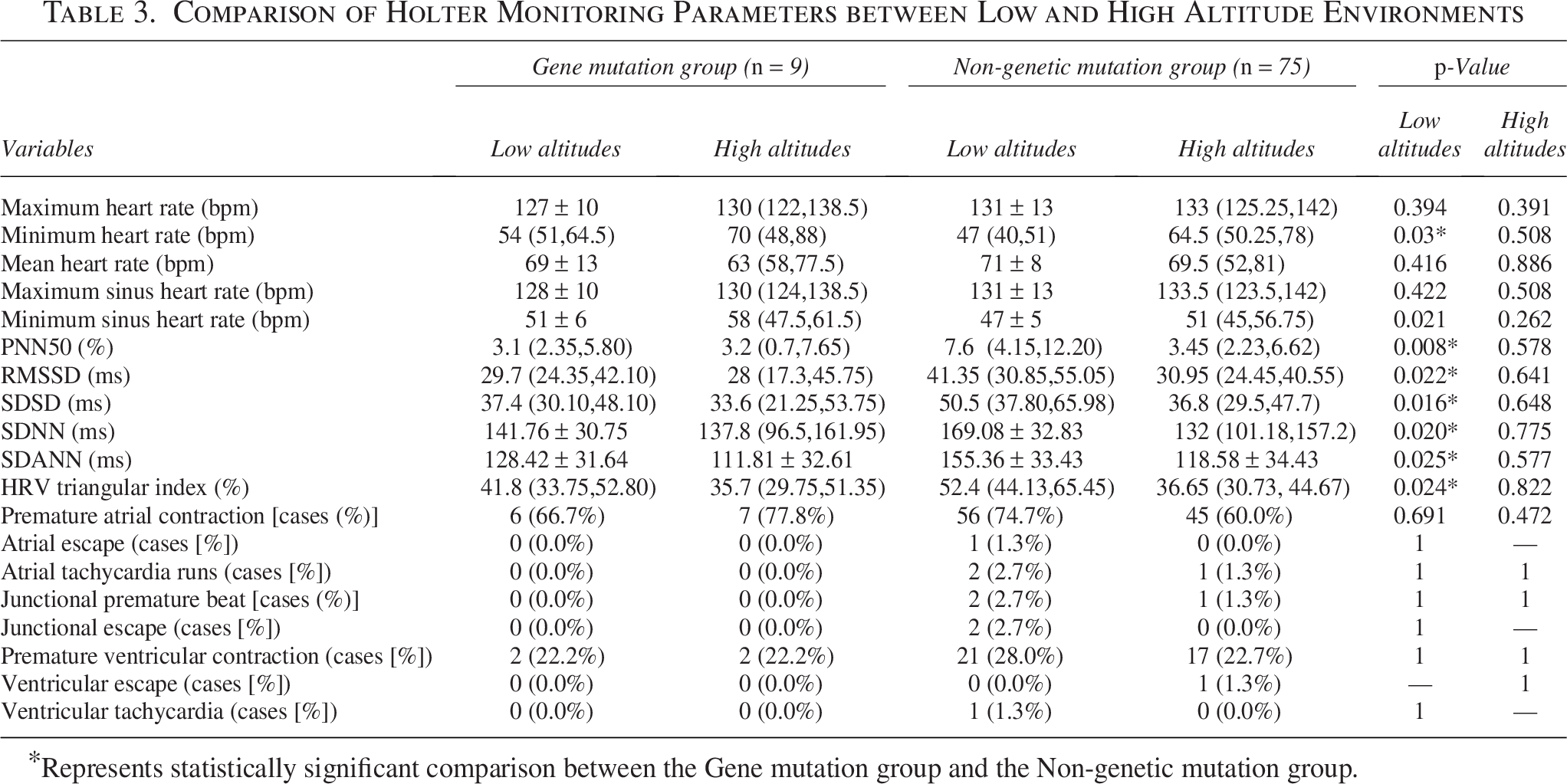
Represents statistically significant comparison between the Gene mutation group and the Non-genetic mutation group.
