
Abstract
Chen, Yanli, Suying Zhu, Doudou Hao, Zhiyou Shi, Yang Zhong, Suyuan Wang, and Yunhong Wu. Time-Dependent Effects of Chronic Hypoxic Exposure on Microarchitecture of Different Skeletal Sites in Mice. High Alt Med Biol. 00:00–00, 2026.—High-altitude hypoxia accelerates bone loss by disrupting bone metabolic homeostasis. Currently, hypoxic effects on bone and their link to exposure time remain undefined. This study aimed to systematically evaluate the dynamic effects of chronic hypoxic exposure on bone homeostasis and bone microstructure in different skeletal sites through in vivo and in vitro experiments. Male C57BL/6J mice were exposed to 5,500 m simulated hypoxia for 16 weeks. Micro-CT analyzed cervical (C3–C4), lumbar (L4–L6) vertebrae, and femurs. Histopathological analysis assessed bone tissue and multiorgan pathology. In vitro experiments, a 5% hypoxic culture system was established to detect the proliferation and differentiation capabilities of MC3T3-E1 osteoblasts. After 12 weeks of hypoxic exposure, compared with the control group, the trabecular separation (Tb.Sp) of the mouse femur was significantly increased, and the bone mineral content was significantly decreased. After 16 weeks of exposure, the trabecular bone volume/tissue volume (Tb.BV/TV) and bone mineral density of the cervical vertebrae, lumbar vertebrae, and femurs were significantly reduced. H&E staining showed osteoblasts in multiple bones of hypoxic mice shifted to quiescent states, and long-term hypoxia induced multi-organ damage (pulmonary, renal, colonic, skeletal muscle). 5% hypoxia significantly downregulated collagen type I and alkaline phosphatase (ALP) mRNA levels in MC3T3-E1 cells, inhibited ALP activity and mineralized nodule formation, and reduced osteoblast differentiation capacity. Chronic hypoxic exposure induces bone loss in multiple skeletal sites of mice in a time-dependent manner, with femur showing significantly higher sensitivity to hypoxic exposure than the cervical and lumbar vertebrae. The underlying mechanism may be related to hypoxia inhibiting osteoblast function and disrupting the balance of bone metabolism. Long-term hypoxia also causes pathological damage to multiple organs. This study provides evidence for clarification of the pathological mechanism of hypoxic bone injury and lays a foundation for the subsequent development of targeted intervention strategies for hypoxia-related bone diseases.
Introduction
With the increase in altitude, the atmospheric oxygen partial pressure decreases in a gradient, leading to a significant reduction in oxygen content per unit volume of air. This hypoxic environment can trigger compensatory changes in multiple systems of the human body (Mallet et al., 2023). Although such adaptive responses help the body maintain basic physiological functions under the conditions of low pressure and hypoxia at high altitudes, they may also exert potential negative effects on physiological processes such as bone metabolic homeostasis (Chen et al., 2022).
The mechanism by which high-altitude hypoxia acts on the skeletal system is relatively complex. Existing studies have shown that hypoxia may disrupt bone metabolic homeostasis by inhibiting osteoblast activity and promoting osteoclast differentiation (Hannah et al., 2021). However, because differences in factors such as hypoxic models, experimental animal species, and intervention durations, current research conclusions remain inconsistent. For instance, most studies have demonstrated that short-term exposure to hypobaric hypoxia can cause a decrease in bone mineral density (BMD) and bone mineral content (BMC) of the femur in rats, as well as damage to bone microstructure (Wang et al., 2020; Wang et al., 2017). In contrast, other studies have found that intermittent hypobaric hypoxia can increase the BMD of the spine in mice (Guner et al., 2013). In in vitro experiments, hypoxia may not only inhibit osteoblast differentiation (Zhang et al., 2024) and promote osteoclast formation (Tan et al., 2021), but also attenuate hypoxia-induced cell death by upregulating the expression of hypoxia-inducible factor-1α (HIF-1α) (Xu et al., 2015). Additionally, HIF-1α can promote angiogenesis and osteogenesis by upregulating the level of vascular endothelial growth factor (Hiraga, 2018; Wang et al., 2007).
Controversies also exist regarding the association between hypoxia and BMD in human studies. Some scholars have found that the BMD of residents living in areas at an altitude of 2,500–4,500 m tends to decrease with increasing altitudes (Zuo et al., 2022; O’Brien et al., 2018). However, other studies have shown that there is no significant difference in BMD between high-altitude residents and plain residents; in some cases, high-altitude residents even have higher BMD (liu, 2017). This contradiction may be related to confounding factors such as the duration of high-altitude adaptation, age, physical activity level, and nutritional status of the population.
Notably, bones in different parts of the human body differ in anatomical structure, weight-bearing function, blood supply, and cell composition, leading to significant site-specific differences in their bone mass regulation mechanisms (Ambrosi et al., 2025). Current studies on high-altitude hypoxia and bone metabolism mostly focus on traditional sites such as the femur, tibia, or lumbar spine (Yan et al., 2022; Brent et al., 2022b; Hao et al., 2023). However, it is particularly important to note that there is an essential difference in the bone loading pattern between humans (bipedal upright posture) and mice (quadrupedal crawling posture) (Reitmaier and Schmidt, 2020). The cervical vertebrae of mice bear significantly higher mechanical loads than other parts because the need to continuously support the weight of the head and the stress generated by movement (Smit, 2002). This unique mechanical environment may result in a distinct response mechanism of the cervical vertebrae to hypoxic stimulation compared with weight-bearing bones (e.g., femur) or the lumbar segment of the spine, making them a key research target for exploring the regional heterogeneity of hypoxic bone injury. Existing studies have initially revealed contradictions regarding such site-specific differences: some experiments have shown that trabecular bone damage in the lumbar spine is the most significant under hypobaric hypoxia (Brent et al., 2022a), while other studies have found that the changes in the microstructure of the cervical vertebrae in mice are more obvious than those in the lumbar spine (Teng et al., 2023). This divergence strongly suggests that the effect of hypoxic exposure on bones not only exhibits time dependence but also has clear site-specific heterogeneity. The difference in responses between weight-bearing bones and axial bones awaits systematic investigation.
Based on the above research background, this study proposes the scientific hypothesis that chronic hypoxic exposure induces bone loss in multiple skeletal sites of mice and disrupts bone metabolic homeostasis in a time-dependent and site-specific manner. In this study, MC3T3 cells were cultured under hypoxic conditions to investigate the effects of hypoxia on osteoblasts, and a normobaric hypoxic environment simulating an altitudes of 5,500 m was established, and 25-week-old male C57BL/6J mice were used as the model. The mice were continuously exposed to hypoxia for 16 weeks. Combined with quantitative analysis using micro-CT, the changes in bone microstructure of the cervical vertebrae, lumbar spine, and femur, as well as pathological changes in multiple tissues, were observed. The objectives of this study were to clarify the damage to the bone microstructure of multiple skeletal sites in mice caused by the hypoxic environment, and to further explore whether the effect of hypoxia on bones is related to osteoblast activity and the expression of key genes. This study aims to elucidate the “time-site” dual regulatory characteristics of hypoxic bone injury and provide experimental evidence and theoretical references for the prevention and treatment of hypoxia-related bone diseases.
Materials and Methods
Experimental animals
Twenty male C57BL/6J mice, aged 25 weeks were purchased from GemPharmatech. (Jiangsu, China). All mice were housed in a standardized animal facility with a temperature of 20 ± 2°C, relative humidity of 60 ± 5%, and a 12-hour light/12-hour dark cycle. They had free access to standard pellet feed (Beijing Ke’ao Xieli Feed Co., Ltd.) and sterile water. After 1 week of adaptive feeding, the mice were randomly divided into two groups using a random number table (n = 10 per group): Normoxic control (NC) group: Housed at an altitude of 500 m (oxygen concentration: 21%), with 5 mice per cage (2 cages total). Hypoxic intervention (HI) group: Housed in a normobaric hypoxic environment simulating an altitude of 5,500 m (oxygen concentration: 10.5%), with 5 mice per cage (2 cages total).
HI protocol and specimen collection
A normobaric hypoxic animal system (Model: Ox-100, Shanghai Tawang Intelligent Technology) was used for HI. This system consists of a hypoxic chamber (dimension: 750 × 505 × 320 mm), a gas concentration controller, and a nitrogen generator. The mouse cages had a size of 318 × 202 × 135 mm. The oxygen concentration in the HI group was maintained at 10.5% by continuous injection of nitrogen into the hypoxic chamber. Parameters such as oxygen concentration, CO2 concentration, and humidity inside the chamber were monitored in real-time. Bedding and cages were changed once a week; during the replacement process, the HI group was briefly exposed to normoxic conditions, while the set hypoxic concentration was maintained continuously at other times. Considering potential individual differences or attrition in animal experiments, the final sample size was rounded up to 10 mice per group. After 16 weeks of HI, the mice were euthanized by cervical dislocation. The cervical vertebrae, lumbar vertebrae, bilateral femurs, and tibias were quickly dissected and fixed in 4% paraformaldehyde for subsequent ex vivo micro-CT scanning, histopathological, and molecular pathological analyses. Meanwhile, tissues including the heart, liver, lung, kidney, colon, rectus femoris muscle, and femur were collected and fixed in 4% paraformaldehyde for histopathological analysis.
In vivo micro-CT scanning and bone microstructure analysis
Before HI (baseline) and at week 12 of intervention, in vivo micro-CT scans were performed, the mice were anesthetized by isoflurane inhalation (induction concentration: 2%, maintenance concentration: 1.5%, RWD Life Science, China), fixed on a 37°C constant-temperature scanning stage, and subjected to in vivo scanning using a high-resolution micro-CT system (NMC-100, Pingsheng Medical Technology Co., Ltd., Kunshan, China). The scanning parameters were set as follows: voltage = 80 kV, current = 0.08 mA, rotation step = 0.36°, voxel size = 25 μm, integration time = 50 ms, and 360° projection images were acquired. After 16 weeks of HI, the fixed samples of cervical vertebrae, lumbar vertebrae, and femurs were subjected to ex vivo scanning using the micro-CT system. The scanning parameters were: voltage = 90 kV, current = 0.05 mA, voxel size = 15 μm, rotation step = 0.36°, integration time = 50 ms, and 360° projection images were acquired. Three-dimensional reconstruction and bone microstructure parameter analysis were performed using Avatar software (Version v1.6.9.3, Pingsheng Medical Technology Co., Ltd.). Image segmentation was conducted using a fixed threshold (800 mg HA/cm³). Regions of interest were selected as follows: For the femur: A 1 mm-high cylindrical region 1 mm below the growth plate of the femoral metaphysis was selected to analyze trabecular bone parameters; a 1 mm-long cortical bone segment 6 mm away from the growth plate was selected to analyze cortical bone parameters. For the cervical and lumbar vertebrae: The middle region of each vertebra was selected to analyze trabecular bone parameters.
The analyzed parameters included trabecular bone mineral density (Tb.BMD), trabecular bone volume/tissue volume (Tb.BV/TV), trabecular number (Tb.N), trabecular thickness (Tb.Th), trabecular separation (Tb.Sp), cortical thickness (Ct.Th), and cortical bone mineral density (Ct.BMD).
Histopathological analysis
Tissue specimens (heart, liver, lung, kidney, colon, rectus femoris muscle, femur, lumbar vertebrae, cervical vertebrae) fixed in 4% paraformaldehyde were subjected to histopathological analysis (n = 4 per group). They were processed using a fully automatic dehydrator according to the following protocol: 70% ethanol for 4 hours, 80% ethanol for 2 hours, 90% ethanol for 2 hours, 95% ethanol for 2 hours, two changes of absolute ethanol (1 hours each), two changes of xylene (30 minutes each), and three changes of paraffin (60°C, 1.5 hours each). After paraffin embedding, 4 μm-thick sections were prepared using a rotary microtome. The sections were deparaffinized with xylene, stained with hematoxylin and eosin (H&E), dehydrated, cleared, and mounted with neutral balsam. Images were acquired and analyzed using a 3DHISTECH (Hungary) Pannoramic SCAN digital slide scanner.