Analysis of results of blood pressure
When in a low-altitude environment, the 24-hour mean systolic blood pressure (118.63 ± 9.54 mmHg vs. 113.39 ± 6.30 mmHg, p = 0.029) and the nocturnal mean diastolic blood pressure (67.37 ± 7.91 mmHg vs. 61.11 ± 7.08 mmHg, p = 0.015) of the mutant volunteers were statistically significant and higher than those of the nonmutants (Table 4). After acute high exposure, the above indicators showed no statistical significance in the comparison between the two groups (Table 4).
Ambulatory Blood Pressure Monitoring in Low and High Altitude Environments
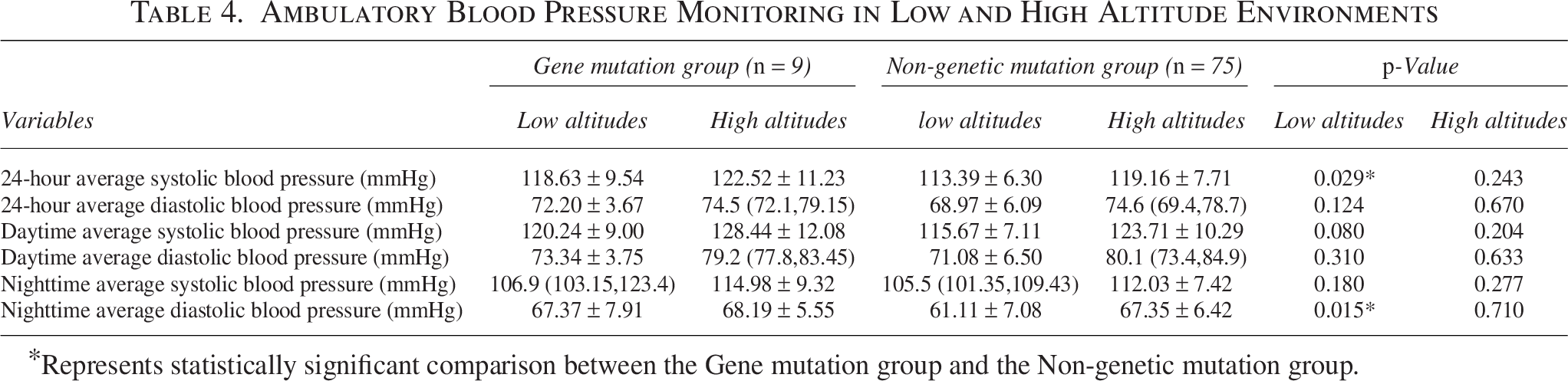
Represents statistically significant comparison between the Gene mutation group and the Non-genetic mutation group.
Analysis of results of blood routine and blood biochemistry
When in a low-altitude environment, platelet ratio (0.23% vs. 0.26%, p = 0.019) and total protein level (72.78 ± 3.84 g/L vs. 75.65 ± 3.0 g/L, p = 0.008) were significantly lower in the mutant group (Table 5). After acute high exposure, the above indicators showed no statistical significance in the comparison between the two groups (Table 5).
Blood Routine and Blood Biochemistry Data in Low and High Altitude
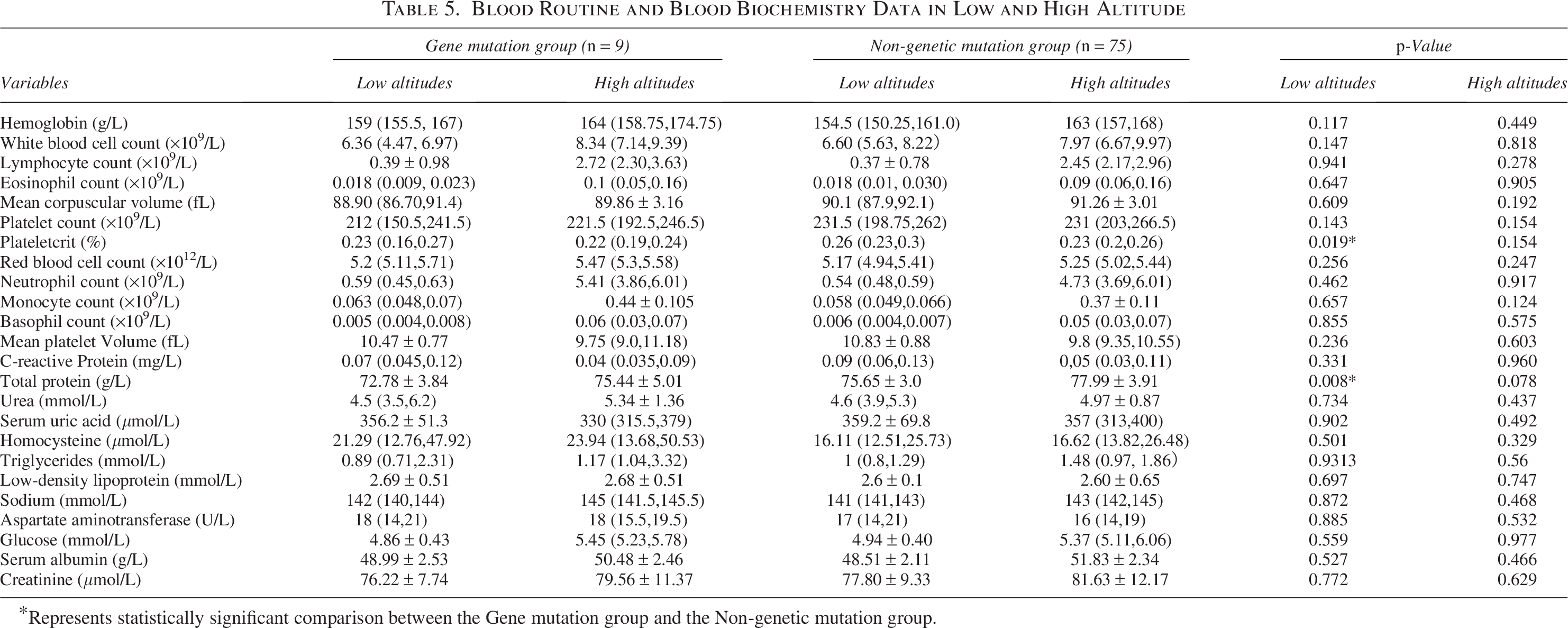
Represents statistically significant comparison between the Gene mutation group and the Non-genetic mutation group.
Analysis of results of ultrasound examination
When in a low-altitude environment, there was no statistically significant difference in systolic pulmonary artery pressure, TAPSE, and other indicators between the two groups of volunteers (Table 6).
Ultrasound Examination in Low and High Altitude

TAPSE, tricuspid annular plane systolic excursion; FAC, fractional area change.
Discussion
In the past, many cohort studies have elucidated the patterns, trends, and risk factors associated with altitude sickness. For example, Professor Shurong Han’s team conducted a retrospective cohort study among maintenance workers of the Qinghai-Tibet Railway, highlighting employment age and workplace altitude as significant risk factors for cardiac abnormalities and recommending intensified cardiovascular monitoring for high-altitude workers (Han et al., 2020). Similarly, Jiucun Wang’s research team carried out an observational cohort study identifying decreased oxygen saturation and altered heart rate as independent risk factors for high-altitude headache after acute high-altitude exposure (Wang et al., 2018). However, these studies did not explore the pathogenesis of AMS from the perspective of mitochondrial genetics or gene-environment dualism. Our study provides evidence for a dual, environment role of the mitochondrial tRNALeu 12320 A>G mutation. At low altitude, mutation carriers exhibited reduced heart rate variability (HRV), elevated blood pressure, and altered hematological parameters (e.g., decreased plateletcrit and total protein), suggesting heightened cardiovascular risk. In contrast, none of the carriers developed AMS at high altitude (>4,000 m), despite HRV and blood pressure levels comparable to non-carriers. This environmental duality underscores the importance of context-dependent genetic effects in physiological adaptation.
The observed reduction in HRV and elevation in blood pressure at low altitude may originate from mitochondrial dysfunction (Vercellino and Sazanov, 2022). The A12320G mutation, located in the conserved TC loop of the MT-TL2 gene, is known to impair tRNALeu stability and aminoacylation efficiency, potentially disrupting CCO assembly and oxidative phosphorylation (Suzuki et al., 2011). We speculate that inefficient ATP production in cardiomyocytes could destabilize cardiac electrical activity, thereby reducing HRV (Li et al., 2020), while compensatory sympathetic activation to meet energy demands may contribute to hypertension (Schroeder et al., 2003). These alterations are consistent with mitochondrial disease phenotypes characterized by arrhythmias and autonomic dysregulation (Yang, 2025). Furthermore, in the low-altitude setting, the decreased plateletcrit in the mutation group may reflect altered blood viscosity, and reduced total protein levels suggest an underlying metabolic abnormality (Semenza, 2012). Together, these findings provide multidimensional support for the elevated cardiovascular risk associated with this mutation under normoxic conditions.