Cell viability assay
Cell viability was evaluated using the Cell Counting Kit-8 (CCK-8, Oriscience Biotechnology Co., Ltd., China). MC3T3-E1 cells were seeded into 96-well plates at a density of 6 × 10³ cells per well, with 100 µl of medium added to each well. The cells were precultured under normoxic conditions for 24 hours, then continued to be cultured under normoxic (21% O2) or hypoxic (5% O2) conditions. At 12, 24, 48, and 72 hours of culture, 10 µl of CCK-8 solution was added to each well, and the plates were incubated at 37°C in the dark for 1.5 hours. The absorbance value at 450 nm was measured using a microplate reader (SpectraMax i3X, Molecular Devices).
Osteogenic induction culture
The mouse preosteoblastic cell line MC3T3-E1 was purchased from the Cell Bank of the Chinese Academy of Sciences. The cells were cultured in α-MEM medium (Gibco, USA) supplemented with 10% fetal bovine serum (FBS, Gibco, USA) and 1% penicillin–streptomycin double antibody (Solarbio, China) in a conventional cell incubator (HealForce Likang, China) at 37°C with 5% CO2. The medium was changed every 2–3 days. The experiment was divided into two groups: Normoxic group: Maintained at 21% O2, 5% CO2, and 37°C. Hypoxic group: Cultured in a tri-gas incubator (Thermo Fisher Scientific, USA), with the oxygen concentration maintained at 5% by nitrogen injection; other conditions were consistent with the normoxic group. When the cell confluency reached 80%, the medium was replaced with osteogenic induction differentiation medium, which consisted of α-MEM basic medium, 10% FBS, 1% penicillin/streptomycin, 50 μg/ml ascorbic acid, and 10 mM β-glycerophosphate sodium (Solarbio, China). Alkaline phosphatase (ALP) staining (#C3250S, Beyotime Biotech Inc.) and Alizarin Red S (ARS) staining (#G1452, Beijing Solarbio Science & Technology Co.) were performed on days 14 and 21 of induction, respectively, to evaluate osteoblastic differentiation and mineralization. On day 14 of induction, total cellular RNA was extracted using a Total RNA Extraction Kit (#NB304-100T, Origene Technologies). cDNA was synthesized by reverse transcription using the PrimeScript RT reagent Kit (Takara, Japan). Quantitative real-time polymerase chain reaction (qPCR) was performed using TB Green Premix Ex Taq II (Takara, Japan) on a real-time fluorescent quantitative PCR system (CFX Connect, Bio-Rad) to detect the mRNA expression levels of osteogenic differentiation and related functional genes, including ALP (alkaline phosphatase), Collagen Type I Alpha-1 Chain (Col1agen-I), and Osteoprotegerin (OPG). β-actin was used as the internal reference gene. Primers were designed by Primerbank and synthesized by Sangon Biotech (Shanghai, China). The primer sequences are listed in Supplementary Table S1. The relative mRNA expression levels of target genes were calculated using the 2−ΔΔCt method.
Statistical analysis
Statistical analysis and graphing were performed using GraphPad Prism 9 software. Measurement data were expressed as mean ± standard deviation. Normality test (Shapiro–Wilk test) and homogeneity of variance test (Levene test) were conducted first. If the data conformed to normal distribution and homogeneous variance, the independent samples t-test was used; otherwise, the Mann–Whitney U test was used. A p-value <0.05 was considered statistically significant. Considering the relatively high rate of missing data that occurred due to animal mortality during the study. We ultimately performed independent-samples analysis rather than paired tests for the baseline and 12-week datasets to avoid statistical bias from sample exclusion and ensure the robustness of our results.
Sample size estimation
Sample size calculation was based on preliminary data regarding the effect of 12-week high-altitude exposure on mouse femoral BMD (86.3 ± 9.52 vs. 74.9 ± 7.23 mg/cm2) (Brent et al., 2022a). First, Cohen’s d effect size was calculated using the mean and standard deviation of each group. Combined with this effect size, a significance level (α = 0.05), and a group allocation ratio of 1:1, the minimum sample size required to achieve 80% statistical power (power = 0.80) (Vahidi et al., 2023) was determined to be 9 mice per group.
Results
Hypoxic exposure inhibits proliferation and differentiation of MC3T3-E1 osteoblasts
To investigate the effect of hypoxia on osteoblast functional activity, a 5% O2 hypoxic culture model was established in vitro, and the viability of MC3T3-E1 cells and indicators related to osteoblastic function were detected. Results from the CCK-8 assay showed that compared with the normoxic group (21% O2), the absorbance value of cells in the hypoxic group was significantly decreased at 72 hours of culture, indicating that hypoxia significantly inhibited the proliferative activity of MC3T3-E1 cells (Fig. 1A). ALP staining and ARS staining results revealed that the area of ALP-positive regions and mineralized nodules in the hypoxic group were significantly reduced, demonstrating that hypoxia markedly suppressed the differentiation and mineralization capabilities of osteoblasts (Fig. 1B,C). qPCR detection further showed that the mRNA expression levels of osteogenic differentiation and related functional genes (Col1α1 and ALP) were significantly downregulated under hypoxic conditions (Fig. 1D–E), while the expression of the OPG gene showed no significant difference (Fig. 1F).