Although there were no significant differences in HRV and blood pressure between mutant and nonmutant groups in the high-altitude environment, the incidence of AMS was significantly lower in the mutant group, a seemingly paradoxical phenomenon that may stem from the following adaptive mechanisms: firstly, anaerobic flexibility: mitochondrial dysfunction caused by the mutation could favor glycolysis over oxidative phosphorylation-a metabolic shift already induced by hypoxia (Gnaiger et al., 1998). This “pre-adaptation” might mitigate energy crises at high altitude, as seen in Sherpa populations with enhanced glycolytic capacity (Horscroft et al., 2017). Secondly, antioxidant defense: although mitochondrial defects often increase reactive oxygen species (ROS), epigenetic upregulation of antioxidant enzymes (e.g., SOD2) has been reported in hypoxia-tolerant models (Kowalczyk et al., 2021). The SOD2 rs4880 polymorphism, linked to AMS susceptibility, supports the role of ROS scavenging in altitudes adaptation (de Oliveira Paludo et al., 2021). Finally, HIF pathway synergy: The mutation might enhance hypoxia-inducible factor (HIF-1α) signaling, promoting erythropoietin and vascular endothelial growth factor expression to improve oxygen transport-a mechanism analogous to EPAS1 adaptations in Tibetans (Chen et al., 2025). However, direct evidence (e.g., measurements of HIF-1α protein levels or activity) is needed to test this hypothesis.
The dual environmental role of the A12320G mutation revealed in this study has important implications for individualized health management and high-altitude medicine. Carriers could be screened for enhanced cardiovascular risk (e.g., via ambulatory blood pressure, HRV screening) in low-altitude environment. Conversely, the potential adaptive advantage in high-altitude environments may inform personalized risk assessments for sojourns to high altitude, though this could be evaluated in combination with other genetic markers (e.g., EPAS1, EGLN1 polymorphisms) to comprehensively predict AMS susceptibility. At the same time, this study provides an empirical case for “opposite effects of the same gene across different environments” and underscores the necessity of incorporate environmental variables-such as altitude and partial pressure of oxygen-into genetic studies to improve phenotypic prediction accuracy. For instance, the mechanism underlying the dual role of the A12320G mutation could be further explored through environment-specific epigenetic analyses. Although our findings suggest a protective effect of the A12320G mutation against AMS at high altitude, several limitations must be acknowledged. Firstly, the small sample size of the mutation group (n = 9) limited statistical power to detect subtle effects in the high-altitude environment; future studies should expand cohorts, especially among AMS-susceptible populations. Secondly, direct mechanistic evidence-such as measurements of respiratory chain complex activity, ROS levels, or HIF pathway protein expression-is lacking and warrants validation via cellular or animal models. Third, the lack of data on smoking history and physical fitness, both known to influence HRV and blood pressure, represents a potential confounding factor that should be addressed in future research. Finally, the study cohort consisted only of Han Chinese males; future work should include females and other ethnic groups to improve generalizability. Integrative multi-omics and gene-environment interaction analyses may help further elucidate these mechanisms.
Conclusion
Our findings are consistent with a dual role for the A12320G mutation: it was associated with an elevated cardiovascular risk profile at low altitude, while being linked to a lower incidence of AMS at high-altitude, possibly due to compensatory adaptations. This finding not only provides a potential target for individual intervention in high altitude medicine but also provides an important reference for the gene-environment research paradigm of complex diseases. In the future, it is necessary to combine multi-omics technology with cross-environmental cohorts to further elucidate the molecular mechanism and clinical translational value.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by grants from Capital’s Funds for Health Improvement and Research (CSH2022-1G-5071) and the Hygiene and Health Development Scientific Research Fostering Plan of Haidian District of Beijing (HP-2021–03-80602). All authors declare that they have no conflicts of interest.