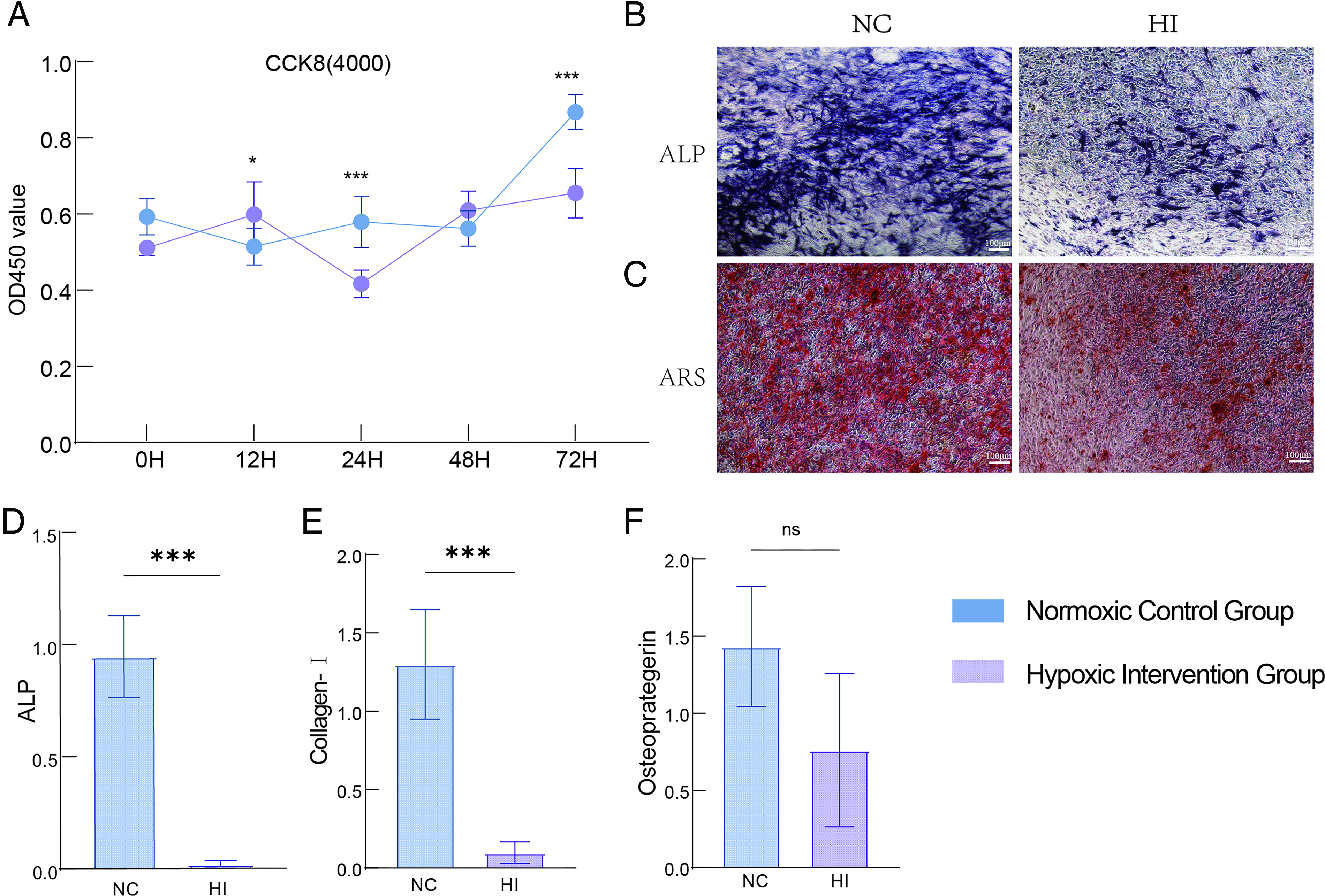
Effects of hypoxia on the viability and osteogenic differentiation of MC3T3-E1 cells.
Animal mortality
During the experiment, a total of 12 mice died, with 8 mice surviving until the end (4 mice per group). The causes of death were as follows: NC group: 1 mouse died of secondary infection due to skin laceration caused by group fighting at week 10; 3 mice died of respiratory depression due to excessive isoflurane anesthesia during in vivo Micro-CT scanning at week 12; 2 mice died within 24 hours after scanning. HI group: 3 mice died of hypoxic adaptation disorder at week 3 of hypoxic exposure; 2 mice died of respiratory depression due to excessive isoflurane anesthesia during in vivo micro-CT scanning at week 12; 1 mouse died within 24 hours after scanning.
Sustained 12-week hypoxic exposure induces femoral bone microstructure damage in mice
Baseline (preintervention) micro-CT scanning results showed no significant differences in bone microstructure parameters of the cervical vertebrae (C3, C4), lumbar vertebrae (L4, L5, L6), and distal femur between the two groups, indicating that the baseline bone status of mice in the two groups was basically consistent (Fig. 2A–H). After 12 weeks of HI, micro-CT quantitative analysis demonstrated that compared with the NC group: In the HI group, Tb.Sp was significantly increased (0.3370 ± 0.07692 vs. 0.3985 ± 0.03011, p = 0.0161) (Fig. 2C); The Tb.BV/TV (0.168 ± 0.075 vs. 0.061 ± 0.012, p <0.05 (Fig. 2B), Tb.BMC (2.256 ± 0.2117 vs. 1.782 ± 0.1850, p < 0.001) (Fig. 2G), Tb.BMD (0.204 ± 0.095 vs. 0.064 ± 0.003, p < 0.05) (Fig. 2H), was significantly decreased; No significant differences were observed in other parameters such as Tb.N. Additionally, all bone microstructure parameters of C3, C4, L4, L5, and L6 in the HI group showed no statistical differences compared with the NC group (p > 0.05) (Fig. 2A–H). These results indicated that 12 weeks of sustained hypoxic exposure mainly affected the femoral microstructure, but had no significant impact on the bone microstructure of the spine (cervical and lumbar vertebrae).
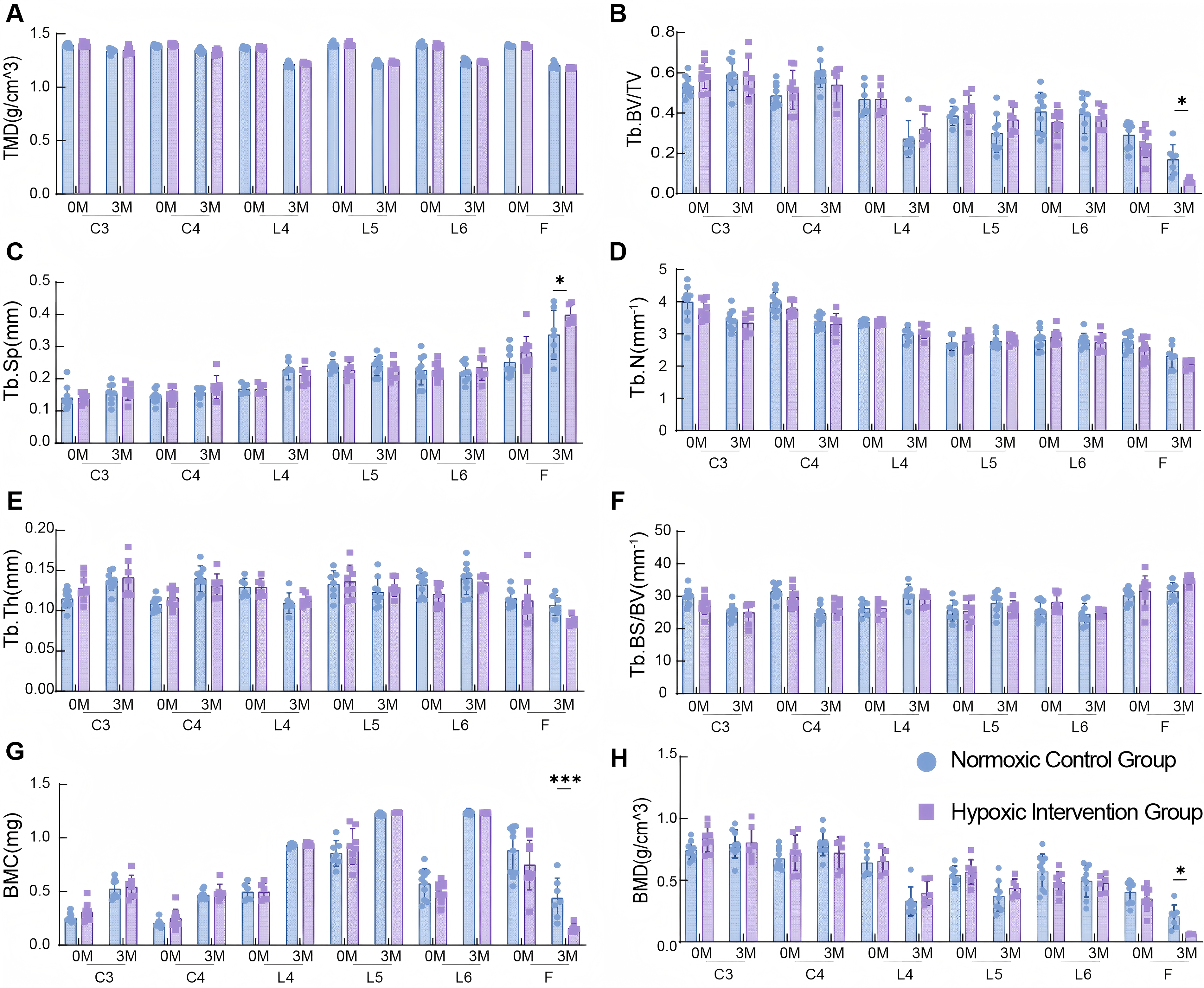
Effects of 12-week hypoxia intervention on bone microstructural parameters in different sites of mice.
Sustained 16-week hypoxic exposure induces Multisite bone microstructure damage in mice
After extending hypoxic exposure to 16 weeks, micro-CT three-dimensional reconstruction images showed that the HI group had bone loss and sparse trabecular structure in the cervical vertebrae, lumbar vertebrae, and femur (Fig. 3A). Specifically, compared with the NC group: In the HI group, the TMD (Fig. 3B), Tb.BV/TV (Fig. 3C), BMC (Fig. 3H), and BMD (Fig. 3I) of cervical vertebrae C3 and C4 were significantly decreased, while the Tb.BS/BV was significantly increased (Fig. 3D); The TMD (Fig. 3B) Tb.BV/TV (Fig. 3C) and BMD (Fig. 3I) of lumbar vertebrae (L4, L5, L6) were significantly reduced, with the most obvious changes in bone microstructure parameters observed in L5; For the femur, the trabecular TMD (Fig. 3B), Tb.BV/TV (Fig. 3C), Tb.Th (Fig. 3F), and BMD (Fig. 3I) in the HI group were significantly decreased, and Tb.BS/BV (Fig. 3D) was significantly increased; cortical bone parameters (Ct.Th, Ct.BMC, and Ct.BMD) also significantly decreased (Fig. 3J–L). No statistical differences in Tb.N and Tb.Sp of the cervical vertebrae, lumbar vertebrae, and femur were found between the two groups (Fig. 3E–F). H&E staining results showed that after 16 weeks of sustained HI: The trabecular bone of the cervical and lumbar vertebrae became thinner and atrophic, with reduced osteoblast activity; Osteoblasts in the femur showed a flattened morphology, suggesting weakened bone anabolic metabolism (Fig. 3M–O). Therefore, 16 weeks of sustained hypoxic exposure could induce bone microstructure damage in multiple sites (cervical vertebrae, lumbar vertebrae, and femur) of mice, with site-specific differences in the degree of damage.
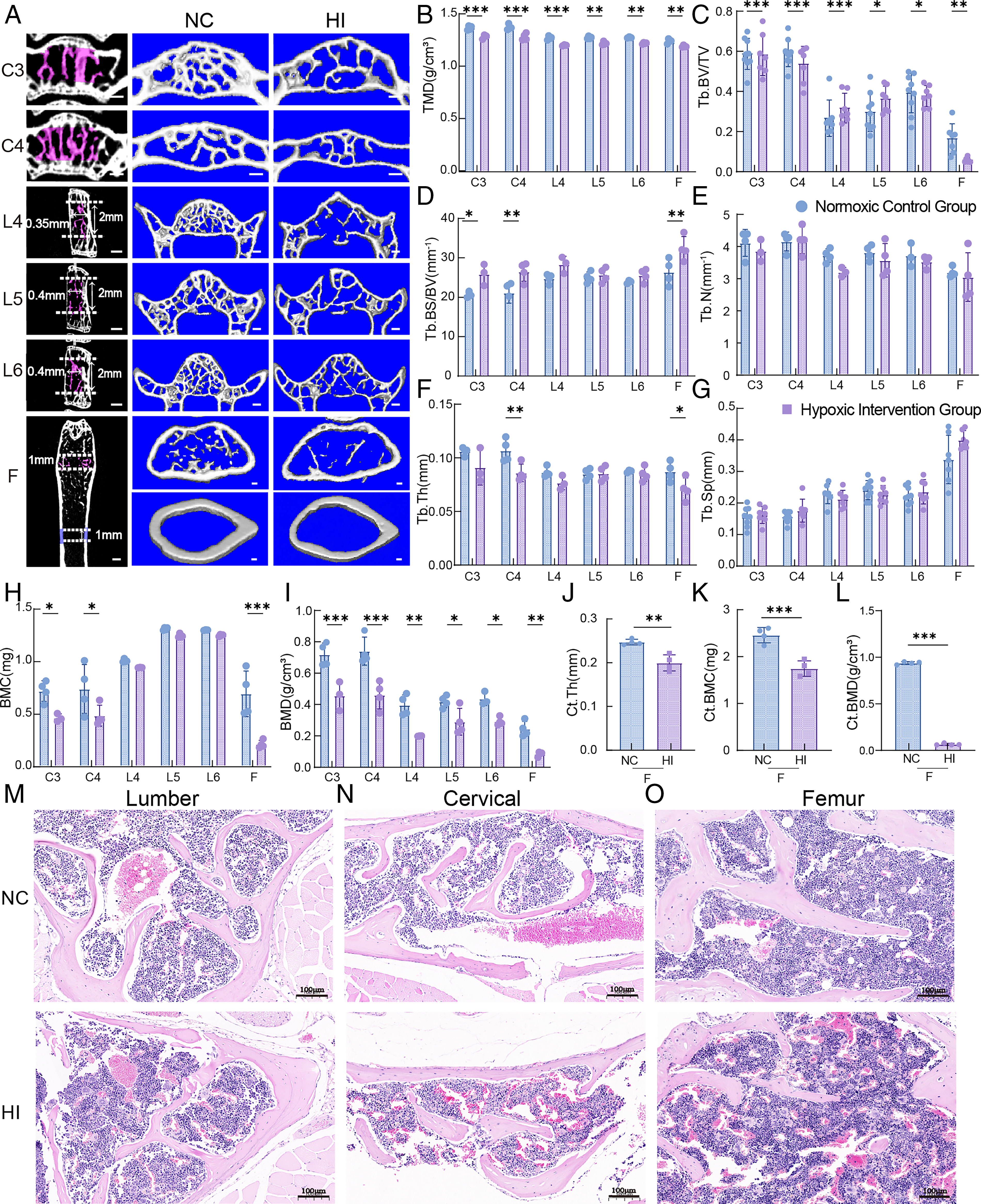
Effects of 16-week hypoxia intervention on bone microstructural parameters and histomorphology in different sites of mice.
Sustained 16-week hypoxic exposure induces pathological damage in multiple organs of mice
H&E staining was used to observe the effect of 16-week hypoxic exposure on major organs of mice. Compared with the NC group, the HI group showed the following pathological changes (Fig. 4A–F): Heart: Disordered arrangement of myocardial fibers (Fig. 4A); Liver: Neatly arranged hepatocytes, clear hepatic lobule structure, and no significant abnormalities (Fig. 4B); Lung: Local congestion or hemorrhage (red arrows), accompanied by a small amount of inflammatory cell infiltration (black arrows) and mild fibrous hyperplasia (dark green arrows) (Fig. 4C); Kidney: Vascular congestion in renal tissue (red arrows), pyknosis of some renal tubular epithelial cells (blue arrows), and eosinophilic substances in the tubular lumen (yellow arrows) (Fig. 4D); Skeletal muscle: A small amount of inflammatory cell infiltration (black arrows) (Fig. 4E); Colon: Significant pathological damage, including local crypt injury and degeneration (light blue arrows), pyknosis of a small number of mucosal epithelial cells (blue arrows), and occasional exfoliated cells in the intestinal lumen (green arrows) (Fig. 4F).
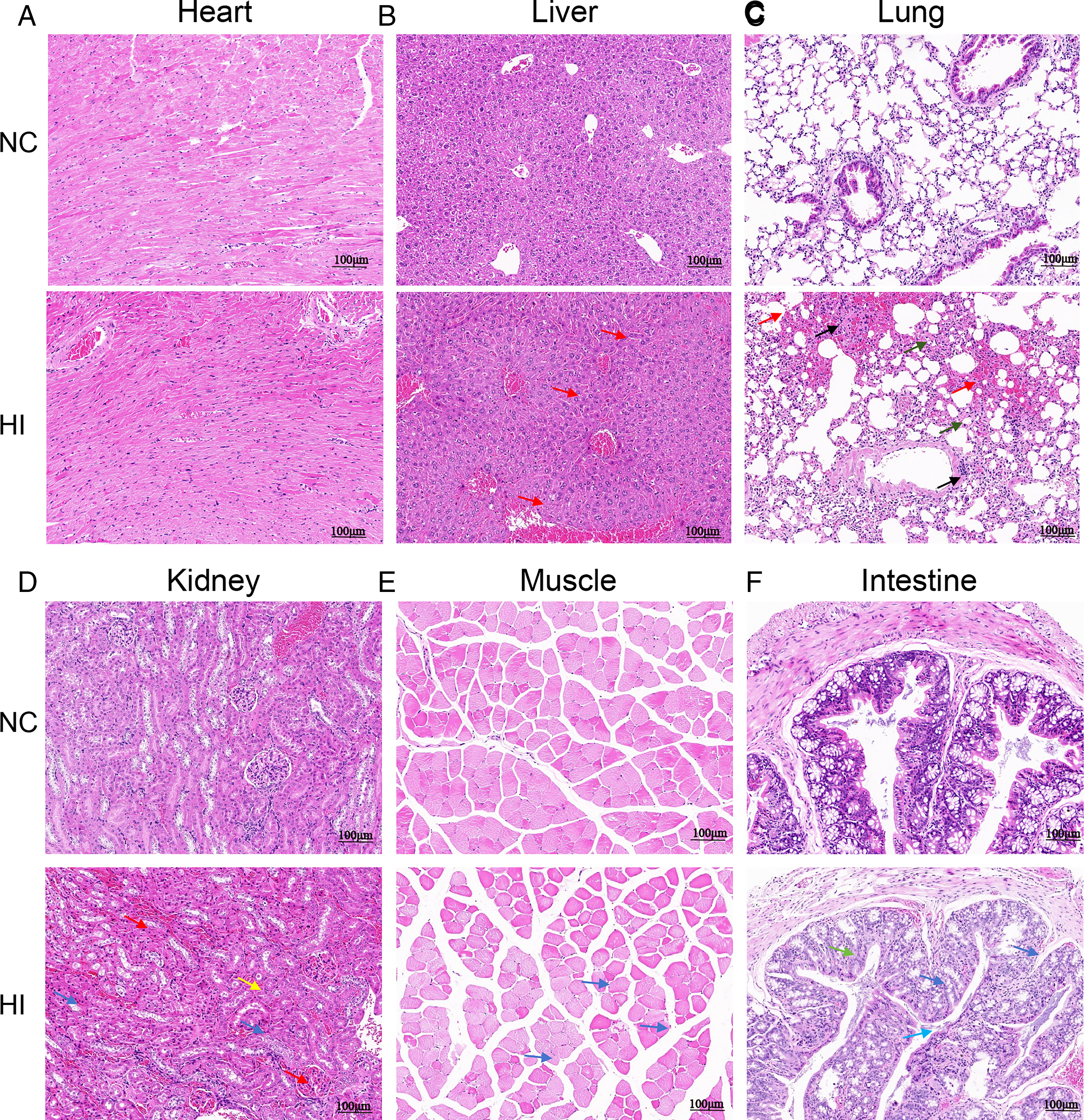
Morphological changes in major organ tissues of mice after 16 weeks of hypoxia exposure.
Discussion
The global aging population has intensified the burden of osteoporosis, making it a severe public health challenge (Adami et al., 2022). In high-altitude regions, the hypobaric and hypoxic environment further increases the risk to bone health (Hao et al., 2025). Therefore, systematically clarifying the effects of hypoxia on bone metabolism and its underlying mechanisms is of great significance for the prevention and treatment of hypoxic bone injury. In this study, 25-week-old male C57BL/6J mice were continuously exposed to a hypoxic environment simulating an altitude of 5,500 m (10.5% O2) for 16 weeks. The results confirmed that long-term hypoxic exposure induces multisite bone loss, with the femur (a weight-bearing bone) showing the highest sensitivity to hypoxia. These findings provide important evidence for research on high-altitude bone injury and the development of intervention strategies.
Previous studies have reported inconsistent conclusions regarding the effect of hypoxia on bone metabolism, which may be attributed to differences in experimental animal strains, ages, and hypoxic intervention conditions (Terrizzi et al., 2021; Yin et al., 2020; Durand et al., 2019). Research has shown that the skeletal development of laboratory mice is age-dependent: mice undergo a rapid growth phase from 0 to 2 months of age, and the peak BMD of trabecular and cortical bone is typically reached between 3 and 6 months (Buettmann et al., 2022). For example, the trabecular volume of C57BL/6 mice peaks at approximately 2 months of age and decreases annually thereafter (Liu et al., 2024); other studies have indicated that the trabecular volume of C57BL/6 mice declines after 6 months of age (Buie et al., 2008). Different mouse strains also exhibit varying sensitivities to bone metabolism interventions: in a glucocorticoid-induced bone loss model, the success rate of modeling in 7-week-old male BALB/c mice is significantly higher than that in age-matched C57BL/6 mice, which is associated with the higher sensitivity of BALB/c mice to glucocorticoids (Shidara et al., 2019). In studies on multiple myeloma-induced bone injury, NSG mice show more severe lumbar and femoral damage compared to C57BL/KaLwRij mice (Mehdi et al., 2021). C57BL/6J mice are widely used in bone metabolism research due to their clear genetic background and high genomic homology with humans. In this study, 25-week-old male C57BL/6J mice were selected as experimental subjects. At this age, mice have passed the rapid skeletal growth phase, and their bone mass is in a relatively stable plateau period. This selection effectively avoids interference from bone mass fluctuations caused by growth and development in young mice, ensuring accurate evaluation of the hypoxic intervention effect (Halloran et al., 2002). Recent studies have demonstrated that the impact of high-altitude hypoxic environments on the skeletal system exhibits significant time and altitude dependence. An experiment involving 6 weeks of intervention under conditions simulating an altitude of 2,900 m showed no significant structural changes in rat bones (Bozzini et al., 2013); however, when the exposure time was extended to 12 weeks and the altitude was increased to 5,500 m, significant femoral bone microstructure damage was observed in rats (Lezon et al., 2016). Consistent with these findings, the present study found that 12 weeks of continuous hypoxic exposure (simulating 5,500 m) caused significant damage to the femoral microstructure in mice. When the exposure time was extended to 16 weeks, the damage further spread to axial bones such as the cervical and lumbar vertebrae—consistent with conclusions from previous studies using SD rat models (Lezon et al., 2016). This suggests that long-term hypoxia disrupts the balance of bone remodeling, leading to systemic bone loss.
The present study revealed that after 12 weeks of hypoxic exposure, bone loss mainly occurred in the femur, with no significant damage observed in the lumbar or cervical vertebrae. This site-specific difference may be related to variations in mechanical load and metabolic demand among bones: as the primary weight-bearing bone, the femur may experience a more significant increase in metabolic demand under hypoxic conditions (Gunderson et al., 2020); in contrast, the metabolic demand of axial bones is relatively lower at rest. Site-dependent responses of bones to stimuli (including hypoxia, aging, and drugs) are widely observed. Studies have shown that age- and sex-related changes in bone properties are more pronounced in weight-bearing bones (e.g., femur and tibia) than in nonweight-bearing sites (e.g., ulna) (Mumtaz et al., 2020). Age-related changes in mouse trabecular bone also exhibit site specificity: for instance, the trabecular bone volume/tissue volume (Tb.BV/TV) of the proximal tibia decreases progressively after 4 months of age, while the age-related decline in the lumbar spine (L5) is more moderate, and the Tb.BV/TV of the caudal vertebra even increases with age (Willinghamm et al., 2010). In drug intervention studies, rapamycin damages the femoral trabecular bone, cortical bone, and L5 lumbar microstructure, with a more significant effect on the femur (Martin et al., 2021); in a rosiglitazone-induced osteoporosis model, sclerostin-neutralizing antibodies show a more potent protective effect on the femur than on the tibia (Farrell et al., 2021); mice with knockout of estrogen receptor α exhibit reduced cancellous and cortical bone mass in the tibia, vertebrae, and femur at 12 and 18 weeks of age (Melville et al., 2014). These studies collectively confirm that weight-bearing bones are more sensitive to interventions, which is consistent with the finding in the present study that the femur was the first site to show damage. However, some studies have reported results inconsistent with the present study. For example, one study found that trabecular bone damage in the lumbar spine was most prominent under hypobaric hypoxic conditions (Brent et al., 2022a). This discrepancy may stem from fundamental differences in experimental models: the aforementioned study used a hypobaric hypoxic model (simultaneously reducing air pressure and oxygen concentration), which differs from the normobaric hypoxic model in the present study (only adjusting oxygen concentration). Additionally, differences in experimental animal species and housing environment parameters may contribute to the divergent results. Another study reported that changes in the cervical vertebra microstructure of mice were more significant than those in the lumbar spine, but femoral bones were not examined, precluding direct comparison with the present study’s conclusion that the femur is the most sensitive site (Teng et al., 2023). These findings suggest that the effect of hypoxia on bones is regulated by multiple factors, including mechanical load, species differences, and hypoxic exposure conditions.
Hypoxia in high-altitude environments is a core pathogenic factor of high-altitude sickness, which can disrupt the body’s compensatory balance (e.g., hyperventilation, polycythemia) and trigger multi-system damage (Mallet et al., 2023). The present study found that 16 weeks of continuous hypoxic exposure caused varying degrees of pathological changes in the heart, lungs, kidneys, colon, and rectus femoris muscle of mice, with the most severe damage observed in the lungs, kidneys, and colon—consistent with previous research results (Ji et al., 2024). Mechanistically, long-term hypoxia can promote the development of pulmonary arterial hypertension (Sydykov et al., 2021) and induce autophagy in alveolar cells (Brito et al., 2020); the infiltration of inflammatory cells observed in high-altitudes pulmonary edema also indicates a close association between hypoxia and inflammation (El Alam et al., 2022). Furthermore, hypoxic exposure can cause kidney damage, which is linked to the development and progression of chronic kidney disease (Wang et al., 2022). In the intestinal tract, high-altitude environments reduce the oxygen concentration in the colonic lumen, and HIF-1α-mediated damage to the intestinal mucosal barrier exacerbates intestinal pathological changes (Cheng et al., 2022; Yan et al., 2024). Collectively, these findings indicate that hypoxia does not exert isolated effects on individual organs but induces significant systemic associations among multiorgan injuries.
This study has several limitations. First, only 25-week-old male C57BL/6J mice were used, and female mice and other strains were not included, which limits the generalizability of the conclusions. Second, because of experimental constraints, only normobaric hypoxia (rather than hypobaric hypoxia) was used, failing to fully simulate the environment at 5,500 m; additionally, only a single oxygen concentration was tested, precluding systematic exploration of the dose–effect relationship between oxygen concentration and bone injury. Third, intermittent “background noise” (50 decibels) generated by equipment during the study may have exerted confounding effects on bone metabolism indicators, leading to data dispersion and reduced result accuracy. Fourth, mouse mortality caused by hypoxia and accidental deaths of both hypoxic and control group mice due to excessive isoflurane anesthesia during micro-CT scanning resulted in insufficient sample size. This may explain why no significant differences were observed in some femoral microstructure parameters (e.g., trabecular number [Tb.N], trabecular separation [Tb.Sp]) between groups after 16 weeks of hypoxic exposure. Finally, considering the scanning time required for multisite in vivo imaging, animal welfare, and scanning resolution, different settings were used between the 12-week in vivo scan and the 16-week ex vivo scan, which limits the comparability of results at different time points to some extent. In future studies, stricter control of potential confounding variables will be implemented to ensure result accuracy and focus on the femur as the key site and conduct more refined in vivo scanning and analyses with a voxel resolution below 15 μm.
In conclusion, this study suggests that in a chronic hypoxic environment, weight-bearing bones such as the femur can serve as sensitive indicators of early bone injury, and the degree of bone loss is positively correlated with hypoxic exposure duration. Therefore, for populations with short-term hypoxic exposure, priority should be given to protecting bone tissues with high metabolic activity; for those with long-term exposure, comprehensive prevention, and control strategies are needed to address multisite bone loss.
Authors’ Contributions
Y.C. and S.Z. contributed equally as co-first authors; they were contributed significantly to the methodology, curated the data, and prepared the initial draft of the writing. D.H. and Z.S. were primarily responsible for conducting the experiments. Y.Z. was dedicated to the curation and management of the data. S.W. reviewed and edited the article. Y.W. conceived and designed the study, acquired funding, and provided overall supervision. All authors concurred and approved the final version of the article.
Data Availability
All data supporting the findings of this study are included within the article and its supplementary materials. Raw micro-CT images, histology data, and additional experimental datasets are available from the corresponding author upon reasonable request.
Footnotes
Acknowledgments
Authors sincerely thank Mr. Guang Yang and Mr. Guangneng Liao from The Experimental Animal Center of West China Hospital, Sichuan University, for their full support and selfless assistance in this research. Meanwhile, authors also express their profound gratitude to Mr. Jiehao Chen for his valuable guidance in MicroCT scanning and data analysis.
Author Disclosure Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Funding Information
This work was financially sponsored and supported by
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.